
Effect of ALDH1A1 Gene Knockout on Drug Resistance in Paclitaxel and Topotecan Resistant Human Ovarian Cancer Cell Lines in 2D and 3D Model

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Results
2.1. Model of the Study
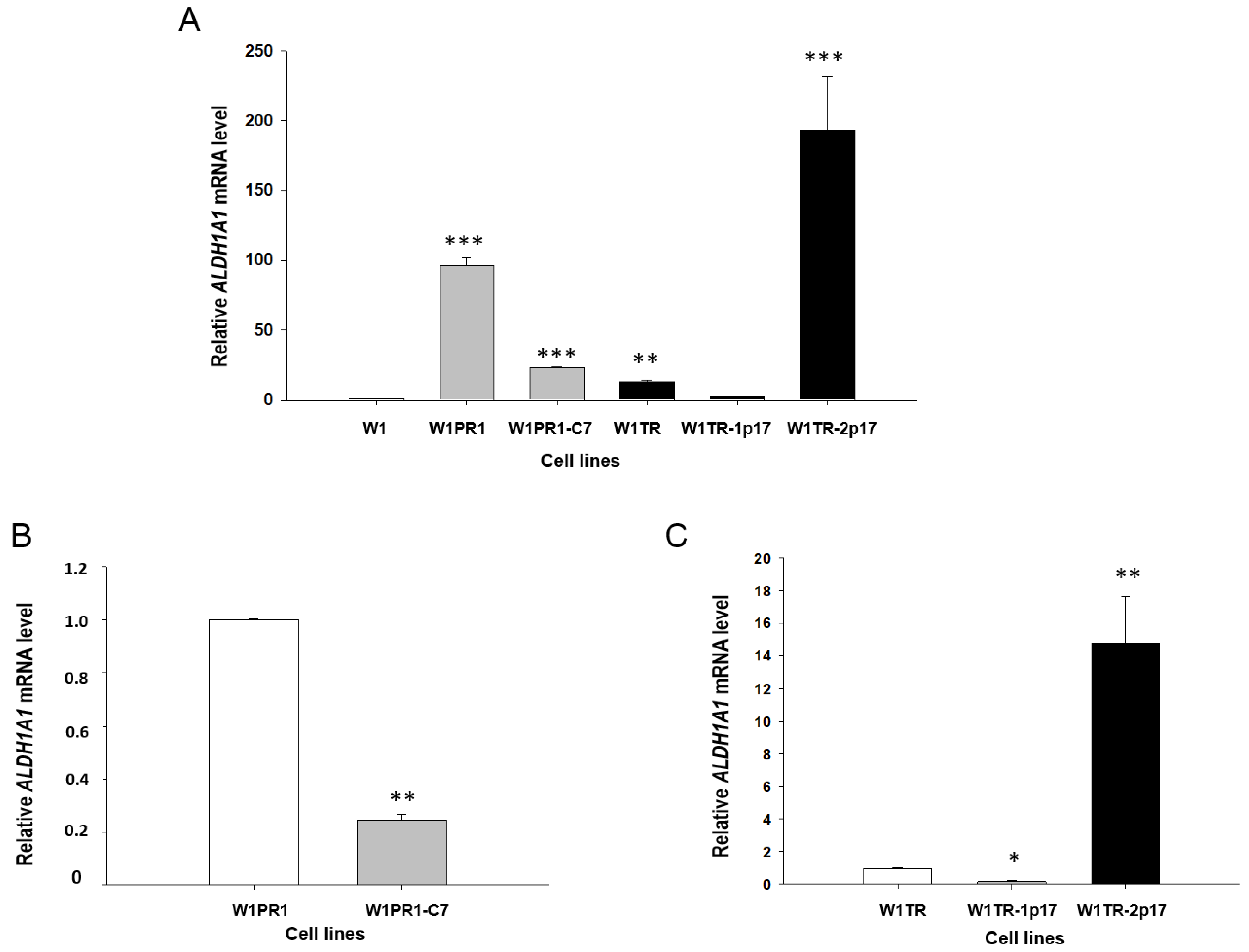
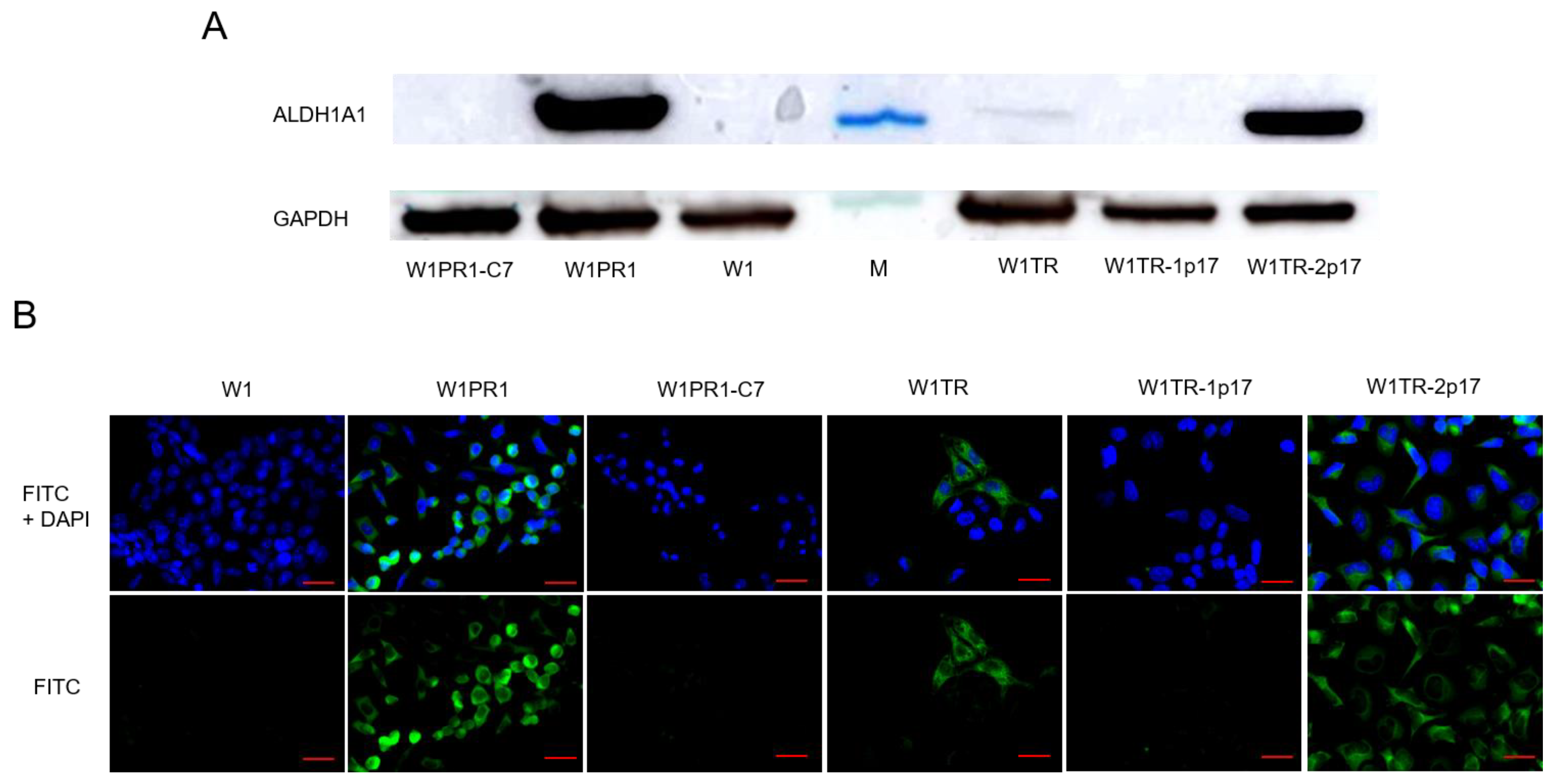
2.2. ALDH1A1 Gene Knockout Using the CRISPR-Cas9 System
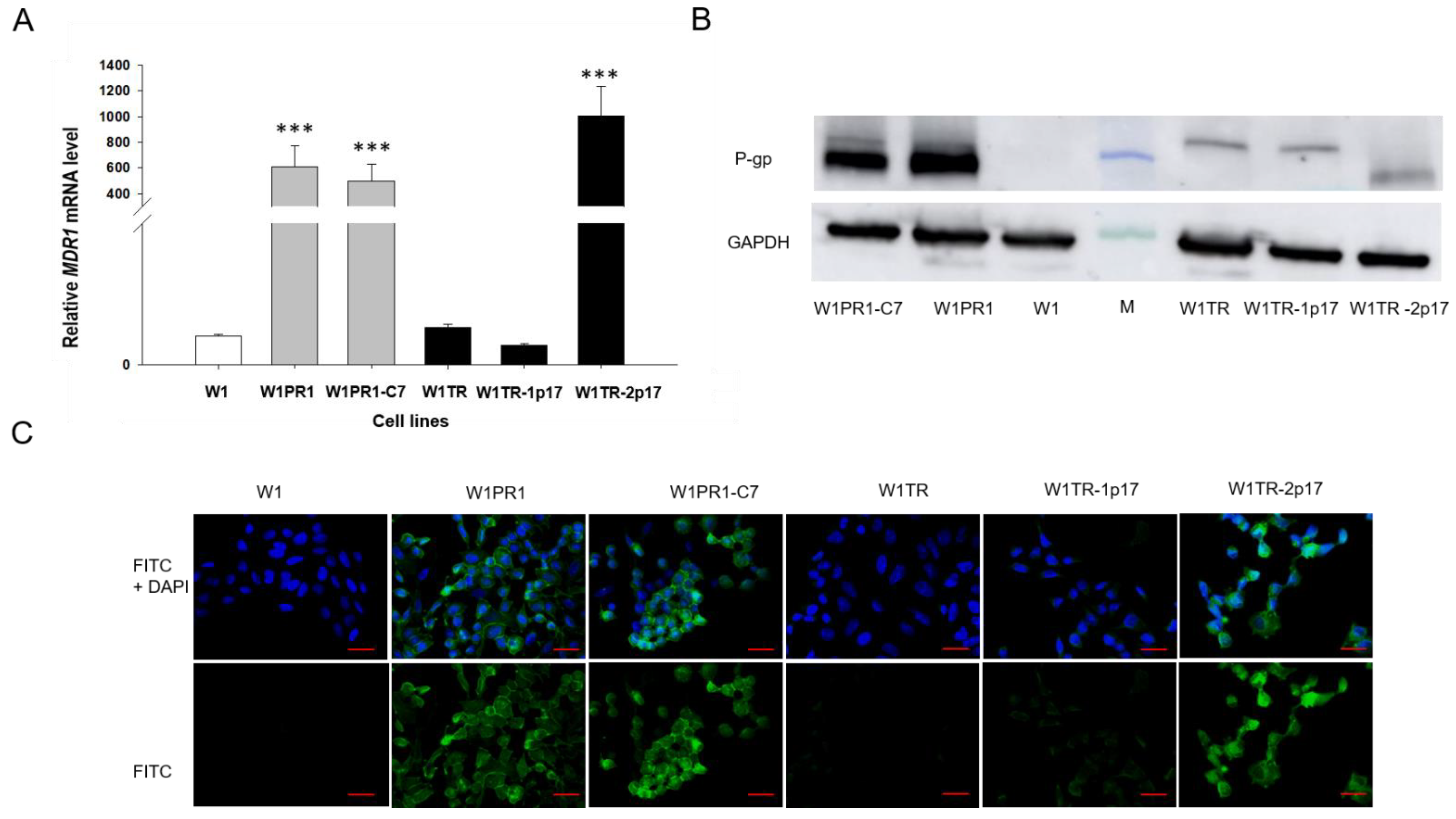
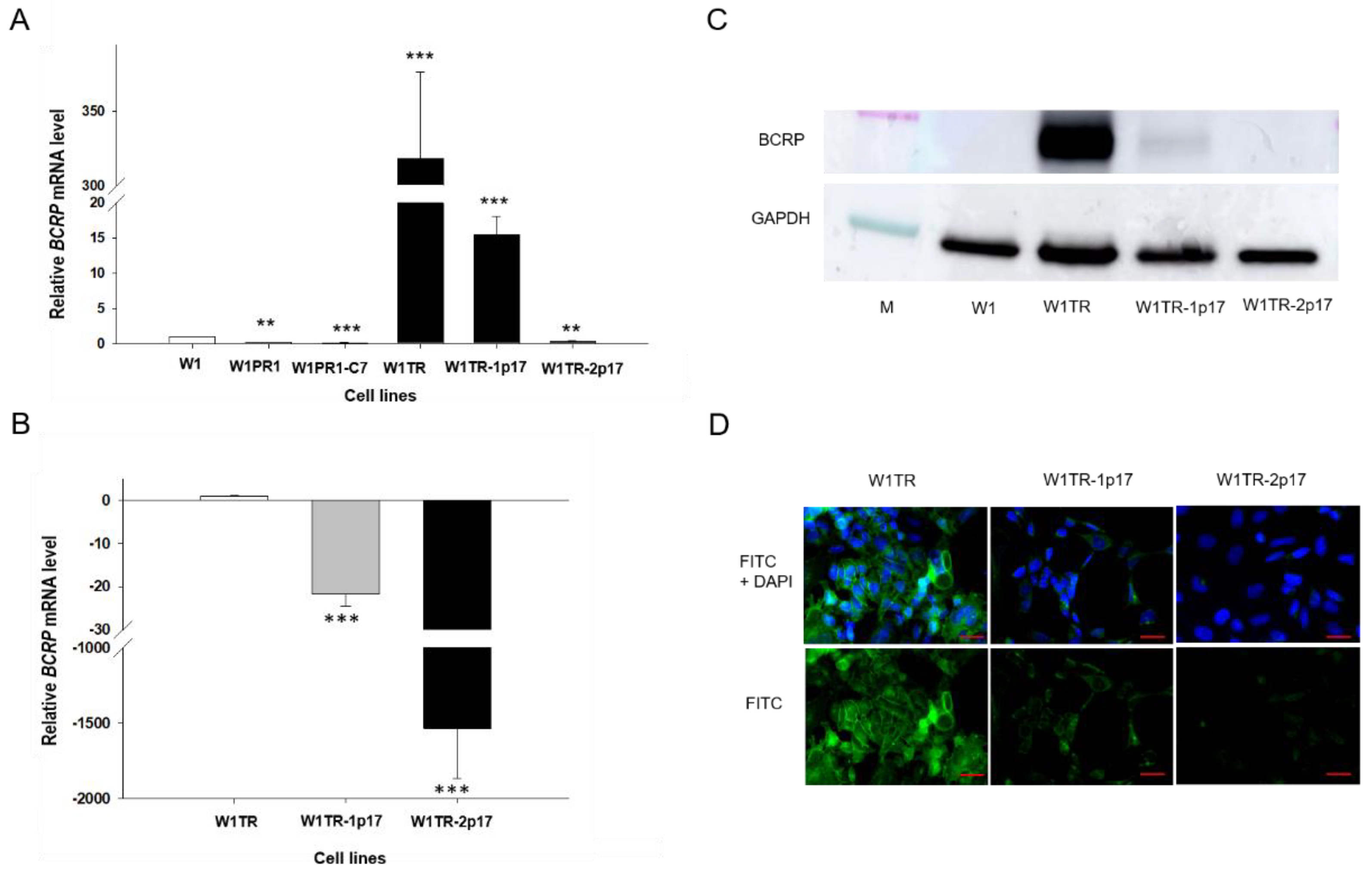
2.3. MDR1/Glycoprotein- P, BCRP, COL3A1 Gene and Protein Expression in Ovarian Cancer Cell Lines
2.3.1. MDR1 Gene and Protein Expression in Investigated Cell Lines
2.3.2. BCRP Gene and Protein Expression in Drug-Resistant Ovarian Cancer Cell Lines
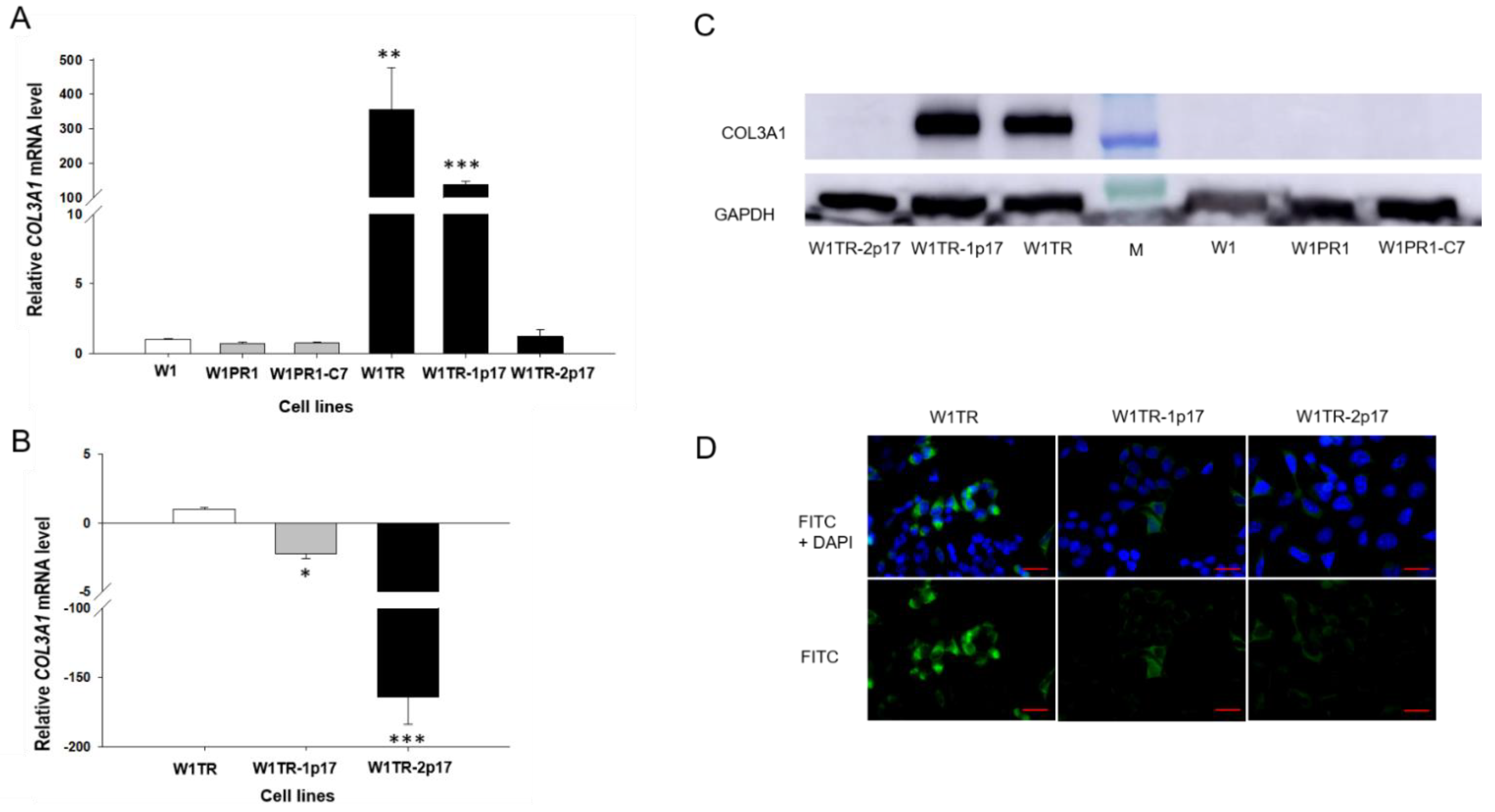
2.3.3. COL3A1 Gene and Protein Expression in Drug-Resistant Ovarian Cancer Cell Lines
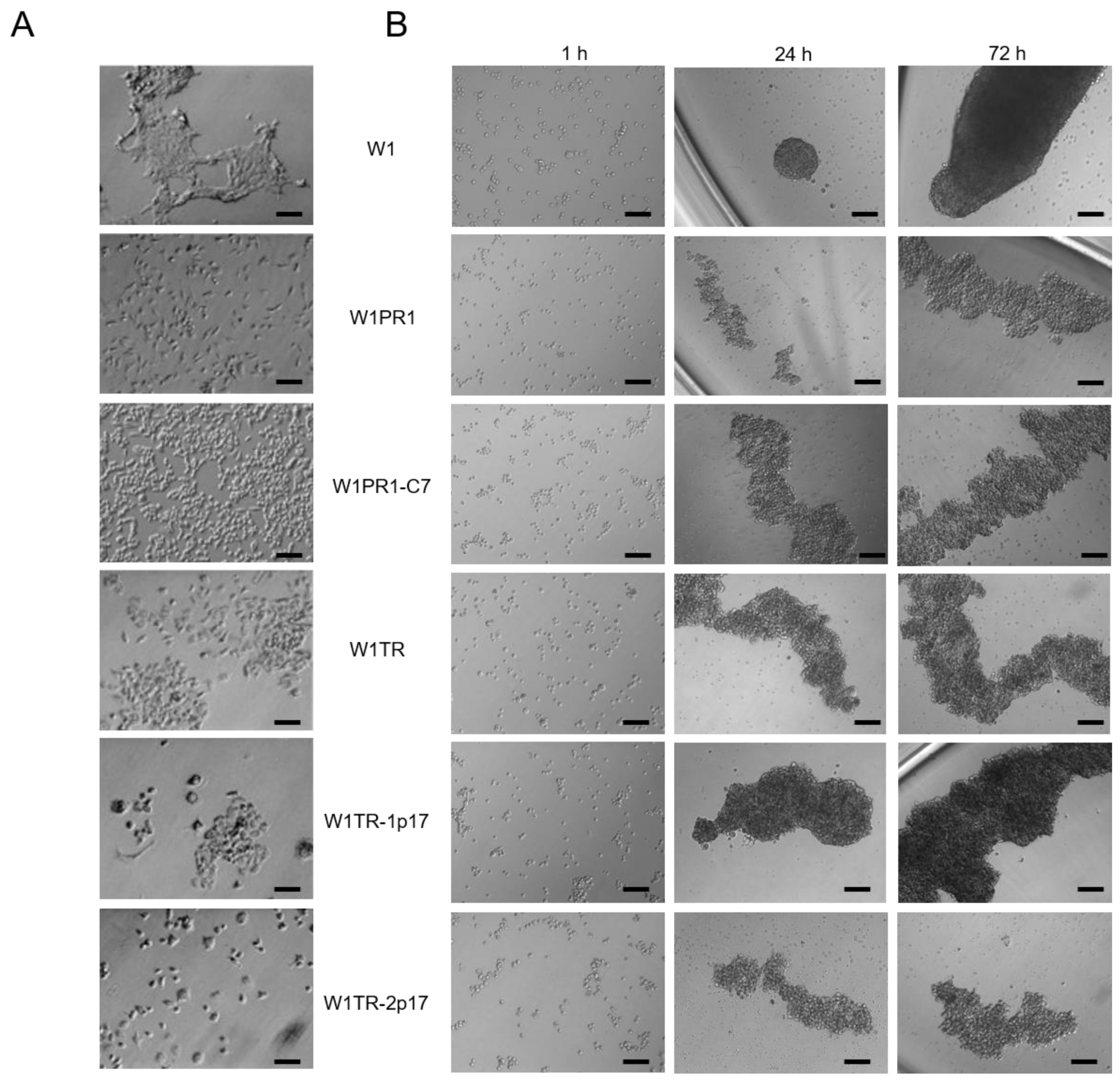
