
Comparison of the Cisterna Maturation-Progression Model with the Kiss-and-Run Model of Intra-Golgi Transport: Role of Cisternal Pores and Cargo Domains †



Abstract
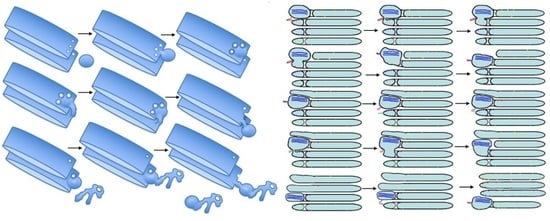
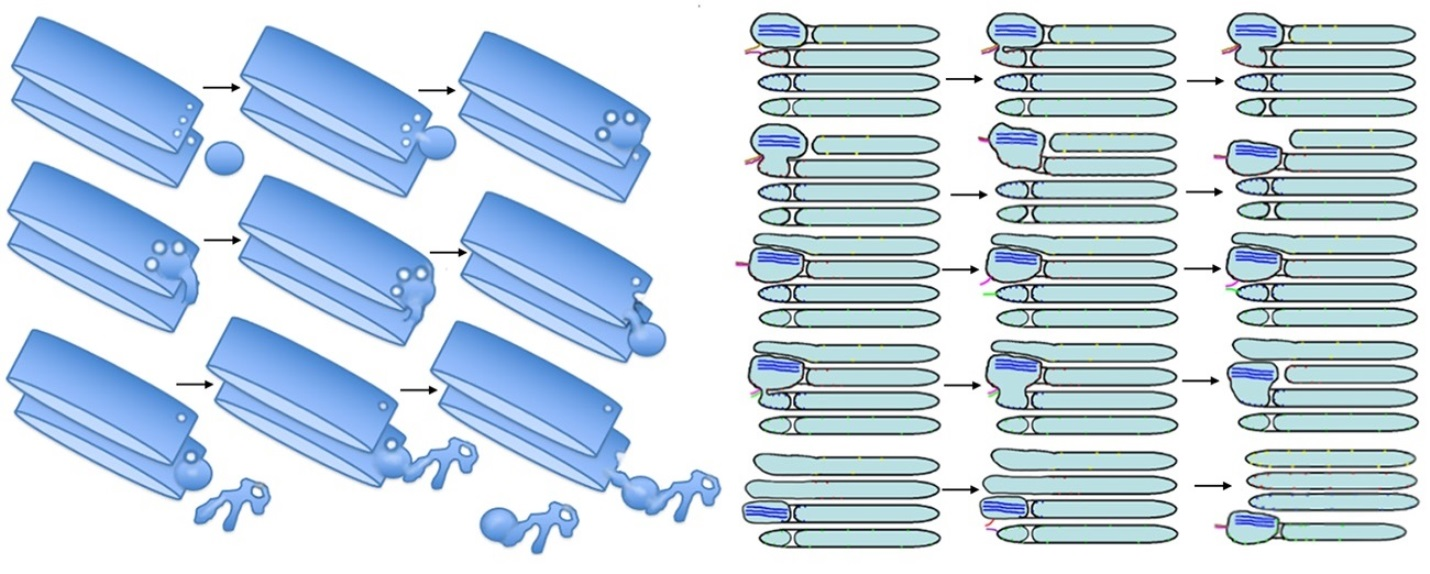
1. Introduction
2. Results
2.1. Background
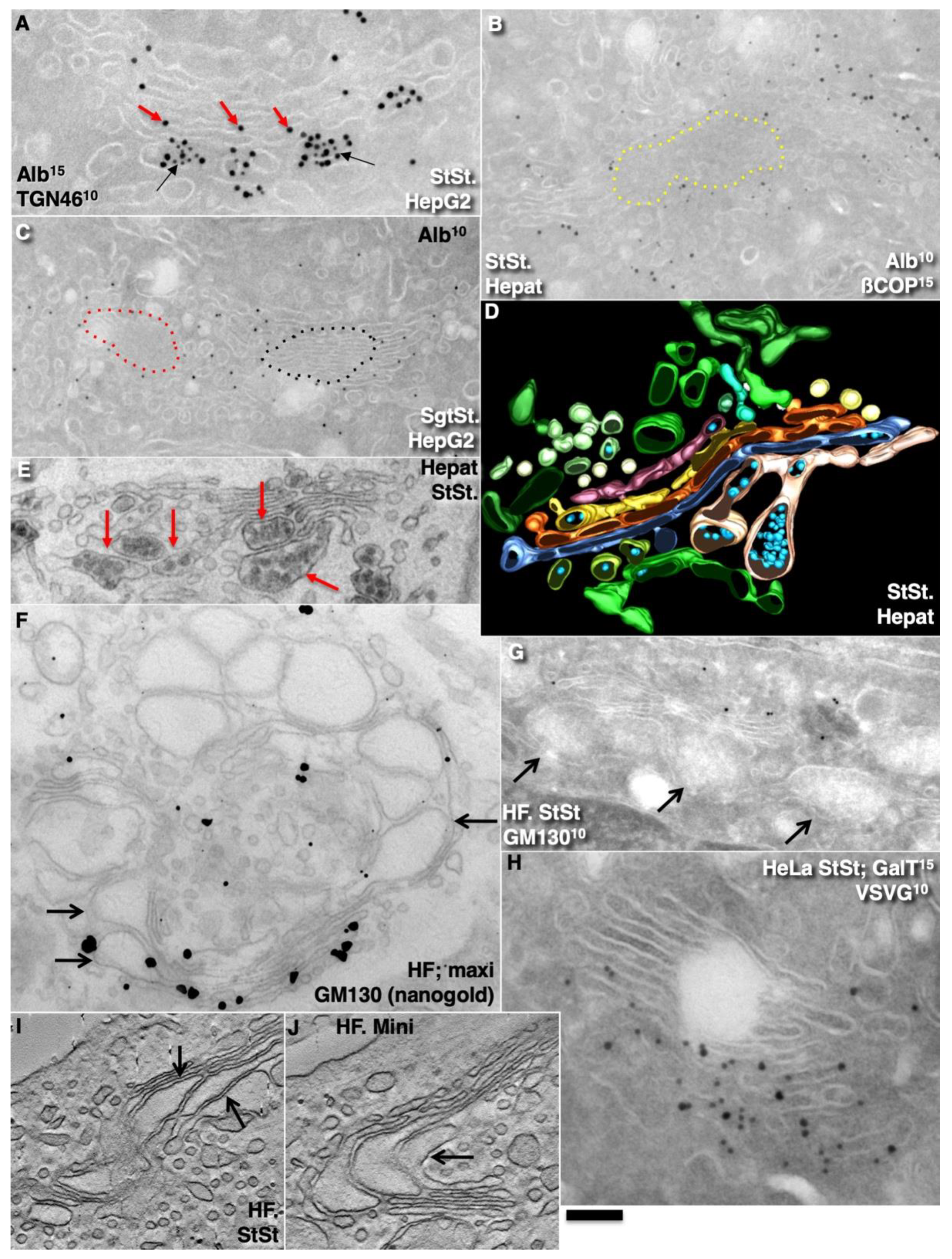
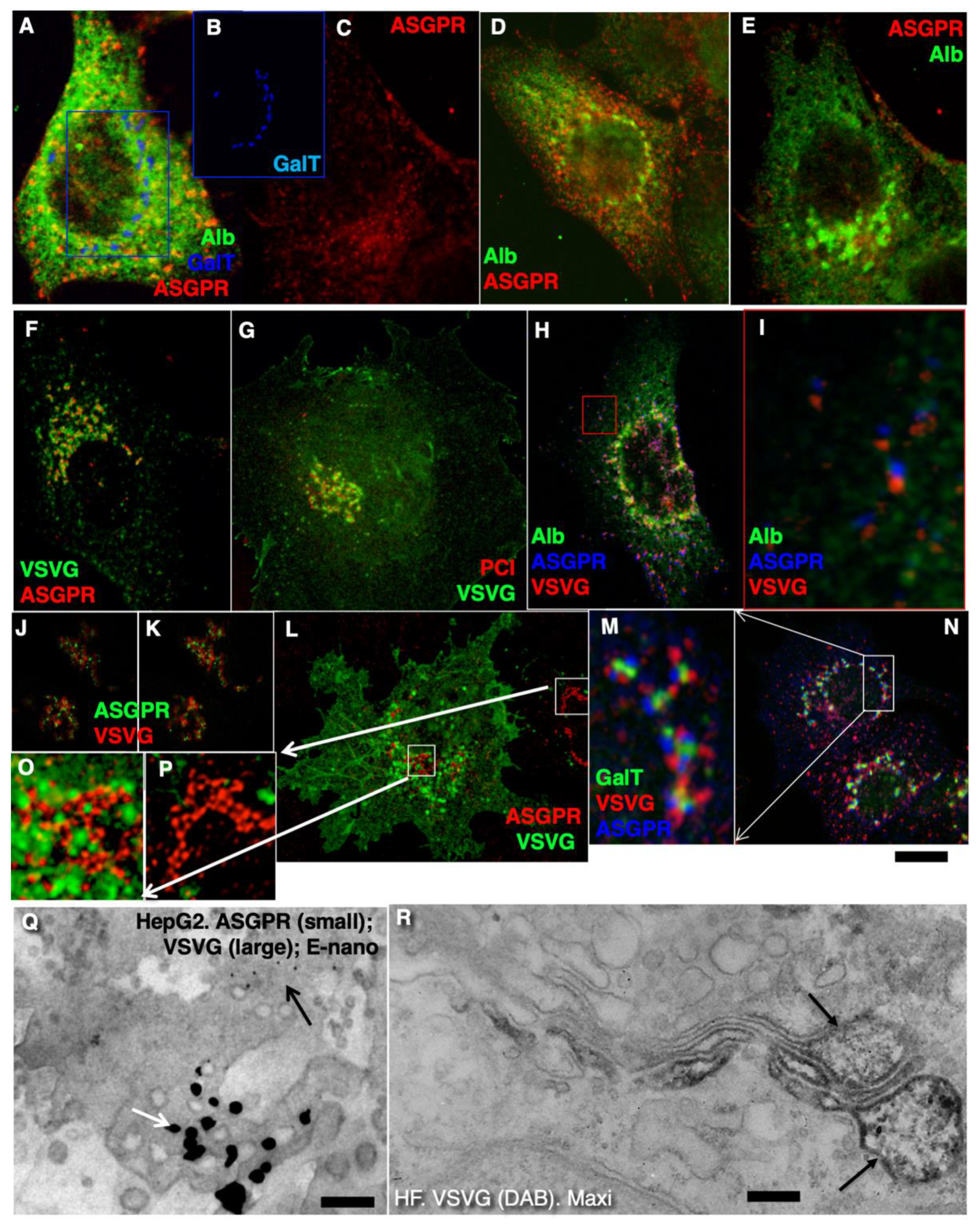
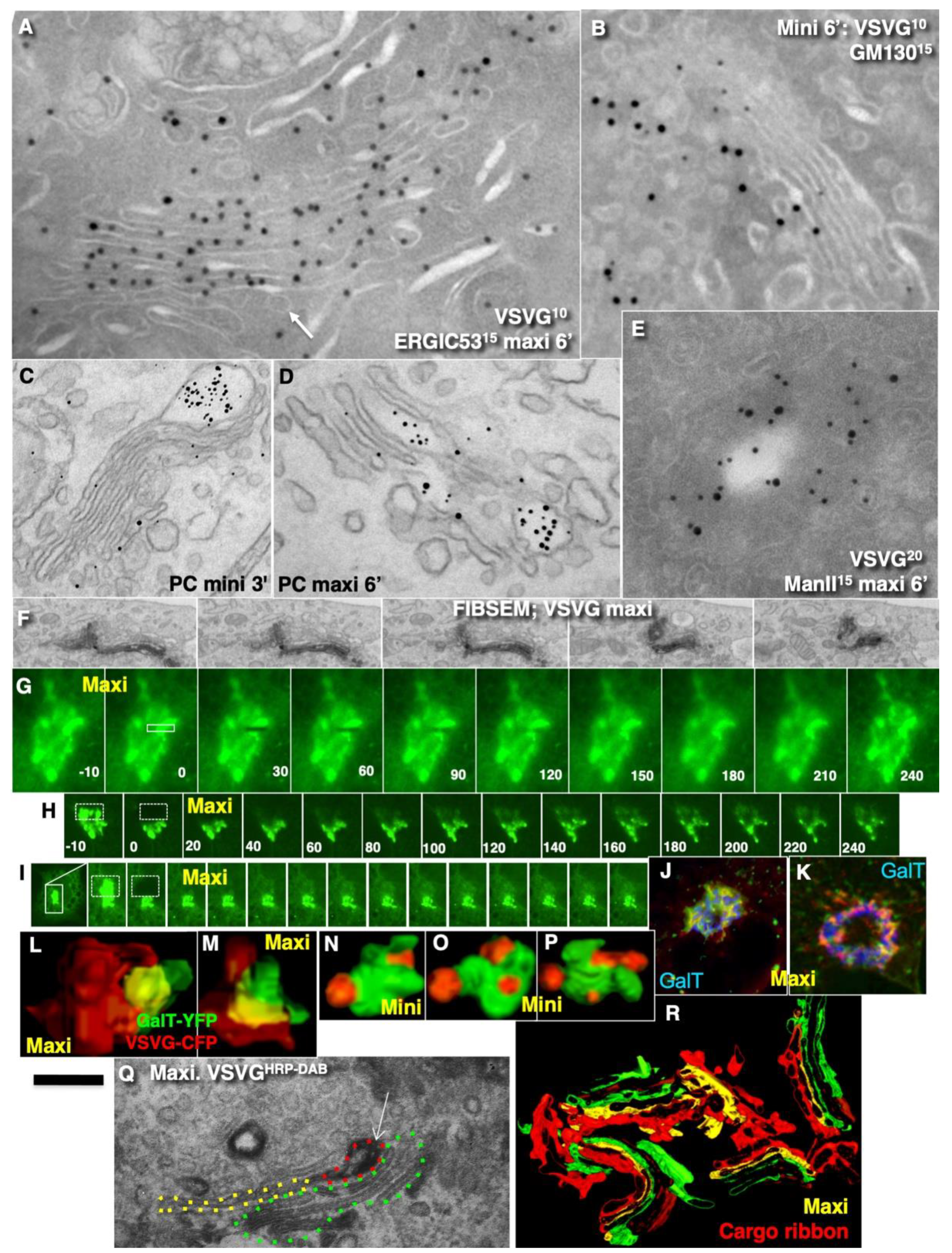
2.2. Conventional Mega-Cargoes Are Concentrated at the Trans-Side of the Golgi Complex
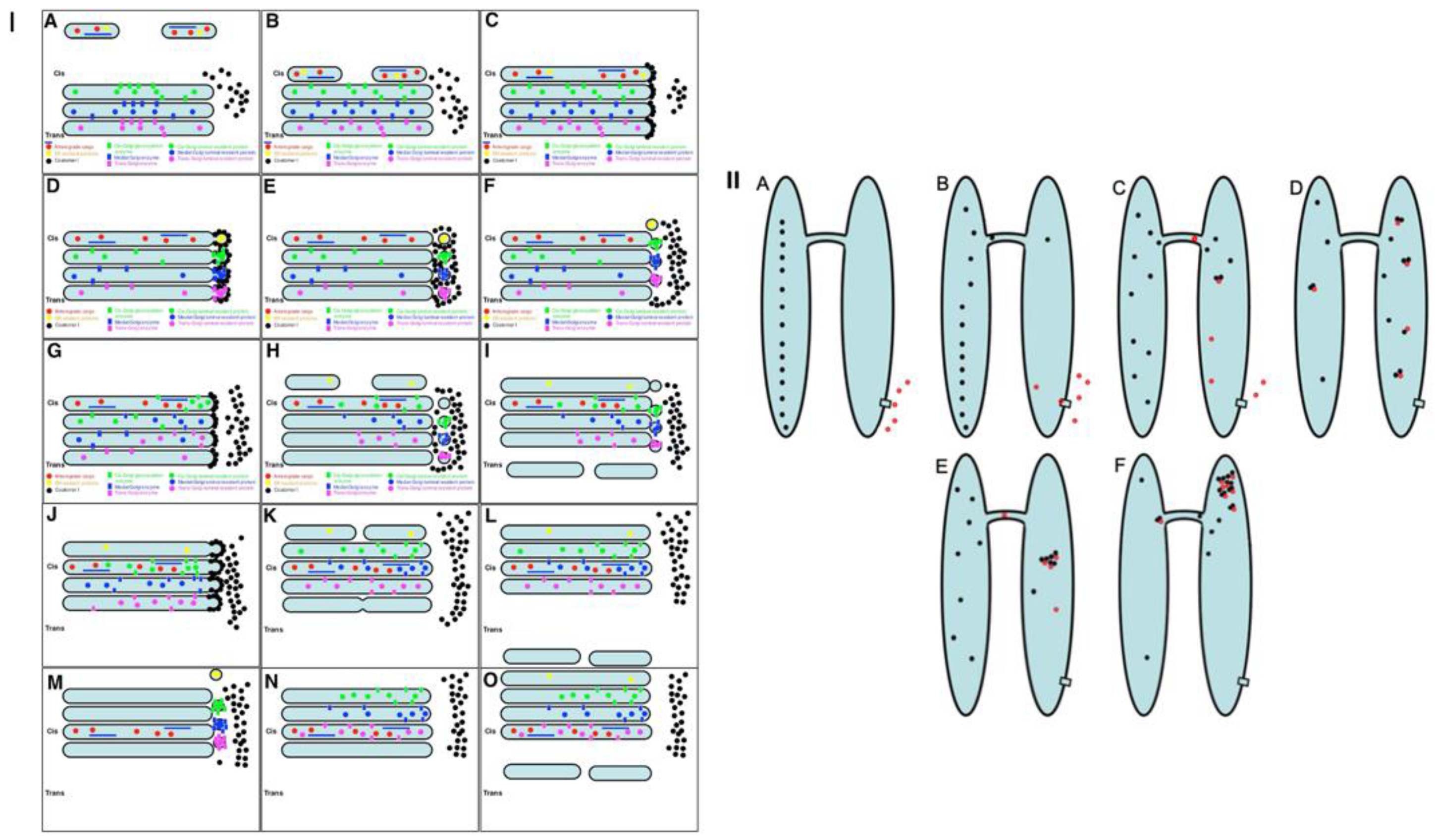
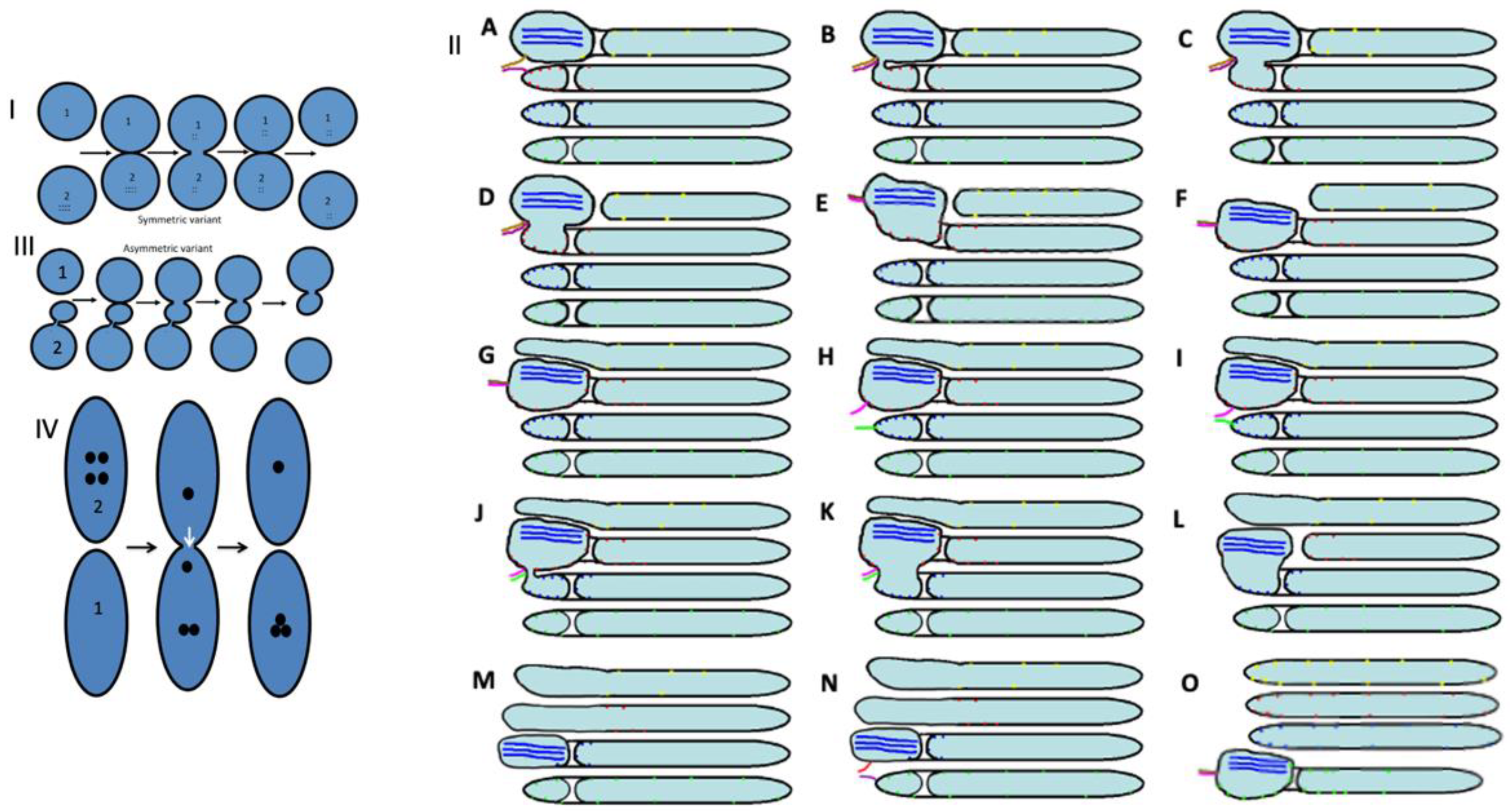
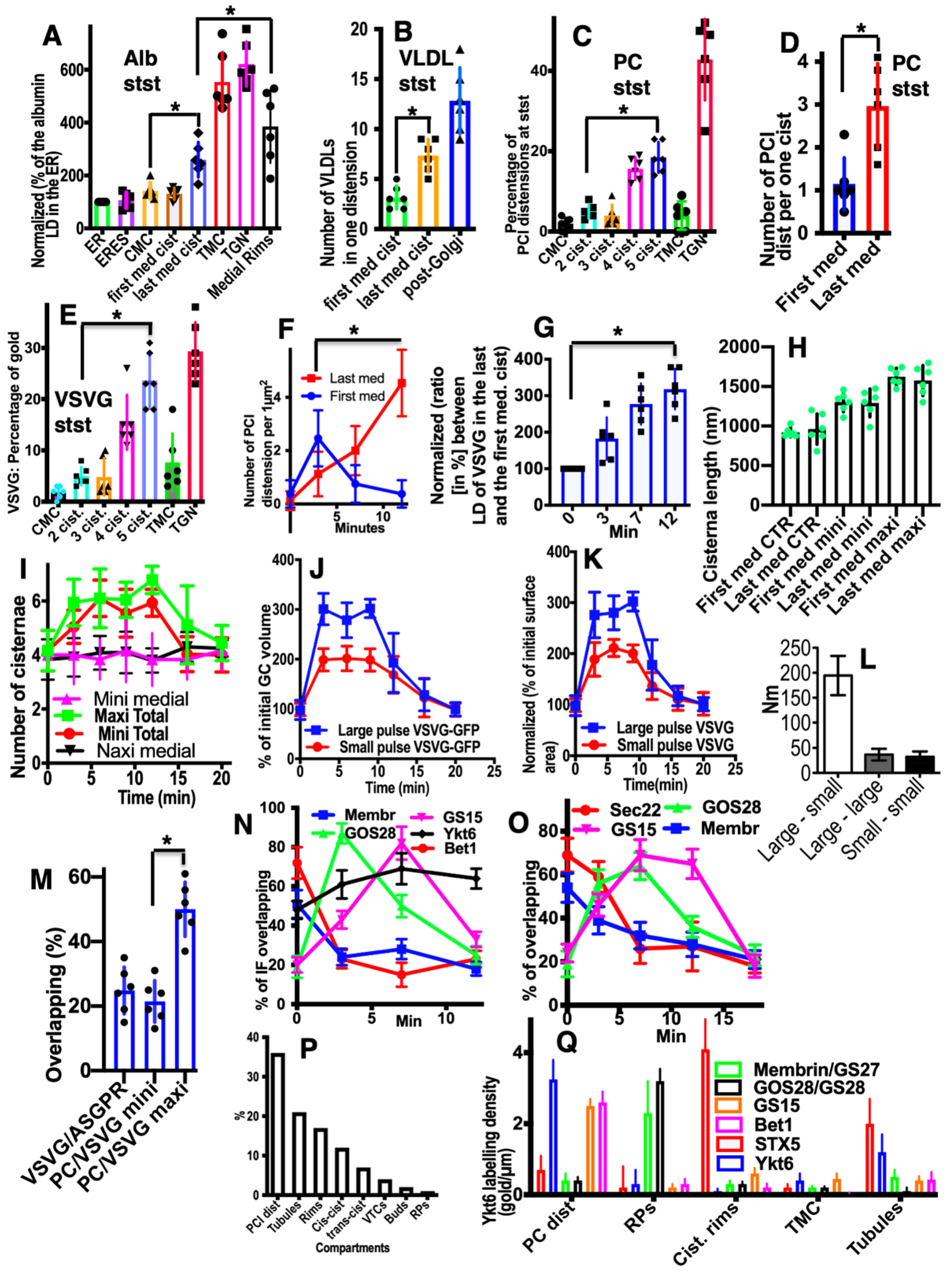
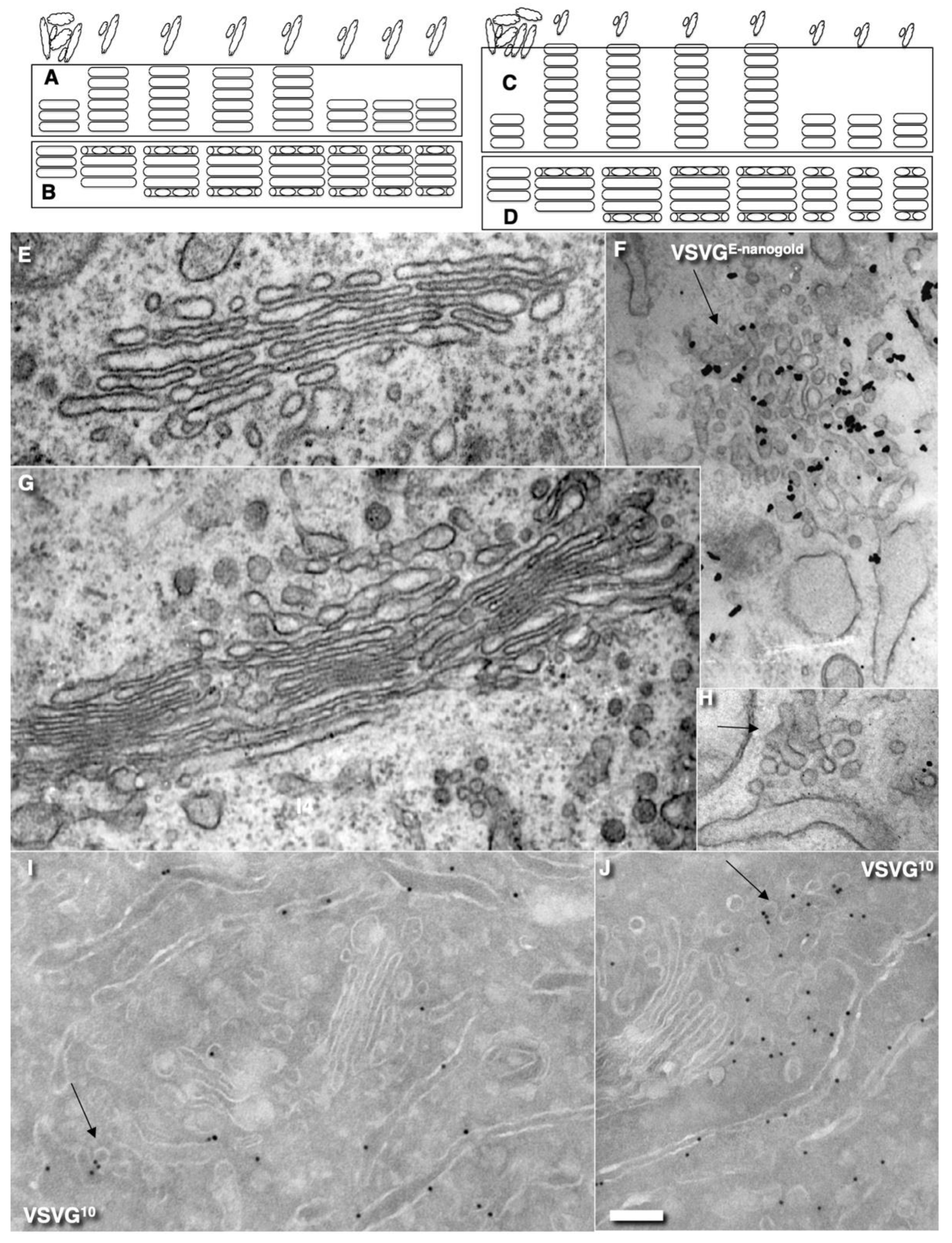
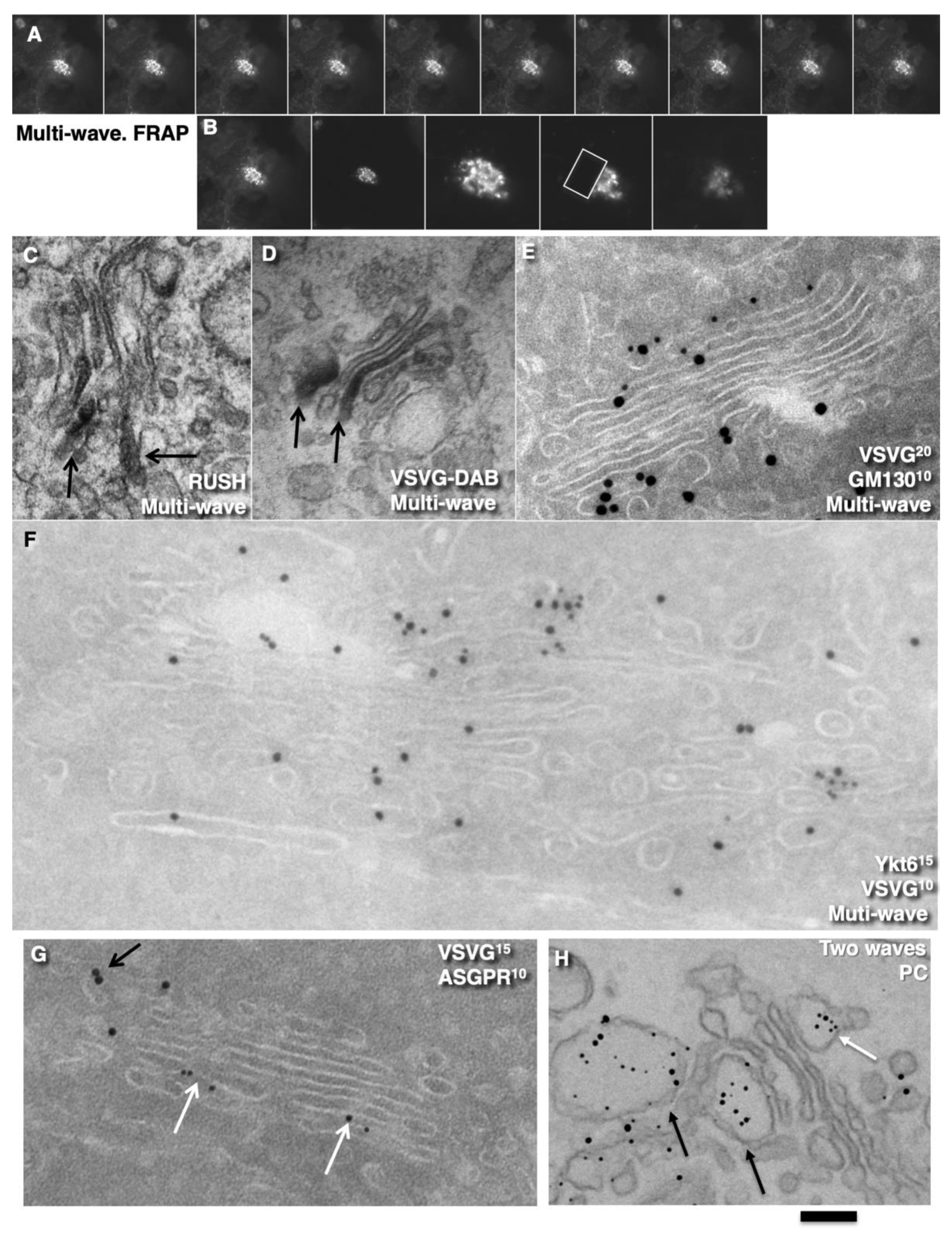
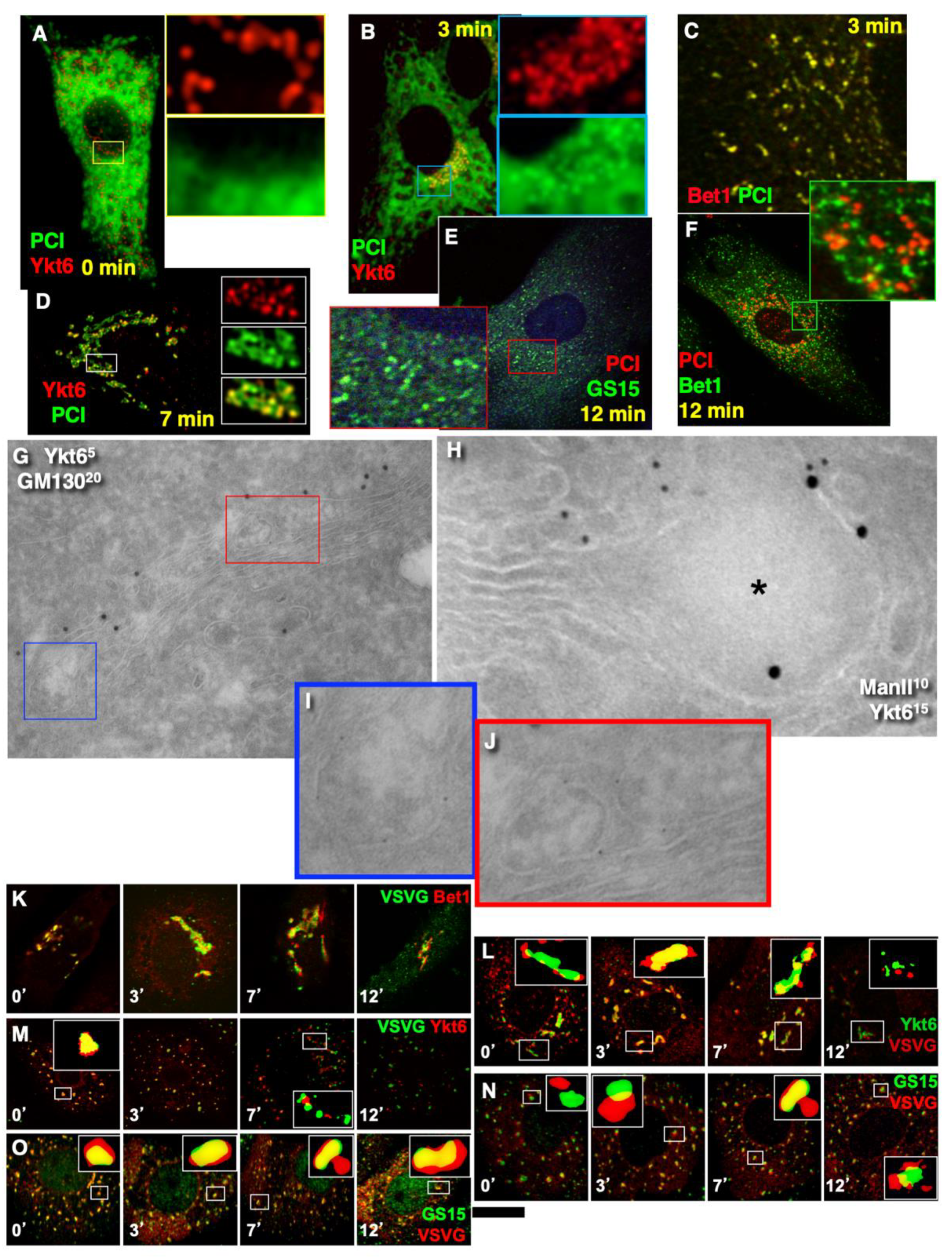
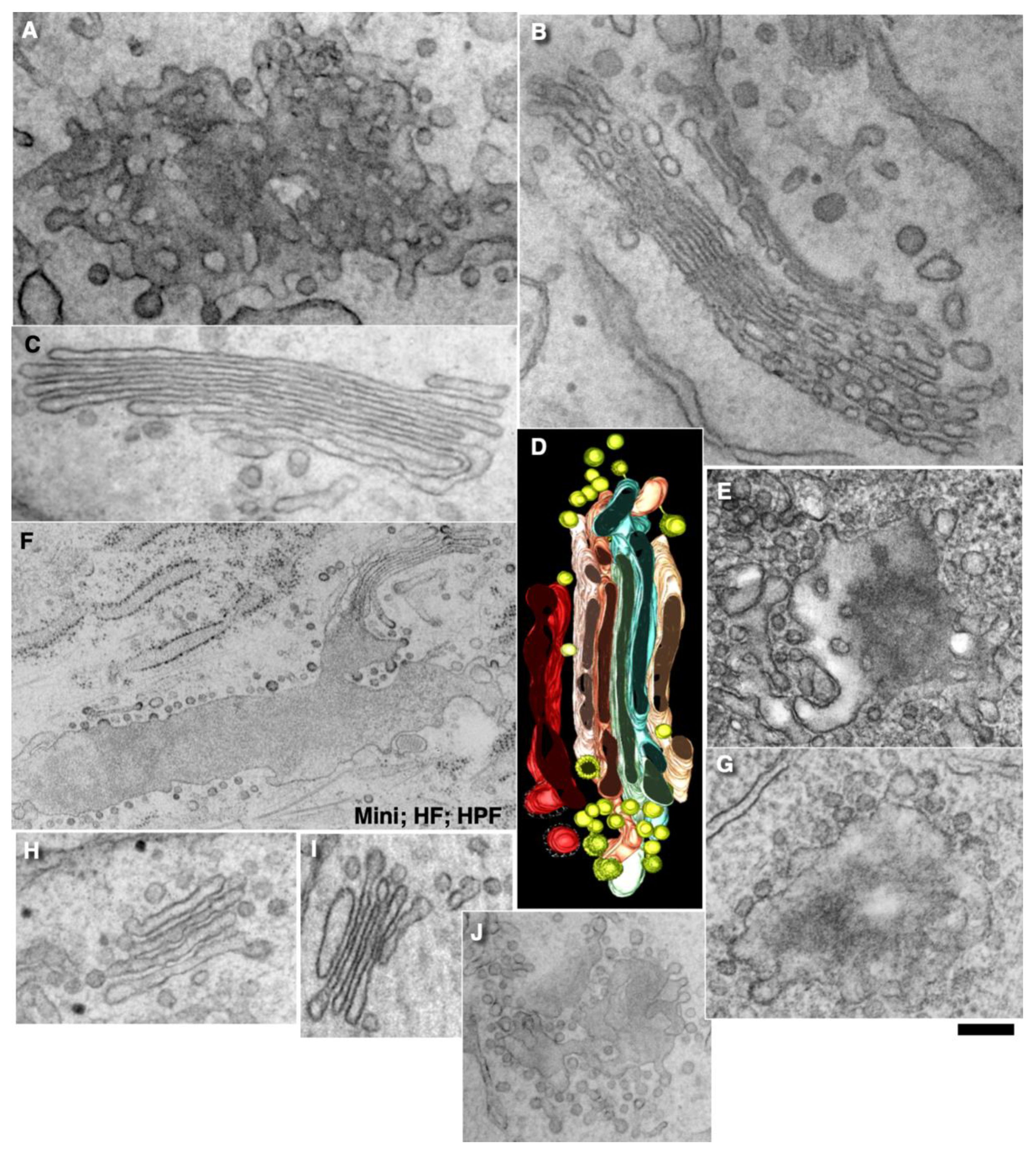
2.3. Membrane Dynamics
2.4. Membrane Cargoes Form Domains
2.5. Different Membrane Cargoes Form Distinct Domains
2.6. SNARE Enrichment over Cargo Domains
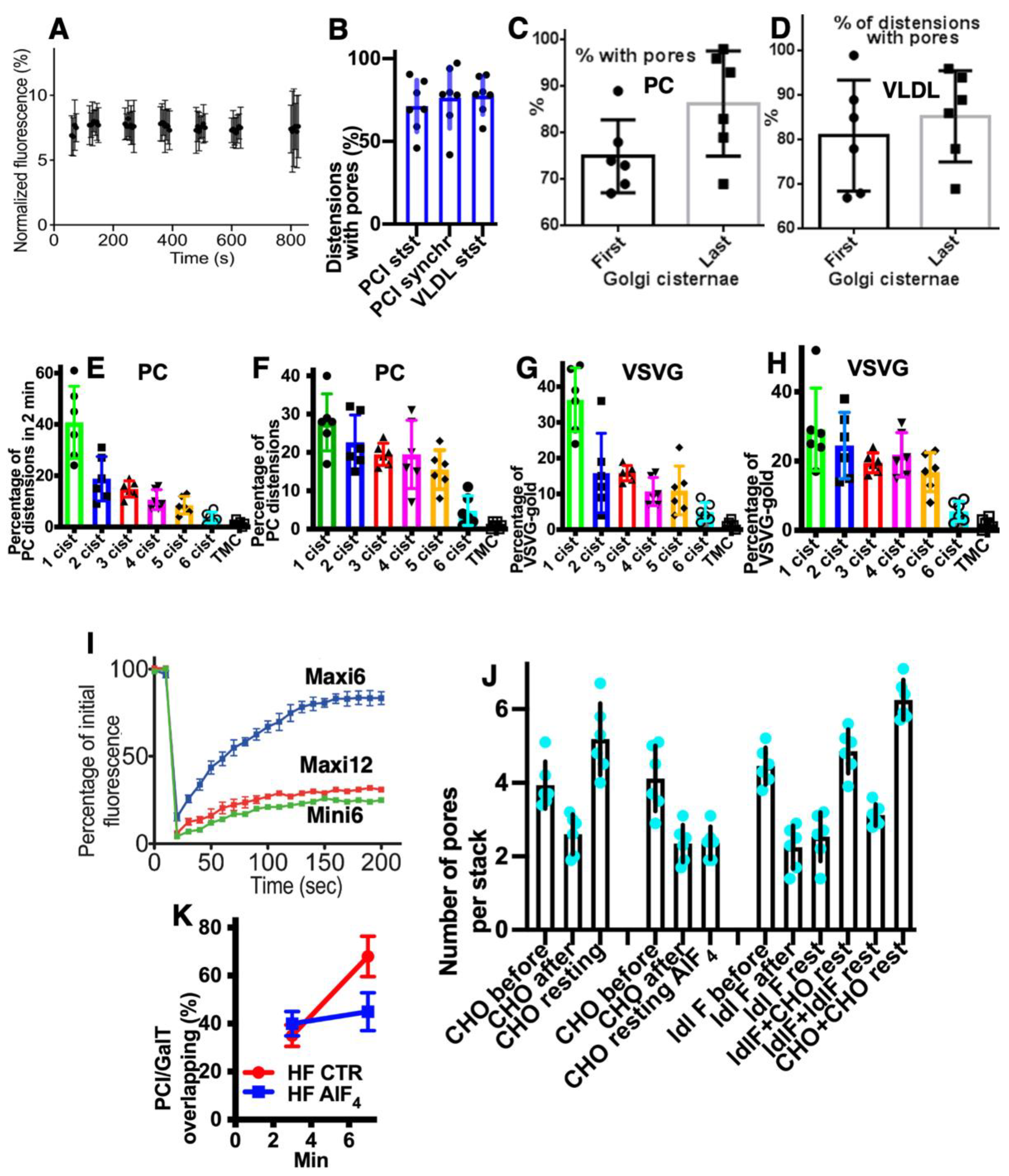
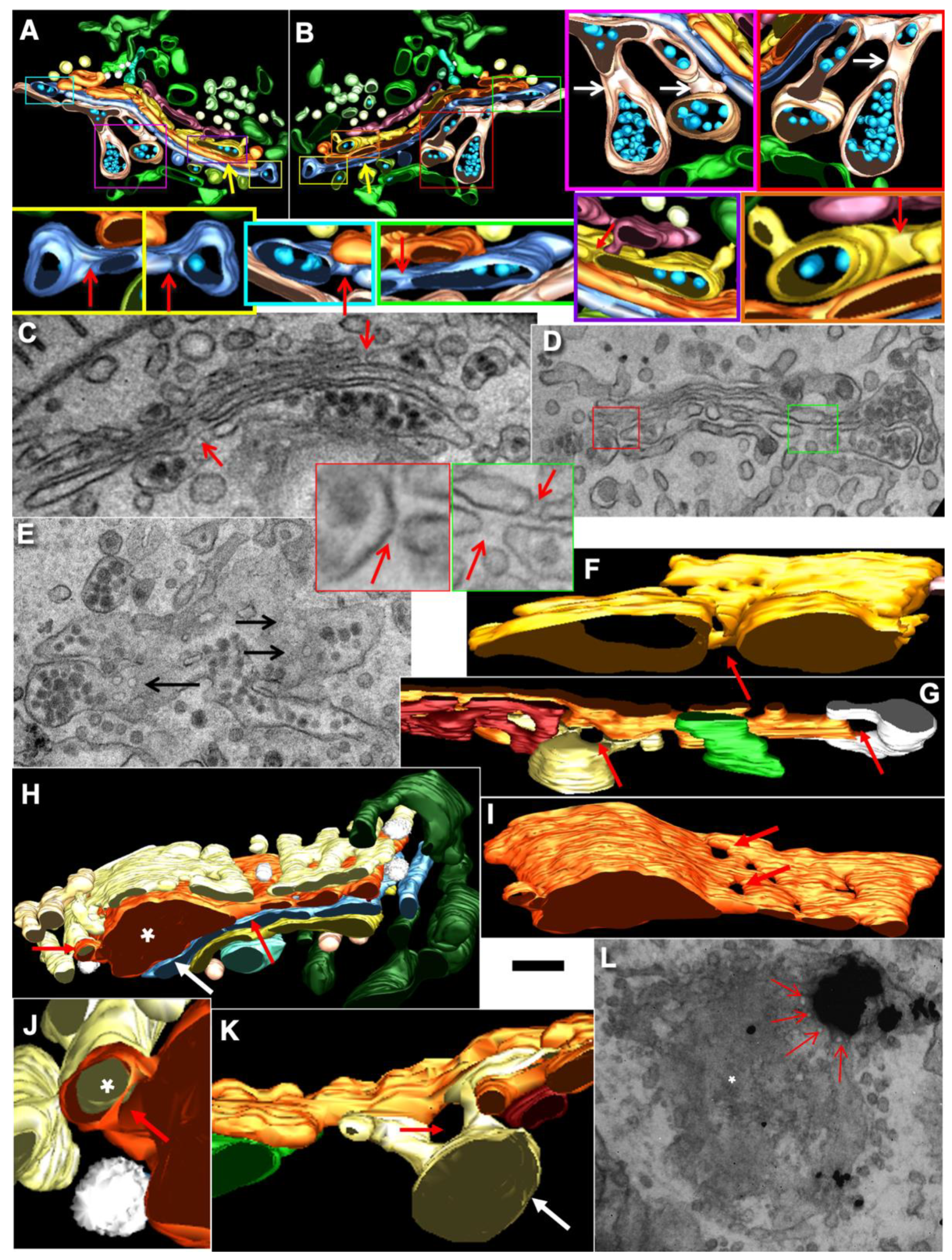
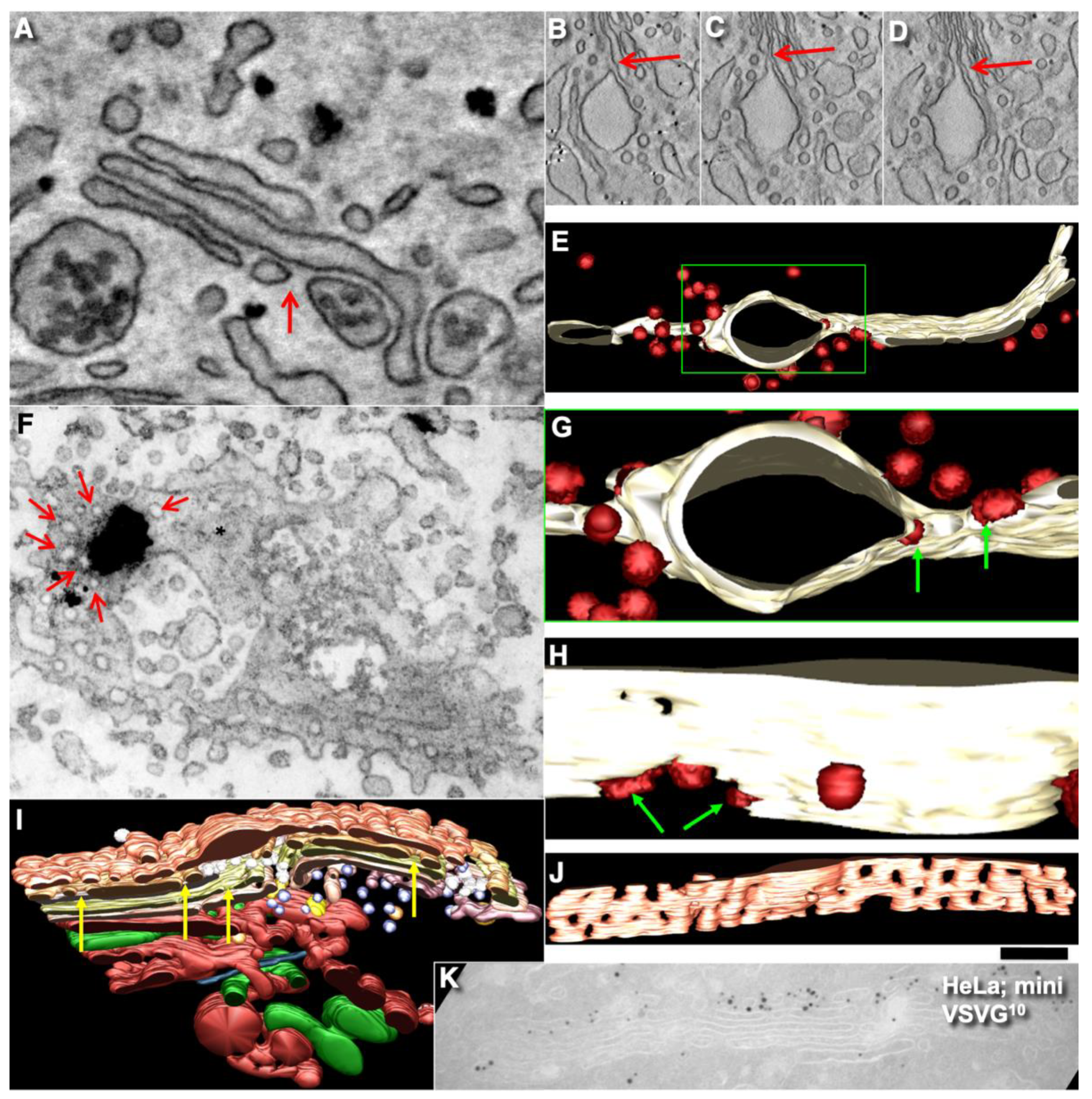
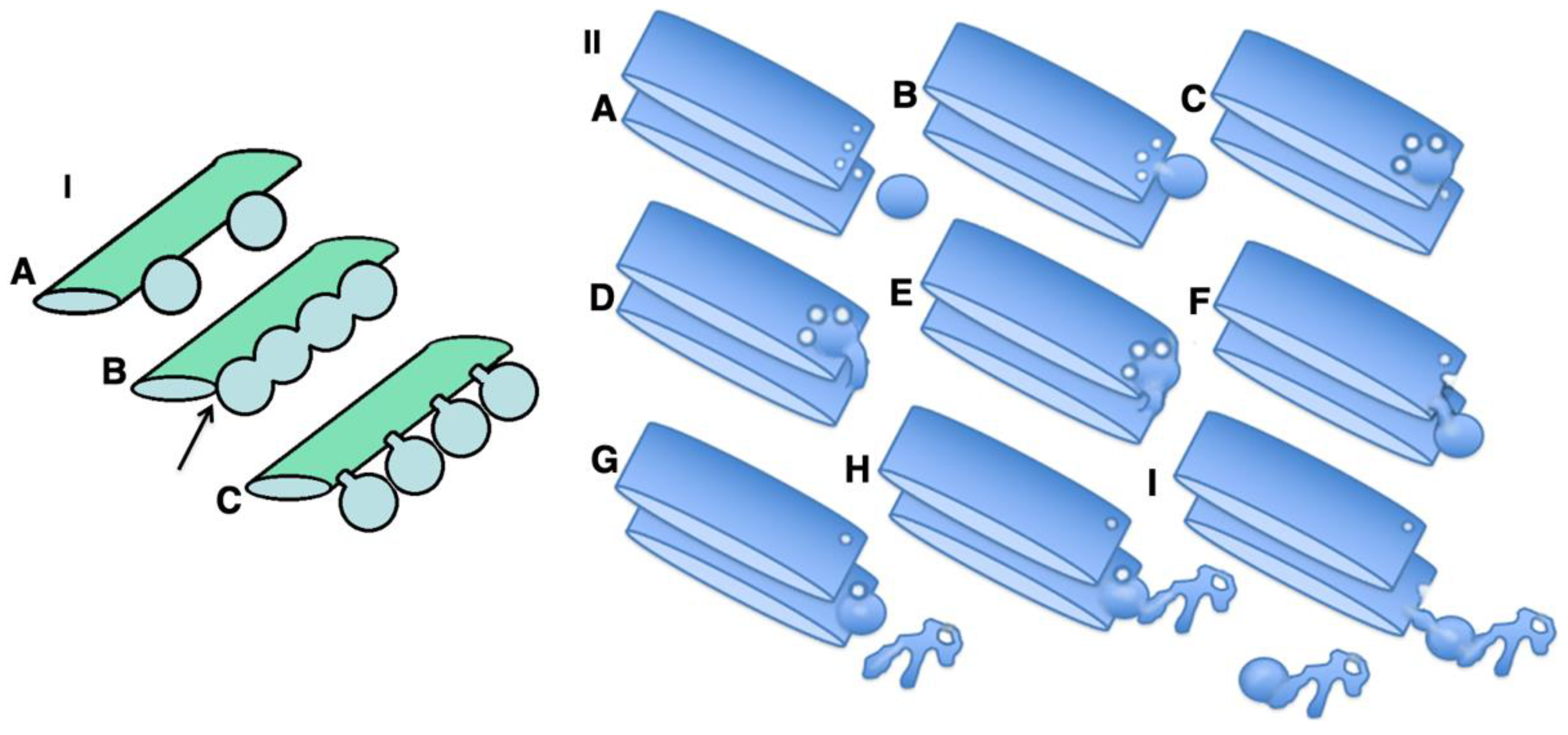
2.7. Pores Separate Cargo Domains and the Rest of the Golgi Cisternae
2.8. Transport of Different Amounts of Membranes
2.9. COPI Is Important for Restoration of Cisternal Pores
3. Discussion
How to Reconcile the Hypothesis with Previous Data and Models
4. Materials and Methods
4.1. Reagents and Treatments

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagent or Resource | Source | Identifier |
|---|---|---|
| Antibodies | ||
| Rabbit polyclonal antibody against GFP | Abcam (Cambridge, UK) | ab6556 |
| Rabbit polyclonal antibody against syntaxin 5 | Abcam (Cambridge, UK) | ab211417 |
| Rabbit polyclonal antibody against syntaxin 5 | Santa Cruz Biotechnology (USA) | Sc365124 |
| Rabbit polyclonal antibody against Bet1 | Abcam (Cambridge, UK) | ab42488 |
| Rabbit polyclonal antibody against GM130 | Abcam (Cambridge, UK) | ab31561 |
| Monoclonal antibody against VSVG (P5D4) | Abcam(Cambridge, UK) | ab50549 |
| Rabbit polyclonal antibody against mannosidase II | Abcam(Cambridge, UK) | ab12277 |
| Rabbit polyclonal against asialoglycoprotein receptor | Abcam(Cambridge, UK) | ab42488 |
| Rabbit polyclonal antibody against albumin | Abcam (Cambridge, UK) | ab42488 |
| Rabbit polyclonal antibody against Ykt6 | Abcam(Cambridge, UK) | ab236583 |
| Rabbit polyclonal anti-human antibody against β-1,3-galactosyltransferase 5 | MyBioSource Inc. (San Diego CA. USA) | MBS2026257 |
| PCI polyclonal against the α1 chain C-terminal domain of PCI | L. W. Fisher (National Institutes of Health) | N/A |
| Affinity-purified polyclonal sheep antibody against human pro-collagen I α1 | R&D Systems, Inc. (NY, USA) | AF6220 |
| Rabbit polyclonal antibody against ßCOP | Dr J. Lippincott-Schwartz (Howard Hughes Medical Institute, Ashburn, VA, USA) | N/A |
| Rabbit polyclonal antibody against GS15 | MyBioSource, Inc. (San Diego CA. USA) | MBS2525056) |
| Rabbit polyclonal antibody against membrin | Biorbyt Ltd. (NY; USA) | orb185573 |
| Rabbit polyclonal antibody against GOSR1(GS28) | Biorbyt Ltd. (NY; USA) | orb326297 |
| Rabbit polyclonal antibody against TGN46 | S Ponnambalam (Leeds University, UK) | N/A |
| GFP-albumin | Beznoussenko et al. [25] | N/A |
| Fab fragments of polyclonal antibodies conjugated with HRP | Jackson ImmunoResearch (West Grove, PA, USA) | Not produced anymore |
| Fab fragments of the polyclonal antibodies against IgGs | Jackson ImmunoResearch (West Grove, PA, USA) | Not produced anymore |
| Mouse IgG rabbit polyclonal antibody | ThermoFisher Scientific (USA; https://www.thermofisher.com, accessed on 15 March 2022) | 31188 |
| Goat anti-rabbit, anti-mouse, and anti-sheep IgG antibodies conjugated with Alexa 488, Alexa 546 | ThermoFisher Scientific (USA; https://www.thermofisher.com, accessed on 15 March 2022) | Many |
| Goat anti-rabbit Fab’ fragments of IgG conjugated with Nanogold | Nanoprobes (Yaphank, NY, USA) | #2004-1 mL |
| Protein A conjugated with gold particles of different sizes | Utrecht Universit. Utrecht; The Neverland) | Direct order |
| Chemicals, peptides, and recombinant proteins | ||
| Gold enhancer | Nanoprobes (Yaphank, NY, USA) | |
| Reagents for electron microscopy | EMS or Sigma (Merck) | Many |
| FUGENE6 transfection reagent | Roche (Monza, Italy). | |
| Recombinant DNA | ||
| Plasmid containing α1-chain of PC tagged with GFP | Beznoussenko et al. [25]; Patterson et al. [28] | N/A |
| Plasmid with GFP-Alb | Beznoussenko et al. [25] | N/A |
| Plasmid with PCIII-GFP | Perinetti et al. [112] | N/A |
| Software and algorithms | ||
| Image J | NIH, Bethesda | N/A |
