
Retinal Organoids and Retinal Prostheses: An Overview



Abstract
1. Introduction
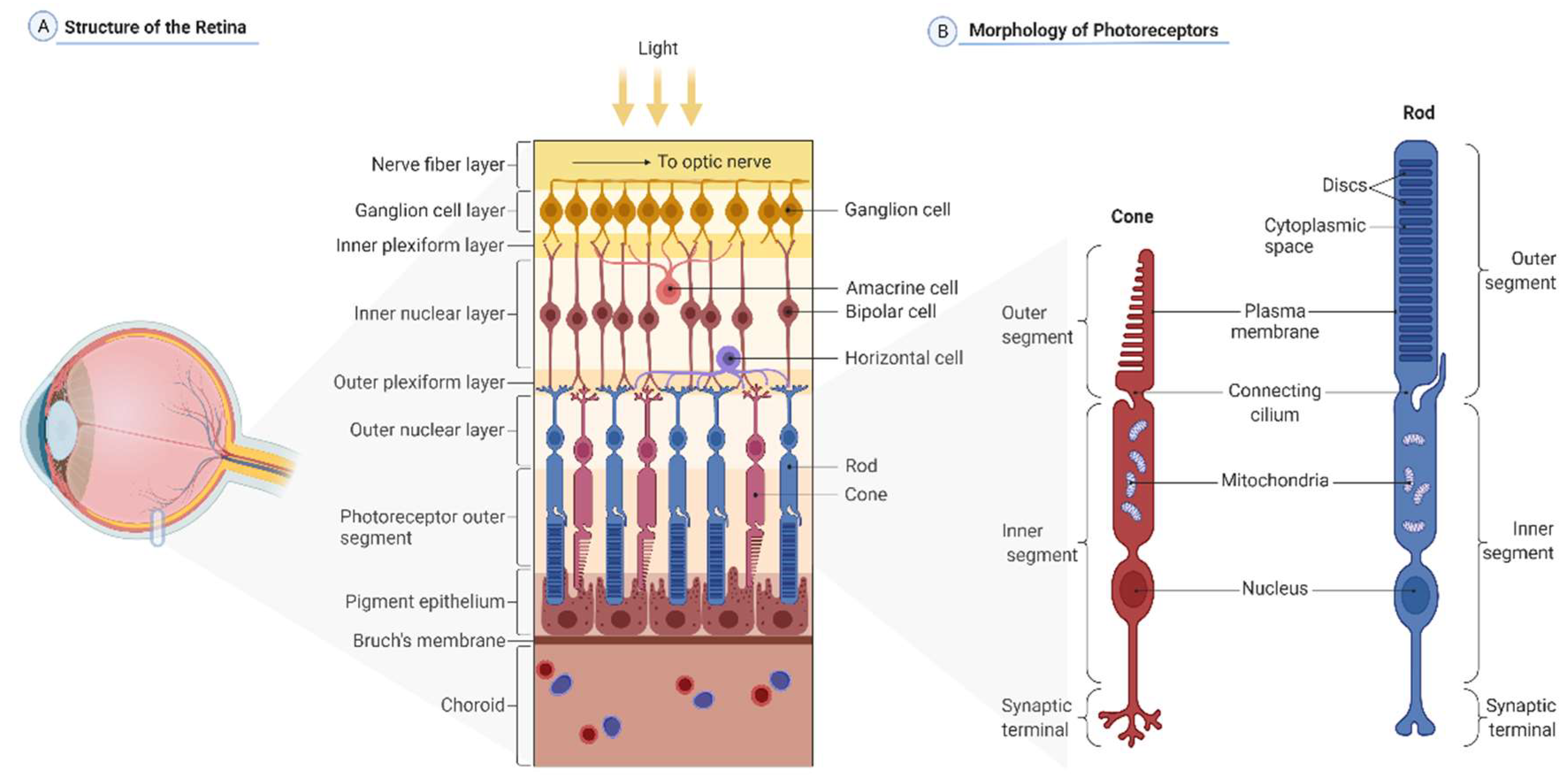
2. General Information about the Retina
2.1. Retinal Cell Population
2.2. Photoreceptors
3. Basics of Eye Development
4. Animal Models
5. Retinal Organoid Protocols
5.1. Two-Dimensional (2D) Retinal Differentiation
5.2. Three-Dimensional (3D) Retinal Differentiation
5.3. 2D/3D Retinal Differentiation Technique
5.4. Protocol Improvement
5.5. Differentiation Phases of 3D Retinal Organoids
5.6. Brain Organoid-Derived Neural Retina
5.7. Limitations of Retinal Organoids
6. Retinal Prosthesis to Restore Vision
6.1. Principle of Electronic Retinal Prosthesis
6.2. Types of Electronic Retinal Prostheses
6.3. Limitations of Electronic Retinal Prostheses
6.4. Organic Retinal Prostheses

{kind=link}
{kind=link}
| Organic Material | Configuration | Purpose | Cytotoxicity | Validation Models | Publication |
|---|---|---|---|---|---|
| P3HT | rrP3HT—el. donor PCBM—el. Acceptor | Neuronal stimulation | Propidium iodine/fluorescein diacetate staining assay and patch-clamp recordings | Primary culture of hippocampal neurons | Ghezzi et al., 2011 [137] |