2.4. The Morphological Characteristics of Cell Lines under 2D and 3D Cell Culture Conditions
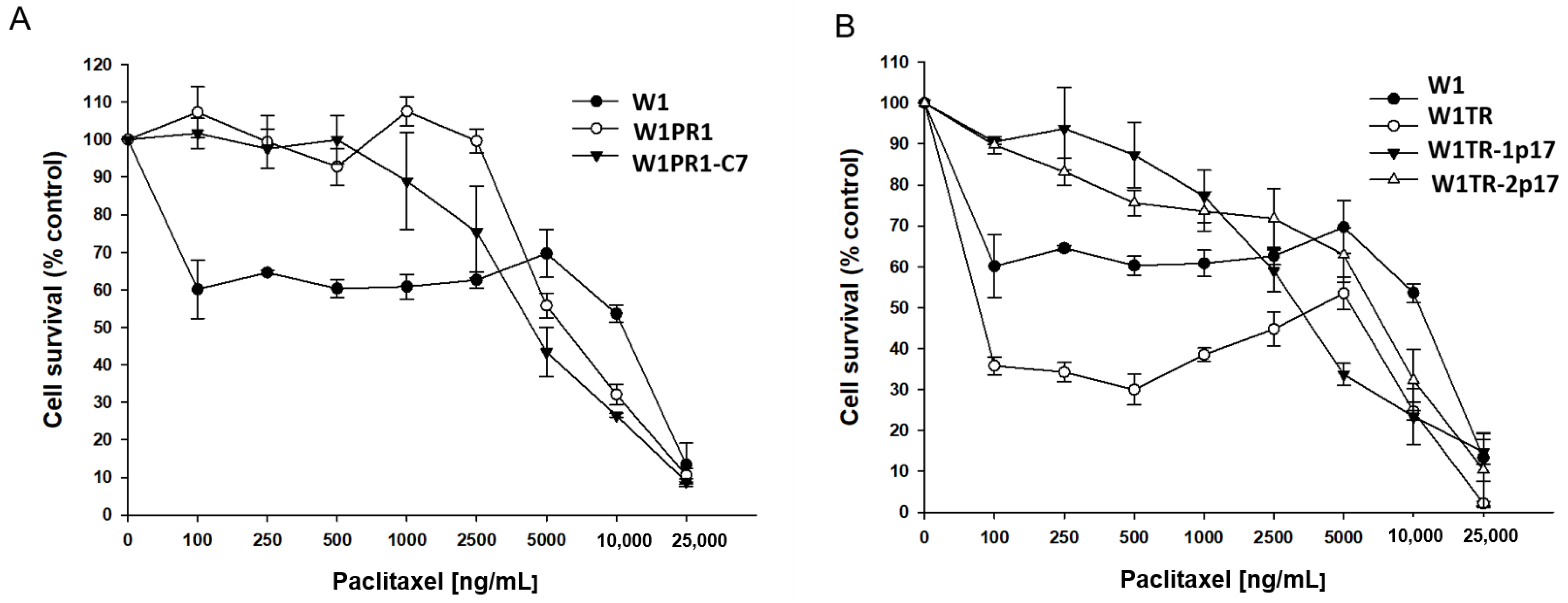
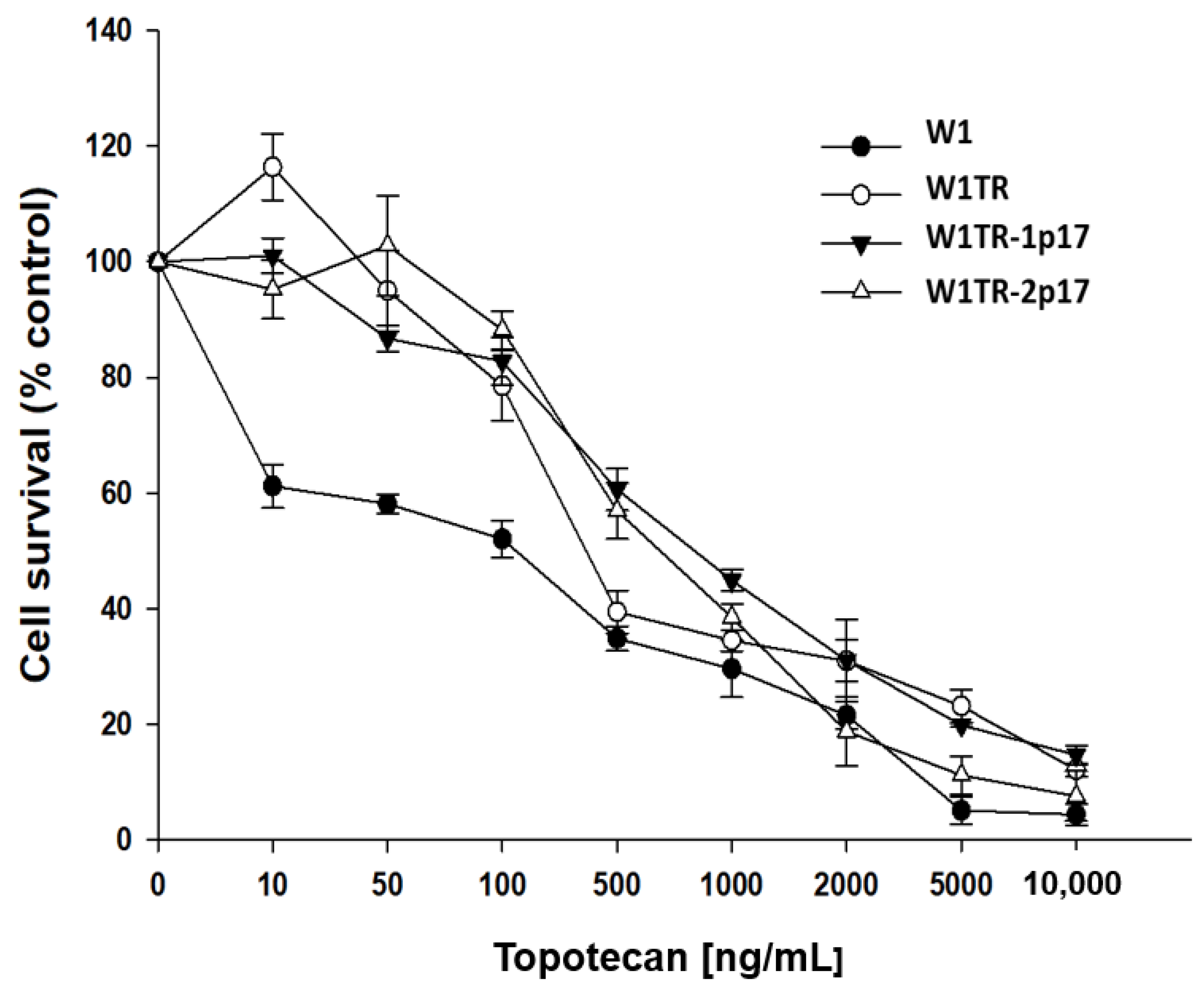
2.5. Analysis of PAC and TOP Resistance of Cells Growing under 2D and 3D Cell Culture Conditions
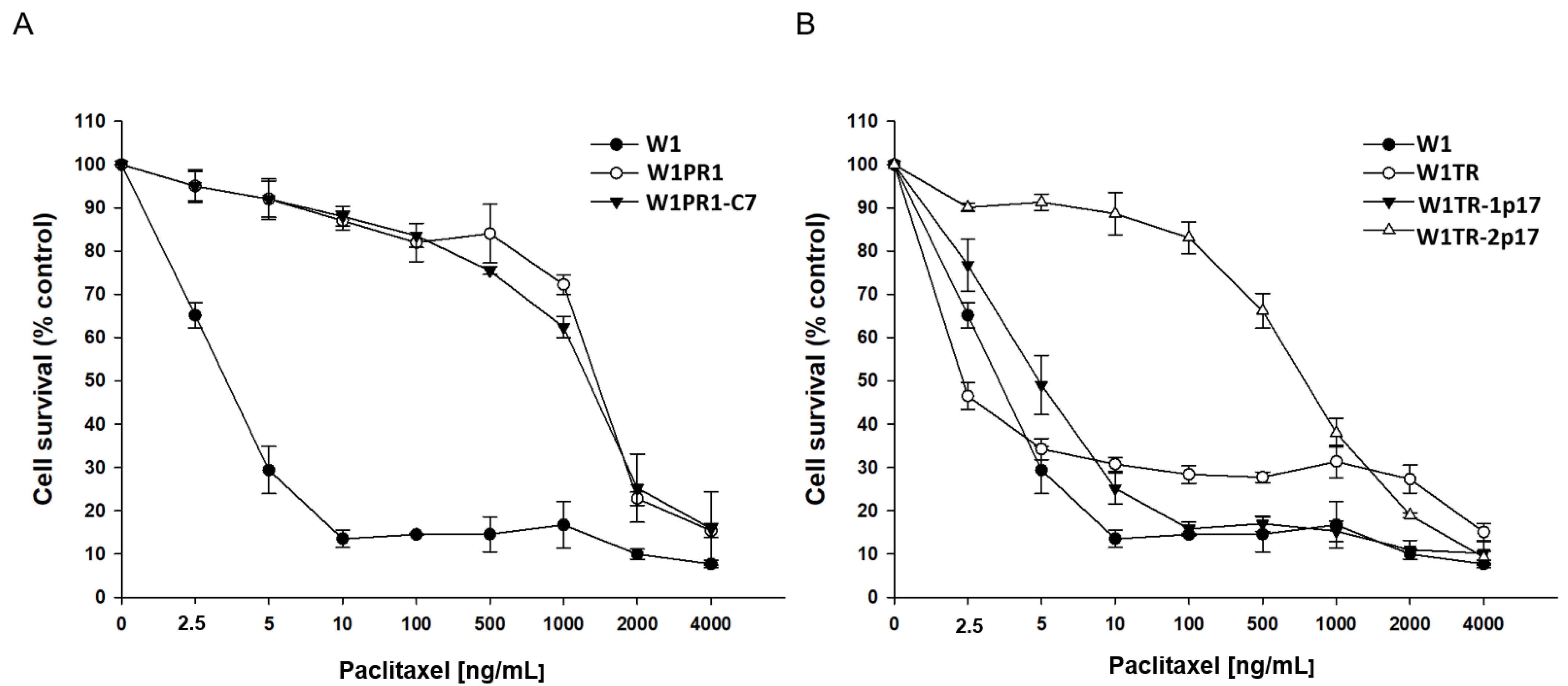
2.5.1. Analysis of PAC and TOP Resistance in 2D Conditions
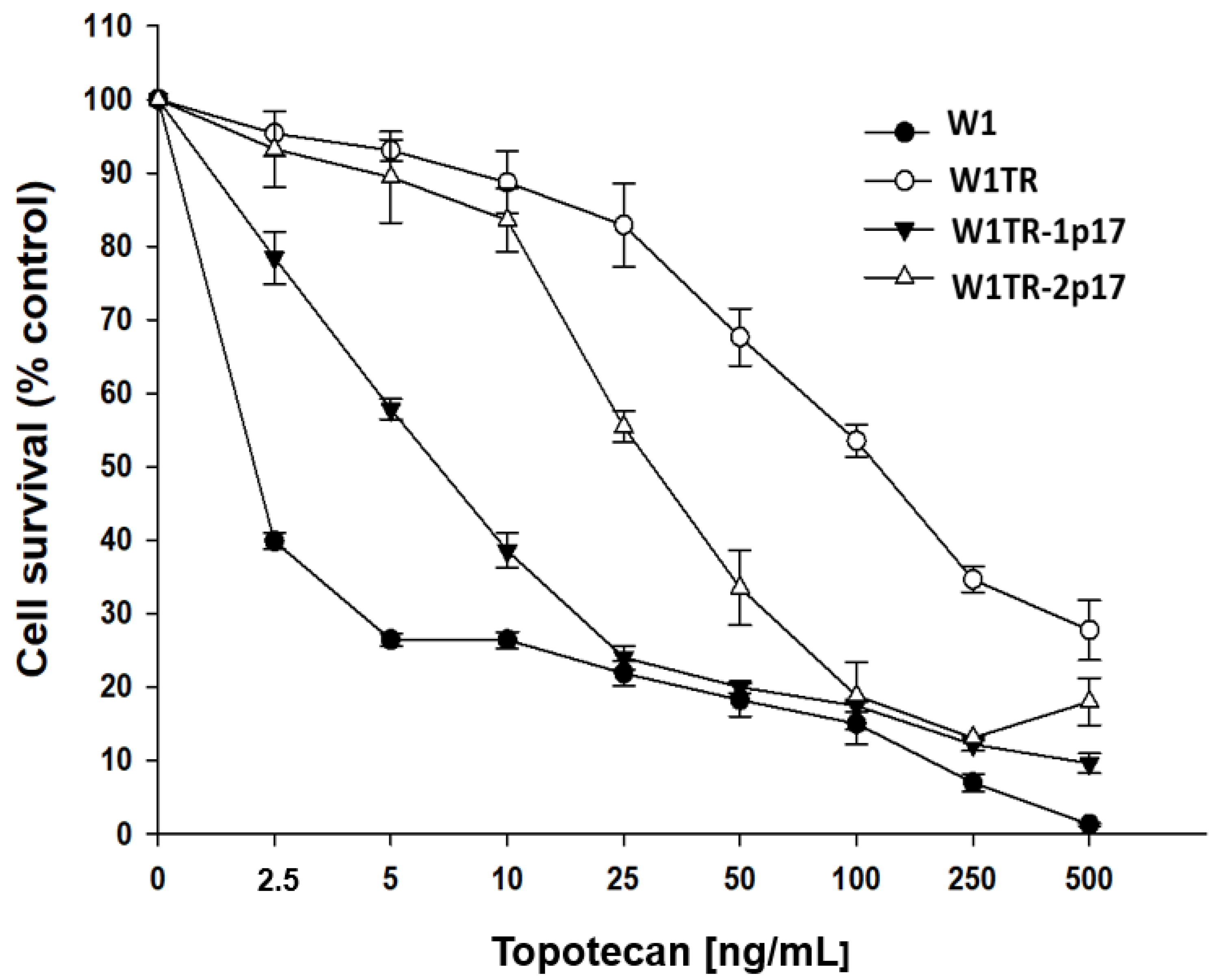
2.5.2. Analysis of PAC and TOP Resistance of Cells Growing under 3D Cell Culture Conditions
3. Discussion
4. Materials and Method
4.1. Cell Lines and Cell Culture
4.2. Stable Transfection with ALDH1A1 Gene Knockout Plasmids
4.3. MTT Assay
4.4. Examination of Gene Expression by Q-PCR
4.5. Western Blot Anlysis
4.6. Immunofluorescence Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Webb, P.M.; Jordan, S.J. Epidemiology of Epithelial Ovarian Cancer. Best Pr. Res. Clin. Obs. Gynaecol. 