
Ocular Surface Infection Mediated Molecular Stress Responses: A Review

 and
and
Abstract
1. Introduction
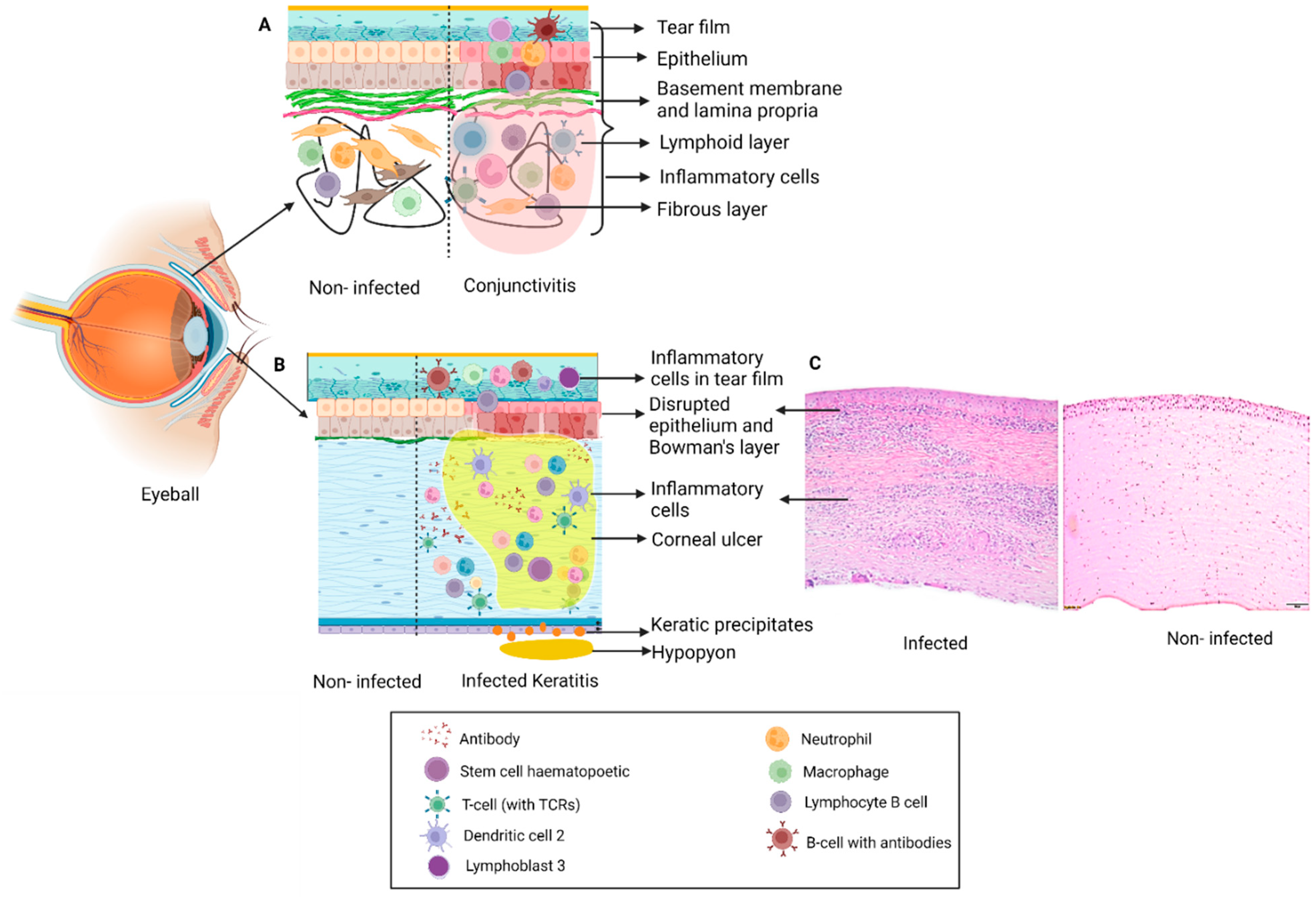
2. Clinical Presentation of Keratitis and Conjunctivitis and Treatment Modalities
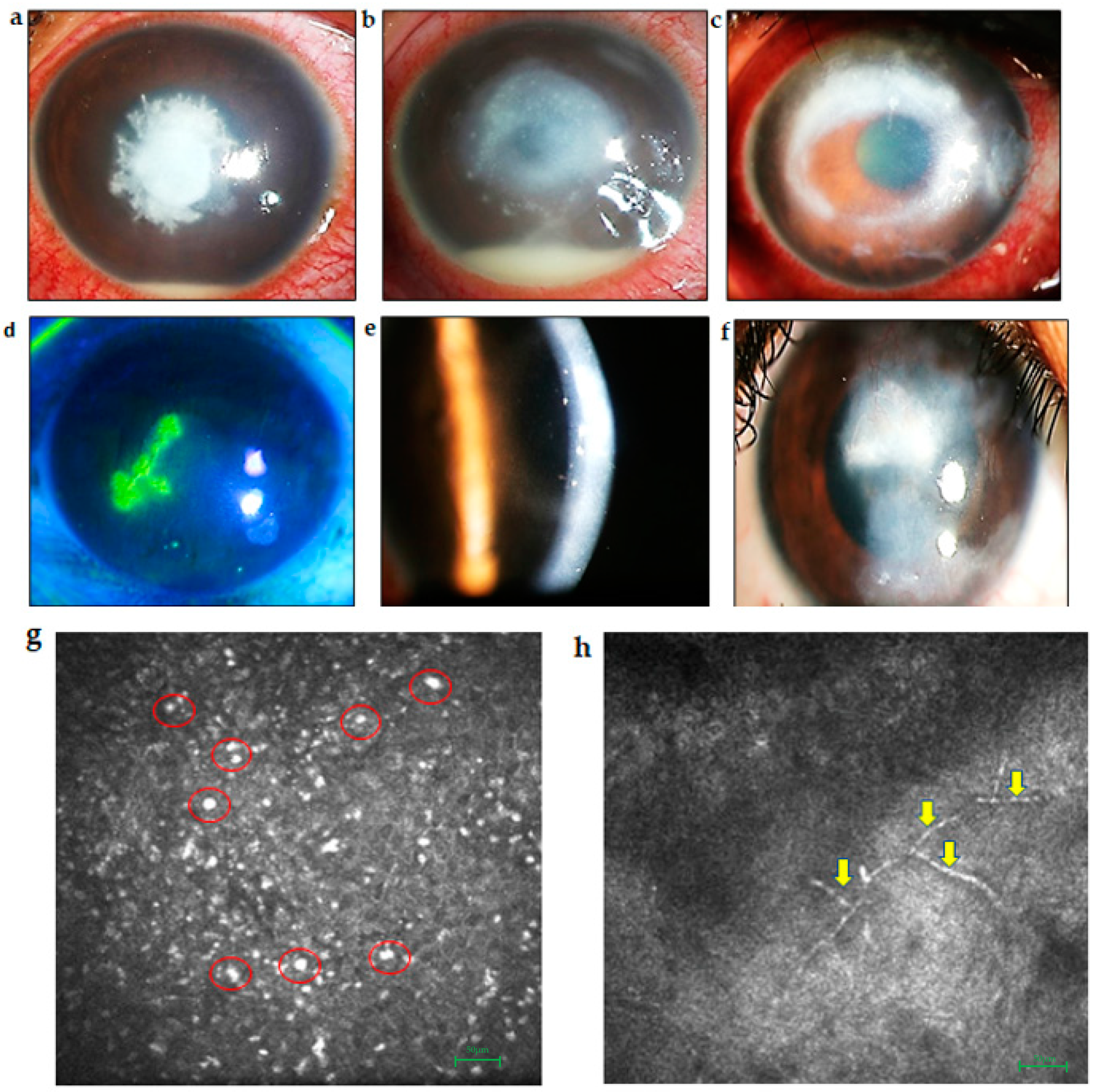
2.1. Keratitis
2.1.1. Bacterial Keratitis
2.1.2. Fungal Keratitis
2.1.3. Viral Keratitis
Herpes Simplex Virus Keratitis
Herpes Zoster Ophthalmicus
2.1.4. Acanthameoba Keratitis
2.2. Conjunctivitis
2.2.1. Bacterial Conjunctivitis
2.2.2. Viral Conjunctivitis
3. Diagnostic Challenges
4. Clinical Need
5. Molecular Status of Ocular Surface Infection Mediated Host Molecular Responses
5.1. Bacterial Keratitis
5.2. Fungal Keratitis
5.3. Viral Infections
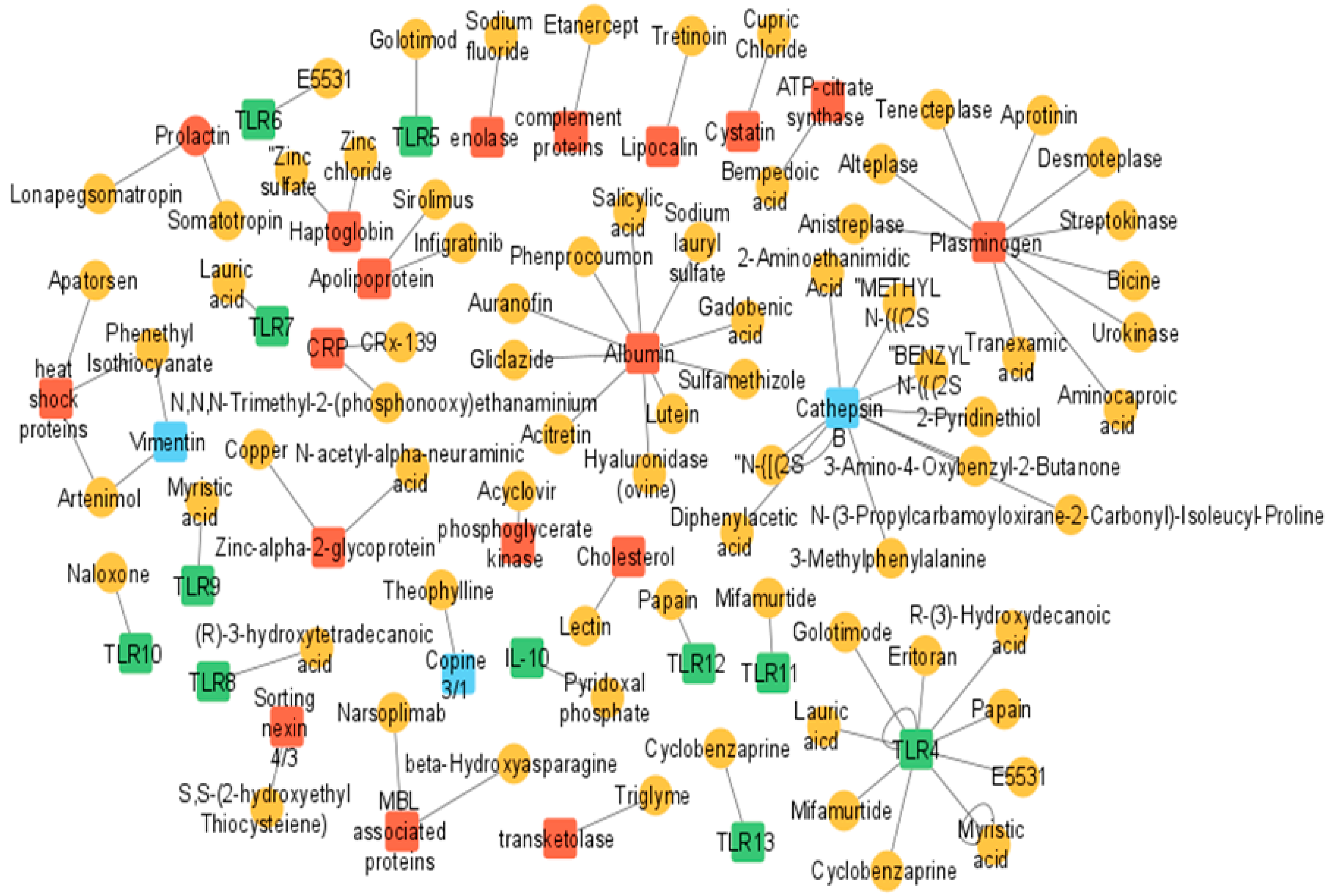
6. Translational and Clinical Relevance
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Cher, I. Ocular surface concepts: Development and citation. Ocul. Surf. 2013, 12, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, M. Infective keratitis: A challenge to Indian ophthalmologists. Indian J. Ophthalmol. 2007, 55, 5–6. [Google Scholar] [CrossRef] [PubMed]
- Marquart, M.E.; O’Callaghan, R.J. Infectious keratitis: Secreted bacterial proteins that mediate corneal damage. J. Ophthalmol. 2013, 2013, 369094. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, M.S. Anatomy of cornea and ocular surface. Indian J. Ophthalmol. 2018, 66, 190. [Google Scholar] [CrossRef] [PubMed]
- Garfunkel, L.C.; Kaczorowski, J.; Christy, C. Pediatric Clinical Advisor E-book: Instant Diagnosis and Treatment; Elsevier Health Sciences: Philadelphia, PA, USA, 2007. [Google Scholar]
- St. Leger, A.J.; Caspi, R.R. Visions of eye commensals: The known and the unknown about how the microbiome affects eye disease. Bioessays 2018, 40, 1800046. [Google Scholar] [CrossRef] [PubMed]
- Amescua, G.; Miller, D.; Alfonso, E. What is causing the corneal ulcer? Management strategies for unresponsive corneal ulceration. Eye 2012, 26, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.; Sachdev, A.; Coggon, D.; Hossain, P. Geographic variations in microbial keratitis: An analysis of the peer-reviewed literature. Br. J. Ophthalmol. 2011, 95, 762–767. [Google Scholar] [CrossRef] [PubMed]
- Garg, P.; Rao, G.N. Corneal ulcer: Diagnosis and management. Community Eye Health 1999, 12, 21. [Google Scholar] [PubMed]
- Azari, A.A.; Barney, N.P. Conjunctivitis: A systematic review of diagnosis and treatment. JAMA 2013, 310, 1721–1730. [Google Scholar] [CrossRef]
- Prajna, V.N.; Nirmalan, P.K.; Saravanan, S.; Srinivasan, M. Economic analysis of corneal ulcers in South India. Cornea 2007, 26, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Go, E.P.; Wikoff, W.R.; Shen, Z.; O’Maille, G.; Morita, H.; Conrads, T.P.; Nordstrom, A.; Trauger, S.A.; Uritboonthai, W.; Lucas, D.A.; et al. Mass spectrometry reveals specific and global molecular transformations during viral infection. J. Proteome Res. 2006, 5, 2405–2416. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kosch, R.; Delarocque, J.; Claus, P.; Stefanie, C.; Jung, B.K. Gene expression profiles in neurological tissues during West Nile virus infection: A critical meta-analysis. BMC Genom. 2018, 19, 530. [Google Scholar] [CrossRef] [PubMed]
- Mejias, A.; Dimo, B.; Suarez, N.M.; Garcia, C.; Suarez-Arrabal, M.C.; Jartti, T.; Blankenship, D.; Jordan-Villegas, A.; Ardura, M.I.; Xu, Z.; et al. Whole blood gene expression profiles to assess pathogenesis and disease severity in infants with respiratory syncytial virus infection. PLoS Med. 2013, 10, e1001549. [Google Scholar] [CrossRef] [PubMed]