| Single (donor)-component P3HT film | Subretinal stimulation | Propidium iodine/fluorescein diacetate staining assay and patch-clamp recordings on primary culture of hippocampal neurons | Sprague–Dawley albino rat retinal explants | Ghezzi et al., 2013 [17] | |
| P3HT—el. donor N2200—el. Acceptor | Epiretinal stimulation | TUNEL assay | Embryonic chick retina | Gautam et al., 2014 [143] | |
| P3HT—semiconductive layer PEDOT:PSS—intermediate conductive layer Silk fibroin—substrate | Fully organic SILK-PEDOT:PSS-P3HT prosthesis | No Inflammation after 6 months in vivo | In vivo RCS rat models | Maya-Vetencourt et al., 2017 [136] | |
| PEDOT:PSS—anode P3HT:PCBM—Semiconductor layer Titanium—cathode PDMS—substrate material; | Foldable, wide-field epiretinal prosthesis POLYRETINA | XTT cell viability assay | Ex vivo explants from rd10 mouse model | Ferlauto et al., 2018 [138] | |
| Conjugated polymer nanoparticles P3HT on PET substrate | Liquid retinal prosthesis (subretinal injection) | No inflammation after 240 DPI | RCS rat retinal explants; In vivo RCS rat model | Maya-Vetencourt et al., 2020 [106] | |
