
A Comparative Transcriptomic Analysis Reveals That HSP90AB1 Is Involved in the Immune and Inflammatory Responses to Porcine Deltacoronavirus Infection

 , ,
, ,
Abstract
:1. Introduction
2. Results
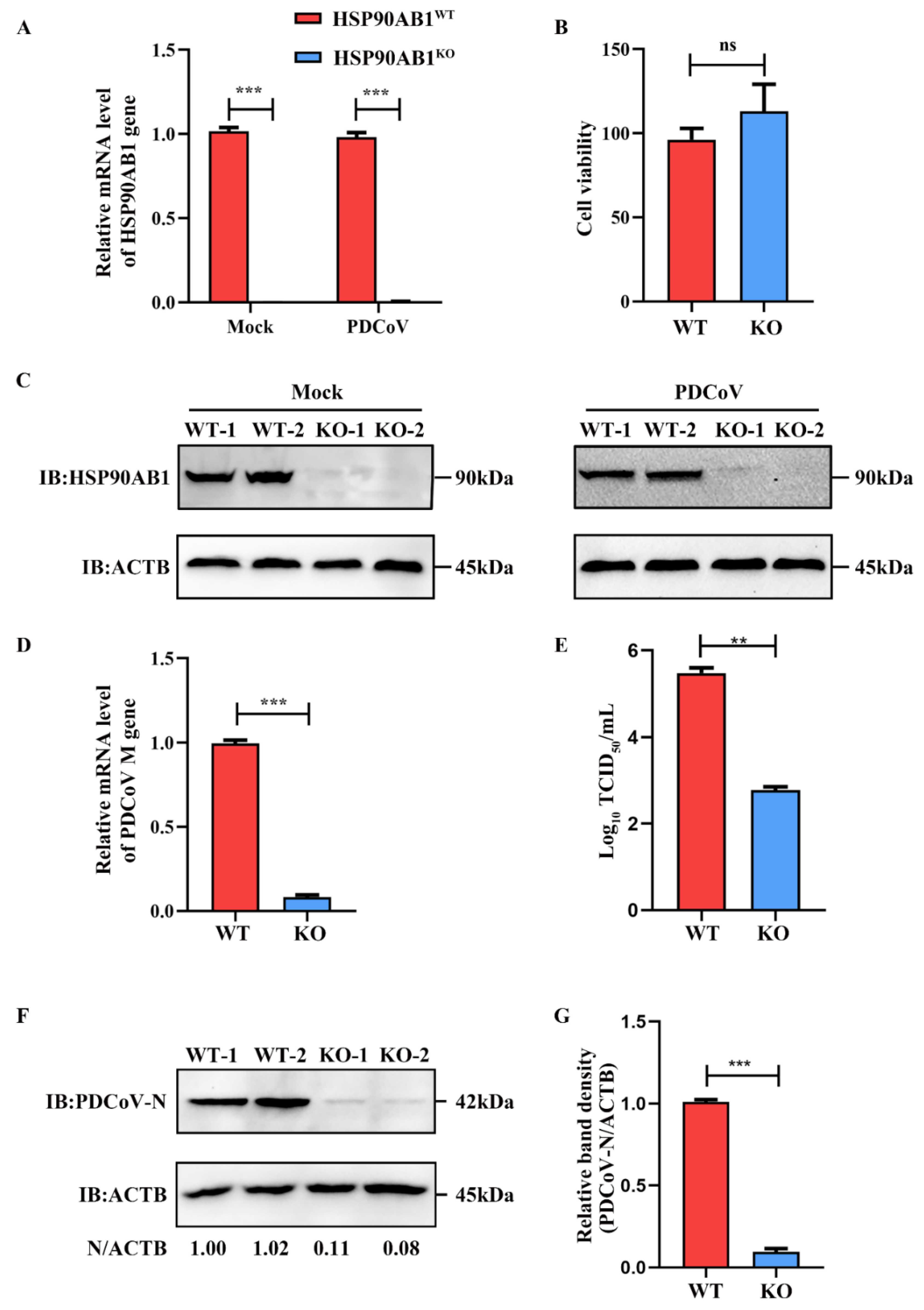
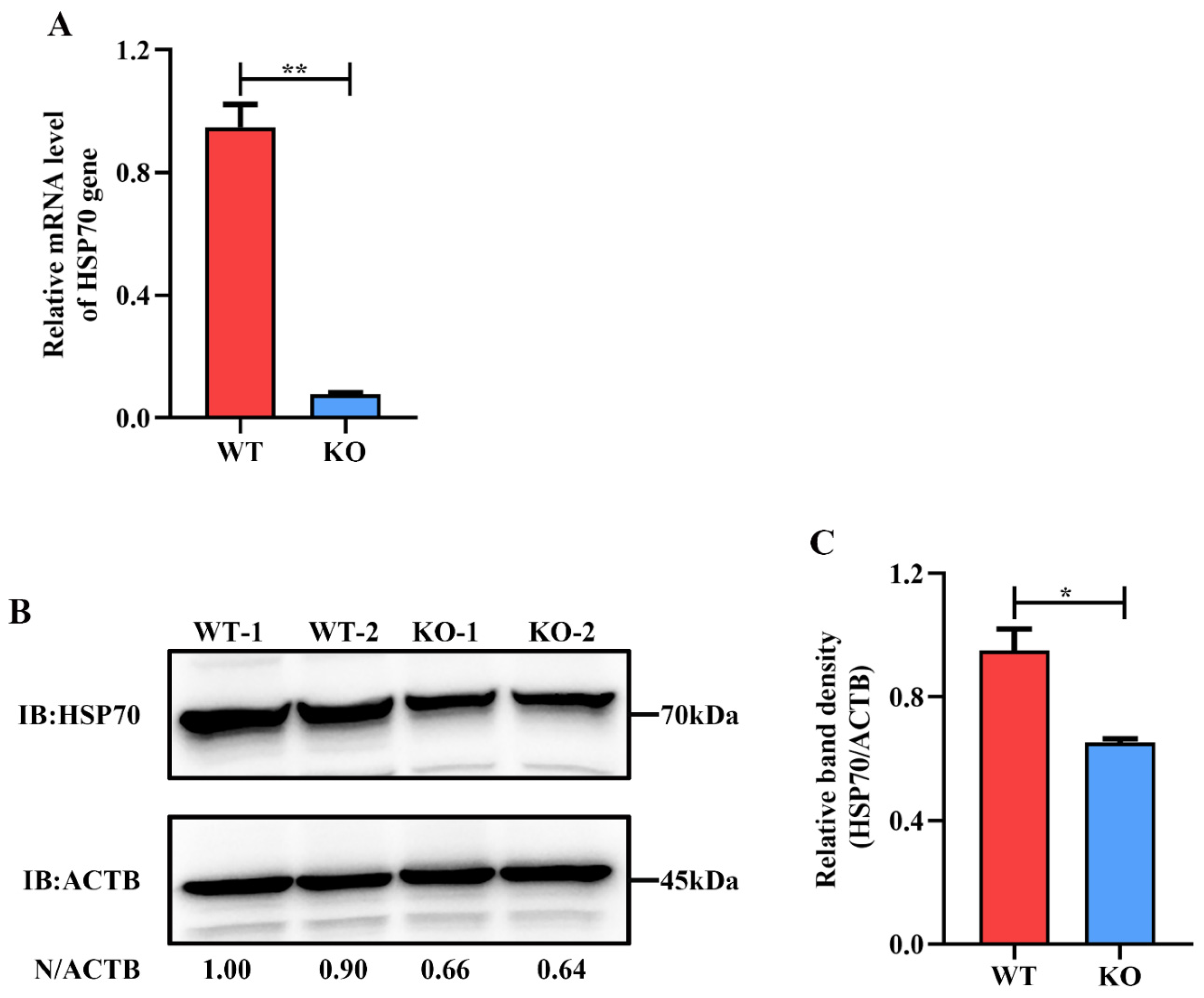
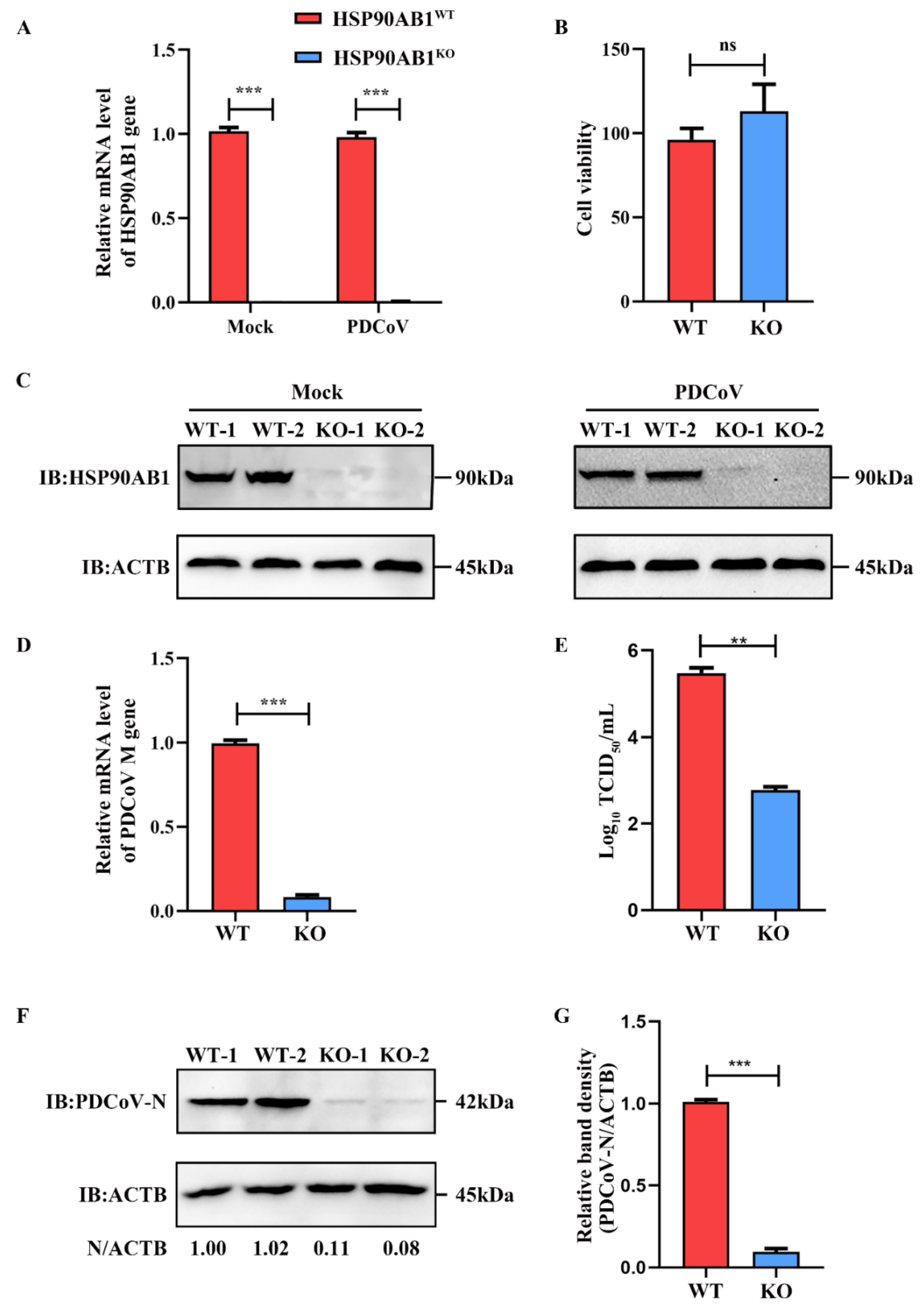
2.1. HSP90AB1 Knockout Reduces PDCoV Infection
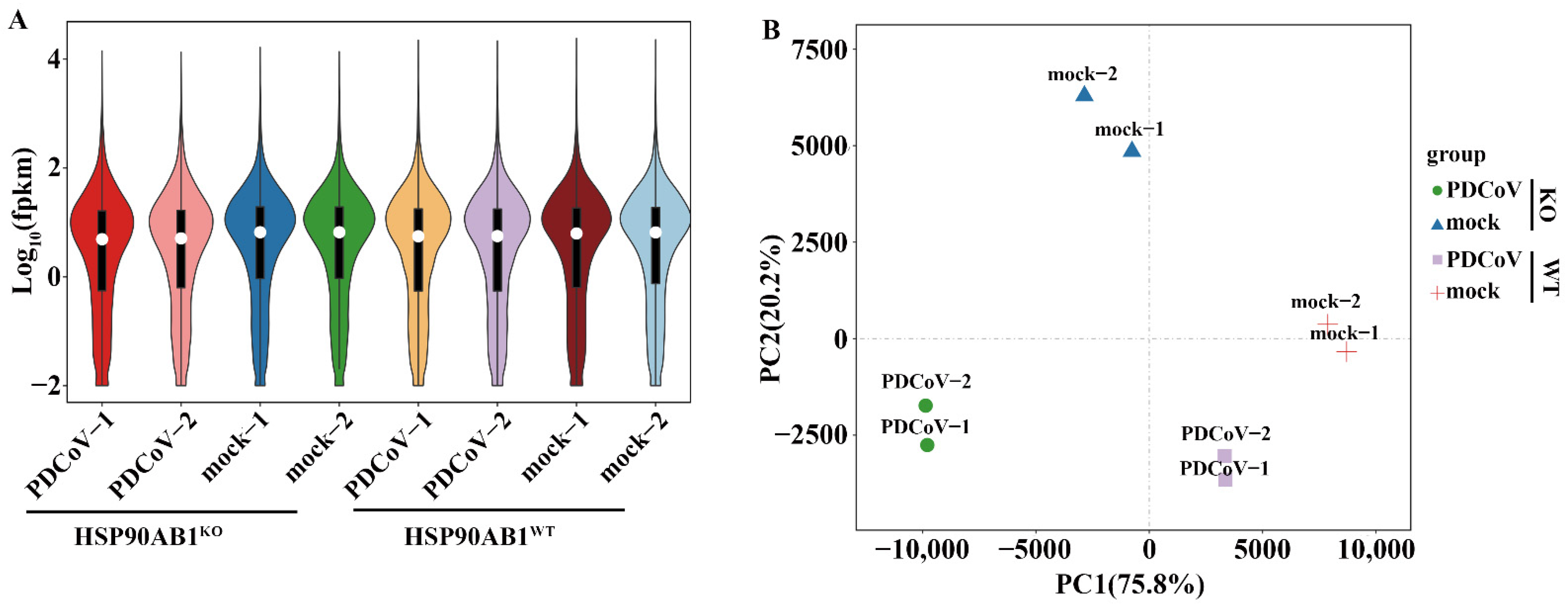
2.2. Quality Control and Mapping of RNA Sequencing Results
2.3. Identification and Analysis of DEGs
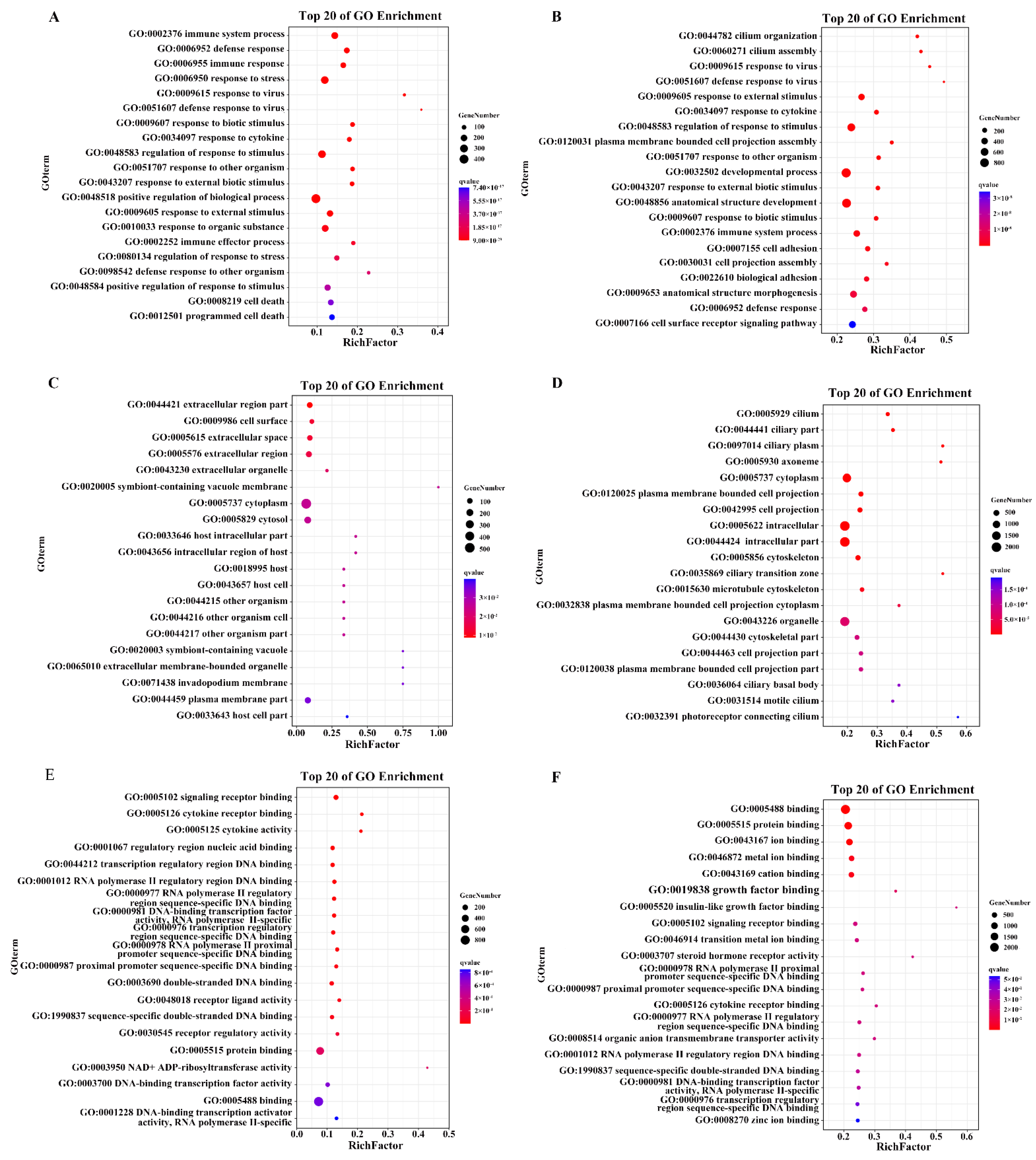
2.4. GO Enrichment Analysis of the DEGs upon PDCoV Infection
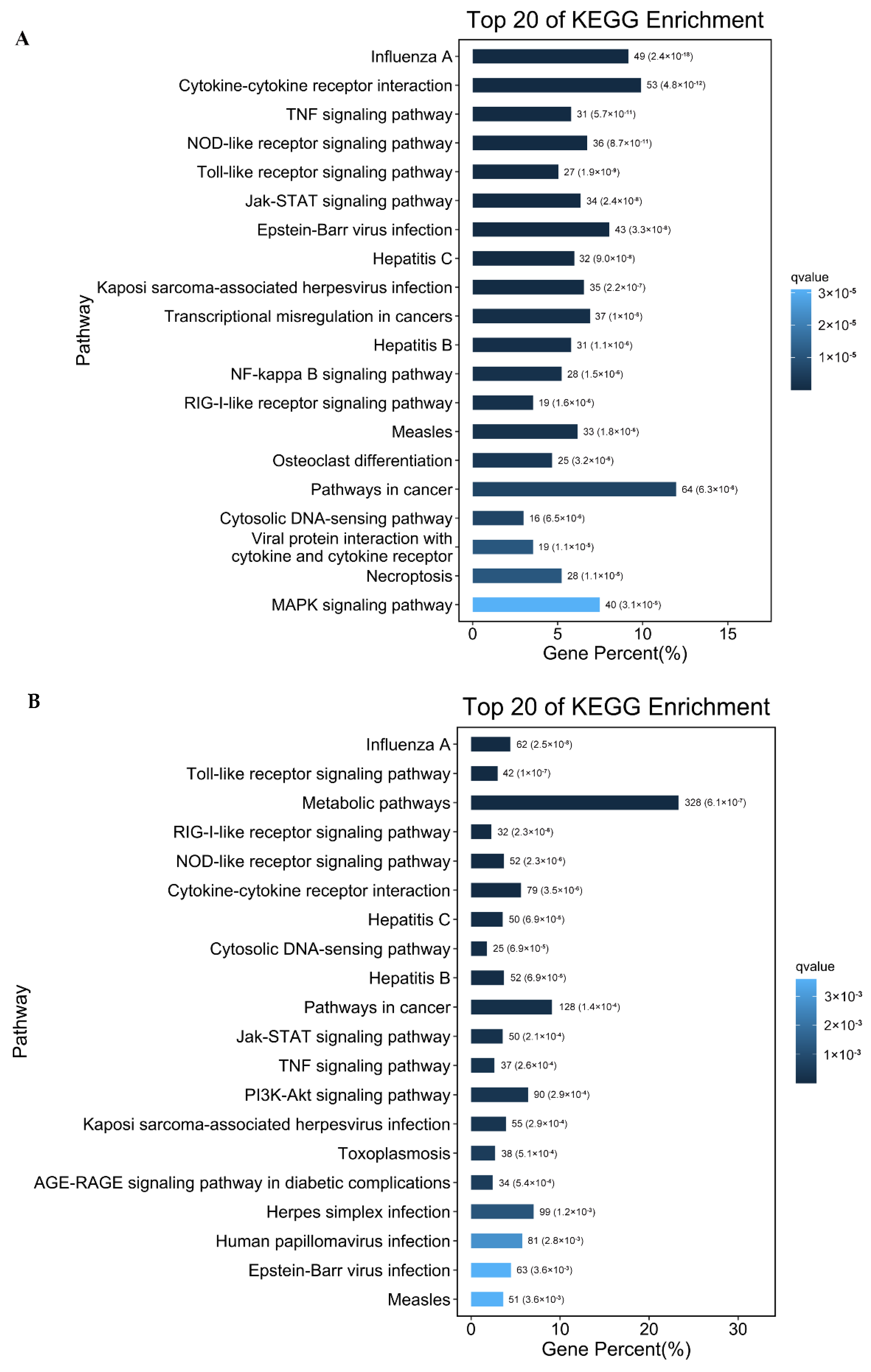
2.5. KEGG Pathway Analysis of the DEGs upon PDCoV Infection
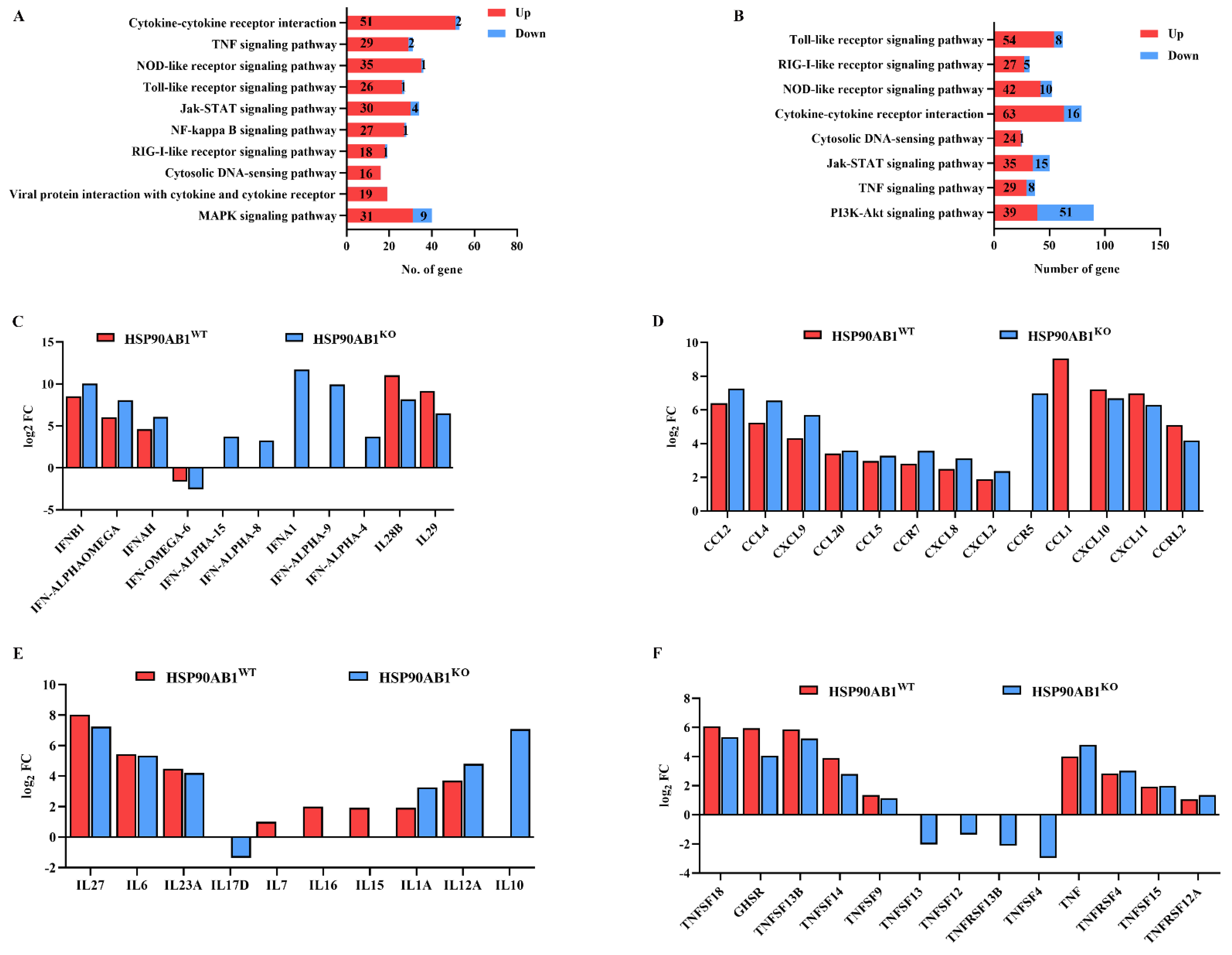
2.6. Expression of DEGs in Immune- and Inflammatory-Related Pathways
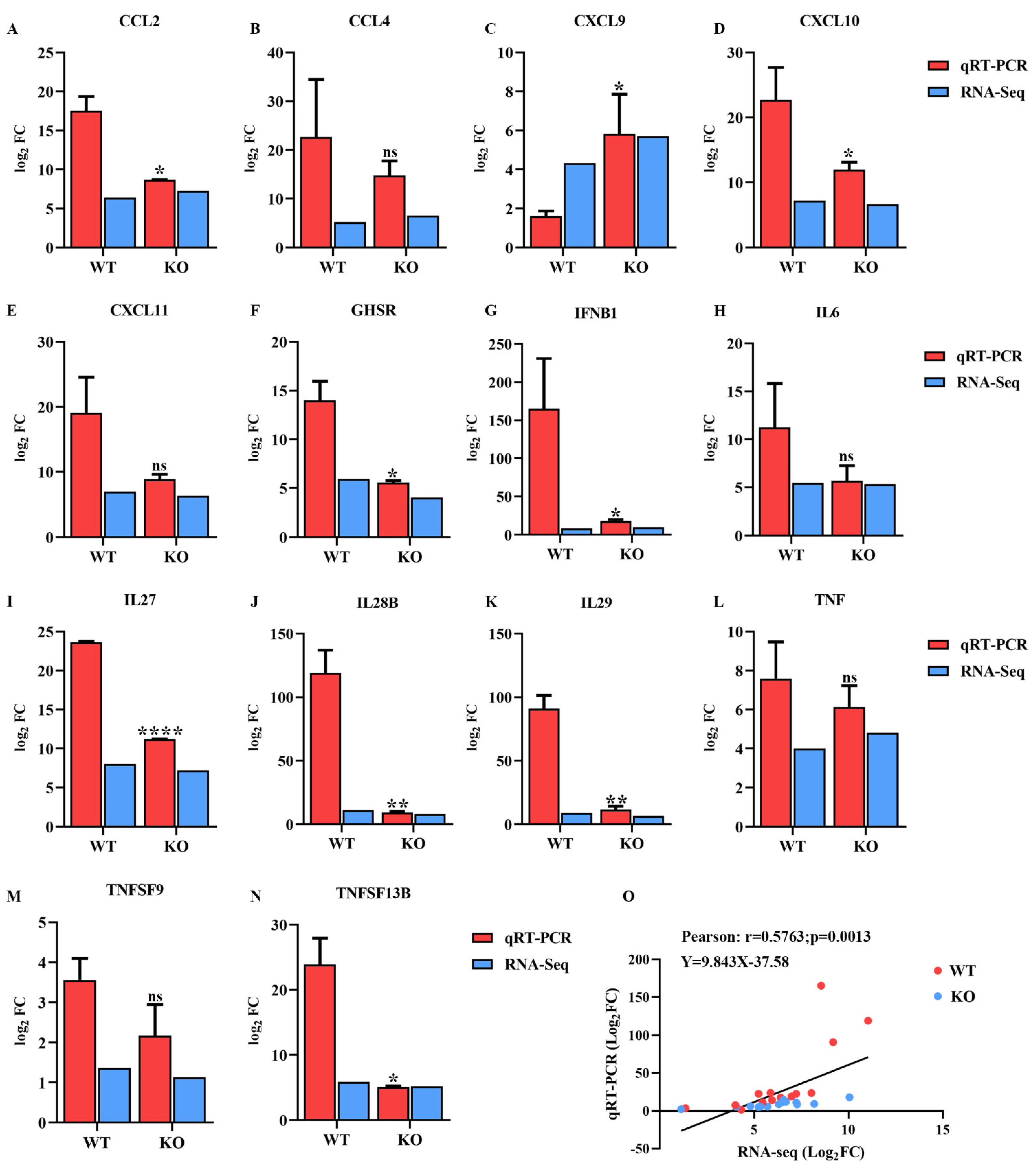
2.7. Validation of Transcriptomic Data Using qRT-PCR
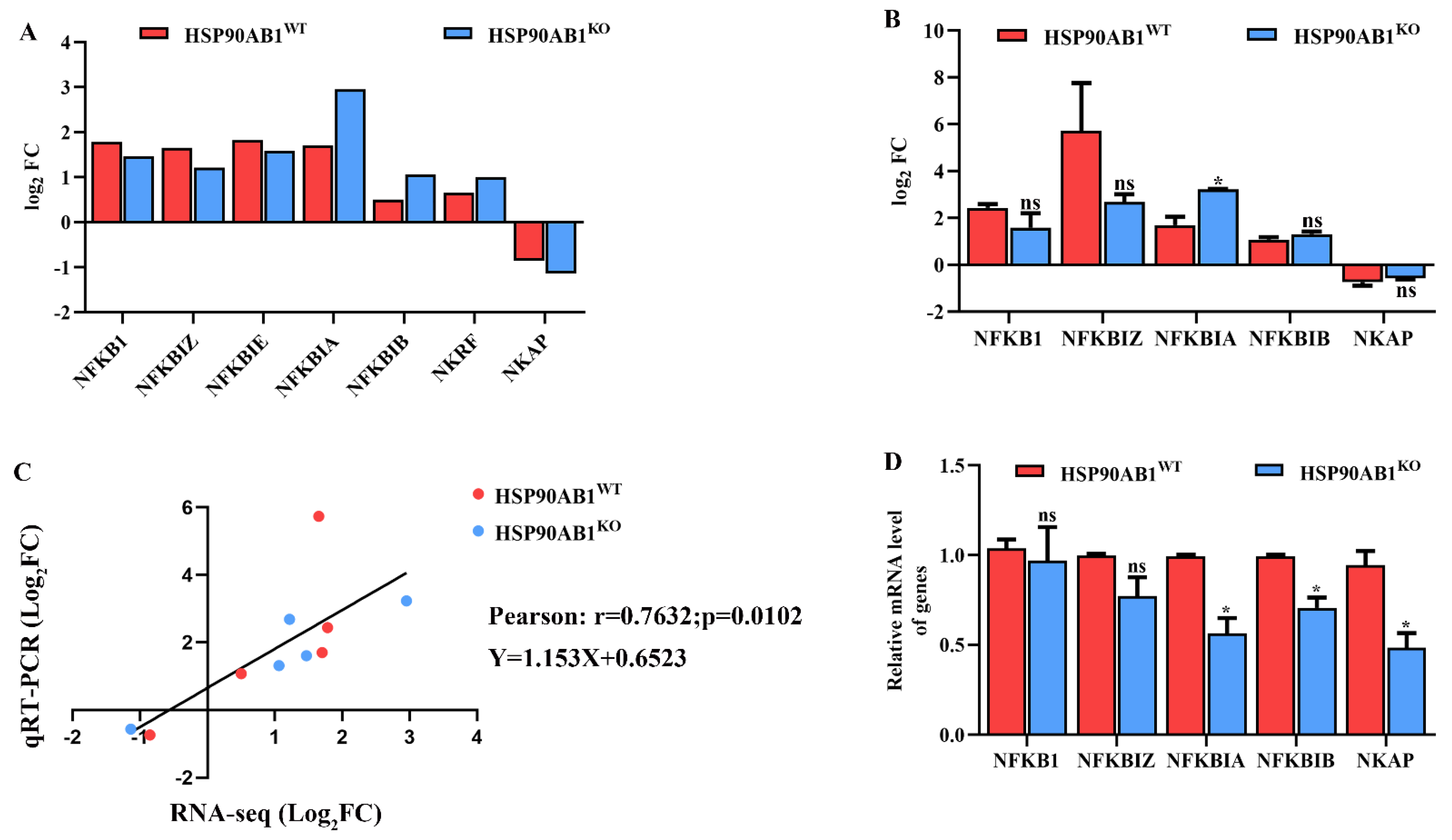
2.8. HSP90AB1 Knockout Modulates NF-κB Activity
3. Discussion
4. Materials and Methods
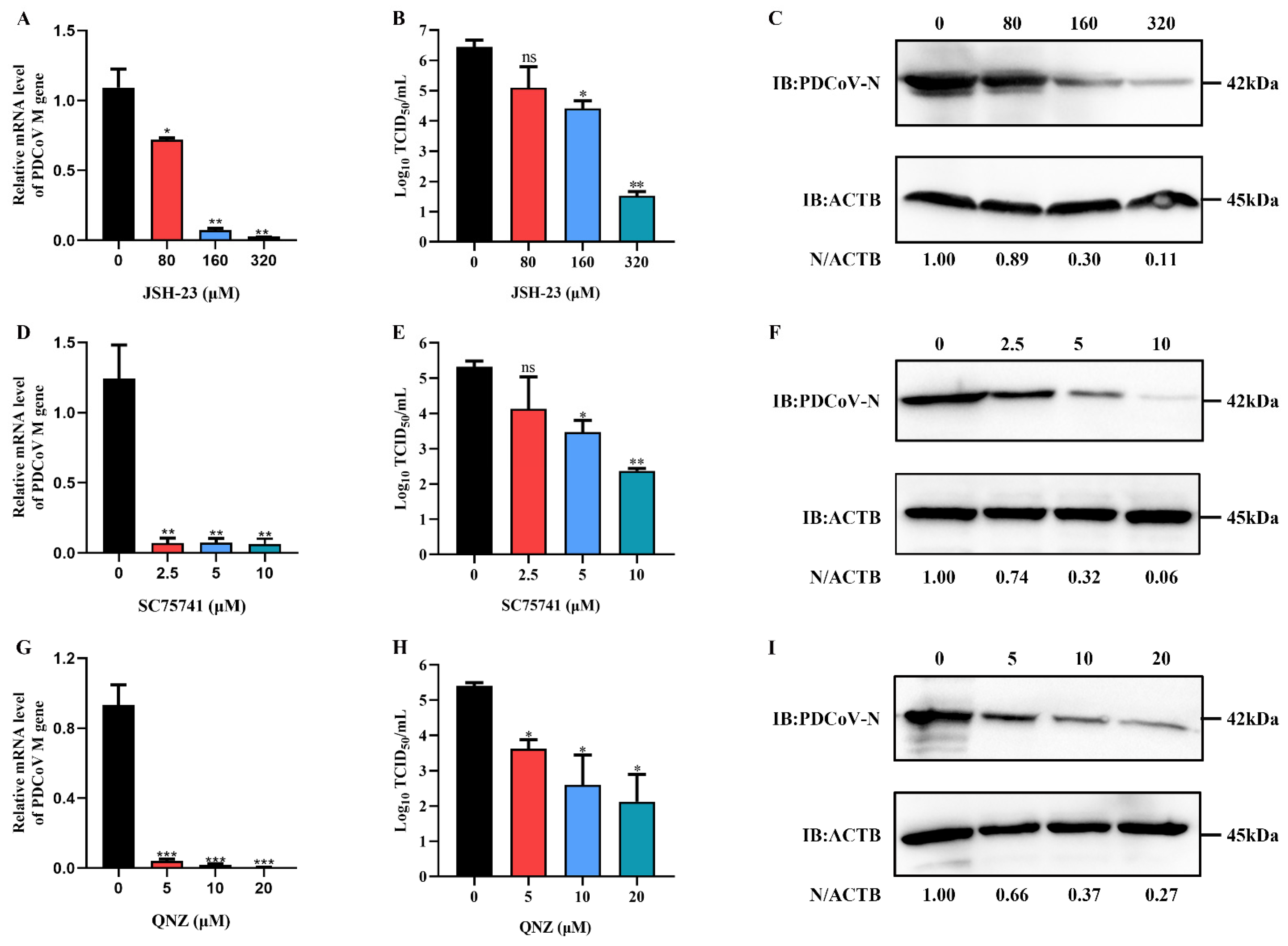
4.1. Cells, Virus, Inhibitors and Antibodies
4.2. Generation of HSP90AB1 Knockout Cells Using the CRISPR/Cas9 System
4.3. Sample Preparation and RNA Extraction
4.4. cDNA Library Preparation and Sequencing
4.5. Read Quality Control and Mapping
4.6. Correlation Analysis of Samples
4.7. Differentially Expressed Gene (DEG) Analysis
4.8. qRT-PCR
4.9. Western Blotting
4.10. TCID50
4.11. Cell Viability Assay
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhao, Y.; Qu, H.; Hu, J.; Fu, J.; Chen, R.; Li, C.; Cao, S.; Wen, Y.; Wu, R.; Zhao, Q.; et al. Characterization and Pathogenicity of the Porcine Deltacoronavirus Isolated in Southwest China. Viruses 2019, 11, 1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leyi, W.; Beverly, B.; Yan, Z. Detection and genetic characterization of deltacoronavirus in pigs, Ohio, USA, 2014. Emerg. Infect. Dis. 2014, 20, 1227–1230. [Google Scholar] [CrossRef]
- Jung, K.; Hu, H.; Saif, L.J. Porcine deltacoronavirus infection: Etiology, cell culture for virus isolation and propagation, molecular epidemiology and pathogenesis. Virus Res. 2016, 226, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Vlasova, A.N.; Kenney, S.P.; Saif, L.J. Emerging and re-emerging coronaviruses in pigs. Curr. Opin. Virol. 2019, 34, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Fang, P.; Fang, L.; Hong, Y.; Liu, X.; Dong, N.; Ma, P.; Bi, J.; Wang, D.; Xiao, S. Discovery of a novel accessory protein NS7a encoded by porcine deltacoronavirus. J. Gen. Virol. 2017, 98, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Hulswit, R.J.G.; Kenney, S.P.; Widjaja, I.; Bosch, B.-J. Broad receptor engagement of an emerging global coronavirus may potentiate its diverse cross-species transmissibility. Proc. Natl. Acad. Sci. USA 2018, 115, E5135–E5143. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Chen, J.; Shi, D.; Shi, H.; Zhang, X.; Liu, J.; Cao, L.; Zhu, X.; Liu, Y.; Wang, X. Porcine deltacoronavirus enters cells via two pathways: A protease-mediated one at the cell surface and another facilitated by cathepsins in the endosome. J. Biol. Chem. 2019, 294, 9830–9843. [Google Scholar] [CrossRef]