- Rahman, Z.A.; Harun, A.; Hasan, H.; Mohamed, Z.; Noor, S.S.; Deris, Z.Z.; Ismail, N.; Hassan, A.S.; Ahmad, F.; Yaakub, A. Ocular surface infections in northeastern state of malaysia: A 10-year review of bacterial isolates and antimicrobial susceptibility. Eye Contact Lens 2013, 39, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, H.E.; Barron, B.A.; McDonald, M.B. The Cornea, on CD-ROM; Butterworth-Heinemann: Oxford, UK, 1998. [Google Scholar]
- Kanski, J.J.; Bowling, B. Clinical Ophthalmology. A Systematic Approach; Neuroophtholmology; Butterworth-Heinemann: Oxford, UK, 2007. [Google Scholar]
- O’brien, T. Management of bacterial keratitis: Beyond exorcism towards consideration of organism and host factors. Eye 2003, 17, 957–974. [Google Scholar] [CrossRef] [PubMed]
- Song, A.; Deshmukh, R.; Lin, H.; Ang, M.; Mehta, J.S.; Chodosh, J.; Said, D.G.; Dua, H.S.; Ting, D. Post-keratoplasty infectious keratitis: Epidemiology, risk factors, management, and outcomes. Front. Med. 2021, 8, 707242. [Google Scholar] [CrossRef] [PubMed]
- Dikmetaş, Ö.; Deniz, Y.; Kocabeyoğlu, S.; Başol, M.; İrkeç, M. The Value of Fortified Aminoglycoside/Cephalosporin Treatment as First-Line Treatment and in Fluoroquinolone-Resistant Bacterial Keratitis. Turk. J. Ophthalmol. 2020, 50, 258. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.; Liu, X.; Ma, Z.; Yin, Y.; Sun, L.; Yang, L.; Zheng, Y. Fungal keratitis: Pathogenesis, diagnosis and prevention. Microb. Pathog. 2020, 138, 103802. [Google Scholar] [CrossRef] [PubMed]
- O’day, D.M.; Head, W.S.; Robinson, R.D.; Clanton, J.A. Corneal penetration of topical amphotericin B and natamycin. Curr. Eye Res. 1986, 5, 877–882. [Google Scholar] [CrossRef] [PubMed]
- Abbouda, A.; Abicca, I.; Alio, J. Infectious keratitis following corneal crosslinking: A systematic review of reported cases: Management, visual outcome, and treatment proposed. In Seminars in Ophthalmology; Taylor & Francis: Abingdon, UK, 2016. [Google Scholar]
- Valerio, G.S.; Lin, C.C. Ocular manifestations of herpes simplex virus. Curr. Opin. Ophthalmol. 2019, 30, 525–531. [Google Scholar] [CrossRef]
- Farooq, A.V.; Shah, A.; Shukla, D. The role of herpesviruses in ocular infections. Virus Adapt. Treat. 2010, 2, 115–123. [Google Scholar]
- Kwon, M.S.; Carnt, N.A.; Truong, N.R.; Pattamatta, U.; White, A.J.; Samarawickrama, C.; Cunningham, A.L. Dendritic cells in the cornea during Herpes simplex viral infection and inflammation. Surv. Ophthalmol. 2018, 63, 565–578. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, B.; Patel, B.C. Herpes simplex keratitis. Prog. Retin. Eye Res. 2019, 25, 355–380. [Google Scholar]
- Liesegang, T.J. Herpes simplex virus epidemiology and ocular importance. Cornea 2001, 20, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Rowe, A.; St Leger, A.J.; Jeon, S.; Dhaliwal, D.K.; Knickelbein, J.E.; Hendricks, R.L. Herpes keratitis. Prog. Retin. Eye Res. 2013, 32, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Szentmáry, N.; Daas, L.; Shi, L.; Laurik, K.L.; Lepper, S.; Milioti, G.; Seitz, B. Acanthamoeba keratitis–Clinical signs, differential diagnosis and treatment. J. Curr. Ophthalmol. 2019, 31, 16–23. [Google Scholar] [CrossRef]
- Lorenzo-Morales, J.; Khan, N.A.; Walochnik, J. An update on Acanthamoeba keratitis: Diagnosis, pathogenesis and treatment. Parasite 2015, 22, 10. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, T.D. Treatment of Acanthamoeba keratitis. Cornea 1998, 17, 11–16. [Google Scholar] [CrossRef] [PubMed]
