
Disruption of Membrane Integrity as a Molecular Initiating Event Determines the Toxicity of Polyhexamethylene Guanidine Phosphate Depending on the Routes of Exposure

 , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. Body and Organ Weights
2.2. Histopathological Evaluation
2.3. Serum Chemistry Analysis
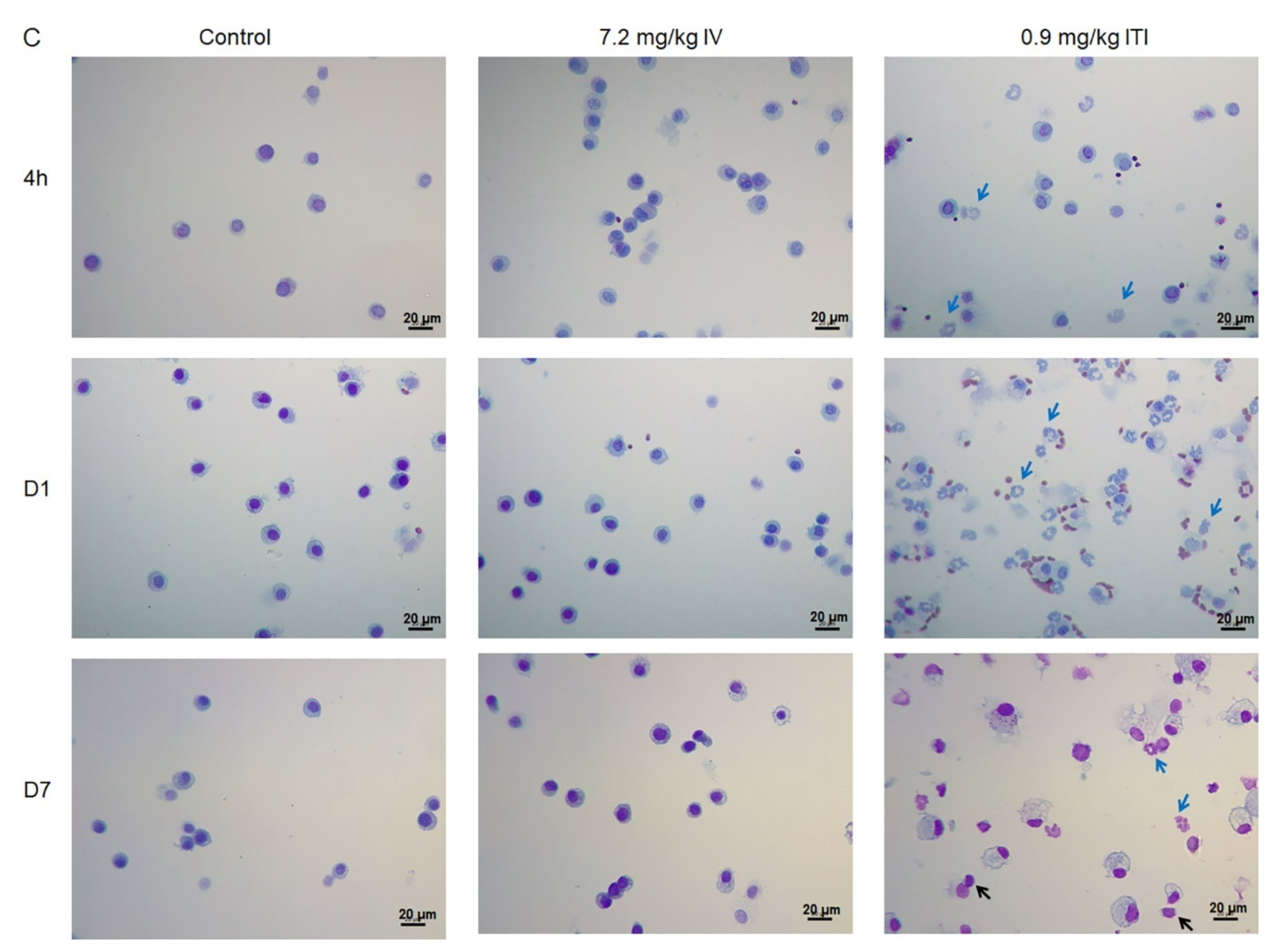
2.4. BAL Total and Differential Cell Counts
2.5. Changes in Cytokine Levels in BAL, the Lungs, and the Liver
2.6. Measurement of HMGB1 and sPLA2 in BAL Fluid
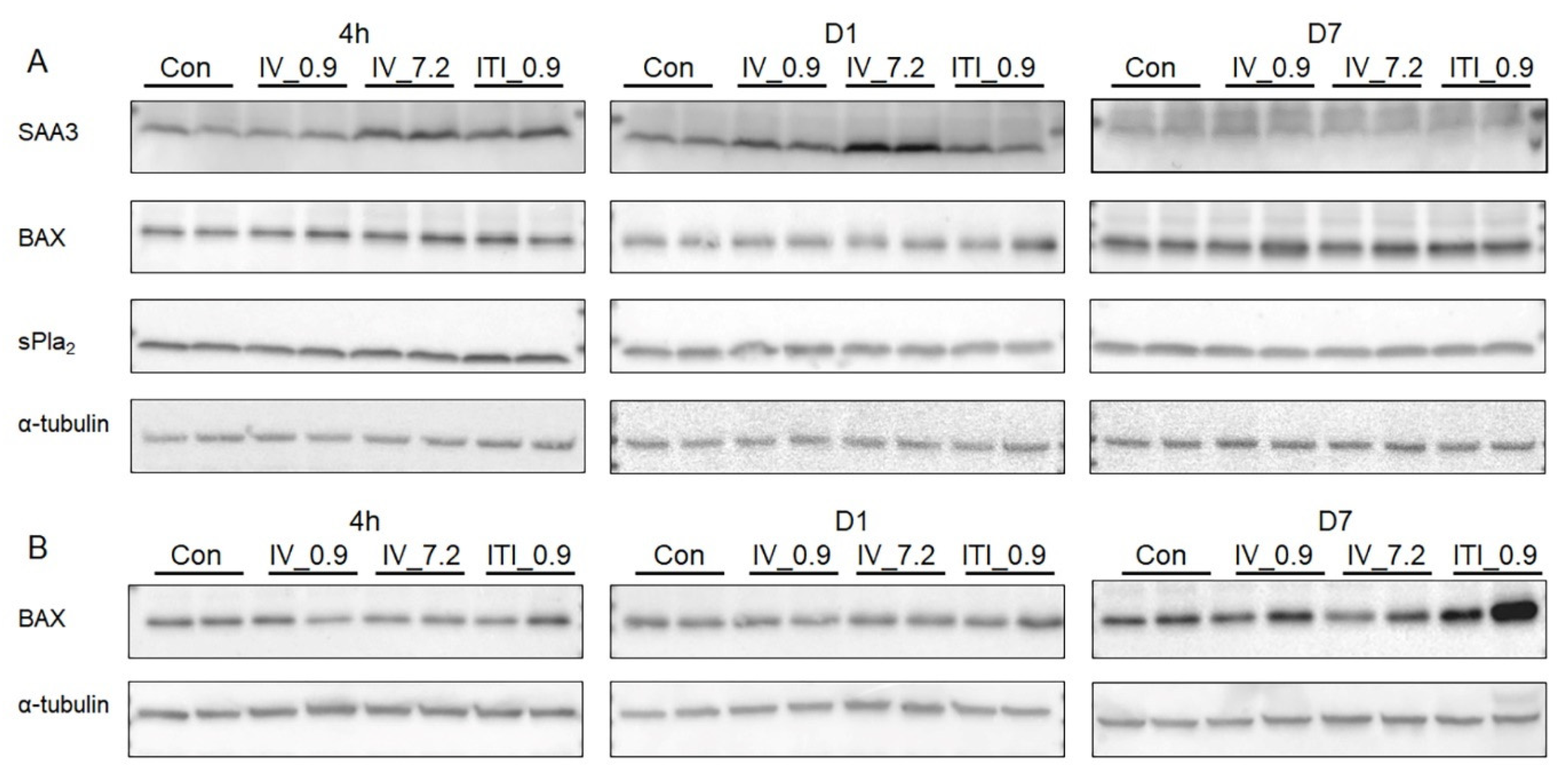
2.7. Measurement of mRNA and Protein Expression Levels in the Liver and Lung Tissues
2.8. Zebrafish Assay
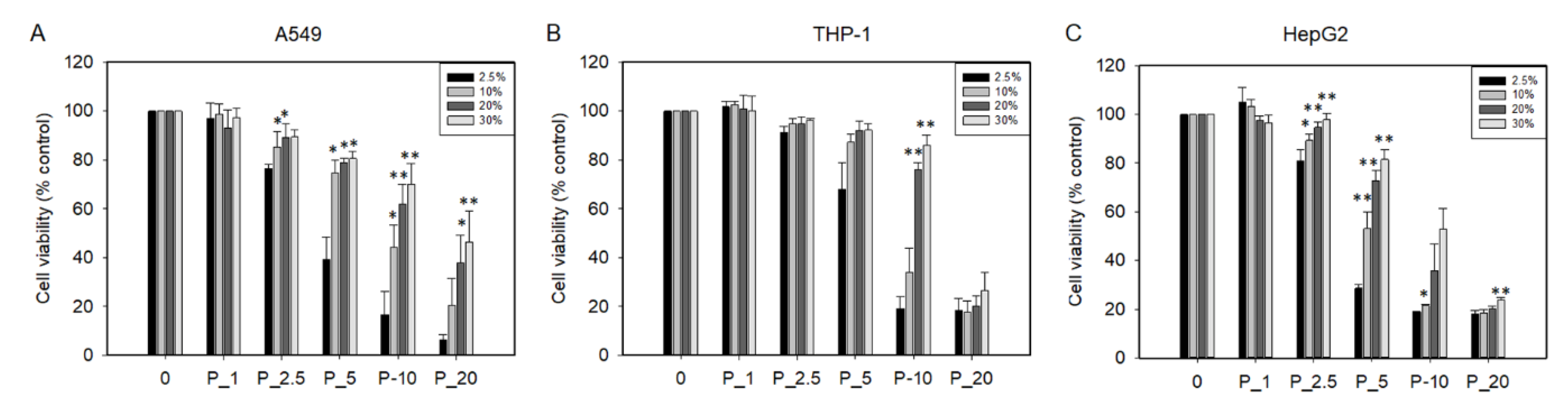
2.9. PHMG-P Induced Cytotoxicity at Various Serum Concentrations
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animal Experiment
4.3. Bronchoalveolar Lavage
4.4. Histopathological Examination
4.5. Serum Chemistry
4.6. Cytokine Measurement in BAL Fluid, the Lungs, and the Liver
4.7. Quantitative Real-Time PCR
4.8. Western Blot Analysis
4.9. Cell Culture
4.10. Zebrafish Assay