- Qin, P.; Luo, W.-T.; Su, Q.; Zhao, P.; Zhang, Y.; Wang, B.; Yang, Y.-L.; Huang, Y.-W. The porcine deltacoronavirus accessory protein NS6 is expressed in vivo and incorporated into virions. Virology 2021, 556, 1–8. [Google Scholar] [CrossRef]
- Gu, W.-Y.; Li, Y.; Liu, B.-J.; Wang, J.; Yuan, G.-F.; Chen, S.-J.; Zuo, Y.-Z.; Fan, J.-H. Short hairpin RNAs targeting M and N genes reduce replication of porcine deltacoronavirus in ST cells. Virus Genes 2019, 55, 795–801. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Chen, J.; Tian, L.; Zhou, Y.; Xu, S.; Long, S.; Wang, D.; Fang, L.; Xiao, S. Porcine Deltacoronavirus nsp5 Cleaves DCP1A To Decrease Its Antiviral Activity. J. Virol. 2020, 94, e02162-19. [Google Scholar] [CrossRef]
- Jiang, S.; Li, F.; Li, X.; Wang, L.; Zhang, L.; Lu, C.; Zheng, L.; Yan, M. Transcriptome analysis of PK-15 cells in innate immune response to porcine deltacoronavirus infection. PLoS ONE 2019, 14, e0223177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.; Zhang, L.; Zhou, P.; Zhang, Y.; Wei, Y.; Wang, Y.; Liu, X. Tandem Mass Tag-Based Quantitative Proteome Analysis of Porcine Deltacoronavirus (PDCoV)-Infected LLC Porcine Kidney Cells. ACS Omega 2020, 5, 21979–21987. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.-H.; Zhang, Y.-F.; Yuan, Y.-X.; Han, L.; Zhang, G.-P.; Hu, H. Isolation, characterization and transcriptome analysis of porcine deltacoronavirus strain HNZK-02 from Henan Province, China. Mol. Immunol. 2021, 134, 86–99. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Pulido, D.; Boley, P.A.; Ouma, W.Z.; Alhamo, M.A.; Saif, L.J.; Kenney, S.P. Comparative Transcriptome Profiling of Human and Pig Intestinal Epithelial Cells after Porcine Deltacoronavirus Infection. Viruses 2021, 13, 292. [Google Scholar] [CrossRef]
- Zhou, X.; Zhou, L.; Ge, X.; Guo, X.; Han, J.; Zhang, Y.; Yang, H. Quantitative Proteomic Analysis of Porcine Intestinal Epithelial Cells Infected with Porcine Deltacoronavirus Using iTRAQ-Coupled LC-MS/MS. J. Proteome Res. 2020, 19, 4470–4485. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Fang, P.; Ke, W.; Wang, J.; Wang, X.; Xiao, S.; Fang, L. Porcine deltacoronavirus (PDCoV) infection antagonizes interferon-λ1 production. Vet. Microbiol. 2020, 247, 108785. [Google Scholar] [CrossRef]
- Liu, X.; Fang, P.; Fang, L.; Hong, Y.; Zhu, X.; Wang, D.; Peng, G.; Xiao, S. Porcine deltacoronavirus nsp15 antagonizes interferon-β production independently of its endoribonuclease activity. Mol. Immunol. 2019, 114, 100–107. [Google Scholar] [CrossRef]
- Likai, J.; Shasha, L.; Wenxian, Z.; Jingjiao, M.; Jianhe, S.; Hengan, W.; Yaxian, Y. Porcine Deltacoronavirus Nucleocapsid Protein Suppressed IFN-β Production by Interfering Porcine RIG-I dsRNA-Binding and K63-Linked Polyubiquitination. Front. Immunol. 2019, 10, 1024. [Google Scholar] [CrossRef] [Green Version]