| PCBM | MEH-PPV—el. donor PCBM—el. Acceptor | Hybrid solid-liquid polymer photodiode | — | Photocurrent action spectrum measurements in cell culture medium working as a cathode | Antognazza et al., 2009 [144] |
| PDPP3T—el. donor PCBM—el. Acceptor | Near-Infrared Tandem Organic Photodiodes | — | Pulsed NIR illumination in a physiological environment | Simone et al., 2018 [145] | |
| PEDOT:PSS—anode PCPDTBT (or P3HT):PC60BM—BHJ Titanium—cathode | NIR-sensitive foldable and photovoltaic wide-field epiretinal prosthesis nirPOLYRETINA | XTT cell viability assay | Ex vivo explants from rd10 mouse models | Airaghi Leccardi et al., 2020 [131] | |
| PTCDI | H2Pc: p-type el. donor PTCDI—n-type el. Acceptor | Epiretinal stimulation | — | Embryonic chicken retina | Rand et al., 2018 [18] |
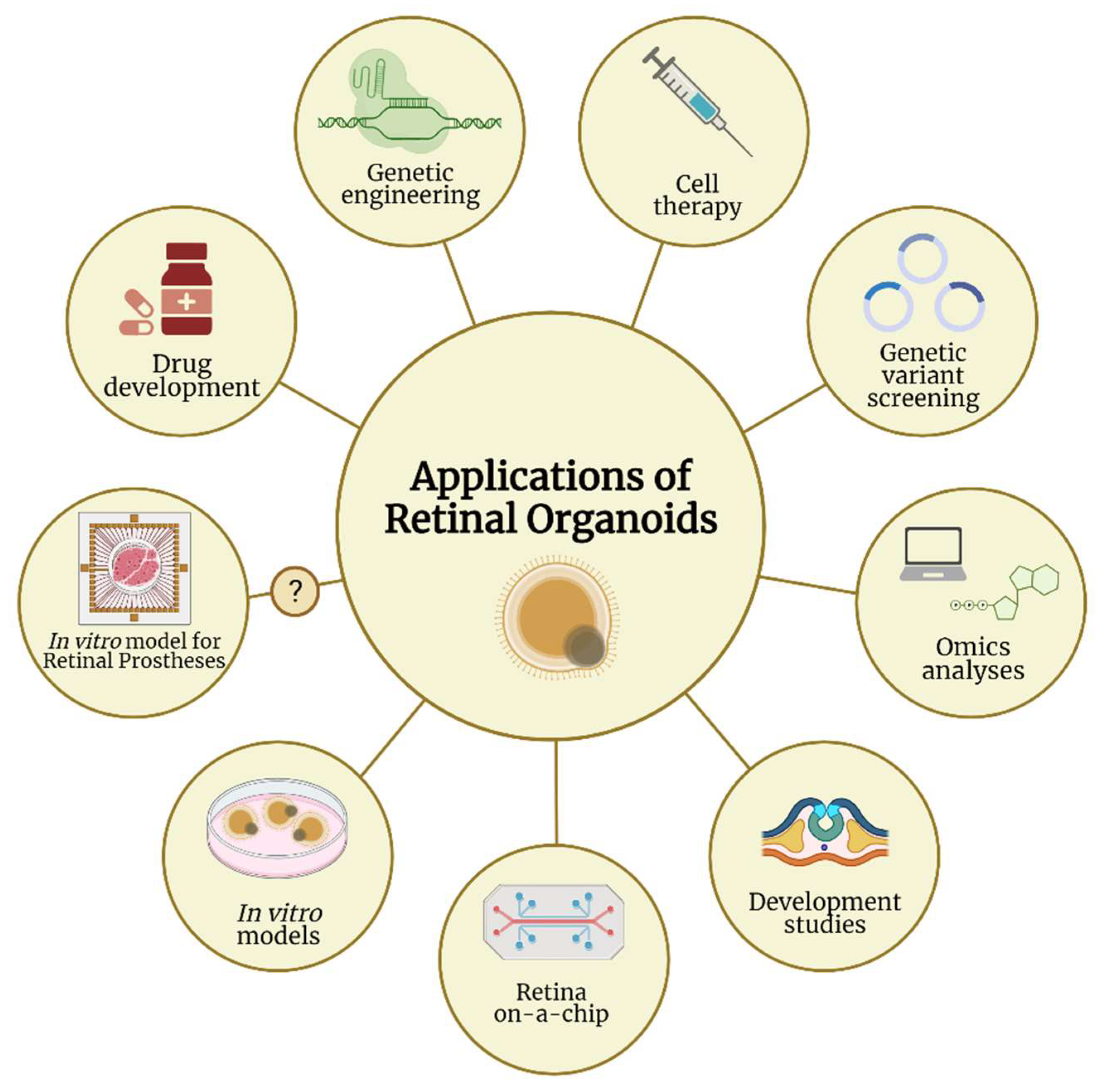
7. Utility of Retinal Organoids
7.1. Cell Replacement and Gene Therapies
7.2. Retinal Organoids as Human In Vitro Models
7.3. Retinal Organoids as In Vitro Models for Retinal Prostheses
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Frings, S.; Müller, F. Visuelles System—Auge und Sehen. In Duale Reihe Physiologie; Thieme-Verlag: Stuttgart, Germany, 2010; Available online: https://juser.fz-juelich.de/record/14543 (accessed on 9 February 2022).