| IMOD 4.0.11 package | The Boulder Laboratory for 3-D Electron Microscopy of Cells. University of Colorado Boulder | http://bio3d.colorado.edu/imod/accessed on 20 January 2020 |
| Prism: Version 9.4.2 package | GraphPad (NY; USA) | https://www.graphpad.com/scientific-software/prism/ accessed on 20 January 2020 |
4.2. Cells
4.3. Animals
4.4. Constructs
4.5. Experiments with Cells
4.6. Heterokaryon Assay
4.7. Fluorescence Microscopy
4.8. Electron Microscopy
4.9. Pre-Embedding Immuno-Electron Microscopy
4.10. Cryosections
4.11. Electron Microscopy Tomography
4.12. Focused Ion Beam Scanning EM
4.13. Sampling
4.14. Stereology
4.15. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Velasco, A.; Hendricks, L.; Moreman, K.W.; Tulsiani, D.R.P.; Touster, O.; Farquhar, M.G. Cell-type dependent variations in the subcellular distribution of a-mannosidase I and II. J. Cell Biol. 1993, 122, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, J.D.; Palade, G.E. Intracellular transport of secretory proteins in the pancreatic exocrine cell. I. Role of the peripheral elements of the Golgi complex. J. Cell Biol. 1967, 34, 577–596. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, J.D.; Palade, G.E. Intracellular transport of secretory proteins in the pancreatic exocrine cell. II. Transport to condensing vacuoles and zymogen granules. J. Cell Biol. 1967, 34, 597–615. [Google Scholar] [CrossRef]
- Jamieson, J.D.; Palade, G.E. Intracellular transport of secretory proteins in the pancreatic exocrine cell. IV. Metabolic requirements. J. Cell Biol. 1968, 39, 589–603. [Google Scholar] [CrossRef]
- Jamieson, J.D.; Palade, G.E. Intracellular transport of secretory proteins in the pancreatic exocrine cell. 3. Dissociation of intracellular transport from protein synthesis. J. Cell Biol. 1968, 39, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Palade, G. Intracellular aspects of the process of protein synthesis. Science 1975, 189, 347–359. [Google Scholar] [CrossRef]
- Farquhar, M.G.; Palade, G.E. The Golgi apparatus (complex)-(1954–1981)-from artifact to center stage. J. Cell Biol. 1981, 91, 77s–103s. [Google Scholar] [CrossRef]
- Rothman, J.E.; Bursztyn-Pettegrew, H.; Fine, R.E. Transport of the membrane glycoprotein of vesicular stomatitis virus to the cell surface in two stages by clathrin-coated vesicles. J. Cell Biol. 1980, 86, 162–171. [Google Scholar] [CrossRef]
- Orci, L.; Glick, B.S.; Rothman, J.E. A new type of coated vesicular carrier that appears not to contain clathrin: Its possible role in protein transport within the Golgi stack. Cell 1986, 46, 171–184. [Google Scholar] [CrossRef]
- Marsh, B.J.; Mastronarde, D.N.; Buttle, K.F.; Howell, K.E.; McIntosh, J.R. Organellar relationships in the Golgi region of pancreatic beta cell line, HIT-T15, visualized by high resolution electron tomography. Proc. Natl. Acad. Sci. USA 2001, 98, 2399–2406. [Google Scholar] [CrossRef]
- Lakatos, I. The Methodology of Scientific Research Programmes; Philosophical Papers Vol. 1; Cambridge University Press: Cambridge, UK, 1978; p. 250. [Google Scholar]
- Mironov, A.A.; Weidman, P.; Luini, A. Variations on the intracellular transport theme: Maturing cisternae and trafficking tubules. J. Cell Biol. 1997, 138, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Mironov, A., Jr.; Luini, A.; Mironov, A. A synthetic model of intra-Golgi traffic. FASEB J. 1998, 12, 249–252. [Google Scholar] [CrossRef] [PubMed]
- Mironov, A.A.; Beznoussenko, G.V.; Polishchuk, R.S.; Trucco, A. Intra-Golgi transport. A way to a new paradigm? BBA-Mol. Cell Res. 2005, 1744, 340–350. [Google Scholar] [CrossRef]
- Beznoussenko, G.V.; Mironov, A.A. Models of intracellular transport and evolution of the Golgi complex. Anat. Rec. 2002, 268, 226–238. [Google Scholar] [CrossRef] [PubMed]
- Bonfanti, L.; Mironov, A.A., Jr.; Martínez-Menárguez, J.A.; Martella, O.; Fusella, A.; Baldassarre, M.; Buccione, R.; Geuze, H.J.; Mironov, A.A.; Luini, A. Procollagen traverses the Golgi stack without leaving the lumen of cisternae: Evidence for cisternal maturation. Cell 1998, 95, 993–1003. [Google Scholar] [CrossRef]
- Volchuk, A.; Amherdt, M.; Ravazzola, M.; Brügger, B.; Rivera, V.M.; Clackson, T.; Perrelet, A.; Söllner, T.H.; Rothman, J.E.; Orci, L. Megavesicles implicated in the rapid transport of intracisternal aggregates across the Golgi stack. Cell 2000, 102, 335–348. [Google Scholar] [CrossRef]
- Mironov, A.A.; Beznoussenko, G.V.; Nicoziani, P.; Martella, O.; Trucco, A.; Kweon, H.-S.; Di Giandomenico, D.; Polishchuk, R.S.; Fusella, A.; Lupetti, P.; et al. Small cargo proteins and large aggregates can traverse the Golgi by a common mechanism without leaving the lumen of cisternae. J. Cell Biol. 2001, 155, 1225–1238. [Google Scholar] [CrossRef]