- Fang, P.; Fang, L.; Ren, J.; Hong, Y.; Liu, X.; Zhao, Y.; Wang, D.; Peng, G.; Xiao, S. Porcine Deltacoronavirus Accessory Protein NS6 Antagonizes Interferon Beta Production by Interfering with the Binding of RIG-I/MDA5 to Double-Stranded RNA. J. Virol. 2018, 92, e00712-18. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Wang, D.; Zhou, J.; Pan, T.; Chen, J.; Yang, Y.; Lv, M.; Ye, X.; Peng, G.; Fang, L.; et al. Porcine Deltacoronavirus nsp5 Antagonizes Type I Interferon Signaling by Cleaving STAT2. J. Virol. 2017, 91, e00003-17. [Google Scholar] [CrossRef] [Green Version]
- Fang, P.; Hong, Y.; Xia, S.; Zhang, J.; Ren, J.; Zhou, Y.; Fang, L.; Xiao, S. Porcine deltacoronavirus nsp10 antagonizes interferon-β production independently of its zinc finger domains. Virology 2021, 559, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Fang, P.; Fang, L.; Xia, S.; Ren, J.; Zhang, J.; Bai, D.; Zhou, Y.; Peng, G.; Zhao, S.; Xiao, S. Porcine Deltacoronavirus Accessory Protein NS7a Antagonizes IFN-β Production by Competing With TRAF3 and IRF3 for Binding to IKKε. Front. Cell. Infect. Microbiol. 2020, 10, 257. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Yu, H.; Gu, W.; Luo, X.; Li, R.; Zhang, J.; Xu, Y.; Yang, L.; Shen, N.; Feng, L.; et al. Autophagy Negatively Regulates Transmissible Gastroenteritis Virus Replication. Sci. Rep. 2016, 6, 23864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richetta, C.; Faure, M. Autophagy in antiviral innate immunity. Cell Microbiol. 2013, 15, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Li, B.; Liu, M.; Zhou, H.; He, K.; Fan, H. Nonstructural protein 6 of porcine epidemic diarrhea virus induces autophagy to promote viral replication via the PI3K/Akt/mTOR axis. Vet. Microbiol. 2020, 244, 108684. [Google Scholar] [CrossRef] [PubMed]
- Hui, X.; Zhang, L.; Cao, L.; Huang, K.; Zhao, Y.; Zhang, Y.; Chen, X.; Lin, X.; Chen, M.; Jin, M. SARS-CoV-2 promote autophagy to suppress type I interferon response. Signal Transduct. Target. Ther. 2021, 6, 180. [Google Scholar] [CrossRef]
- Qin, P.; Du, E.-Z.; Luo, W.-T.; Yang, Y.-L.; Zhang, Y.-Q.; Wang, B.; Huang, Y.-W. Characteristics of the Life Cycle of Porcine Deltacoronavirus (PDCoV) In Vitro: Replication Kinetics, Cellular Ultrastructure and Virion Morphology, and Evidence of Inducing Autophagy. Viruses 2019, 11, 455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, C.; Liu, Y.; Hao, Z.; Wang, J. Ergosterol peroxide suppresses porcine deltacoronavirus (PDCoV)-induced autophagy to inhibit virus replication via p38 signaling pathway. Vet. Microbiol. 2021, 257, 109068. [Google Scholar] [CrossRef]
- Hoter, A.; El-Sabban, M.E.; Naim, H.Y. The HSP90 Family: Structure, Regulation, Function, and Implications in Health and Disease. Int. J. Mol. Sci. 2018, 19, 2560. [Google Scholar] [CrossRef] [Green Version]