- Aumüller, G.; Aust, G.; Engele, J.; Kirsch, J.; Maio, G.; Mayerhofer, A.; Mense, S.; Reißig, D.; Salvetter, J.; Schmidt, W.; et al. Duale Reihe Anatomie; Thieme: Stuttgart, Germany, 2017. [Google Scholar] [CrossRef]
- Vu, H.T.V.; Keeffe, J.E.; McCarty, C.A.; Taylor, H.R. Impact of unilateral and bilateral vision loss on quality of life. Br. J. Ophthalmol. 2005, 89, 360–363. [Google Scholar] [CrossRef] [PubMed]
- Scholl, H.P.N.; Strauss, R.W.; Singh, M.S.; Dalkara, D.; Roska, B.; Picaud, S.; Sahel, J.A. Emerging therapies for inherited retinal degeneration. Sci. Transl. Med. 2016, 8, rv6–rv368. [Google Scholar] [CrossRef] [PubMed]
- Sahel, J.A.; Marazova, K.; Audo, I. Clinical characteristics and current therapies for inherited retinal degenerations. Cold Spring Harb. Perspect. Med. 2014, 5, a017111. [Google Scholar] [CrossRef] [PubMed]
- Verbakel, S.K.; van Huet, R.A.C.; Boon, C.J.F.; den Hollander, A.I.; Collin, R.W.J.; Klaver, C.C.W.; Hoyng, C.B.; Roepman, R.; Klevering, B.J. Non-syndromic retinitis pigmentosa. Prog. Retin. Eye Res. 2018, 66, 157–186. [Google Scholar] [CrossRef]
- Parmeggiani, F. Clinics, epidemiology and genetics of retinitis pigmentosa. Curr. Genom. 2011, 12, 236–237. [Google Scholar] [CrossRef] [PubMed]
- Roska, B.; Sahel, J.A. Restoring vision. Nature 2018, 557, 359–367. [Google Scholar] [CrossRef]
- Frederick, J.; Bronson, J.D.; Baehr, W. Animal models of inherited retinal diseases. Methods Enzymol. 2000, 316, 515–526. [Google Scholar]
- Tang, Z.; Zhang, Y.; Wang, Y.; Zhang, D.; Shen, B.; Luo, M.; Gu, P. Progress of stem/progenitor cell-based therapy for retinal degeneration. J. Transl. Med. 2017, 15, 99. [Google Scholar] [CrossRef]
- Artero Castro, A.; Rodríguez Jimenez, F.J.; Jendelova, P.; Erceg, S. Deciphering retinal diseases through the generation of three dimensional stem cell-derived organoids: Concise Review. Stem Cells 2019, 37, 1496–1504. [Google Scholar] [CrossRef]
- Kruczek, K.; Swaroop, A. Pluripotent stem cell-derived retinal organoids for disease modeling and development of therapies. Stem Cells 2020, 38, 1206–1215. [Google Scholar] [CrossRef]
- Garg, S.J.; Federman, J. Optogenetics, visual prosthesis and electrostimulation for retinal dystrophies. Curr. Opin. Ophthalmol. 2013, 24, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.H.L.; Da Cruz, L. A review and update on the current status of retinal prostheses (bionic eye). Br. Med. Bull. 2014, 109, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Shim, S.; Eom, K.; Jeong, J.; Kim, S.J. Retinal Prosthetic Approaches to Enhance Visual Perception for Blind Patients. Micromachines 2020, 11, 535. [Google Scholar] [CrossRef] [PubMed]
- Benfenati, F.; Lanzani, G. New technologies for developing second generation retinal prostheses. Lab. Anim. 2018, 47, 71–75. [Google Scholar] [CrossRef]
- Ghezzi, D.; Antognazza, M.R.; MacCarone, R.; Bellani, S.; Lanzarini, E.; Martino, N.; Mete, M.; Pertile, G.; Bisti, S.; Lanzani, G.; et al. A polymer optoelectronic interface restores light sensitivity in blind rat retinas. Nat. Photonics 2013, 7, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Rand, D.; Jakešová, M.; Lubin, G.; Vėbraitė, I.; David-Pur, M.; Đerek, V.; Cramer, T.; Sariciftci, N.S.; Hanein, Y.; Głowacki, E.D. Direct Electrical Neurostimulation with Organic Pigment Photocapacitors. Adv. Mater. 2018, 30, 1–11. [Google Scholar] [CrossRef]
- Kolb, H. Simple Anatomy of the Retina. 2012. Available online: https://www.ncbi.nlm.nih.gov/books/NBK11533/ (accessed on 9 February 2022).