- American Optometric Association. Care of the Patient with Hyperopia; American Optometric Association: St. Louis, MO, USA, 2010. [Google Scholar]
- Rietveld, R.P.; Riet, G.; Bindels, P.J.; Sloos, J.H.; van Weert, H.C. Predicting bacterial cause in infectious conjunctivitis: Cohort study on informativeness of combinations of signs and symptoms. BMJ 2004, 329, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Satpathy, G.; Behera, H.S.; Ahmed, N.H. Chlamydial eye infections: Current perspectives. Indian J. Ophthalmol. 2017, 65, 97. [Google Scholar] [CrossRef] [PubMed]
- Deschênes, J.; Seamone, C.; Baines, M. The ocular manifestations of sexually transmitted diseases. Canadian journal of ophthalmology. J. Can. D’ophtalmol. 1990, 25, 177–185. [Google Scholar]
- Sheikh, A.; Hurwitz, B.; van Schayck, C.P.; McLean, S.; Nurmatov, U. Antibiotics versus placebo for acute bacterial conjunctivitis. Cochrane Database Syst. Rev. 2012, 9. [Google Scholar] [CrossRef] [PubMed]
- Hu, V.H.; Holland, M.J.; Burton, M.J. Trachoma: Protective and pathogenic ocular immune responses to Chlamydia trachomatis. PLoS Negl. Trop. Dis. 2013, 7, e2020. [Google Scholar] [CrossRef] [PubMed]
- Sow, A.; Kane, H.; Ka, A.M.; Hanne, F.T.; Ndiaye, J.M.M.; Diagne, J.P.; Nguer, M.; Sow, S.; Saheli, Y.; Sy, E.H.M.; et al. Senegalese experience with acute viral conjunctivitis. J. Fr. D’ophtalmol. 2017, 40, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Haq, A.; Wardak, H.; Kraskian, N. Infective conjunctivitis–its pathogenesis, management and complications. In Common Eye Infections; Intechopen: Vienna, Austria, 2013; p. 21. [Google Scholar]
- Hierholzer, J.C.; Wigand, R.; Anderson, L.J.; Adrian, T.; Gold, J.W. Adenoviruses from patients with AIDS: A plethora of serotypes and a description of five new serotypes of subgenus D (types 43–47). J. Infect. Dis. 1988, 158, 804–813. [Google Scholar] [CrossRef] [PubMed]
- Solano, D.; Fu, L.; Czyz, C.N. Viral Conjunctivitis; StatPearls Publishing: Treasure Island, FL, USA, 2017. [Google Scholar]
- Gordon, J. Adenovirus and other non-herpetic viral diseases. In The Cornea, 3rd ed.; Little, Brown & Company: Boston, MA, USA, 1994; pp. 215–222. [Google Scholar]
- McLeod, S.D. The role of cultures in the management of ulcerative keratitis. Cornea 1997, 16, 381–382. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S. Diagnosis of infectious diseases of the eye. Eye 2012, 26, 177–184. [Google Scholar] [CrossRef] [PubMed]
- McLeod, S.D.; Kolahdouz-Isfahani, A.; Rostamian, K.; Flowers, C.W.; Lee, P.P.; McDonnell, P.J. The role of smears, cultures, and antibiotic sensitivity testing in the management of suspected infectious keratitis. Ophthalmology 1996, 103, 23–28. [Google Scholar] [CrossRef]
- Kim, E.; Chidambaram, J.D.; Srinivasan, M.; Lalitha, P.; Wee, D.; Lietman, T.M.; Whitcher, J.P.; Van Gelder, R.N. Prospective comparison of microbial culture and polymerase chain reaction in the diagnosis of corneal ulcer. Am. J. Ophthalmol. 2008, 146, 714–723.e1. [Google Scholar] [CrossRef] [PubMed]