- Genest, O.; Wickner, S.; Doyle, S.M. Hsp90 and Hsp70 chaperones: Collaborators in protein remodeling. J. Biol. Chem. 2019, 294, 2109–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, P.; Gaur, P.; Kumari, R.; Lal, S.K. Influenza A virus neuraminidase protein interacts with Hsp90, to stabilize itself and enhance cell survival. J. Cell. Biochem. 2019, 120, 6449–6458. [Google Scholar] [CrossRef]
- Sakata, M.; Katoh, H.; Otsuki, N.; Okamoto, K.; Mori, Y. Heat Shock Protein 90 Ensures the Integrity of Rubella Virus p150 Protein and Supports Viral Replication. J. Virol. 2019, 93, e01142-19. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Chu, H.; Liu, X.; Chiu, M.C.; Zhao, X.; Wang, D.; Wei, Y.; Hou, Y.; Shuai, H.; Cai, J.; et al. Human coronavirus dependency on host heat shock protein 90 reveals an antiviral target. Emerg. Microbes Infect. 2020, 9, 2663–2672. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, R.; Li, F.; Wang, Y.; Zhang, Z.; Wang, Q.; Ren, Z.; Jin, F.; Kitazato, K.; Wang, Y. Heat-shock protein 90α is involved in maintaining the stability of VP16 and VP16-mediated transactivation of α genes from herpes simplex virus-1. Mol. Med. 2018, 24, 65. [Google Scholar] [CrossRef] [Green Version]
- Roby, J.A.; Esser-Nobis, K.; Dewey-Verstelle, E.C.; Fairgrieve, M.R.; Schwerk, J.; Lu, A.Y.; Soveg, F.W.; Hemann, E.A.; Hatfield, L.D.; Keller, B.C.; et al. Flavivirus Nonstructural Protein NS5 Dysregulates HSP90 to Broadly Inhibit JAK/STAT Signaling. Cells 2020, 9, 899. [Google Scholar] [CrossRef]
- Sato, S.; Li, K.; Sakurai, N.; Hashizume, M.; Baidya, S.; Nonaka, H.; Noguchi, K.; Ishikawa, K.; Obuse, C.; Takaoka, A. Regulation of an adaptor protein STING by Hsp90β to enhance innate immune responses against microbial infections. Cell. Immunol. 2020, 356, 104188. [Google Scholar] [CrossRef]
- Liu, D.; Wu, A.; Cui, L.; Hao, R.; Wang, Y.; He, J.; Guo, D. Hepatitis B virus polymerase suppresses NF-κB signaling by inhibiting the activity of IKKs via interaction with Hsp90β. PLoS ONE 2014, 9, e91658. [Google Scholar] [CrossRef] [Green Version]
- Qu, H.; Wen, Y.; Hu, J.; Xiao, D.; Li, S.; Zhang, L.; Liao, Y.; Chen, R.; Zhao, Y.; Wen, Y.; et al. Study of the inhibitory effect of STAT1 on PDCoV infection. Vet. Microbiol. 2022, 266, 109333. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Xiao, D.; Zhang, L.; Song, D.; Chen, R.; Li, S.; Liao, Y.; Wen, Y.; Liu, W.; Yu, E.; et al. HSP90 inhibitors 17-AAG and VER-82576 inhibit porcine deltacoronavirus replication in vitro. Vet. Microbiol. 2022, 265, 109316. [Google Scholar] [CrossRef]
- Jung, K.; Hu, H.; Saif, L.J. Calves are susceptible to infection with the newly emerged porcine deltacoronavirus, but not with the swine enteric alphacoronavirus, porcine epidemic diarrhea virus. Arch. Virol. 2017, 162, 2357–2362. [Google Scholar] [CrossRef] [PubMed]