2017, 41, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Parmar, M.K.B.; Ledermann, J.A.; Colombo, N.; du Bois, A.; Delaloye, J.-F.; Kristensen, G.B.; Wheeler, S.; Swart, A.M.; Qian, W.; Torri, V.; et al. Paclitaxel plus Platinum-Based Chemotherapy versus Conventional Platinum-Based Chemotherapy in Women with Relapsed Ovarian Cancer: The ICON4/AGO-OVAR-2.2 Trial. Lancet 2003, 361, 2099–2106. [Google Scholar] [CrossRef] [PubMed]
- Webber, K.; Friedlander, M. Chemotherapy for Epithelial Ovarian, Fallopian Tube and Primary Peritoneal Cancer. Best Pract. Res. Clin. Obstet. Gynaecol. 2017, 41, 126–138. [Google Scholar] [CrossRef] [PubMed]
- Norouzi-Barough, L.; Sarookhani, M.R.; Sharifi, M.; Moghbelinejad, S.; Jangjoo, S.; Salehi, R. Molecular Mechanisms of Drug Resistance in Ovarian Cancer. J. Cell. Physiol. 2018, 233, 4546–4562. [Google Scholar] [CrossRef] [PubMed]
- Leslie, E.M.; Deeley, R.G.; Cole, S.P.C. Multidrug Resistance Proteins: Role of P-Glycoprotein, MRP1, MRP2, and BCRP (ABCG2) in Tissue Defense. Toxicol. Appl. Pharmacol. 2005, 204, 216–237. [Google Scholar] [CrossRef]
- Wu, C.-P.; Calcagno, A.M.; Ambudkar, S.V. Reversal of ABC Drug Transporter-Mediated Multidrug Resistance in Cancer Cells: Evaluation of Current Strategies. Curr. Mol. Pharm. 2008, 1, 93–105. [Google Scholar] [CrossRef]
- Amawi, H.; Sim, H.-M.; Tiwari, A.K.; Ambudkar, S.V.; Shukla, S. ABC Transporter-Mediated Multidrug-Resistant Cancer. In Drug Transporters in Drug Disposition, Effects and Toxicity; Liu, X., Pan, G., Eds.; Advances in Experimental Medicine and Biology; Springer: Singapore, 2019; pp. 549–580. ISBN 9789811376474. [Google Scholar]
- Lehne, G. P-Glycoprotein as a Drug Target in the Treatment of Multidrug Resistant Cancer. Curr. Drug Targets 2000, 1, 85–99. [Google Scholar] [CrossRef]
- Maliepaard, M.; van Gastelen, M.A.; de Jong, L.A.; Pluim, D.; Ruevekamp-Helmers, M.C.; Floot, B.G.J.; Schellens, J.H.M. Overexpression of the BCRP/MXR/ABCP Gene in a Topotecan-Selected Ovarian Tumor Cell Line. 6. Cancer Res. 1999, 59, 4559–4563. [Google Scholar]
- Correia, A.L.; Bissell, M.J. The Tumor Microenvironment Is a Dominant Force in Multidrug Resistance. Drug Resist. Updates 2012, 15, 39–49. [Google Scholar] [CrossRef]
- Cancer-Associated Fibroblasts in Tumor Microenvironment—Accomplices in Tumor Malignancy|Elsevier Enhanced Reader. Available online: https://reader.elsevier.com/reader/sd/pii/S0008874917302228?token=3F5F2EC8C1AA1D8ABBD54E618E3B9C0F1CBD41F79941B230D5E78EB2CF6534A367D905C2C59367DEE3A088B8BAAC2159&originRegion=eu-west-1&originCreation=20210914142221 (accessed on 14 September 2021).