- Fuhrmann, S. Eye morphogenesis and patterning of the optic vesicle. Curr. Top. Dev. Biol. 2010, 93, 61–84. [Google Scholar] [CrossRef]
- Curcio, C.A.; Sloan, K.R.; Kalina, R.E.; Hendrickson, A.E. Human photoreceptor topography. J. Comp. Neurol. 1990, 292, 497–523. [Google Scholar] [CrossRef]
- Ahmad, K.M.; Klug, K.; Herr, S.; Sterling, P.; Schein, S. Cell density ratios in a foveal patch in macaque retina. Vis. Neurosci. 2003, 20, 189–209. [Google Scholar] [CrossRef]
- Anderson, D.H.; Fisher, S.K. The photoreceptors of diurnal squirrels: Outer segment structure, disc shedding, and protein renewal. J. Ultrastruct. Res. 1976, 55, 119–141. [Google Scholar] [CrossRef]
- Villegas, G.M. Ultrastructure of the human retina. J. Anat. 1964, 98, 501. [Google Scholar] [PubMed]
- De Robertis, E. Some observations on the ultrastructure and morphogenesis of photoreceptors. J. Gen. Physiol. 1960, 43, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Papermaster, D.; Schneider, B.G.; Desharse, J.C. Vesicular transport of newly synthesized opsin from the Golgi apparatus toward the rod outer segment. Ultrastructural immunocytochemical and autoradiographic evidence in Xenopus retinas. Investig. Ophthalmol. Vis. Sci. 1985, 26, 1386–1404. [Google Scholar]
- Steinberg, R.H.; Fisher, S.K.; Anderson, D.H. Disc morphogenesis in vertebrate photoreceptors. J. Comp. Neurol. 1980, 190, 501–518. [Google Scholar] [CrossRef]
- Bowmaker, J.K.; Dartnall, H.J. Visual pigments of rods and cones in a human retina. J. Physiol. 1980, 298, 501–511. [Google Scholar] [CrossRef]
- Hecht, S.; Shlaer, S.; Pirenne, M.H. Energy, quanta, and vision. J. Gen. Physiol. 1942, 25, 819–840. [Google Scholar] [CrossRef]
- Pepperberg, D.R. Rhodopsin and visual adaptation: Analysis of photoreceptor thresholds in the isolated skate retina. Vis. Res. 1984, 24, 357–366. [Google Scholar] [CrossRef]
- Bowmaker, J.K.; Dartnall, H.J.; Mollon, J.D. Microspectrophotometric demonstration of four classes of photoreceptor in an old world primate, Macaca fascicularis. J. Physiol. 1980, 298, 131–143. [Google Scholar] [CrossRef]
- Nathans, J.; Thomas, D.; Hogness, D.S. Molecular genetics of human color vision: The genes encoding blue, green, and red pigments. Science 1986, 232, 196–202. [Google Scholar] [CrossRef]
- Chow, R.L.; Altmann, C.R.; Lang, R.A.; Hemmati-Brivanlou, A. Pax6 induces ectopic eyes in a vertebrate. Development 1999, 126, 4213–4222. [Google Scholar] [CrossRef]
- Mathers, P.H.; Grinberg, A.; Mahon, K.A.; Jamrich, M. The Rx homeobox gene is essential for vertebrate eye development. Nature 1997, 387, 603–607. [Google Scholar] [CrossRef] [PubMed]
- O’Hara-Wright, M.; Gonzalez-Cordero, A. Retinal organoids: A window into human retinal development. Development 2020, 147, dev189746. [Google Scholar] [CrossRef] [PubMed]
- Hyer, J.; Kuhlman, J.; Afif, E.; Mikawa, T. Optic cup morphogenesis requires pre-lens ectoderm but not lens differentiation. Dev. Biol. 2003, 259, 351–363. [Google Scholar] [CrossRef]