4.11. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Korea Centers for Disease Control and Prevention. Interim report of epidemiological investigation on lung injury with unknown cause in Korea. Public Health Wkly. Rep. KCDC 2011, 4, 817–832. [Google Scholar]
- Kim, S.; Park, S.; Jo, H.; Song, S.; Ham, S.; Yoon, C. Behavioral characteristics of polyhexamethyleneguanidine (PHMG) particles in aqueous solution and air when sprayed into an ultrasonic humidifier. Environ. Res. 2020, 182, 109078. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Jung, K.J.; Yang, M.J.; Han, S.C.; Lee, K. Assessment of acute and repeated pulmonary toxicities of oligo(2-(2-ethoxy)ethoxyethyl guanidium chloride in mice. Toxicol. Res. 2021, 37, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Lee, K.; Lee, E.J.; Lee, S.Y.; In, K.H.; Kim, H.K.; Kang, M.S. Humidifier disinfectant-associated interstitial lung disease in an animal model induced by polyhexamethylene guanidine aerosol. Am. J. Respir. Crit. Care Med. 2014, 190, 706–708. [Google Scholar] [CrossRef]
- Huh, J.W.; Hong, S.B.; Do, K.H.; Koo, H.J.; Jang, S.J.; Lee, M.S.; Paek, D.; Park, D.U.; Lim, C.M.; Koh, Y. Inhalation lung injury associated with humidifier disinfectants in adults. J. Korean Med. Sci. 2016, 31, 1857–1862. [Google Scholar] [CrossRef]
- Lee, E.; Lee, S.Y.; Hong, S.J. The past, present, and future of humidifier disinfectant-associated interstitial lung diseases in children. Clin. Exp. Pediatr. 2020, 63, 251–258. [Google Scholar] [CrossRef]
- Paek, D.; Koh, Y.; Park, D.U.; Cheong, H.K.; Do, K.H.; Lim, C.M.; Hong, S.J.; Kim, Y.H.; Leem, J.H.; Chung, K.H.; et al. Nationwide study of humidifier disinfectant lung injury in South Korea, 1994–2011. Incidence and Dose-Response Relationships. Ann. Am. Thorac. Soc. 2015, 12, 1813–1821. [Google Scholar] [CrossRef]
- Ministry of Environment Press Release, 26 May 2021. Available online: https://www.me.go.kr/home/web/board/read.do?pagerOffset=0&maxPageItems=10&maxIndexPages=10&searchKey=title&searchValue=%EA%B0%80%EC%8A%B5%EA%B8%B0&menuId=286&orgCd=&boardId=1455075&boardMasterId=1&boardCategoryId=&decorator= (accessed on 18 September 2021).