- Yoder, J.S.; Verani, J.; Heidman, N.; Hoppe-Bauer, J.; Alfonso, E.C.; Miller, D.; Jones, D.B.; Bruckner, D.; Langston, R.; Jeng, B.H.; et al. Acanthamoeba keratitis: The persistence of cases following a multistate outbreak. Ophthalmic Epidemiol. 2012, 19, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Miserocchi, E.; Fogliato, G.; Bianchi, I.; Bandello, F.; Modorati, G. Clinical features of ocular herpetic infection in an Italian referral center. Cornea 2014, 33, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Baratz, K.H. The role of antiviral therapy after the resolution of acute herpes simplex keratitis or acute herpes zoster ophthalmicus. Arch. Ophthalmol. 2012, 130, 108–110. [Google Scholar] [CrossRef] [PubMed]
- Liesegang, T.J. Herpes zoster virus infection. Curr. Opin. Ophthalmol. 2004, 15, 531–536. [Google Scholar] [CrossRef] [PubMed]
- Edell, A.R.; Cohen, E.J. Herpes simplex and herpes zoster eye disease: Presentation and management at a city hospital for the underserved in the United States. Eye Contact Lens 2013, 39, 311–314. [Google Scholar] [CrossRef] [PubMed]
- El-Aal, A.M.A.; El Sayed, M.; Mohammed, E.; Ahmed, M.; Fathy, M. Evaluation of herpes simplex detection in corneal scrapings by three molecular methods. Curr. Microbiol. 2006, 52, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Satpathy, G.; Mishra, A.K.; Tandon, R.; Sharma, M.K.; Sharma, A.; Nayak, N.; Titiyal, J.S.; Sharma, N. Evaluation of tear samples for Herpes Simplex Virus 1 (HSV) detection in suspected cases of viral keratitis using PCR assay and conventional laboratory diagnostic tools. Br. J. Ophthalmol. 2011, 95, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, R.P.; Gordon, Y.J.; Romanowski, E.G.; Araullo-Cruz, T.; Kinchington, P.R. A comparison of enzyme immunoassay and polymerase chain reaction with the clinical examination for diagnosing ocular herpetic disease. Ophthalmology 1993, 100, 530–533. [Google Scholar] [CrossRef]
- McGilligan, V.; Moore, J.E.; Tallouzi, M.; Atkinson, S.D.; Neill, H.O.; Feeney, S.; Novitskaya, E.S.; Sharma, A.; Shah, S.; Jackson, J.A. A comparison of the clinical and molecular diagnosis of herpes simplex keratitis. Open J. Ophthalmol. 2014, 4, 65–74. [Google Scholar] [CrossRef]
- Bartee, E.; McCormack, A.; Früh, K. Quantitative membrane proteomics reveals new cellular targets of viral immune modulators. PLoS Pathog. 2006, 2, e107. [Google Scholar] [CrossRef] [PubMed]
- Callegan, M.C.; Engel, L.S.; Hill, J.M.; O’Callaghan, R.J. Corneal virulence of Staphylococcus aureus: Roles of alpha-toxin and protein A in pathogenesis. Infect. Immun. 1994, 62, 2478–2482. [Google Scholar] [CrossRef]
- Strateva, T.; Mitov, I. Contribution of an arsenal of virulence factors to pathogenesis of Pseudomonas aeruginosa infections. Ann. Microbiol. 2011, 61, 717–732. [Google Scholar] [CrossRef]
- Sewell, A.; Dunmire, J.; Wehmann, M.; Rowe, T.; Bouhenni, R. Proteomic analysis of keratitis-associated Pseudomonas aeruginosa. Mol. Vis. 2014, 20, 1182. [Google Scholar] [PubMed]
- Hilliam, Y.; Kaye, S.; Winstanley, C. Pseudomonas aeruginosa and microbial keratitis. J. Med. Microbiol. 2020, 69, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Sack, R.; Sathe, S.; Beaton, A.R.; McNamara, N.; Fleiszig, S.; Ni, M. Protein array characterization of bioactive proteins secreted by immortalized human corneal epithelium in response to pseudomonas constituents. Curr. Eye Res. 2009, 34, 92–98. [Google Scholar] [CrossRef]
- Bouhenni, R.; Dunmire, J.; Rowe, T.; Bates, J. Proteomics in the study of bacterial keratitis. Proteomes 2015, 3, 496–511. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Kong, Q.; Dong, Z.; Dong, W.; Fu, X.; Su, L.; Tan, X. NLRC3 promotes host resistance against Pseudomonas aeruginosa-induced keratitis by promoting the degradation of IRAK1. Int. J. Mol. Med. 2017, 40, 898–906. [Google Scholar] [CrossRef] [PubMed]