- Nguyen, M.T.T.; Arnheiter, H. Signaling and transcriptional regulation in early mammalian eye development: A link between FGF and MITF. Development 2000, 127, 3581–3591. [Google Scholar] [CrossRef]
- Aparicio, J.G.; Hopp, H.; Choi, A.; Mandayam Comar, J.; Liao, V.C.; Harutyunyan, N.; Lee, T.C. Temporal expression of CD184(CXCR4) and CD171(L1CAM) identifies distinct early developmental stages of human retinal ganglion cells in embryonic stem cell derived retina. Exp. Eye Res. 2017, 154, 177–189. [Google Scholar] [CrossRef]
- Horsford, D.J.; Nguyen, M.T.T.; Sellar, G.C.; Kothary, R.; Arnheiter, H.; McInnes, R.R. Chx10 repression of Mitf is required for the maintenance of mammalian neuroretinal identity. Development 2005, 132, 177–187. [Google Scholar] [CrossRef]
- Martínez-Morales, J.R.; Rodrigo, I.; Bovolenta, P. Eye development: A view from the retina pigmented epithelium. Bioessays 2004, 26, 766–777. [Google Scholar] [CrossRef]
- Chauhan, B.K.; Disanza, A.; Choi, S.Y.; Faber, S.C.; Lou, M.; Beggs, H.E.; Scita, G.; Zheng, Y.; Lang, R.A. Cdc42- and IRSp53-dependent contractile filopodia tether presumptive lens and retina to coordinate epithelial invagination. Development 2009, 136, 3657–3667. [Google Scholar] [CrossRef]
- Schnichels, S.; Paquet-Durand, F.; Löscher, M.; Tsai, T.; Hurst, J.; Joachim, S.C.; Klettner, A. Retina in a dish: Cell cultures, retinal explants and animal models for common diseases of the retina. Prog. Retin. Eye Res. 2021, 81, 100880. [Google Scholar] [CrossRef]
- Ericsson, A.C.; Crim, M.J.; Franklin, C.L. A Brief History of Animal Modeling. Mo. Med. 2013, 110, 201. [Google Scholar]
- Verra, D.M.; Sajdak, B.S.; Merriman, D.K.; Hicks, D. Diurnal rodents as pertinent animal models of human retinal physiology and pathology. Prog. Retin. Eye Res. 2020, 74, 100776. [Google Scholar] [CrossRef] [PubMed]
- Alfonsetti, M.; Castelli, V.; D’angelo, M.; Benedetti, E.; Allegretti, M.; Barboni, B.; Cimini, A. Looking for In Vitro Models for Retinal Diseases. Int. J. Mol. Sci. 2021, 22, 10334. [Google Scholar] [CrossRef] [PubMed]
- Fan, N.; Lai, L. Genetically modified pig models for human diseases. J. Genet. Genom. 2013, 40, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.M.; McGill, T.J.; Puthussery, T.; Stoddard, J.; Renner, L.; Lewis, A.D.; Colgin, L.M.A.; Gayet, J.; Wang, X.; Prongay, K.; et al. Bardet-Biedl Syndrome in rhesus macaques: A nonhuman primate model of retinitis pigmentosa. Exp. Eye Res. 2019, 189, 107825. [Google Scholar] [CrossRef] [PubMed]
- Finger, R.P.; Fimmers, R.; Holz, F.G.; Scholl, H.P.N. Prevalence and causes of registered blindness in the largest federal state of Germany. Br. J. Ophthalmol. 2011, 95, 1061–1067. [Google Scholar] [CrossRef]
- Li, X.; Li, M.J.; Yang, Y.; Bai, Y. Effects of reprogramming on genomic imprinting and the application of pluripotent stem cells. Stem Cell Res. 2019, 41, 101655. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Knoblich, J.A. Generation of cerebral organoids from human pluripotent stem cells. Nat. Protoc. 2014, 9, 2329–2340. [Google Scholar] [CrossRef]