- KEITI Supporting Portal Site for Humidifier Disinfectant Victim. Available online: https://www.healthrelief.or.kr/home/content/stats01/view.do (accessed on 31 October 2021).
- Liu, J.; Li, X.W.; Guo, Y.W. Recent Advances in the Isolation, Synthesis and Biological Activity of Marine Guanidine Alkaloids. Mar. Drugs. 2017, 15, 324. [Google Scholar] [CrossRef]
- Ranaldi, G.; Marigliano, I.; Vespignani, I.; Perozzi, G.; Sambuy, Y. The effect of chitosan and other polycations on tight junction permeability in the human intestinal Caco-2 cell line. J. Nutr. Biochem. 2002, 13, 157–167. [Google Scholar] [CrossRef]
- Creppy, E.E.; Diallo, A.; Moukha, S.; Eklu-Gadegbeku, C.; Cros, D. Study of epigenetic properties of Poly(HexaMethylene Biguanide) hydrochloride (PHMB). Int. J. Environ. Res. Public Health 2014, 11, 8069–8092. [Google Scholar] [CrossRef]
- Song, J.; Jung, K.J.; Yoon, S.J.; Lee, K.; Kim, B. Polyhexamethyleneguanidine phosphate induces cytotoxicity through disruption of membrane integrity. Toxicology 2019, 414, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Gad, S.C. Routes in toxicology: An overview. J. Am. Coll. Toxicol. 1994, 13, 34–39. [Google Scholar] [CrossRef]
- Casarett, L.J.; Klaassen, C.D.; Amdur, M.O.; Doull, J. Casarett and Duoll’s Toxicology: The Basic Science of Poisons, 8th ed.; McGraw-Hill Education: New York, NY, USA, 2013; pp. 20–25. [Google Scholar]
- Vitt, A.; Sofrata, A.; Slizen, V.; Sugars, R.V.; Gustafsson, A.; Gudkova, E.I.; Kazeko, L.A.; Ramberg, P.; Buhlin, K. Antimicrobial activity of polyhexamethylene guanidine phosphate in comparison to chlorhexidine using the quantitative suspension method. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 36. [Google Scholar] [CrossRef] [PubMed]
- Muller, G.; Kramer, A. Effect of selected wound antiseptics on adult articular cartilage (bovine sesamoid bone) in the presence of Escherichia coli and Staphylococcus aureus. J. Orthop. Res. 2005, 23, 127–133. [Google Scholar] [CrossRef]
- Kari, G.; Rodeck, U.; Dicker, A.P. Zebrafish: An emerging model system for human disease and drug discovery. Clin. Pharmacol. Ther. 2007, 82, 70–80. [Google Scholar] [CrossRef]
- Hill, A.J.; Teraoka, H.; Heideman, W.; Peterson, R.E. Zebrafish as a model vertebrate for investigating chemical toxicity. Toxicol. Sci. 2005, 86, 6–19. [Google Scholar] [CrossRef]
- Kim, H.R.; Shin, D.Y.; Chung, K.H. The role of NF-kappaB signaling pathway in polyhexamethylene guanidine phosphate induced inflammatory response in mouse macrophage RAW264.7 cells. Toxicol. Lett. 2015, 233, 148–155. [Google Scholar] [CrossRef]
- Jin, S.W.; Lee, G.H.; Pham, H.T.; Choi, J.H.; Jeong, H.G. Polyhexamethylene Guanidine Phosphate Damages Tight Junctions and the F-Actin Architecture by Activating Calpain-1 via the P2RX7/Ca(2+) Signaling Pathway. Cells 2019, 9, 59. [Google Scholar] [CrossRef]