- Hazlett, L.D.; McClellan, S.; Somayajulu, M.; Bessert, D. Targeting Inflammation Driven by HMGB1 in Bacterial Keratitis—A Review. Pathogens 2021, 10, 1235. [Google Scholar] [CrossRef] [PubMed]
- Taube, M.; Cendra, D.M.; Elsahn, A. Pattern recognition receptors in microbial keratitis. Eye 2015, 29, 1399–1415. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Karmakar, M.; Roy, S.; Ramadan, R.T.; Williams, S.R.; Howell, S.; Shive, C.L.; Han, Y.; Stopford, C.M.; Rietsch, A.; et al. TLR4 and TLR5 on corneal macrophages regulate Pseudomonas aeruginosa keratitis by signaling through MyD88-dependent and-independent pathways. J. Immunol. 2010, 185, 4272–4283. [Google Scholar] [CrossRef]
- Ross, B.X.; Gao, N.; Cui, X.; Standiford, T.J.; Xu, J.; Yu, F.X. IL-24 promotes Pseudomonas aeruginosa keratitis in C57BL/6 mouse corneas. J. Immunol. 2017, 198, 3536–3547. [Google Scholar] [CrossRef]
- Willcox, M.D. Pseudomonas aeruginosa infection and inflammation during contact lens wear: A review. Optom. Vis. Sci. 2007, 84, 273–278. [Google Scholar] [CrossRef]
- Sueke, H.; Kaye, S.B.; Neal, T.; Hall, A.; Tuft, S.; Parry, C.M. An in vitro investigation of synergy or antagonism between antimicrobial combinations against isolates from bacterial keratitis. Investig. Ophthalmol. Vis. Sci. 2010, 51, 4151–4155. [Google Scholar] [CrossRef] [PubMed]
- Boyd, K.; Pagan-Duran, B.; Pink Eye (Conjunctivitis). American Academy of Ophthalmology. EyeSmart® Eye Health. Available online: https://www.aao.org/eye-health/diseases/pink-eye-conjunctivitis-list (accessed on 22 March 2019).
- Tian, R.; Zou, H.; Wang, L.; Liu, L.; Song, M.; Zhang, H. Analysis of differentially expressed genes in bacterial and fungal keratitis. Indian J. Ophthalmol. 2020, 68, 39–46. [Google Scholar] [PubMed]
- Carnt, N.A.; Cipriani, V.; Stapleton, F.J.; Calder, V.; Willcox, M.D. Association study of single nucleotide polymorphisms in IL-10 and IL-17 genes with the severity of microbial keratitis. Contact Lens Anterior Eye 2019, 42, 658–661. [Google Scholar] [CrossRef] [PubMed]
- Chidambaram, J.D.; Kannambath, S.; Srikanthi, P.; Shah, M.; Lalitha, P.; Elakkiya, S.; Bauer, J.; Prajna, N.V.; Holland, M.J.; Burton, M.J. Persistence of innate immune pathways in late stage human bacterial and fungal keratitis: Results from a comparative transcriptome analysis. Front. Cell. Infect. Microbiol. 2017, 7, 193. [Google Scholar] [CrossRef] [PubMed]
- Gowda, R.N.; Redfern, R.; Frikeche, J.; Pinglay, S.; Foster, J.W. Functions of peptidoglycan recognition proteins (Pglyrps) at the ocular surface: Bacterial keratitis in gene-targeted mice deficient in Pglyrp-2,-3 and-4. PLoS ONE 2015, 10, e0137129. [Google Scholar]
- Kuo, M.-T.; Chen, J.L.; Hsu, S.L.; Chen, A.; You, H.L. An omics approach to diagnosing or investigating fungal keratitis. Int. J. Mol. Sci. 2019, 20, 3631. [Google Scholar] [CrossRef]
- Ananthi, S.; Chitra, T.; Bini, R.; Prajna, N.V.; Lalitha, P.; Dharmalingam, K. Comparative analysis of the tear protein profile in mycotic keratitis patients. Mol. Vis. 2008, 14, 500–507. [Google Scholar] [PubMed]
- Ananthi, S.; Venkatesh, P.N.; Lalitha, P.; Valarnila, M.; Dharmalingam, K. Pathogen induced changes in the protein profile of human tears from Fusarium keratitis patients. PLoS ONE 2013, 8, e53018. [Google Scholar] [CrossRef]