- Eiraku, M.; Watanabe, K.; Matsuo-Takasaki, M.; Kawada, M.; Yonemura, S.; Matsumura, M.; Wataya, T.; Nishiyama, A.; Muguruma, K.; Sasai, Y. Self-organized formation of polarized cortical tissues from ESCs and its active manipulation by extrinsic signals. Cell Stem Cell 2008, 3, 519–532. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Renner, M.; Martin, C.-A.; Wenzel, D.; Bicknell, L.S.; Hurles, M.E.; Homfray, T.; Penninger, J.M.; Jackson, A.P.; Knoblich, J.A. Cerebral organoids model human brain development and microcephaly. Nature 2013, 501, 373–379. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Corsini, N.S.; Wolfinger, S.; Gustafson, E.H.; Phillips, A.W.; Burkard, T.R.; Otani, T.; Livesey, F.J.; Knoblich, J.A. Guided self-organization and cortical plate formation in human brain organoids. Nat. Biotechnol. 2017, 35, 659–666. [Google Scholar] [CrossRef]
- Schwank, G.; Koo, B.K.; Sasselli, V.; Dekkers, J.F.; Heo, I.; Demircan, T.; Sasaki, N.; Boymans, S.; Cuppen, E.; Van Der Ent, C.K.; et al. Functional repair of CFTR by CRISPR/Cas9 in intestinal stem cell organoids of cystic fibrosis patients. Cell Stem Cell 2013, 13, 653–658. [Google Scholar] [CrossRef] [PubMed]
- Dekkers, J.F.; Wiegerinck, C.L.; De Jonge, H.R.; Bronsveld, I.; Janssens, H.M.; De Winter-De Groot, K.M.; Brandsma, A.M.; De Jong, N.W.M.; Bijvelds, M.J.C.; Scholte, B.J.; et al. A functional CFTR assay using primary cystic fibrosis intestinal organoids. Nat. Med. 2013, 19, 939–945. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.P.; Bear, C.E.; Chin, S.; Pasceri, P.; Thompson, T.O.; Huan, L.J.; Ratjen, F.; Ellis, J.; Rossant, J. Directed differentiation of human pluripotent stem cells into mature airway epithelia expressing functional CFTR protein. Nat. Biotechnol. 2012, 30, 876–882. [Google Scholar] [CrossRef] [PubMed]
- Low, J.H.; Li, P.; Chew, E.G.Y.; Zhou, B.; Suzuki, K.; Zhang, T.; Lian, M.M.; Liu, M.; Aizawa, E.; Rodriguez Esteban, C.; et al. Generation of Human PSC-Derived Kidney Organoids with Patterned Nephron Segments and a De Novo Vascular Network. Cell Stem Cell 2019, 25, 373–387.e9. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell 2006, 2, 663–676. [Google Scholar] [CrossRef]
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic stem cell lines derived from human blastocysts. Science 1998, 282, 1145–1147. [Google Scholar] [CrossRef]
- Osakada, F.; Ikeda, H.; Sasai, Y.; Takahashi, M. Stepwise differentiation of pluripotent stem cells into retinal cells. Nat. Protoc. 2009, 4, 811–824. [Google Scholar] [CrossRef]
- Osakada, F.; Ikeda, H.; Mandai, M.; Wataya, T.; Watanabe, K.; Yoshimura, N.; Akaike, A.; Sasai, Y.; Takahashi, M. Toward the generation of rod and cone photoreceptors from mouse, monkey and human embryonic stem cells. Nat. Biotechnol. 2008, 26, 215–224. [Google Scholar] [CrossRef]
- Lamba, D.A.; Karl, M.O.; Ware, C.B.; Reh, T.A. Efficient generation of retinal progenitor cells from human embryonic stem cells. Proc. Natl. Acad. Sci. USA 2006, 103, 12769–12774. [Google Scholar] [CrossRef]