- Hong, S.; Bielinska, A.U.; Mecke, A.; Keszler, B.; Beals, J.L.; Shi, X.; Balogh, L.; Orr, B.G.; Baker, J.R., Jr.; Banaszak Holl, M.M. Interaction of poly(amidoamine) dendrimers with supported lipid bilayers and cells: Hole formation and the relation to transport. Bioconjug. Chem. 2004, 15, 774–782. [Google Scholar] [CrossRef]
- Paliienko, K.O.; Veklich, T.O.; Shatursky, O.Y.; Shkrabak, O.A.; Pastukhov, A.O.; Galkin, M.O.; Krisanova, N.V.; Chunikhin, A.J.; Rebriev, A.V.; Lysytsya, A.V.; et al. Membrane action of polyhexamethylene guanidine hydrochloride revealed on smooth muscle cells, nerve tissue and rat blood platelets: A biocide driven pore-formation in phospholipid bilayers. Toxicol. Vitro. 2019, 60, 389–399. [Google Scholar] [CrossRef]
- Andrews, N.W.; Almeida, P.E.; Corrotte, M. Damage control: Cellular mechanisms of plasma membrane repair. Trends Cell. Biol. 2014, 24, 734–742. [Google Scholar] [CrossRef] [PubMed]
- Draeger, A.; Monastyrskaya, K.; Babiychuk, E.B. Plasma membrane repair and cellular damage control: The annexin survival kit. Biochem. Pharmacol. 2011, 81, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Mellgren, R.L. A new twist on plasma membrane repair. Commun. Integr. Biol. 2011, 4, 198–200. [Google Scholar] [CrossRef] [PubMed]
- Busher, J.T. Serum Albumin and Globulin. In Clinical Methods: The History, Physical, and Laboratory Examinations; Walker, H.K., Hall, W.D., Hurst, J.W., Eds.; Butterworths: Boston, MA, USA, 1990. [Google Scholar]
- Zaias, J.; Mineau, M.; Cray, C.; Yoon, D.; Altman, N.H. Reference values for serum proteins of common laboratory rodent strains. J. Am. Assoc. Lab. Anim. Sci. 2009, 48, 387–390. [Google Scholar]
- Moliva, J.I.; Rajaram, M.V.; Sidiki, S.; Sasindran, S.J.; Guirado, E.; Pan, X.J.; Wang, S.H.; Ross, P., Jr.; Lafuse, W.P.; Schlesinger, L.S.; et al. Molecular composition of the alveolar lining fluid in the aging lung. Age 2014, 36, 9633. [Google Scholar] [CrossRef]
- Kim, S.; Paek, D. Humidifier disinfectant disaster: What is known and what needs to be clarified. Environ. Health Toxicol. 2016, 31, e2016025. [Google Scholar] [CrossRef]
- Asiedu-Gyekye, I.J.; Mahmood, S.A.; Awortwe, C.; Nyarko, A.K. A preliminary safety evaluation of polyhexamethylene guanidine hydrochloride. Int. J. Toxicol. 2014, 33, 523–531. [Google Scholar] [CrossRef]
- Song, J.A.; Park, H.J.; Yang, M.J.; Jung, K.J.; Yang, H.S.; Song, C.W.; Lee, K. Polyhexamethyleneguanidine phosphate induces severe lung inflammation, fibrosis, and thymic atrophy. Food Chem. Toxicol. 2014, 69, 267–275. [Google Scholar] [CrossRef]
- Kratz, F.; Elsadek, B. Clinical impact of serum proteins on drug delivery. J. Control. Release 2012, 161, 429–445. [Google Scholar] [CrossRef]