- Netti, P.A.; Berk, D.A.; Swartz, M.A.; Grodzinsky, A.J.; Jain, R.K. Role of Extracellular Matrix Assembly in Interstitial Transport in Solid Tumors. Cancer Res. 2000, 60, 2497–2503. [Google Scholar]
- Grantab, R.; Sivananthan, S.; Tannock, I.F. The Penetration of Anticancer Drugs through Tumor Tissue as a Function of Cellular Adhesion and Packing Density of Tumor Cells. Cancer Res. 2006, 66, 1033–1039. [Google Scholar] [CrossRef] [PubMed]
- Wantoch von Rekowski, K.; König, P.; Henze, S.; Schlesinger, M.; Zawierucha, P.; Januchowski, R.; Bendas, G. The Impact of Integrin-Mediated Matrix Adhesion on Cisplatin Resistance of W1 Ovarian Cancer Cells. Biomolecules 2019, 9, 788. [Google Scholar] [CrossRef] [PubMed]
- Wantoch von Rekowski, K.; König, P.; Henze, S.; Schlesinger, M.; Zawierucha, P.; Januchowski, R.; Bendas, G. Insight into Cisplatin-Resistance Signaling of W1 Ovarian Cancer Cells Emerges MTOR and HSP27 as Targets for Sensitization Strategies. Int. J. Mol. Sci. 2020, 21, 9240. [Google Scholar] [CrossRef] [PubMed]
- Sterzyńska, K.; Klejewski, A.; Wojtowicz, K.; Świerczewska, M.; Andrzejewska, M.; Rusek, D.; Sobkowski, M.; Kędzia, W.; Brązert, J.; Nowicki, M.; et al. The Role of Matrix Gla Protein (MGP) Expression in Paclitaxel and Topotecan Resistant Ovarian Cancer Cell Lines. Int. J. Mol. Sci. 2018, 19, 2901. [Google Scholar] [CrossRef]
- Senthebane, D.A.; Jonker, T.; Rowe, A.; Thomford, N.E.; Munro, D.; Dandara, C.; Wonkam, A.; Govender, D.; Calder, B.; Soares, N.C.; et al. The Role of Tumor Microenvironment in Chemoresistance: 3D Extracellular Matrices as Accomplices. Int. J. Mol. Sci. 2018, 19, 2861. [Google Scholar] [CrossRef]
- Senthebane, D.A.; Rowe, A.; Thomford, N.E.; Shipanga, H.; Munro, D.; Mazeedi, M.A.M.A.; Almazyadi, H.A.M.; Kallmeyer, K.; Dandara, C.; Pepper, M.S.; et al. The Role of Tumor Microenvironment in Chemoresistance: To Survive, Keep Your Enemies Closer. Int. J. Mol. Sci. 2017, 18, 1586. [Google Scholar] [CrossRef]
- Sterzyńska, K.; Klejewski, A.; Wojtowicz, K.; Świerczewska, M.; Nowacka, M.; Kaźmierczak, D.; Andrzejewska, M.; Rusek, D.; Brązert, M.; Brązert, J.; et al. Mutual Expression of ALDH1A1, LOX, and Collagens in Ovarian Cancer Cell Lines as Combined CSCs- and ECM-Related Models of Drug Resistance Development. Int. J. Mol. Sci. 2018, 20, 54. [Google Scholar] [CrossRef]
- Sterzyńska, K.; Kaźmierczak, D.; Klejewski, A.; Świerczewska, M.; Wojtowicz, K.; Nowacka, M.; Brązert, J.; Nowicki, M.; Januchowski, R. Expression of Osteoblast-Specific Factor 2 (OSF-2, Periostin) Is Associated with Drug Resistance in Ovarian Cancer Cell Lines. Int. J. Mol. Sci. 2019, 20, 3927. [Google Scholar] [CrossRef]
- Klejewski, A.; Sterzyńska, K.; Wojtowicz, K.; Świerczewska, M.; Partyka, M.; Brązert, M.; Nowicki, M.; Zabel, M.; Januchowski, R. The Significance of Lumican Expression in Ovarian Cancer Drug-Resistant Cell Lines. Oncotarget 2017, 8, 74466–74478. [Google Scholar] [CrossRef]
- Januchowski, R.; Świerczewska, M.; Sterzyńska, K.; Wojtowicz, K.; Nowicki, M.; Zabel, M. Increased Expression of Several Collagen Genes Is Associated with Drug Resistance in Ovarian Cancer Cell Lines. J. Cancer 2016, 7, 1295–1310. [Google Scholar] [CrossRef]
- Moitra, K.; Lou, H.; Dean, M. Multidrug Efflux Pumps and Cancer Stem Cells: Insights into Multidrug Resistance and Therapeutic Development. Clin. Pharmacol. Ther. 2011, 89, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Al-Alem, L.F.; Pandya, U.M.; Baker, A.T.; Bellio, C.; Zarrella, B.D.; Clark, J.; DiGloria, C.M.; Rueda, B.R. Ovarian Cancer Stem Cells: What Progress Have We Made? Int. J. Biochem. Cell Biol. 2019, 107, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Nwani, N.G.; Condello, S.; Wang, Y.; Swetzig, W.M.; Barber, E.; Hurley, T.; Matei, D. A Novel ALDH1A1 Inhibitor Targets Cells with Stem Cell Characteristics in Ovarian Cancer. Cancers 2019, 11, 502. [Google Scholar] [CrossRef]
- Clevers, H. The Cancer Stem Cell: Premises, Promises and Challenges. Nat. Med. 2011, 17, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.H.; Kim, B.; Cho, U.; Azmi, A.S.; Song, Y.S. Association of ALDH1A1-NEK-2 Axis in Cisplatin Resistance in Ovarian Cancer Cells. Heliyon 2020, 6, e05442. [Google Scholar] [CrossRef] [PubMed]
- ALDH1A1+ Ovarian Cancer Stem Cells Co-Expressing Surface Markers CD24, EPHA1 and CD9 Form Tumours In Vivo|Elsevier Enhanced Reader. Available online: https://reader.elsevier.com/reader/sd/pii/S0014482720302317?token=DD92BF30B6651F26F7E847502B429C4CE1C245C8A5E0414F6BBF46C22C8C8CF7E41FDC753F8B8734448BA207DE975CD0&originRegion=eu-west-1&originCreation=20210914145051 (accessed on 14 September 2021).
- Inhibition of ALDH1A1 Activity Decreases Expression of Drug Transporters and Reduces Chemotherapy Resistance in Ovarian Cancer Cell Lines|Elsevier Enhanced Reader. Available online: https://reader.elsevier.com/reader/sd/pii/S1357272516301911?token=CB7EE6B98150BD6462E88D85FC4AA3B748FB5F72D780DFAD04CD00C30DD384310E7E36A1268C867CB80A624F6B06B7E0 (accessed on 17 September 2020).
- The Stem Cell Markers Oct4A, Nanog and c-Myc Are Expressed in Ascites Cells and Tumor Tissue of Ovarian Cancer Patients|SpringerLink. Available online: https://link.springer.com/article/10.1007%2Fs13402-013-0142-8 (accessed on 20 September 2021).