- Boley, P.A.; Alhamo, M.A.; Lossie, G.; Yadav, K.K.; Vasquez-Lee, M.; Saif, L.J.; Kenney, S.P. Porcine Deltacoronavirus Infection and Transmission in Poultry, United States(1). Emerg. Infect. Dis. 2020, 26, 255–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Q.; Zhang, H.; Li, B.; Ding, Q.; Wang, Y.; Gao, W.; Guo, D.; Wei, Z.; Hu, H. Susceptibility of Chickens to Porcine Deltacoronavirus Infection. Viruses 2019, 11, 573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Jia, K.; Jia, P.; Xiang, Y.; Lu, X.; Liu, W.; Yi, M. Marine medaka heat shock protein 90ab1 is a receptor for red-spotted grouper nervous necrosis virus and promotes virus internalization through clathrin-mediated endocytosis. PLoS Pathog. 2020, 16, e1008668. [Google Scholar] [CrossRef] [PubMed]
- Wyler, E.; Mösbauer, K.; Franke, V.; Diag, A.; Gottula, L.T.; Arsiè, R.; Klironomos, F.; Koppstein, D.; Hönzke, K.; Ayoub, S.; et al. Transcriptomic profiling of SARS-CoV-2 infected human cell lines identifies HSP90 as target for COVID-19 therapy. IScience 2021, 24, 102151. [Google Scholar] [CrossRef]
- Zhang, W.J.; Wang, R.Q.; Li, L.T.; Fu, W.; Chen, H.C.; Liu, Z.F. Hsp90 is involved in pseudorabies virus virion assembly via stabilizing major capsid protein VP5. Virology 2021, 553, 70–80. [Google Scholar] [CrossRef]
- Zhang, J.; Huang, Y.; Li, L.; Dong, J.; Liao, M.; Sun, M. Transcriptome Analysis Reveals the Neuro-Immune Interactions in Duck Tembusu Virus-Infected Brain. Int. J. Mol. Sci. 2020, 21, 2402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Mao, L.; Shu, X.; Liu, R.; Hao, F.; Li, J.; Liu, M.; Yang, L.; Zhang, W.; Sun, M.; et al. Transcriptome analysis reveals differential immune related genes expression in bovine viral diarrhea virus-2 infected goat peripheral blood mononuclear cells (PBMCs). BMC Genom. 2019, 20, 516. [Google Scholar] [CrossRef]
- Cheng, Y.; Sun, F.; Wang, L.; Gao, M.; Xie, Y.; Sun, Y.; Liu, H.; Yuan, Y.; Yi, W.; Huang, Z.; et al. Virus-induced p38 MAPK activation facilitates viral infection. Theranostics 2020, 10, 12223–12240. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, A.; Sun, Y.; Zhang, S.; Xia, T.; Guo, T.; Hao, Z.; Sun, L.; Jiang, Y.; Qiao, X.; et al. Porcine transmissible gastroenteritis virus inhibits NF-κB activity via nonstructural protein 3 to evade host immune system. Virol. J. 2019, 16, 97. [Google Scholar] [CrossRef] [Green Version]
- Regan, A.D.; Cohen, R.D.; Whittaker, G.R. Activation of p38 MAPK by feline infectious peritonitis virus regulates pro-inflammatory cytokine production in primary blood-derived feline mononuclear cells. Virology 2009, 384, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, M.; Guo, S.; Guo, Y.F.; Zahoor, A.; Shaukat, A.; Chen, Y.; Umar, T.; Deng, P.G.; Guo, M. Upregulated-gene expression of pro-inflammatory cytokines (TNF-α, IL-1β and IL-6) via TLRs following NF-κB and MAPKs in bovine mastitis. Acta. Trop. 2020, 207, 105458. [Google Scholar] [CrossRef] [PubMed]