- Otagiri, M. A molecular functional study on the interactions of drugs with plasma proteins. Drug Metab. Pharmacokinet. 2005, 20, 309–323. [Google Scholar] [CrossRef]
- Kragh-Hansen, U. Molecular aspects of ligand binding to serum albumin. Pharmacol. Rev. 1981, 33, 17–53. [Google Scholar] [PubMed]
- Bewersdorff, T.; Vonnemann, J.; Kanik, A.; Haag, R.; Haase, A. The influence of surface charge on serum protein interaction and cellular uptake: Studies with dendritic polyglycerols and dendritic polyglycerol-coated gold nanoparticles. Int. J. Nanomed. 2017, 12, 2001–2019. [Google Scholar] [CrossRef] [PubMed]
- Ostapenko, Y.N.; Brusin, K.M.; Zobnin, Y.V.; Shchupak, A.Y.; Vishnevetskiy, M.K.; Sentsov, V.G.; Novikova, O.V.; Alekseenko, S.A.; Lebed’ko, O.A.; Puchkov, Y.B. Acute cholestatic liver injury caused by polyhexamethyleneguanidine hydrochloride admixed to ethyl alcohol. Clin. Toxicol. 2011, 49, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Shim, H.E.; Lee, J.Y.; Lee, C.H.; Mushtaq, S.; Song, H.Y.; Song, L.; Choi, S.J.; Lee, K.; Jeon, J. Quantification of inhaled aerosol particles composed of toxic household disinfectant using radioanalytical method. Chemosphere 2018, 207, 649–654. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Seo, D.S. Toxicity of humidifier disinfectant polyhexamethylene guanidine hydrochloride by two-week whole body-inhalation exposure in rats. J. Toxicol. Pathol. 2020, 33, 265–277. [Google Scholar] [CrossRef]
- Giannini, E.G.; Testa, R.; Savarino, V. Liver enzyme alteration: A guide for clinicians. CMAJ 2005, 172, 367–379. [Google Scholar] [CrossRef]
- Ather, J.L.; Dienz, O.; Boyson, J.E.; Anathy, V.; Amiel, E.; Poynter, M.E. Serum Amyloid A3 is required for normal lung development and survival following influenza infection. Sci. Rep. 2018, 8, 16571. [Google Scholar] [CrossRef] [PubMed]
- Bjorkman, L.; Karlsson, J.; Karlsson, A.; Rabiet, M.J.; Boulay, F.; Fu, H.; Bylund, J.; Dahlgren, C. Serum amyloid A mediates human neutrophil production of reactive oxygen species through a receptor independent of formyl peptide receptor like-1. J. Leukoc. Biol. 2008, 83, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Uhlar, C.M.; Whitehead, A.S. Serum amyloid A, the major vertebrate acute-phase reactant. Eur. J. Biochem. 1999, 265, 501–523. [Google Scholar] [CrossRef]
- Saber, A.T.; Lamson, J.S.; Jacobsen, N.R.; Ravn-Haren, G.; Hougaard, K.S.; Nyendi, A.N.; Wahlberg, P.; Madsen, A.M.; Jackson, P.; Wallin, H.; et al. Particle-induced pulmonary acute phase response correlates with neutrophil influx linking inhaled particles and cardiovascular risk. PLoS ONE 2013, 8, e69020. [Google Scholar] [CrossRef]
- Jorgensen, K.; Davidsen, J.; Mouritsen, O.G. Biophysical mechanisms of phospholipase A2 activation and their use in liposome-based drug delivery. FEBS Lett. 2002, 531, 23–27. [Google Scholar] [CrossRef]