- Mironov, A.A.; Beznoussenko, G.V. Intra-Golgi transport. In The Golgi Apparatus. State of the Art 110 Years after Camillo Golgi’s Discovery; Mironov, A.A., Pavelka, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 342–357. [Google Scholar]
- Glick, B.S.; Elston, T.; Oster, G. A cisternal maturation mechanism can explain the asymmetry of the Golgi stack. FEBS Lett. 1997, 414, 177–181. [Google Scholar]
- Kweon, H.S.; Beznoussenko, G.V.; Micaroni, M.; Polishchuk, R.S.; Trucco, A.; Martella, O.; Di Giandomenico, D.; Marra, P.; Fusella, A.; Di Pentima, A.; et al. Golgi enzymes are enriched in perforated zones of Golgi cisternae but are depleted in COPI vesicles. Mol. Biol. Cell 2004, 15, 4710–4724. [Google Scholar] [CrossRef]
- Fusella, A.; Micaroni, M.; Di Giandomenico, D.; Mironov, A.A.; Beznoussenko, G.V. Segregation of the Qb-SNAREs GS27 and GS28 into Golgi vesicles regulates intra-Golgi transport. Traffic 2013, 14, 568–584. [Google Scholar] [CrossRef]
- Beznoussenko, G.V.; Ragnini-Wilson, A.; Wilson, C.; Mironov, A.A. Three-dimensional and immune electron microscopic analysis of the secretory pathway in Saccharomyces cerevisiae. Histochem. Cell Biol. 2016, 146, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Trucco, A.; Polishchuk, R.S.; Martella, O.; Di Pentima, A.; Fusella, A.; Di Giandomenico, D.; San Pietro, E.; Beznoussenko, G.V.; Polishchuk, E.V.; Baldassarre, M.; et al. Secretory traffic triggers the formation of tubular continuities across Golgi sub-compartments. Nat. Cell Biol. 2004, 6, 1071–1081. [Google Scholar] [CrossRef] [PubMed]
- Beznoussenko, G.V.; Parashuraman, S.; Rizzo, R.; Polishchuk, R.; Martella, O.; Di Giandomenico, D.; Fusella, A.; Spaar, A.; Sallese, M.; Capestrano, M.G.; et al. Transport of soluble proteins through the Golgi occurs by diffusion via continuities across cisternae. eLife 2014, 3, e02009. [Google Scholar] [CrossRef] [PubMed]
- Sleight, R.G.; Pagano, R.E. Rapid appearance of newly synthesized phosphatidylethanolamine at the plasma membrane. J. Biol. Chem. 1983, 258, 9050–9058. [Google Scholar] [CrossRef]
- Pagano, R.E.; Longmuir, K.J. Phosphorylation, transbilayer movement, and facilitated intracellular transport of diacylglycerol are involved in the uptake of a fluorescent analog of phosphatidic acid by cultured fibroblasts. J. Biol. Chem. 1985, 260, 1909–1916. [Google Scholar] [CrossRef]
- Patterson, G.H.; Hirschberg, K.; Polishchuk, R.S.; Gerlich, D.; Phair, R.D.; Lippincott-Schwartz, J. Transport through the Golgi apparatus by rapid partitioning within a two-phase membrane system. Cell 2008, 133, 1055–1067. [Google Scholar] [CrossRef]
- Pagano, R.E.; Sepanski, M.A.; Martin, O.C. Molecular trapping of a fluorescent ceramide analogue at the Golgi apparatus of fixed cells: Interaction with endogenous lipids provides a trans-Golgi marker for both light and electron microscopy. J. Cell Biol. 1989, 109, 2067–2079. [Google Scholar] [CrossRef]
- Presley, J.F.; Cole, N.B.; Schroer, T.A.; Hirschberg, K.; Zaal, K.J.; Lippincott-Schwartz, J. ER-to-Golgi transport visualized in living cells. Nature 1997, 389, 81–85. [Google Scholar] [CrossRef]
- Mironov, A.A.; Colanzi, A.; Polishchuk, R.S.; Beznoussenko, G.V.; Mironov, A.A., Jr.; Fusella, A.; Di Tullio, G.; Silletta, M.G.; Corda, D.; De Matteis, M.A.; et al. Dicumarol, an inhibitor of ADP-ribosylation of CtBP3/BARS, fragments Golgi non-compact tubular zones and inhibits intra-Golgi transport. Eur. J. Cell Biol. 2004, 83, 263–279. [Google Scholar] [CrossRef]
- Mavillard, F.; Hidalgo, J.; Megias, D.; Levitsky, K.L.; Velasco, A. PKA-mediated Golgi remodeling during cAMP signal transmission. Traffic 2010, 11, 90–109. [Google Scholar] [CrossRef]
- Valtorta, F.; Meldolesi, J.; Fesce, R. Synaptic vesicles: Is kissing a matter of competence? Trends Cell Biol. 2001, 11, 324–328. [Google Scholar] [CrossRef]
- Mironov, A.A.; Beznoussenko, G.V. The kiss-and-run model of intra-Golgi transport. Int. J. Mol. Sci. 2012, 13, 6800–6819. [Google Scholar] [CrossRef] [PubMed]
- Mironov, A.A.; Sesorova, I.V.; Beznoussenko, G.V. Golgi’s way: A long path toward the new paradigm of the intra-Golgi transport. Histochem. Cell Biol. 2013, 140, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Mironov, A.A.; Komissarchik, Y.Y.; Mironov, A.A., Jr.; Snigirevskaya, E.S.; Luini, A. Currents concepts of structure and function of the Golgi complex. On the 100th anniversary of the discovery by Camillo Golgi. Tsitologiia 1998, 40, 483–496. [Google Scholar] [PubMed]
- Mironov, A.A.; Dimov, I.D.; Beznoussenko, G.V. Role of Intracellular Transport in the Centriole-Dependent Formation of Golgi Ribbon. Results Probl. Cell Differ. 2019, 67, 49–79. [Google Scholar] [PubMed]