- Landen, C.N.; Goodman, B.; Katre, A.A.; Steg, A.D.; Nick, A.M.; Stone, R.L.; Miller, L.D.; Mejia, P.V.; Jennings, N.B.; Gershenson, D.M.; et al. Targeting Aldehyde Dehydrogenase Cancer Stem Cells in Ovarian Cancer. Mol Cancer 2010, 9, 3186–3199. [Google Scholar] [CrossRef]
- Kaipio, K.; Chen, P.; Roering, P.; Huhtinen, K.; Mikkonen, P.; Östling, P.; Lehtinen, L.; Mansuri, N.; Korpela, T.; Potdar, S.; et al. ALDH1A1-Related Stemness in High-Grade Serous Ovarian Cancer Is a Negative Prognostic Indicator but Potentially Targetable by EGFR/MTOR-PI3K/Aurora Kinase Inhibitors. J. Pathol. 2020, 250, 159–169. [Google Scholar] [CrossRef]
- Sherman-Baust, C.A.; Weeraratna, A.T.; Rangel, L.B.A.; Pizer, E.S.; Cho, K.R.; Schwartz, D.R.; Shock, T.; Morin, P.J. Remodeling of the Extracellular Matrix through Overexpression of Collagen VI Contributes to Cisplatin Resistance in Ovarian Cancer Cells. Cancer Cell 2003, 3, 377–386. [Google Scholar] [CrossRef]
- Morin, P.J. Drug Resistance and the Microenvironment: Nature and Nurture. Drug Resist. Updates 2003, 6, 169–172. [Google Scholar] [CrossRef]
- Helleman, J.; Jansen, M.P.H.M.; Span, P.N.; van Staveren, I.L.; Massuger, L.F.A.G.; Gelder, M.E.M.; Sweep, F.C.G.J.; Ewing, P.C.; van der Burg, M.E.L.; Stoter, G.; et al. Molecular Profiling of Platinum Resistant Ovarian Cancer. Int. J. Cancer 2006, 118, 1963–1971. [Google Scholar] [CrossRef]
- Chien, J.; Kuang, R.; Landen, C.; Shridhar, V. Platinum-Sensitive Recurrence in Ovarian Cancer: The Role of Tumor Microenvironment. Front. Oncol. 2013, 3, 251. [Google Scholar] [CrossRef] [PubMed]
- Januchowski, R.; Zawierucha, P.; Andrzejewska, M.; Ruciński, M.; Zabel, M. Microarray-Based Detection and Expression Analysis of ABC and SLC Transporters in Drug-Resistant Ovarian Cancer Cell Lines. Biomed. Pharmacother. 2013, 67, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Januchowski, R.; Wojtowicz, K.; Sujka-Kordowska, P.; Andrzejewska, M.; Zabel, M. MDR Gene Expression Analysis of Six Drug-Resistant Ovarian Cancer Cell Lines. BioMed Res. Int. 2013, 2013, 241763. [Google Scholar] [CrossRef] [PubMed]
- Januchowski, R.; Zawierucha, P.; Ruciński, M.; Zabel, M. Microarray-Based Detection and Expression Analysis of Extracellular Matrix Proteins in Drug-resistant Ovarian Cancer Cell Lines. Oncol. Rep. 2014, 32, 1981–1990. [Google Scholar] [CrossRef] [PubMed]
- Sterzyńska, K.; Klejewski, A.; Wojtowicz, K.; Świerczewska, M.; Nowicki, M.; Brązert, J.; Januchowski, R. Myotilin, a New Topotecan Resistant Protein in Ovarian Cancer Cell Lines. J. Cancer 2018, 9, 4413–4421. [Google Scholar] [CrossRef] [PubMed]
- Nowacka, M.; Sterzynska, K.; Andrzejewska, M.; Nowicki, M.; Januchowski, R. Drug Resistance Evaluation in Novel 3D In Vitro Model. Biomed. Pharmacother. 2021, 138, 111536. [Google Scholar] [CrossRef] [PubMed]
- Ozben, T. Mechanisms and Strategies to Overcome Multiple Drug Resistance in Cancer. FEBS Lett. 2006, 580, 2903–2909. [Google Scholar] [CrossRef]
- Pignata, S.; C Cecere, S.; Du Bois, A.; Harter, P.; Heitz, F. Treatment of Recurrent Ovarian Cancer. Ann. Oncol. 2017, 28, viii51–viii56. [Google Scholar] [CrossRef]
- Tirino, V.; Desiderio, V.; Paino, F.; De Rosa, A.; Papaccio, F.; La Noce, M.; Laino, L.; De Francesco, F.; Papaccio, G. Cancer Stem Cells in Solid Tumors: An Overview and New Approaches for Their Isolation and Characterization. FASEB J. 2013, 27, 13–24. [Google Scholar] [CrossRef]
- The Role of Aldehyde Dehydrogenase (ALDH) in Cancer Drug Resistance|Elsevier Enhanced Reader. Available online: https://reader.elsevier.com/reader/sd/pii/S0753332213000590?token=4EA5C8DCA1128518F46D9140972528C5B0C4F11C7D449671190A08CED4E87DB0F507FD83E1D48578A546C0197167A6D3&originRegion=eu-west-1&originCreation=20211103135350 (accessed on 3 November 2021).
- Steg, A.D.; Bevis, K.S.; Katre, A.A.; Ziebarth, A.; Dobbin, Z.C.; Alvarez, R.D.; Zhang, K.; Conner, M.; Landen, C.N. Stem Cell Pathways Contribute to Clinical Chemoresistance in Ovarian Cancer. Clin. Cancer Res. 2012, 18, 869–881. [Google Scholar] [CrossRef]
- Liu, S.; Liu, C.; Min, X.; Ji, Y.; Wang, N.; Liu, D.; Cai, J.; Li, K. Prognostic Value of Cancer Stem Cell Marker Aldehyde Dehydrogenase in Ovarian Cancer: A Meta-Analysis. PLoS ONE 2013, 8, e81050. [Google Scholar] [CrossRef] [PubMed]
- Aldehyde Dehydrogenase in Combination with CD133 Defines Angiogenic Ovarian Cancer Stem Cells That Portend Poor Patient Survival|Cancer Research. Available online: https://cancerres.aacrjournals.org/content/71/11/3991.long (accessed on 13 October 2021).
- ALDH1-Bright Epithelial Ovarian Cancer Cells Are Associated with CD44 Expression, Drug Resistance, and Poor Clinical Outcome-ClinicalKey. Available online: https://www.clinicalkey.com/#!/content/playContent/1-s2.0-S0002944011010790?returnurl=https:%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS0002944011010790%3Fshowall%3Dtrue&referrer= (accessed on 17 September 2020).
- Aldehyde Dehydrogenase 1A1 (ALDH1A1) Expression by Immunohistochemistry Is Associated with Chemo-Refractoriness in Patients with High-Grade Ovarian Serous Carcinoma|Elsevier Enhanced Reader. Available online: https://reader.elsevier.com/reader/sd/pii/S0046817717302976?token=8A99D1BB1D312814F964D405FC5A06B2279D90C7336AA2E75172655CF61CBA3A8FD1E98E87FAE6120CFC613D720977EC&originRegion=eu-west-1&originCreation=20211013083007 (accessed on 13 October 2021).
- Ayub, T.H.; Keyver-Paik, M.-D.; Debald, M.; Rostamzadeh, B.; Thiesler, T.; Schröder, L.; Barchet, W.; Abramian, A.; Kaiser, C.; Kristiansen, G.; et al. Accumulation of ALDH1-Positive Cells after Neoadjuvant Chemotherapy Predicts Treatment Resistance and Prognosticates Poor Outcome in Ovarian Cancer. Oncotarget 2015, 6, 16437–16448. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Du, F.; Jiang, L.; Zhu, Y.; Chen, Z.; Liu, Y.; Hong, T.; Wang, T.; Mao, Y.; Wu, X.; et al. A2780 Human Ovarian Cancer Cells with Acquired Paclitaxel Resistance Display Cancer Stem Cell Properties. Oncol. Lett. 2013, 6, 1295–1298. [Google Scholar] [CrossRef]
- Hilton, J. Role of Aldehyde Dehydrogenase in Cyclophosphamide-Resistant L1210 Leukemia. Cancer Res. 1984, 44, 6. [Google Scholar]
- Moreb, J.; Zucali, J.R.; Zhang, Y.; Colvin, M.O.; Gross, M.A. Role of Aldehyde Dehydrogenase in the Protection of Hematopoietic Progenitor Cells from 4-Hydroperoxycyclophosphamide by Inter Leukin Ißand Tumor Necrosis Factor. Cancer Res. 1992, 52, 1770–1774. [Google Scholar] [PubMed]
- Alison, M.R.; Lin, W.-R.; Lim, S.M.L.; Nicholson, L.J. Cancer Stem Cells: In the Line of Fire. Cancer Treat. Rev. 2012, 38, 589–598. [Google Scholar] [CrossRef]
- Yang, Y.; Zhou, W.; Xia, J.; Gu, Z.; Wendlandt, E.; Zhan, X.; Janz, S.; Tricot, G.; Zhan, F. NEK2 Mediates ALDH1A1-Dependent Drug Resistance in Multiple Myeloma. Oncotarget 2014, 5, 11986–11997. [Google Scholar] [CrossRef]
- Kim, D.; Choi, B.; Ryoo, I.; Kwak, M.-K. High NRF2 Level Mediates Cancer Stem Cell-like Properties of Aldehyde Dehydrogenase (ALDH)-High Ovarian Cancer Cells: Inhibitory Role of All-Trans Retinoic Acid in ALDH/NRF2 Signaling. Cell Death Dis. 2018, 9, 896. [Google Scholar] [CrossRef]