- Duan, C.; Ge, X.; Wang, J.; Wei, Z.; Feng, W.H.; Wang, J. Ergosterol peroxide exhibits antiviral and immunomodulatory abilities against porcine deltacoronavirus (PDCoV) via suppression of NF-κB and p38/MAPK signaling pathways in vitro. Int. Immunopharmacol. 2021, 93, 107317. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Song, X.; Su, G.; Wang, Y.; Wang, Z.; Qing, S.; Jia, J.; Wang, Y.; Huang, L.; Zheng, K.; et al. AT-533, a Hsp90 inhibitor, attenuates HSV-1-induced inflammation. Biochem. Pharmacol. 2019, 166, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Cai, D.; Wang, M.; He, T.; Deng, L.; Wu, L.; Jia, R.; Zhu, D.; Liu, M.; Chen, S.; et al. SC75741 antagonizes vesicular stomatitis virus, duck Tembusu virus, and duck plague virus infection in duck cells through promoting innate immune responses. Poult. Sci. 2021, 100, 101085. [Google Scholar] [CrossRef] [PubMed]
- Haasbach, E.; Reiling, S.J.; Ehrhardt, C.; Droebner, K.; Rückle, A.; Hrincius, E.R.; Leban, J.; Strobl, S.; Vitt, D.; Ludwig, S.; et al. The NF-kappaB inhibitor SC75741 protects mice against highly pathogenic avian influenza A virus. Antivir. Res. 2013, 99, 336–344. [Google Scholar] [CrossRef]
- Mendoza, C.A.; Yamaoka, S.; Tsuda, Y.; Matsuno, K.; Weisend, C.M.; Ebihara, H. The NF-κB inhibitor, SC75741, is a novel antiviral against emerging tick-borne bandaviruses. Antivir. Res. 2021, 185, 104993. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Zhen, Z.D.; Fan, D.Y.; Wang, P.G.; An, J. Transcriptomic Analysis Suggests the M1 Polarization and Launch of Diverse Programmed Cell Death Pathways in Japanese Encephalitis Virus-Infected Macrophages. Viruses 2020, 12, 356. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Zhong, H.; Huang, S.; Zhou, Q.; Du, Y.; Chen, L.; Xue, C.; Cao, Y. Porcine deltacoronavirus induces TLR3, IL-12, IFN-α, IFN-β and PKR mRNA expression in infected Peyer’s patches in vivo. Vet. Microbiol. 2019, 228, 226–233. [Google Scholar] [CrossRef]
- Jung, K.; Miyazaki, A.; Hui, H.; Saif, L.J. Susceptibility of porcine IPEC-J2 intestinal epithelial cells to infection with porcine deltacoronavirus (PDCoV) and serum cytokine responses of gnotobiotic pigs to acute infection with IPEC-J2 cell culture-passaged PDCoV. Vet. Microbiol. 2018, 221, 49–58. [Google Scholar] [CrossRef]
- Li, H.-Y.; Zhang, H.-L.; Zhao, F.-J.; Wang, S.-Q.; Wang, Z.-X.; Wei, Z.-Y. Modulation of Gut Microbiota, Short-Chain Fatty Acid Production, and Inflammatory Cytokine Expression in the Cecum of Porcine Deltacoronavirus-Infected Chicks. Fron. Microbiol. 2020, 11, 897. [Google Scholar] [CrossRef]
- Zhang, H.; Han, F.; Shu, X.; Li, Q.; Ding, Q.; Hao, C.; Yan, X.; Xu, M.; Hu, H. Co-infection of porcine epidemic diarrhoea virus and porcine deltacoronavirus enhances the disease severity in piglets. Transbound. Emerg. Dis. 2021, 14, 144. [Google Scholar] [CrossRef] [PubMed]