- Mironov, A.A.; Beznoussenko, G.V. Models of Intracellular Transport: Pros and Cons. Front. Cell Dev. Biol. 2019, 7, 146. [Google Scholar] [CrossRef]
- Popper, K. The Logic of Scientific Discovery; Huttchinson: London, UK, 1959; p. 475. [Google Scholar]
- Kuhn, T.S. The Structure of Scientific Revolutions; University of Chicago Press: Chicago, IL, USA, 1962; p. 172. [Google Scholar]
- Sesorova, I.S.; Karelina, N.R.; Kazakova, T.E.; Parashuraman, S.; Zdorikova, M.A.; Dimov, I.D.; Seliverstova, E.V.; Beznoussenko, G.V.; Mironov, A.A. Structure of the enterocyte transcytosis compartments during lipid absorption. Histochem. Cell Biol. 2020, 153, 413–429. [Google Scholar] [CrossRef]
- Leblond, C.P. Synthesis and secretion of collagen by cells of connective tissue, bone, and dentin. Anat. Rec. 1989, 224, 123–138. [Google Scholar] [CrossRef]
- Jäntti, J.; Hildén, P.; Rönkä, H.; Mäkiranta, V.; Keränen, S.; Kuismanen, E. Immunocytochemical analysis of Uukuniemi virus budding compartments: Role of the intermediate compartment and the Golgi stack in virus maturation. J. Virol. 1997, 71, 1162–1172. [Google Scholar] [CrossRef]
- Orci, L.; Stamnes, M.; Ravazzola, M.; Amherdt, M.; Perrelet, A.; Sollner, T.H.; Rothman, J.E. Bidirectional transport by distinct populations of COPI-coated vesicles. Cell 1997, 90, 335–349. [Google Scholar] [CrossRef]
- Vetterlein, M.; Ellinger, A.; Neumuller, J.; Pavelka, M. Golgi apparatus and TGN during endocytosis. Histochem. Cell Biol. 2002, 117, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Orci, L.; Perrelet, A.; Rothman, J.E. Vesicles on strings: Morphological evidence for processive transport within the Golgi stack. Proc. Natl. Acad. Sci. USA 1998, 95, 2279–2283. [Google Scholar] [CrossRef] [PubMed]
- Dancourt, J.; Zheng, H.; Bottanelli, F.; Allgeyer, E.S.; Bewersdorf, J.; Graham, M.; Liu, X.; Rothman, J.E.; Lavieu, G. Small cargoes pass through synthetically glued Golgi stacks. FEBS Lett. 2016, 590, 1675–1686. [Google Scholar] [CrossRef] [PubMed]
- Presley, J.F.; Smith, C.; Hirschberg, K.; Miller, C.; Cole, N.B.; Zaal, K.J.; Lippincott-Schwartz, J. Golgi membrane dynamics. Mol. Biol. Cell 1998, 9, 1617–1626. [Google Scholar] [CrossRef]
- Polishchuk, R.S.; Polishchuk, E.V.; Mironov, A.A. Coalescence of Golgi fragments in microtubule-deprived living cells. Eur. J. Cell Biol. 1999, 78, 170–185. [Google Scholar] [CrossRef]
- Denisova, G.N.; Dimov, I.D.; Zaitseva, A.V.; Artiux, L.J.; Mironov, A.A.; Karelina, N.R. Overloading of differentiated Caco-2 cells during lipid transcytosis induces glycosylation mistakes in the Golgi complex. Biocell 2021, 45, 773–783. [Google Scholar] [CrossRef]
- Gilchrist, A.; Au, C.E.; Hiding, J.; Bell, A.W.; Fernandez-Rodriguez, J.; Lesimple, S.; Nagaya, H.; Roy, L.; Gosline, S.J.; Hallett, M.; et al. Quantitative proteomics analysis of the secretory pathway. Cell 2006, 127, 1265–1281. [Google Scholar] [CrossRef]
- Martinez-Menarguez, J.A.; Prekeris, R.; Oorschot, V.M.; Scheller, R.; Slot, J.W.; Geuze, H.J.; Klumperman, J. Peri-Golgi vesicles contain retrograde but not anterograde proteins consistent with the cisternal progression model of intra-Golgi transport. J. Cell Biol. 2001, 155, 1213–1224. [Google Scholar] [CrossRef]
- Rizzo, R.; Parashuraman, S.; Mirabelli, P.; Puri, C.; Lucocq, J.; Luini, A. The dynamics of engineered resident proteins in the mammalian Golgi complex relies on cisternal maturation. J. Cell Biol. 2013, 201, 1027–1036. [Google Scholar] [CrossRef]
- Pothukuchi, P.; Agliarulo, I.; Pirozzi, M.; Rizzo, R.; Russo, D.; Turacchio, G.; Nüchel, J.; Yang, J.; Gehin, C.; Capolupo, L.; et al. GRASP55 regulates intra-Golgi localization of glycosylation enzymes to control glycosphingolipid biosynthesis. EMBO J. 2021, 40, e107766. [Google Scholar] [CrossRef]
- Claude, A. Growth and differentiation of cytoplasmic membranes in the course of lipoprotein granule synthesis in the hepatic cell. I. Elaboration of elements of the Golgi complex. J. Cell Biol. 1970, 47, 745–766. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, S.; Tashiro, Y. Immuno-electron-microscopic studies of endoplasmic reticulum-Golgi relationships in the intracellular transport process of lipoprotein particles in rat hepatocytes. J. Cell Sci. 1979, 39, 273–290. [Google Scholar] [CrossRef] [PubMed]
- Pavelka, M.; Roth, J. Functional Ultrastructure: Atlas of Tissue Biology and Pathology; Springer: Berlin/Heidelberg, Germany, 2005; p. 326. [Google Scholar]
- Rhodin, J.A.G. Histology: A Text and Atlas; Oxford University Press: New York, NY, USA, 1974; p. 803. [Google Scholar]
- Glaumann, H.; Bergstrand, A.; Ericsson, J.L. Studies on the synthesis and intracellular transport of lipoprotein particles in rat liver. J Cell Biol. 1975, 64, 356–377. [Google Scholar] [CrossRef] [PubMed]