- Kandhavelu, J.; Demonte, N.L.; Namperumalsamy, V.P.; Prajna, L.; Thangavel, C.; Jayapal, J.M.; Kuppamuthu, D. Aspergillus flavus induced alterations in tear protein profile reveal pathogen-induced host response to fungal infection. J. Proteom. 2017, 152, 13–21. [Google Scholar] [CrossRef]
- Calvillo-Medina, R.P.; Reyes-Grajeda, J.P.; Barba-Escoto, L.; Bautista-Hernandez, L.A.; Campos-Guillén, J.; Jones, G.H.; Bautista-de Lucio, V.M. Proteome analysis of biofilm produced by a Fusarium falciforme keratitis infectious agent. Microb. Pathog. 2019, 130, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Maharana, P.K.; Sharma, N.; Nagpal, R.; Jhanji, V.; Das, S.; Vajpayee, R.B. Recent advances in diagnosis and management of Mycotic Keratitis. Indian J. Ophthalmol. 2016, 64, 346. [Google Scholar] [PubMed]
- Parthiban, N.; Sampath, N.L.; Jeya Maheshwari, J.; Prajna, N.V.; Lalitha, P.; Dharmalingam, K. Quantitative profiling of tear proteome reveals down regulation of zinc alpha-2 glycoprotein in Aspergillus flavus keratitis patients. Exp. Eye Res. 2019, 186, 107700. [Google Scholar] [CrossRef] [PubMed]
- Ranjith, K.; Kalyana, C.S.; Adicherla, H.; Sharma, S.; Shivaji, S. Temporal expression of genes in biofilm-forming ocular Candida albicans isolated from patients with keratitis and orbital cellulitis. Investig. Ophthalmol. Vis. Sci. 2018, 59, 528–538. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhang, J.; Gong, M.; Pan, R.; Liu, Y.; Tao, L.; He, K. Transcriptome analysis of the gene expression profiles associated with fungal keratitis in mice based on RNA-Seq. Investig. Ophthalmol. Vis. Sci. 2020, 61, 32. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liang, Q.; Liu, Y.; Pan, Z.; Baudouin, C.; Labbé, A.; Lu, Q. Expression of cytokines in aqueous humor from fungal keratitis patients. BMC Ophthalmol. 2018, 18, 105. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Xu, Q.; Lin, J.; Chen, W.; Cui, T.; Hu, L.; Jiang, N. The role of Mincle in innate immune to fungal keratitis. J. Infect. Dev. Ctries. 2017, 11, 89–97. [Google Scholar] [CrossRef]
- Cui, H.; Liu, Y.; Huang, Y. Roles of TRIM32 in corneal epithelial cells after infection with herpes simplex virus. Cell. Physiol. Biochem. 2017, 43, 801–811. [Google Scholar] [CrossRef]
- Berard, A.R.; Coombs, K.M.; Severini, A. Quantification of the host response proteome after herpes simplex virus type 1 infection. J. Proteome Res. 2015, 14, 2121–2142. [Google Scholar] [CrossRef]
- Wang, L.; Wang, R.; Xu, C.; Zhou, H. Pathogenesis of herpes stromal keratitis: Immune inflammatory response mediated by inflammatory regulators. Front. Immunol. 2020, 11, 766. [Google Scholar] [CrossRef]
- Maertzdorf, J.; Osterhaus, A.D.; Verjans, G.M. IL-17 expression in human herpetic stromal keratitis: Modulatory effects on chemokine production by corneal fibroblasts. J. Immunol. 2002, 169, 5897–5903. [Google Scholar] [CrossRef]
- Nicoll, M.P.; Proença, J.T.; Efstathiou, S. The molecular basis of herpes simplex virus latency. FEMS Microbiol. Rev. 2012, 36, 684–705. [Google Scholar] [CrossRef] [PubMed]