- Saeng-Chuto, K.; Madapong, A.; Kaeoket, K.; Piñeyro, P.E.; Tantituvanont, A.; Nilubol, D. Coinfection of porcine deltacoronavirus and porcine epidemic diarrhea virus increases disease severity, cell trophism and earlier upregulation of IFN-α and IL12. Sci. Rep. 2021, 11, 3040. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.X.; Nie, F.R.; Huang, A.Q.; Wang, R.N.; Li, M.Y.; Deng, H.Y.; Zhou, Y.Z.; Zhou, X.M.; Huang, Y.K.; Zhou, J.; et al. Transcriptomic analysis of chicken immune response to infection of different doses of Newcastle disease vaccine. Gene 2021, 766, 145077. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Österlund, P.; Fagerlund, R.; Rios, D.N.; Hoffmann, A.; Poranen, M.M.; Bamford, D.H.; Julkunen, I. MAP kinase p38α regulates type III interferon (IFN-λ1) gene expression in human monocyte-derived dendritic cells in response to RNA stimulation. J. Leukoc. Biol. 2015, 97, 307–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, J.; Chen, R.; Hu, J.; Qu, H.; Zhao, Y.; Cao, S.; Wen, X.; Wen, Y.; Wu, R.; Zhao, Q.; et al. Identification of a Novel Linear B-Cell Epitope on the Nucleocapsid Protein of Porcine Deltacoronavirus. Int. J. Mol. Sci. 2020, 21, 648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, L.J.; Muench, H. A Simple Method Of Estimating Fifty Per Cent Endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Reads | Clean Reads | Raw Bases (G) | Clean Bases (G) | Q20 (%) | Q30 (%) | GC Content (%) |
|---|---|---|---|---|---|---|---|
| KO-PDCoV-1 | 89,417,288 | 89,203,130 | 13.41 | 13.28 | 97.68 | 93.27 | 48.14 |
| KO-PDCoV-2 | 73,552,754 | 73,402,918 | 11.03 | 10.93 | 97.73 | 93.40 | 48.58 |
| KO-mock-1 | 69,435,150 | 69,244,218 | 10.42 | 10.32 | 97.57 | 93.06 | 48.68 |
| KO-mock-2 | 60,095,972 | 59,938,292 | 9.01 | 8.93 | 97.61 | 93.14 | 48.79 |
| WT-PDCoV-1 | 68,873,684 | 68,723,094 | 10.33 | 10.24 | 97.55 | 92.88 | 48.10 |
| WT-PDCoV-2 | 72,214,636 | 72,048,968 | 10.83 | 10.74 | 97.60 | 93.06 | 48.03 |
| WT-mock-1 | 84,349,756 | 84,131,302 | 12.65 | 12.53 | 97.52 | 92.90 | 48.49 |
| WT-mock-2 | 67,038,012 | 66,866,762 | 10.06 | 9.95 | 97.60 | 93.05 | 48.88 |
| Sample | Total Reads 1 | Total Mapped Reads (%) | Unique Mapped Reads (%) | Exon (%) | Intron (%) | Intergenic (%) | Expressed Genes | Novel Genes |
|---|---|---|---|---|---|---|---|---|
| KO-PDCoV-1 | 89,079,788 | 85.68 | 83.21 | 76.03 | 20.96 | 3.01 | 15,299 | 581 |
| KO-PDCoV-2 | 73,291,444 | 85.19 | 82.75 | 77.71 | 19.47 | 2.82 | 15,073 | 588 |
| KO-mock-1 | 69,123,276 | 94.99 | 92.47 | 78.22 | 18.27 | 3.51 | 15,260 | 592 |
| KO-mock-2 | 59,835,550 | 95.11 | 92.63 | 78.09 | 18.43 | 3.48 | 15,246 | 587 |
| WT-PDCoV-1 | 68,625,598 | 86.24 | 83.86 | 79.22 | 17.75 | 3.04 | 15,634 | 589 |
| WT-PDCoV-2 | 71,947,570 | 87.61 | 85.25 | 79.09 | 17.82 | 3.09 | 15,745 | 587 |
| WT-mock-1 | 84,004,888 | 95.48 | 92.86 | 79.09 | 17.70 | 3.21 | 15,786 | 586 |
| WT-mock-2 | 66,758,268 | 95.30 | 92.64 | 79.72 | 17.16 | 3.12 | 15,485 | 579 |
| Genes | Sequences (5′–3′) | Size (bp) |
|---|---|---|
| PDCoV M | F: CCAATGGGTACATGGAGGT | 130 |
| R: GTGGCGGATTTCTAACTGA | ||
| HSP90AB1 | F: TGGAGAGTTCTACAAAAGCCTG | 304 |
| R: CTTCAAAATCTTGCTCTGCTG | ||
| HSP70 | F: GCACGAGGAAAGCCTTAGAG | 166 |