- Oprins, A.; Rabouille, C.; Posthuma, G.; Klumperman, J.; Geuze, H.J.; Slot, J.W. The ER to Golgi interface is the major concentration site of secretory proteins in the exocrine pancreatic cell. Traffic 2001, 2, 831–838. [Google Scholar] [CrossRef]
- Dahan, S.; Ahluwalia, J.P.; Wong, L.; Posner, B.I.; Bergeron, J.J. Concentration of intracellular hepatic apolipoprotein E in Golgi apparatus saccular distensions and endosomes. J. Cell Biol. 1994, 127, 1859–1869. [Google Scholar] [CrossRef]
- Klumperman, J.; Locker, J.K.; Meijer, A.; Horzinek, M.C.; Geuze, H.J.; Rottier, P.J. Coronavirus M proteins accumulate in the Golgi complex beyond the site of virion budding. J. Virol. 1994, 68, 6523–6534. [Google Scholar] [CrossRef]
- Taylor, R.S.; Jones, S.M.; Dahl, R.H.; Nordeen, M.H.; Howell, K.E. Characterization of the Golgi complex cleared of proteins in transit and examination of calcium uptake activities. Mol. Biol. Cell 1997, 8, 1911–1931. [Google Scholar] [CrossRef]
- Wild, P.; Kaech, A.; Schraner, E.M.; Walser, L.; Ackermann, M. Endoplasmic reticulum-to-Golgi transitions upon herpes virus infection [version 1]. F1000Research 2017, 6, 1804. [Google Scholar] [CrossRef]
- Cortese, M.; Lee, J.Y.; Cerikan, B.; Neufeldt, C.J.; Oorschot, V.M.J.; Köhrer, S.; Hennies, J.; Schieber, N.L.; Ronchi, P.; Mizzon, G.; et al. Integrative Imaging Reveals SARS-CoV-2-Induced Reshaping of Subcellular Morphologies. Cell Host Microbe 2020, 28, 853–866. [Google Scholar] [CrossRef]
- Clermont, Y.; Xia, L.; Rambourg, A.; Turner, J.D.; Hermo, L. Transport of casein submicelles and formation of secretion granules in the Golgi apparatus of epithelial cells of the lactating mammary gland of rat. Anat. Rec. 1993, 235, 363–373. [Google Scholar] [CrossRef]
- Rambourg, A.; Clermont, Y.; Chrétien, M.; Olivier, L. Modulation of the Golgi apparatus in stimulated and nonstimulated prolactin cells of female rats. Anat Rec. 1993, 235, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Mironov, A.A.; Luini, A.; Buccione, R. Constitutive transport between the trans-Golgi network and the plasma membrane according to the maturation model. A hypothesis. FEBS Lett. 1998, 440, 99–102. [Google Scholar] [CrossRef]
- Mironov, A.A.; Sesorova, I.S.; Dimov, I.D.; Karelina, N.R.; Beznoussenko, G.V. Intracellular transports and atherogenesis. Front. Biosci. 2020, 25, 1230–1258. [Google Scholar] [CrossRef] [PubMed]
- Boncompain, G.; Divoux, S.; Gareil, N.; de Forges, H.; Lescure, A.; Latreche, L.; Mercanti, V.; Jollivet, F.; Raposo, G.; Perez, F. Synchronization of secretory protein traffic in populations of cells. Nat. Methods 2012, 9, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Mironov, A.A.; Mironov, A.A., Jr.; Beznoussenko, G.V.; Trucco, A.; Lupetti, P.; Smith, J.D.; Geerts, W.J.C.; Koster, A.J.; Burger, K.N.J.; Martone, M.E.; et al. ER-to-Golgi carriers arise through direct en bloc protrusion and multistage maturation of specialized ER exit domains. Dev. Cell 2003, 5, 583–594. [Google Scholar] [CrossRef]
- Griffiths, G.; Fuller, S.D.; Back, R.; Hollinshead, M.; Pfeiffer, S.; Simons, K. The dynamic nature of the Golgi complex. J. Cell Biol. 1989, 108, 277–297. [Google Scholar] [CrossRef]
- Bergmann, J.E. Using temperature-sensitive mutants of VSV to study membrane protein biogenesis. Methods Cell Biol. 1989, 32, 85–110. [Google Scholar]
- Polishchuk, R.S.; Polishchuk, E.V.; Marra, P.; Alberti, S.; Buccione, R.; Luini, A.; Mironov, A.A. Correlative light-electron microscopy reveals the tubular-saccular ultrastructure of carriers operating between Golgi apparatus and plasma membrane. J. Cell Biol. 2000, 148, 45–58. [Google Scholar] [CrossRef]
- Hirschberg, K.; Miller, C.M.; Ellenberg, J.; Presley, J.F.; Siggia, E.D.; Phair, R.B.; Lippincott-Schwartz, J. Kinetic analysis of secretory protein traffic and characterization of Golgi to plasma membrane transport in living cells. J. Cell Biol. 1998, 143, 1485–1503. [Google Scholar] [CrossRef]
- Luini, A.; Mironov, A.A.; Polishchuk, E.V.; Polishchuk, R.S. Morphogenesis of post-Golgi transport carriers. Histochem. Cell Biol. 2008, 129, 153–161. [Google Scholar] [CrossRef]
- Mironov, A.A.; Sesorova, I.S.; Seliverstova, E.V.; Beznoussenko, G.V. Different Golgi ultrastructure across species and tissues: Implications under functional and pathological conditions, and an attempt at classification. Tissue Cell 2017, 49, 186–201. [Google Scholar] [CrossRef] [PubMed]
- Sabesin, S.M.; Frase, S. Electron microscopic studies of the assembly, intracellular transport, and secretion of chylomicrons by rat intestine. J. Lipid Res. 1977, 18, 496–511. [Google Scholar] [CrossRef]