- Atsumi, G.; Murakami, M.; Tajima, M.; Shimbara, S.; Hara, N.; Kudo, I. The perturbed membrane of cells undergoing apoptosis is susceptible to type II secretory phospholipase A2 to liberate arachidonic acid. Biochim. Biophys. Acta 1997, 1349, 43–54. [Google Scholar] [CrossRef]
- Nelson, J.; Gibbons, E.; Pickett, K.R.; Streeter, M.; Warcup, A.O.; Yeung, C.H.; Judd, A.M.; Bell, J.D. Relationship between membrane permeability and specificity of human secretory phospholipase A(2) isoforms during cell death. Biochim. Biophys. Acta 2011, 1808, 1913–1920. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bezzine, S.; Bollinger, J.G.; Singer, A.G.; Veatch, S.L.; Keller, S.L.; Gelb, M.H. On the binding preference of human groups IIA and X phospholipases A2 for membranes with anionic phospholipids. J. Biol. Chem. 2002, 277, 48523–48534. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.K.; Farnbach, A.R.; Harris, F.M.; Hawes, A.C.; Jackson, L.R.; Judd, A.M.; Vest, R.S.; Sanchez, S.; Bell, J.D. Mechanisms by which intracellular calcium induces susceptibility to secretory phospholipase A2 in human erythrocytes. J. Biol. Chem. 2001, 276, 22732–22741. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day | Control | 0.9 mg/kg IV | 7.2 mg/kg IV | 0.9 mg/kg ITI |
|---|---|---|---|---|
| −3 | 21.80 ± 0.75 a | 22.04 ± 0.63 | 21.95 ± 0.80 | 21.80 ± 0.68 |
| 1 | 22.68 ± 0.86 | 23.19 ± 0.76 | 20.32 ± 1.12 ** | 19.49 ± 0.71 ** |
| 2 | 22.02 ± 0.85 | 22.48 ± 0.71 | 19.87 ± 1.17 ** | 17.36 ± 0.65 ** |
| 4 | 22.71 ± 0.90 | 23.53 ± 0.80 | 20.16 ± 1.29 ** | 17.07 ± 0.78 ** |
| 7 | 23.04 ± 0.97 | 24.13 ± 0.83 | 20.66 ± 0.97 ** | 18.64 ± 1.60 ** |
| 4 h | Control | 0.9 mg/kg IV | 7.2 mg/kg IV | 0.9 mg/kg ITI |
|---|---|---|---|---|
| BUN a (mg/dL) | 22.80 ± 1.74 | 20.30 ± 2.82 | 26.30 ± 3.11 * | 23.20 ± 1.67 |
| CREA (mg/dL) | 0.28 ± 0.04 | 0.29 ± 0.02 | 0.29 ± 0.03 | 0.23 ± 0.02 * |
| TP (g/dL) | 5.11 ± 0.18 | 4.87 ± 0.22 | 5.85 ± 0.28 ** | 5.54 ± 0.38 * |
| ALB (g/dL) | 3.31 ± 0.11 | 3.15 ± 0.19 | 3.67 ± 0.16 ** | 3.52 ± 0.19 * |
| AST (IU/L) | 55.60 ± 15.73 | 55.20 ± 4.77 | 92.10 ± 8.20 ** | 108.90 ± 31.72 ** |
| ALT (IU/L) | 28.80 ± 5.30 | 29.20 ± 3.7 | 45.20 ± 11.35 * | 62.70 ± 34.23 |
| GGT (IU/L) | 1.85 ± 0.27 | 1.82 ± 0.35 | 2.06 ± 0.37 | 2.09 ± 0.59 |
| ALP (IU/L) | 485.10 ± 36.46 | 491.90 ± 39.36 | 541.50 ± 45.88 * | 483.90 ± 49.16 |
| TCHO (mg/dL) | 84.10 ± 4.59 | 82.90 ± 5.99 | 98.10 ± 7.82** | 88.80 ± 5.36 |
| TG (mg/dL) | 46.00 ± 7.12 | 66.80 ± 20.17 | 164.70 ± 94.69 * | 77.70 ± 13.22 ** |
| D1 | Control | 0.9 mg/kg IV | 7.2 mg/kg IV | 0.9 mg/kg ITI |
| BUN (mg/dL) | 27.20 ± 3.98 | 28.60 ± 4.30 | 20.20 ± 2.68 ** | 21.50 ± 2.46 * |
| CREA (mg/dL) | 0.33 ± 0.04 | 0.34 ± 0.04 | 0.30 ± 0.03 | 0.28 ± 0.02 |
| TP (g/dL) | 5.34 ± 0.33 | 5.30 ± 0.30 | 5.60 ± 0.25 | 5.51 ± 0.30 |
| ALB (g/dL) | 3.45 ± 0.16 | 3.42 ± 0.12 | 3.63 ± 0.19 | 3.59 ± 0.16 |