| R: GGAGAAGATGGGACGACAAA | ||
| CCL4 | F: CTCTCCTCCAGCAAGACCAT | 296 |
| R: CAGTTCAGTTCCAAGTCATCCA | ||
| CXCL10 | F: AATCTACCTCTGCCATCATCTC | 373 |
| R: AGTAGAAGCCCACGGAGTAAAG | ||
| CXCL11 | F: AACTATTCAAGGCTTCCCCAT | 201 |
| R: ACATTTGCTTGCTTTGATTTG | ||
| GHSR | F: TCCCACCGATGACGAGAG | 276 |
| R: CTTGATGATTCCCAGGAGTTT | ||
| CCL2 | F: GAAGAGTCACCAGCAGCAAG | 144 |
| R: AAGGCTTCGGAGTTTGGTT | ||
| CXCL9 | F: GCTTTTGGGTATCATCTTCCT | 266 |
| R: TCTTTAGGCTGACCTGTTTCT | ||
| IFNB1 | F: ACCAACAAAGGAGCAGCAA | 266 |
| R: ATCCATCTGCCCATCAAGTT | ||
| IL27 | F: CCAGATGGCAGACGACCT | 185 |
| R: GGACCCGAACCTCAGACAG | ||
| IL28B | F: GTGCTGATGACGGTGGCT | 164 |
| R: AGGGACTCTTCAAAGGCATC | ||
| IL29 | F: CTTTCAAGCCCACCACAAC | 241 |
| R: GCCCTTAGGACCTTCAGAGT | ||
| IL6 | F: GATGCTTCCAATCTGGGTTC | 218 |
| R: ATTTGTGGTGGGGTTAGGG | ||
| TNF | F: TCCTCACTCACACCATCAGC | 222 |
| R: GCCCAGATTCAGCAAAGTCC | ||
| TNFSF9 | F: GGCTTCCCAGACTATCCAC | 124 |
| R: CACCTTTCATCCCTGGCTC | ||
| TNFSF13B | F: CCGATAACACCTTTGCCAT | 124 |
| R: TTATTGGGTAGTGTTTCAGGC | ||
| NFKB1 | F: CGGTGAAGGTCTGAACGC | 188 |
| R: GGGCATCACCCTCCAAGA | ||
| NFKBIA | F: CACCAACCAGCCAGAAATC | 261 |
| R: GCACCCAAAGACACCAACA | ||
| NFKBIZ | F: CAAGTAGAGCAGGAAGAAAGCA | 195 |
| R: CTGAAAGGCACTCTGTCCATTA | ||
| NFKBIB | F: CTCTGACCATACCCCTGACA | 176 |
| R: GGACCATCTCGGCATCTT | ||
| NKAP | F: AGAGTCCCAGGAAGAGTTGC | 153 |
| R: CCTTCACCAGGTAACAGAGC | ||
| Porcine ACTB | F: CTTCCTGGGCATGGAGTCC | 201 |
| R: GGCGCGATGATCTTGATCTTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Chen, R.; Xiao, D.; Zhang, L.; Song, D.; Wen, Y.; Wu, R.; Zhao, Q.; Du, S.; Wen, X.; et al. A Comparative Transcriptomic Analysis Reveals That HSP90AB1 Is Involved in the Immune and Inflammatory Responses to Porcine Deltacoronavirus Infection. Int. J. Mol. Sci. 2022, 23, 3280. https://doi.org/10.3390/ijms23063280
Zhao Y, Chen R, Xiao D, Zhang L, Song D, Wen Y, Wu R, Zhao Q, Du S, Wen X, et al. A Comparative Transcriptomic Analysis Reveals That HSP90AB1 Is Involved in the Immune and Inflammatory Responses to Porcine Deltacoronavirus Infection. International Journal of Molecular Sciences. 2022; 23(6):3280. https://doi.org/10.3390/ijms23063280
Chicago/Turabian StyleZhao, Yujia, Rui Chen, Dai Xiao, Luwen Zhang, Daili Song, Yiping Wen, Rui Wu, Qin Zhao, Senyan Du, Xintian Wen, and et al. 2022. "A Comparative Transcriptomic Analysis Reveals That HSP90AB1 Is Involved in the Immune and Inflammatory Responses to Porcine Deltacoronavirus Infection" International Journal of Molecular Sciences 23, no. 6: 3280. https://doi.org/10.3390/ijms23063280
APA StyleZhao, Y., Chen, R., Xiao, D., Zhang, L., Song, D., Wen, Y., Wu, R., Zhao, Q., Du, S., Wen, X., Cao, S., & Huang, X. (2022). A Comparative Transcriptomic Analysis Reveals That HSP90AB1 Is Involved in the Immune and Inflammatory Responses to Porcine Deltacoronavirus Infection. International Journal of Molecular Sciences, 23(6), 3280. https://doi.org/10.3390/ijms23063280