- Sesso, A.; de Faria, F.P.; Iwamura, E.S.; Corrêa, H. A three-dimensional reconstruction study of the rough ER-Golgi interface in serial thin sections of the pancreatic acinar cell of the rat. J. Cell Sci. 1994, 107, 517–528. [Google Scholar] [CrossRef]
- Sesorova, I.S.; Dimov, I.D.; Kashin, A.D.; Sesorov, V.V.; Karelina, N.R.; Zdorikova, M.A.; Beznoussenko, G.V.; Mironov, A.A. Cellular and sub-cellular mechanisms of lipid transport from gut to lymph. Tissue Cell 2021, 72, 101529. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.D.; Elamawi, R.; Bubeck, J.; Pepperkok, R.; Ritzenthaler, C.; Robinson, D.G. Dynamics of COPII vesicles and the Golgi apparatus in cultured Nicotiana tabacum BY-2 cells provides evidence for transient association of Golgi stacks with endoplasmic reticulum exit sites. Plant Cell 2005, 17, 1513–1531. [Google Scholar] [CrossRef]
- Yang, J.S.; Lee, S.Y.; Gao, M.; Bourgoin, S.; Randazzo, P.A.; Premont, R.T.; Hsu, V.W. ARFGAP1 promotes the formation of COPI vesicles, suggesting function as a component of the coat. J. Cell Biol. 2002, 159, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Orci, L.; Schekman, R.; Perrelet, A. Interleaflet clear space is reduced in the membrane of COPI and COPII-coated buds/vesicles. Proc. Natl. Acad. Sci. USA 1996, 93, 8968–8970. [Google Scholar] [CrossRef]
- Yang, J.S.; Valente, C.; Polishchuk, R.S.; Turacchio, G.; Layre, E.; Moody, D.B.; Leslie, C.C.; Gelb, M.H.; Brown, W.J.; Corda, D.; et al. COPI acts in both vesicular and tubular transport. Nat. Cell Biol. 2011, 13, 996–1003. [Google Scholar] [CrossRef]
- Park, S.Y.; Yang, J.S.; Schmider, A.B.; Soberman, R.J.; Hsu, V.W. Coordinated regulation of bidirectional COPI transport at the Golgi by CDC42. Nature 2015, 521, 529–532. [Google Scholar] [CrossRef]
- Pepperkok, R.; Lowe, M.; Burke, B.; Kreis, T.E. Three distinct steps in transport of vesicular stomatitis virus glycoprotein from the ER to the cell surface in vivo with differential sensitivities to GTP gamma S. J. Cell Sci. 1998, 111, 1877–1888. [Google Scholar] [CrossRef]
- Weigert, R.; Colanzi, A.; Mironov, A.; Buccione, R.; Cericola, C.; Sciulli, M.G.; Santini, G.; Flati, S.; Fusella, A.; Donaldson, J.G.; et al. Characterization of chemical inhibitors of brefeldin A-activated mono-ADP-ribosylation. J. Biol. Chem. 1997, 272, 14200–14207. [Google Scholar] [CrossRef] [PubMed]
- Capaci, V.; Bascetta, L.; Fantuz, M.; Beznoussenko, G.V.; Sommaggio, R.; Cancila, V.; Bisso, A.; Campaner, E.; Mironov, A.A.; Wiśniewski, J.R.; et al. Mutant p53 induces Golgi tubulo-vesiculation driving a prometastatic secretome. Nat. Commun. 2020, 11, 3945. [Google Scholar] [CrossRef] [PubMed]
- Shima, D.T.; Scales, S.J.; Kreis, T.; Pepperkok, R. Segregation of COPI-rich and anterograde-cargo rich domains in endoplasmic-reticulum-to-Golgi complexes. Curr. Biol. 1999, 9, 821–824. [Google Scholar] [CrossRef]
- Beznoussenko, G.V.; Pilyugin, S.S.; Geerts, W.J.; Kozlov, M.M.; Burger, K.N.; Luini, A.; Derganc, J.; Mironov, A.A. Trans-membrane area asymmetry controls the shape of cellular organelles. Int. J. Mol. Sci. 2015, 16, 5299–5333. [Google Scholar] [CrossRef] [PubMed]
- Mironov, A.A.; Mironov, A.; Derganc, J.; Beznoussenko, G.V. Membrane Curvature, Trans-Membrane Area Asymmetry, Budding, Fission and Organelle Geometry. Int. J. Mol. Sci. 2020, 21, 7594. [Google Scholar] [CrossRef]
- Morelli, E.; Speranza, E.A.; Pellegrino, E.; Beznoussenko, G.V.; Carminati, F.; Garré, M.; Mironov, A.A.; Onorati, M.; Vaccari, T. Activity of the SNARE Protein SNAP29 at the Endoplasmic Reticulum and Golgi Apparatus. Front. Cell Dev. Biol. 2021, 9, 637565. [Google Scholar] [CrossRef]
- Gomez, M.; Scales, S.J.; Kreis, T.E.; Perez, F. Membrane recruitment of coatomer and binding to dilysine signals are separate events. J. Biol. Chem. 2000, 275, 29162–29169. [Google Scholar] [CrossRef]
- Kurokawa, K.; Osakada, H.; Kojidani, T.; Waga, M.; Suda, Y.; Asakawa, H.; Haraguchi, T.; Nakano, A. Visualization of secretory cargo transport within the Golgi apparatus. J. Cell Biol. 2019, 218, 1602–1618. [Google Scholar] [CrossRef]
- Casler, J.C.; Papanikou, E.; Barrero, J.J.; Glick, B.S. Maturation-driven transport and AP-1-dependent recycling of a secretory cargo in the Golgi. J. Cell Biol. 2019, 218, 1582–1601. [Google Scholar] [CrossRef]
- Dunlop, M.H.; Ernst, A.M.; Schroeder, L.K.; Toomre, D.K.; Lavieu, G.; Rothman, J.E. Land-locked mammalian Golgi reveals cargo transport between stable cisternae. Nat. Commun. 2017, 8, 432. [Google Scholar] [CrossRef]