- Tormanen, K.; Allen, S.; Mott, K.R.; Ghiasi, H. The latency-associated transcript inhibits apoptosis via downregulation of components of the type I interferon pathway during latent herpes simplex virus 1 ocular infection. J. Virol. 2019, 93, e00103-19. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Schambach, F.; DeJong, C.S.; Hammond, S.M.; Reiner, S.L. Micro-RNA-155 inhibits IFN-γ signaling in CD4+ T cells. Eur. J. Immunol. 2010, 40, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Huffaker, T.B.; Hu, R.; Runtsch, M.C.; Bake, E.; Chen, X.; Zhao, J.; Round, J.L.; Baltimore, D.; O’Connell, R.M. Epistasis between microRNAs 155 and 146a during T cell-mediated antitumor immunity. Cell Rep. 2012, 2, 1697–1709. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.; Majeti, B.K.; Acevedo, L.M.; Murphy, E.A.; Mukthavaram, R.; Scheppke, L.; Huang, M.; Shields, D.J.; Lindquist, J.N.; Lapinski, P.E.; et al. MicroRNA-132–mediated loss of p120RasGAP activates the endothelium to facilitate pathological angiogenesis. Nat. Med. 2010, 16, 909–914. [Google Scholar] [CrossRef] [PubMed]
- Jaishankar, D.; Yakoub, A.M.; Yadavalli, T.; Agelidis, A.; Thakkar, N.; Hadigal, S.; Ames, J.; Shukla, D. An off-target effect of BX795 blocks herpes simplex virus type 1 infection of the eye. Sci. Transl. Med. 2018, 10, eaan5861. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Pan, Z.-Q.; Zhai, C.-B. Adenovirus-mediated RNA interference against herpes simplex virus infection in vitro. Folia Histochem. Cytobiol. 2021, 59, 302–310. [Google Scholar] [CrossRef]
- Yang, H.; Yang, X.; Wang, Y.; Zheng, X.; Zhang, Y.; Shao, Y. Comparative analysis of the tear protein profile in herpes simplex virus type 1 epithelial keratitis. BMC Ophthalmol. 2020, 20, 355. [Google Scholar] [CrossRef] [PubMed]
- Tumpey, T.M.; Cheng, H.; Cook, D.N.; Smithies, O.; Oakes, J.E.; Lausch, R.N. Absence of macrophage inflammatory protein-1α prevents the development of blinding herpes stromal keratitis. J. Virol. 1998, 72, 3705–3710. [Google Scholar] [CrossRef]
- Twardy, B.S.; Channappanavar, R.; Suvas, S. Substance P in the corneal stroma regulates the severity of herpetic stromal keratitis lesions. Investig. Ophthalmol. Vis. Sci. 2011, 52, 8604–8613. [Google Scholar] [CrossRef] [PubMed]
- Hurt, M.; Apte, S.; Leher, H.; Howard, K.; Niederkorn, J.; Alizadeh, H. Exacerbation of Acanthamoeba keratitis in animals treated with anti-macrophage inflammatory protein 2 or antineutrophil antibodies. Infect. Immun. 2001, 69, 2988–2995. [Google Scholar] [CrossRef] [PubMed]
- Dreyfuss, J.L.; Regatieri, C.V.; Coelho, B.; Barbosa, J.B.; De Freitas, D.; Nader, H.B.; Martins, J.R. Altered hyaluronic acid content in tear fluid of patients with adenoviral conjunctivitis. An. Acad. Bras. Ciências 2015, 87, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Feunang, Y.D.; Guo, A.C.; Lo, E.J.; Marcu, A.; Grant, J.R.; Sajed, T.; Johnson, D.; Li, C.; Sayeeda, Z.; et al. DrugBank 5.0: A major update to the DrugBank database for 2018. Nucleic Acids Res. 2018, 46, D1074–D1082. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Ocular Infection | Causative Agent | Sign/Symptoms | Treatment |
|---|---|---|---|
| Bacterial Keratitis |
| Symptoms: Eye pain, blurred vision, photophobia, and discharge Signs: Corneal infiltrate of varying involvement of corneal stromal depth and diameter with overlying epithelial defect. Severe infections can be associated with anterior chamber inflammation and hypopyon formation. Progressive ulceration can result in complications including corneal perforation, scleritis, and endophthalmitis | Topical antibiotics are used based on microbiological sensitivity. Combination of medications may be required |
| Fungal Keratitis |
| Symptoms: Eye pain, blurred vision, photophobia, and discharge Signs: Dry looking base of corneal ulcer base with feathery margins. Can also have satellite lesions, endoexudates and hypopyon | Topical antifungal medications as per severity of infections. Natamycin and voriconazole are the most common medications used. Oral antifungals of the azole group are also given for more severe infections |