| AST (IU/L) | 48.20 ± 6.31 | 45.40 ± 4.13 | 53.8 ± 5.94 | 52.7 ± 4.79 |
| ALT (IU/L) | 28.10 ± 4.39 | 28.90 ± 3.43 | 23.30 ± 1.45 * | 23.10 ± 3.50 * |
| GGT (IU/L) | 1.96 ± 0.14 | 1.93 ± 0.40 | 2.08 ± 0.29 | 1.92 ± 0.34 |
| ALP (IU/L) | 485.30 ± 57.82 | 534.80 ± 51.46 | 457.60 ± 52.09 | 480.80 ± 51.26 |
| TCHO (mg/dL) | 85.40 ± 4.90 | 88.60 ± 5.39 | 101.60 ± 9.48 ** | 95.80 ± 4.76 ** |
| TG (mg/dL) | 66.50 ± 15.83 | 64.80 ± 13.22 | 46.50 ± 10.52 * | 57.00 ± 19.31 |
| D7 | Control | 0.9 mg/kg IV | 7.2 mg/kg IV | 0.9 mg/kg ITI |
| BUN (mg/dL) | 30.00 ± 4.13 | 29.80 ± 3.66 | 35.30 ± 6.82 | 30.30 ± 4.24 |
| CREA (mg/dL) | 0.31 ± 0.04 | 0.31 ± 0.03 | 0.32 ± 0.03 | 0.29 ± 0.03 |
| TP (g/dL) | 5.53 ± 0.15 | 5.59 ± 0.29 | 5.51 ± 0.30 | 5.51 ± 0.34 |
| ALB (g/dL) | 3.41 ± 0.09 | 3.39 ± 0.15 | 3.47 ± 0.18 | 3.49 ± 0.18 |
| AST (IU/L) | 48.20 ± 2.77 | 47.40 ± 4.58 | 57.40 ± 6.51 * | 61.30 ± 10.44 ** |
| ALT (IU/L) | 31.10 ± 1.78 | 32.00 ± 5.33 | 31.60 ± 3.60 | 38.60 ± 8.29 |
| GGT (IU/L) | 1.66 ± 0.32 | 1.77 ± 0.21 | 1.91 ± 0.32 | 1.78 ± 0.19 |
| ALP (IU/L) | 426.90 ± 51.04 | 450.80 ± 49.71 | 405.80 ± 30.36 | 376.90 ± 40.01 |
| TCHO (mg/dL) | 84.89 ± 7.72 | 83.22 ± 6.40 | 93.33 ± 6.89 | 96.11 ± 8.59 * |
| TG (mg/dL) | 94.47 ± 24.30 | 72.33 ± 12.31 | 90.33 ± 31.97 | 80.58 ± 22.88 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, J.; Jung, K.-J.; Yang, M.-J.; Kim, W.; Lee, B.-S.; Choe, S.-K.; Kim, S.-J.; Hwang, J.-H. Disruption of Membrane Integrity as a Molecular Initiating Event Determines the Toxicity of Polyhexamethylene Guanidine Phosphate Depending on the Routes of Exposure. Int. J. Mol. Sci. 2022, 23, 3289. https://doi.org/10.3390/ijms23063289
Song J, Jung K-J, Yang M-J, Kim W, Lee B-S, Choe S-K, Kim S-J, Hwang J-H. Disruption of Membrane Integrity as a Molecular Initiating Event Determines the Toxicity of Polyhexamethylene Guanidine Phosphate Depending on the Routes of Exposure. International Journal of Molecular Sciences. 2022; 23(6):3289. https://doi.org/10.3390/ijms23063289
Chicago/Turabian StyleSong, Jeongah, Kyung-Jin Jung, Mi-Jin Yang, Woojin Kim, Byoung-Seok Lee, Seong-Kyu Choe, Seong-Jin Kim, and Jeong-Ho Hwang. 2022. "Disruption of Membrane Integrity as a Molecular Initiating Event Determines the Toxicity of Polyhexamethylene Guanidine Phosphate Depending on the Routes of Exposure" International Journal of Molecular Sciences 23, no. 6: 3289. https://doi.org/10.3390/ijms23063289
APA StyleSong, J., Jung, K.-J., Yang, M.-J., Kim, W., Lee, B.-S., Choe, S.-K., Kim, S.-J., & Hwang, J.-H. (2022). Disruption of Membrane Integrity as a Molecular Initiating Event Determines the Toxicity of Polyhexamethylene Guanidine Phosphate Depending on the Routes of Exposure. International Journal of Molecular Sciences, 23(6), 3289. https://doi.org/10.3390/ijms23063289