- Xiang, Y.; Zhang, X.; Nix, D.B.; Katoh, T.; Aoki, K.; Tiemeyer, M.; Wang, Y. Regulation of protein glycosylation and sorting by the Golgi matrix proteins GRASP55/65. Nat. Commun. 2013, 4, 1659. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.; Tiwari, N.; Dunlop, M.H.; Graham, M.; Liu, X.; Rothman, J.E. Membrane adhesion dictates Golgi stacking and cisternal morphology. Proc. Natl. Acad. Sci. USA 2014, 111, 1849–1854. [Google Scholar] [CrossRef] [PubMed]
- Mironov, A.A.; Beznoussenko, G.V. Molecular mechanisms responsible for formation of Golgi ribbon. Histol. Histopathol. 2011, 26, 117–133. [Google Scholar] [PubMed]
- Aw, R.; De Wachter, C.; Laukens, B.; De Rycke, R.; De Bruyne, M.; Bell, D.; Callewaert, N.; Polizzi, K.M. Knockout of RSN1, TVP18 or CSC1-2 causes perturbation of Golgi cisternae in Pichia pastoris. Traffic 2021, 22, 48–63. [Google Scholar] [CrossRef] [PubMed]
- Micaroni, M.; Perinetti, G.; Di Giandomenico, D.; Bianchi, K.; Spaar, A.; Mironov, A.A. Synchronous intra-Golgi transport induces the release of Ca2+ from the Golgi apparatus. Exp. Cell Res. 2010, 316, 2071–2086. [Google Scholar] [CrossRef] [PubMed]
- Micaroni, M.; Perinetti, G.; Berrie, C.P.; Mironov, A.A. The SPCA1 Ca2+ pump and intracellular membrane trafficking. Traffic 2010, 11, 1315–1333. [Google Scholar] [CrossRef]
- Ayscough, K.; Warren, G. Inhibition of protein synthesis disrupts the Golgi apparatus in the fission yeast, Schizosaccharomyces pombe. Yeast 1994, 10, 1–11. [Google Scholar] [CrossRef]
- Morin-Ganet, M.N.; Rambourg, A.; Deitz, S.B.; Franzusoff, A.; Kepes, F. Morphogenesis and dynamics of the yeast Golgi apparatus. Traffic 2000, 1, 56–68. [Google Scholar] [CrossRef]
- Mironov, A.A.; Banin, V.V.; Sesorova, I.S.; Dolgikh, V.V.; Luini, A.; Beznoussenko, G.V. Evolution of the endoplasmic reticulum and the Golgi complex. Adv. Exp. Med. Biol. 2007, 607, 61–72. [Google Scholar]
- Mironov, A.A., Jr.; Beznusenko, G.V.; Sesorova, I.S.; Banin, V.V. How to measure structures, or new stereology: III. Stereology in electron microscopy. Morfologija 2006, 129, 72–75. [Google Scholar]
- Tiwari, N.; Graham, M.; Liu, X.; Yue, X.; Zhu, L.; Meshram, D.; Choi, S.; Qian, Y.; Rothman, J.E.; Lee, I. Golgin45-Syntaxin5 Interaction Contributes to Structural Integrity of the Golgi Stack. Sci. Rep. 2019, 9, 12465. [Google Scholar] [CrossRef] [PubMed]
- Weidman, P.; Roth, R.; Heuser, J. Golgi membrane dynamics imaged by freeze-etch electron microscopy: Views of different membrane coatings involved in tubulation versus vesiculation. Cell 1993, 75, 123–133. [Google Scholar] [CrossRef]
- Jollivet, F.; Raposo, G.; Dimitrov, A.; Sougrat, R.; Goud, B.; Perez, F. Analysis of de novo Golgi complex formation after enzyme-based inactivation. Mol. Biol. Cell 2007, 18, 4637–4647. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Xiang, Y.; Wang, Y. GRASP55 and GRASP65 play complementary and essential roles in Golgi cisternal stacking. J. Cell Biol. 2010, 188, 237–251. [Google Scholar] [CrossRef] [PubMed]
- Ladinsky, M.S.; Wu, C.C.; McIntosh, S.; McIntosh, J.R.; Howell, K. Structure of the Golgi and distribution of reporter molecules at 20 °C reveals the complexity of the exit compartments. Mol. Biol. Cell 2002, 13, 2810–2825. [Google Scholar] [CrossRef]
- Perinetti, G.; Müller, T.; Spaar, A.; Polishchuk, R.; Luini, A.; Egner, A. Correlation of 4Pi- and electron microscopy to study transport through single Golgi stacks in living cells with super resolution. Traffic 2009, 10, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Vasile, E.; Krieger, M. Disruptions in Golgi structure and membrane traffic in a conditional lethal mammalian cell mutant are corrected by epsilon-COP. J. Cell Biol. 1994, 125, 1213–1224. [Google Scholar] [CrossRef]
- Cutrona, M.B.; Beznoussenko, G.V.; Fusella, A.; Martella, O.; Moral, P.; Mironov, A.A. Silencing of the mammalian Sar1 isoforms reveals COPII-independent protein sorting and transport. Traffic 2013, 14, 691–708. [Google Scholar] [CrossRef]
- Kreft, M.E.; Di Giandomenico, D.; Beznoussenko, G.V.; Resnik, N.; Mironov, A.A.; Jezernik, K. Golgi apparatus fragmentation as a mechanism responsible for uniform delivery of uroplakins to the apical plasma membrane of uroepithelial cells. Biol. Cell 2010, 102, 593–607. [Google Scholar] [CrossRef]
- Mironov, A.A.; Colanzi, A.; Silletta, M.G.; Fiucci, G.; Flati, S.; Fusella, A.; Polishchuk, R.S.; Mironov, A.A., Jr.; Di Tillio, D.; Weigert, R.; et al. Role of NAD+ and ADP-ribosylation in the maintenance of the Golgi structure. J. Cell Biol. 1997, 139, 1109–1118. [Google Scholar] [CrossRef]
- Beznoussenko, G.V.; Mironov, A.A. Correlative video-light-electron microscopy of mobile organelles. Methods Mol. Biol. 2015, 1270, 321–346. [Google Scholar] [PubMed]
- Burry, R.W.; Vandré, D.D.; Hayes, D.M. Silver enhancement of gold antibody probes in pre-embedding electron microscopic immunocytochemistry. J. Histochem. Cytochem. 1992, 40, 1849–1856. [Google Scholar] [CrossRef] [PubMed]