- Januchowski, R.; Zawierucha, P.; Ruciński, M.; Andrzejewska, M.; Wojtowicz, K.; Nowicki, M.; Zabel, M. Drug Transporter Expression Profiling in Chemoresistant Variants of the A2780 Ovarian Cancer Cell Line. Biomed. Pharmacother. 2014, 68, 447–453. [Google Scholar] [CrossRef]
- Januchowski, R.; Wojtowicz, K.; Andrzejewska, M.; Zabel, M. Expression of MDR1 and MDR3 Gene Products in Paclitaxel-, Doxorubicin- and Vincristine-Resistant Cell Lines. Biomed. Pharmacother. 2014, 68, 111–117. [Google Scholar] [CrossRef]
- Vanhoefer, U.; Müller, M.R.; Hilger, R.A.; Lindtner, B.; Klaassen, U.; Schleucher, N.; Rustum, Y.M.; Seeber, S.; Harstrick, A. Reversal of MDR1-Associated Resistance to Topotecan by PAK-200S, a New Dihydropyridine Analogue, in Human Cancer Cell Lines. Br. J. Cancer 1999, 81, 1304–1310. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Januchowski, R.; Sterzyńska, K.; Zaorska, K.; Sosińska, P.; Klejewski, A.; Brązert, M.; Nowicki, M.; Zabel, M. Analysis of MDR Genes Expression and Cross-Resistance in Eight Drug Resistant Ovarian Cancer Cell Lines. J. Ovarian Res. 2016, 9, 65. [Google Scholar] [CrossRef] [PubMed]
- Weiswald, L.-B.; Bellet, D.; Dangles-Marie, V. Spherical Cancer Models in Tumor Biology. Neoplasia 2015, 17, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, G. Multicellular Spheroids as an in Vitro Tumor Model. Cancer Lett. 1998, 131, 29–34. [Google Scholar] [CrossRef]
- Erlanson, M.; Daniel-Szolgay, E.; Carlsson, J. Relations between the Penetration, Binding and Average Concentration of Cytostatic Drugs in Human Tumour Spheroids. Cancer Chemother. Pharmacol. 1992, 29, 343–353. [Google Scholar] [CrossRef]
- Wang, J.; Lu, Z.; Gao, Y.; Wientjes, M.G.; Au, J.L.-S. Improving Delivery and Efficacy of Nanomedicines in Solid Tumors: Role of Tumor Priming. Nanomedicine 2011, 6, 1605–1620. [Google Scholar] [CrossRef]
- Pommier, Y.; Sun, Y.; Huang, S.N.; Nitiss, J.L. Roes of Eukaryotic Topoisomerases in Transcription, Replication and Genomic Stability. Nat. Rev. Mol. Cell Biol. 2016, 17, 703–721. [Google Scholar] [CrossRef]
- Staker, B.L.; Hjerrild, K.; Feese, M.D.; Behnke, C.A.; Burgin, A.B.; Stewart, L. The Mechanism of Topoisomerase I Poisoning by a Camptothecin Analog. Proc. Natl. Acad. Sci. USA 2002, 99, 15387–15392. [Google Scholar] [CrossRef]
- Uhm, J.H.; Dooley, N.P.; Kyritsis, A.P.; Rao, J.S.; Gladson, C.L. Vitronectin, a Glioma-Derived Extracellular Matrix Protein, Protects Tumor Cells from Apoptotic Death. Clin. Cancer Res. 1999, 5, 1587–1594. [Google Scholar]
- Stylianopoulos, T.; Diop-Frimpong, B.; Munn, L.L.; Jain, R.K. Diffusion Anisotropy in Collagen Gels and Tumors: The Effect of Fiber Network Orientation. Biophys. J. 2010, 99, 3119–3128. [Google Scholar] [CrossRef]
- Nederman, T.; Carlsson, J. Penetration and Binding of Vinblastine and 5-Fluorouracil in Cellular Spheroids. Cancer Chemother. Pharmacol. 1984, 13, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Imamura, Y.; Mukohara, T.; Shimono, Y.; Funakoshi, Y.; Chayahara, N.; Toyoda, M.; Kiyota, N.; Takao, S.; Kono, S.; Nakatsura, T.; et al. Comparison of 2D- and 3D-Culture Models as Drug-Testing Platforms in Breast Cancer. Oncol. Rep. 2015, 33, 1837–1843. [Google Scholar] [CrossRef] [PubMed]
- Myungjin Lee, J.; Mhawech-Fauceglia, P.; Lee, N.; Cristina Parsanian, L.; Gail Lin, Y.; Andrew Gayther, S.; Lawrenson, K. A Three-Dimensional Microenvironment Alters Protein Expression and Chemosensitivity of Epithelial Ovarian Cancer Cells in Vitro. Lab. Invest. 2013, 93, 528–542. [Google Scholar] [CrossRef] [PubMed]
- Comparison of 2D and 3D Cell Culture Models for Cell Growth, Gene Expression and Drug Resistance|Elsevier Enhanced Reader. Available online: https://reader.elsevier.com/reader/sd/pii/S0928493119305934?token=A81072D392015C0F3853335103228D48E9C60158BAE61A69162483955AB135BBAA3EB87B3FA7A19190F78188B23943EC&originRegion=eu-west-1&originCreation=20211103150017 (accessed on 3 November 2021).











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | PAC IC50 (ng/mL) | TOP IC50 (ng/mL) |
|---|---|---|
| W1 | 3.37 (3.18–3.62) | 2.06 (1.94–2.18) |
| W1PR1 | 1671 *** (1526–1788) | - |
| W1PR1-C7 | 1342 ** ^ (1234–1451) | - |
| W1TR | 3.96 (2.34–5.20) | 111 ** (98.4–123) |
| W1TR-1p17 | 5.53 * (4.05–8.43) | 7.18 *** ^^ (5.59–8.70) |
| W1TR-2p17 | 787 ** ^^ (682–843) | 31.1 *** ^^ (27.5–34.9) |
| Cell Line | PAC IC50 (ng/mL) | TOP IC50 (ng/mL) |
|---|---|---|
| W1 | 11,883 (9548–15,833) | 154 (65–279) |
| W1PR1 | 6022 ** (5302–7488) | - |
| W1PR1-C7 | 3971 *** (2933–5696) ^ p = 0.06 | - |
| W1TR | 6099 ** (5551–7353) | 384 * (242–473) |
| W1TR-1p17 | 3283 *** ^^ (2831–4218) | 814 *** ^^^ (629–1016) |
| W1TR-2p17 | 7183 * (5095–9544) | 697 (406–797) *** ^ |
| Transcript | Sequence (5′-3′ Direction) | ENST Number http://www.ensembl.org accessed date: 10 September 2021 | Product Size (bp) |
|---|---|---|---|
| MDR1 | TGACAGCTACAGCACGGAAG TCTTCACCTCCAGGCTCAGT | 00000265724 | 131 bp |
| ALDH1A1 | GTTGTCAAACCAGCAGAGCA CTGTAGGCCCATAACCAGGA | 00000165092 | 115 bp |
| BCRP | TTCGGCTTGCAACAACTATG TCCAGACACACCACGGATAA | 00000237612 | 128 bp |
| COL3A1 | AAGGTCCAGCTGGGATACCT CACCCTTTAATCCAGGAGCA | 00000304636 | 105 bp |
| GAPDH | GAAGGTGAAGGTCGGAGTCA GACAAGCTTCCCGTTCTCAG | 00000229239 | 199 bp |
| β-actin | TCTGGCACCACACCTTCTAC GATAGCACAGCCTGGATAGC | 00000331789 | 169 bp |
| HRPT1 | CTGAGGATTTGGAAAGGGTG AATCCAGCAGGTCAGCAAAG | 00000298556 | 156 bp |
| β2M | CGCTACTCTCTCTTTCTGGC ATGTCGGATGGATGAAACCC | 00000558401 | 133 bp |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nowacka, M.; Ginter-Matuszewska, B.; Świerczewska, M.; Sterzyńska, K.; Nowicki, M.; Januchowski, R. Effect of ALDH1A1 Gene Knockout on Drug Resistance in Paclitaxel and Topotecan Resistant Human Ovarian Cancer Cell Lines in 2D and 3D Model. Int. J. Mol. Sci. 2022, 23, 3036. https://doi.org/10.3390/ijms23063036
Nowacka M, Ginter-Matuszewska B, Świerczewska M, Sterzyńska K, Nowicki M, Januchowski R. Effect of ALDH1A1 Gene Knockout on Drug Resistance in Paclitaxel and Topotecan Resistant Human Ovarian Cancer Cell Lines in 2D and 3D Model. International Journal of Molecular Sciences. 2022; 23(6):3036. https://doi.org/10.3390/ijms23063036
Chicago/Turabian StyleNowacka, Marta, Barbara Ginter-Matuszewska, Monika Świerczewska, Karolina Sterzyńska, Michał Nowicki, and Radosław Januchowski. 2022. "Effect of ALDH1A1 Gene Knockout on Drug Resistance in Paclitaxel and Topotecan Resistant Human Ovarian Cancer Cell Lines in 2D and 3D Model" International Journal of Molecular Sciences 23, no. 6: 3036. https://doi.org/10.3390/ijms23063036
APA StyleNowacka, M., Ginter-Matuszewska, B., Świerczewska, M., Sterzyńska, K., Nowicki, M., & Januchowski, R. (2022). Effect of ALDH1A1 Gene Knockout on Drug Resistance in Paclitaxel and Topotecan Resistant Human Ovarian Cancer Cell Lines in 2D and 3D Model. International Journal of Molecular Sciences, 23(6), 3036. https://doi.org/10.3390/ijms23063036