| Viral Keratitis |
| Symptoms: Discomfort, watering, itching, burning, and pain in the eye Signs: Would depend on the form of the herpes viral involvement | Topical acyclovir and ganciclovir for epithelial forms of disease. Stromal viral keratitis requires topical steroids. Viral endothelitis and recurrent stromal involvement require oral antiviral medications |
| Bacterial Conjunctivitis |
| Symptoms: Unilateral or bilateral redness, classically purulent discharge, photophobia, tearing, irritation, stinging, burning, and discomfort Signs: Diffuse bulbar conjunctival injection. Discharge is initially watery, becomes purulent as infection progresses | Acute bacterial conjunctivitis is usually treated with topical fluoroquinolones, macrolides, and aminoglycosides. Tetracycline and macrolides are used to treat chlamydia |
| Trachoma | Chlamydia trachomatis serotypes A, B, Ba, C |
| SAFE strategy: S: Surgery in case of trichiasis, A: Antibiotics (azithromycin and erythromycin) for C. trachomatis infection, F: Facial cleanliness, E: Environmental change for sanitation |
| Viral Conjunctivitis |
| Symptoms: Unilateral or bilateral watering, redness, discomfort, and photophobia Signs: Eyelid edema, follicular conjunctivitis, tender preauricular lymphadenopathy. Chronic cases can have mild conjunctival scarring | Symptomatic relief and cold compresses. Topical antibiotics to prevent secondary infection and topical steroids for severe inflammation in conjunctivitis |
| Ocular Disease | Techniques Used | Host Molecular Responses | References |
|---|---|---|---|
| Bacterial keratitis |
|
|
|
| Fungal keratitis |
|
|
|
| Viral keratitis |
|
|
|
| Viral conjunctivitis | ELISA | Elevated levels of hyaluronic acid (HA) can acts as a rapid diagnostic marker | Dreyfuss, J.L. et al Juliana (2015) |
| Ocular Infection | Techniques | Host Molecular Responses | References |
|---|---|---|---|
| Viral Keratitis |
|
|
|
| Ebola virus uveitis | RNA sequencing, RT-qPCR | Type I interferon (IFN) | Smith, J. R. et al. (2017) |
| Bacterial keratitis |
|
|
|
| Fungal keratitis |
|
|
|
| Acanthameoba keratitis | MIP-2 and myeloperoxidase (MPO) assays | MIP-2 induces neutrophil infiltration which acts as a therapeutic strategy in Acanthameoba keratitis | Hurt, M. et al. (2001) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Das, S.; D’Souza, S.; Gorimanipalli, B.; Shetty, R.; Ghosh, A.; Deshpande, V. Ocular Surface Infection Mediated Molecular Stress Responses: A Review. Int. J. Mol. Sci. 2022, 23, 3111. https://doi.org/10.3390/ijms23063111
Das S, D’Souza S, Gorimanipalli B, Shetty R, Ghosh A, Deshpande V. Ocular Surface Infection Mediated Molecular Stress Responses: A Review. International Journal of Molecular Sciences. 2022; 23(6):3111. https://doi.org/10.3390/ijms23063111
Chicago/Turabian StyleDas, Samayitree, Sharon D’Souza, Bhavya Gorimanipalli, Rohit Shetty, Arkasubhra Ghosh, and Vrushali Deshpande. 2022. "Ocular Surface Infection Mediated Molecular Stress Responses: A Review" International Journal of Molecular Sciences 23, no. 6: 3111. https://doi.org/10.3390/ijms23063111
APA StyleDas, S., D’Souza, S., Gorimanipalli, B., Shetty, R., Ghosh, A., & Deshpande, V. (2022). Ocular Surface Infection Mediated Molecular Stress Responses: A Review. International Journal of Molecular Sciences, 23(6), 3111. https://doi.org/10.3390/ijms23063111