- Mironov, A.A., Jr.; Mironov, A.A. Estimation of subcellular organelle volume from ultrathin sections through centrioles with a discretized version of vertical rotator. J. Microsc. 1998, 192, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Lucocq, J.M.; Gawden-Bone, C. A stereological approach for estimation of cellular immunogold labeling and its spatial distribution in oriented sections using the rotator. J. Histochem. Cytochem. 2009, 57, 709–719. [Google Scholar] [CrossRef] [PubMed]















| Observation | Model | ||
|---|---|---|---|
| CMPM | KARM | Experiment | |
| Domain formation | Neutral | Desired | Shown |
| Increased concentration of SNAREs in the membrane of cargo domains | Neutral | Desired | Shown |
| Pores around cargo domains | Neutral | Desired | Shown |
| Cargo enrichment at the trans-side of the Golgi complex | Prohibitive | Possible | Shown |
| Stability of the number of medial cisternae during intra-Golgi transport | Prohibitive | Desired | Shown |
| Increased concentration of Golgi enzymes in COPI vesicles | Desired | Neutral | Not shown |
| Complete isolation of Golgi compartments in S. cerevisiae | Neutral | Prohibitive | Not shown |
| Temporal fusion of heterogenous Golgi compartment in S. cerevisiae | Neutral | Desired | Shown |
| Protocol | State of Golgi | Steps of Cargo Synchronisation | |||
|---|---|---|---|---|---|
| Complex | Accumulation of Cargo in ER Exit Sites and Emptying of Golgi | Duration of Cargo Pulse | Block of Delivery of Visible Cargoes to the Golgi | Beginning of Video Recording | |
| 1. Small pulse (40-15-40) | |||||
| A. E-40-15-40-small (the mini-wave) | Empty | 16 h at 40 °C | 15 min at 15 °C | Shift back to 40 °C | 2 min after shift to 40 °C |
| B. F-40-15-40-small (the mini-wave) | Full | 10 min after shift to 40 °C | |||
| 2. Large pulse (40-15-40) | |||||
| A. E-40-15-40-large (the maxi-wave) | Empty | 16 h at 40 °C | 2 h at 15 °C | Shift back to 40 °C | 2 min after shift to 40 °C |
| B. F-40-15-40-large (the maxi-wave) | Full | 10 min after shift to 40 °C | |||
| 3. Pulse (40-32-40) | |||||
| A. E-40-32-40-small (the 40-32 mini-wave) | Empty | 16 h at 40 °C | 5 min at 32 °C | Shift back to 40 °C | 2 min after shift to 40 °C |
| B. F-40-32-40-small | Full | 10 min after shift to 40 °C | |||
| 4. Emptying–pulse (32[CHM]-15-32[CHM]) | |||||
| A. E-CHM-15-CHM | Empty | 16 h at 40 °C, then CHM for 3 h at 32 °C | 2 h at 15 °C (without CHM) | Re-addition of CHM, shift back to 32 °C | 2 min after shift to 40 °C |
| B. F-CHM-15-CHM | Full | 10 min after shift to 40 °C | |||
| 5. ER accumulation–chase (40-32) | |||||
| A. E-40-32 (iFRAP) | Empty | 16 h at 40 °C | None | Bleaching of whole cell less the Golgi area | 5 min after shift to 32 °C |
| B. E-40-32 (piFRAP) | 3 min | ||||
| C. F-40-32 (iFRAP) | Full | None | 30 min after shift to 32 °C | ||
| D. F-40-32 (piFRAP) | 3 min | ||||
| 6. Steady-state iFRAP | |||||
| A. Steady-state iFRAP | Full | 16 h at 40 °C | None | Bleaching of whole cell less the Golgi area | Immediately after bleaching |
| B. Steady-state piFRAP | Full | 3 min | |||
| 7. Pulse–outcoming wave | |||||
| A. F-40-32-40 outcoming | Full | 16 h at 40 °C | 30 min at 32 °C | Shift to 40 °C for 30 min | 0, 2, 4, 6, 8, 10, 12 min after shift |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beznoussenko, G.V.; Kweon, H.-S.; Sesorova, I.S.; Mironov, A.A. Comparison of the Cisterna Maturation-Progression Model with the Kiss-and-Run Model of Intra-Golgi Transport: Role of Cisternal Pores and Cargo Domains. Int. J. Mol. Sci. 2022, 23, 3590. https://doi.org/10.3390/ijms23073590
Beznoussenko GV, Kweon H-S, Sesorova IS, Mironov AA. Comparison of the Cisterna Maturation-Progression Model with the Kiss-and-Run Model of Intra-Golgi Transport: Role of Cisternal Pores and Cargo Domains. International Journal of Molecular Sciences. 2022; 23(7):3590. https://doi.org/10.3390/ijms23073590
Chicago/Turabian StyleBeznoussenko, Galina V., Hee-Seok Kweon, Irina S. Sesorova, and Alexander A. Mironov. 2022. "Comparison of the Cisterna Maturation-Progression Model with the Kiss-and-Run Model of Intra-Golgi Transport: Role of Cisternal Pores and Cargo Domains" International Journal of Molecular Sciences 23, no. 7: 3590. https://doi.org/10.3390/ijms23073590
APA StyleBeznoussenko, G. V., Kweon, H.-S., Sesorova, I. S., & Mironov, A. A. (2022). Comparison of the Cisterna Maturation-Progression Model with the Kiss-and-Run Model of Intra-Golgi Transport: Role of Cisternal Pores and Cargo Domains. International Journal of Molecular Sciences, 23(7), 3590. https://doi.org/10.3390/ijms23073590