- Meyer, J.S.; Shearer, R.L.; Capowski, E.E.; Wright, L.S.; Wallace, K.A.; McMillan, E.L.; Zhang, S.C.; Gamm, D.M. Modeling early retinal development with human embryonic and induced pluripotent stem cells. Proc. Natl. Acad. Sci. USA 2009, 106, 16698–16703. [Google Scholar] [CrossRef]
- Watanabe, K.; Kamiya, D.; Nishiyama, A.; Katayama, T.; Nozaki, S.; Kawasaki, H.; Watanabe, Y.; Mizuseki, K.; Sasai, Y. Directed differentiation of telencephalic precursors from embryonic stem cells. Nat. Neurosci. 2005, 8, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Eiraku, M.; Takata, N.; Ishibashi, H.; Kawada, M.; Sakakura, E.; Okuda, S.; Sekiguchi, K.; Adachi, T.; Sasai, Y. Self-organizing optic-cup morphogenesis in three-dimensional culture. Nature 2011, 472, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Ando, S.; Takata, N.; Kawada, M.; Muguruma, K.; Sekiguchi, K.; Saito, K.; Yonemura, S.; Eiraku, M.; Sasai, Y. Self-formation of optic cups and storable stratified neural retina from human ESCs. Cell Stem Cell 2012, 10, 771–785. [Google Scholar] [CrossRef]
- Zhong, X.; Gutierrez, C.; Xue, T.; Hampton, C.; Vergara, M.N.; Cao, L.H.; Peters, A.; Park, T.S.; Zambidis, E.T.; Meyer, J.S.; et al. Generation of three-dimensional retinal tissue with functional photoreceptors from human iPSCs. Nat. Commun. 2014, 5, 4047. [Google Scholar] [CrossRef]
- Wahlin, K.J.; Maruotti, J.A.; Sripathi, S.R.; Ball, J.; Angueyra, J.M.; Kim, C.; Grebe, R.; Li, W.; Jones, B.W.; Zack, D.J. Photoreceptor Outer Segment-like Structures in Long-Term 3D Retinas from Human Pluripotent Stem Cells. Sci. Rep. 2017, 7, 766. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xiong, K.; Lin, C.; Lv, L.; Chen, J.; Xu, C.; Wang, S.; Gu, D.; Zheng, H.; Yu, H.; et al. New medium used in the differentiation of human pluripotent stem cells to retinal cells is comparable to fetal human eye tissue. Biomaterials 2015, 53, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Flamier, A.; Abdouh, M.; Tétreault, N.; Barabino, A.; Wadhwa, S.; Bernier, G. Differentiation of human embryonic stem cells into cone photoreceptors through simultaneous inhibition of BMP, TGFβ and Wnt signaling. Development 2015, 142, 3294–3306. [Google Scholar] [CrossRef]
- Phillips, M.J.; Wallace, K.A.; Dickerson, S.J.; Miller, M.J.; Verhoeven, A.D.; Martin, J.M.; Wright, L.S.; Shen, W.; Capowski, E.E.; Percin, E.F.; et al. Blood-derived human iPS cells generate optic vesicle-like structures with the capacity to form retinal laminae and develop synapses. Investig. Ophthalmol. Vis. Sci. 2012, 53, 2007–2019. [Google Scholar] [CrossRef]
- Reichman, S.; Terray, A.; Slembrouck, A.; Nanteau, C.; Orieux, G.; Habeler, W.; Nandrot, E.F.; Sahel, J.A.; Monville, C.; Goureaua, O. From confluent human iPS cells to self-forming neural retina and retinal pigmented epithelium. Proc. Natl. Acad. Sci. USA 2014, 111, 8518–8523. [Google Scholar] [CrossRef]
- Singh, R.K.; Mallela, R.K.; Cornuet, P.K.; Reifler, A.N.; Chervenak, A.P.; West, M.D.; Wong, K.Y.; Nasonkin, I.O. Characterization of Three-Dimensional Retinal Tissue Derived from Human Embryonic Stem Cells in Adherent Monolayer Cultures. Stem Cells Dev. 2015, 24, 2778–2795. [Google Scholar] [CrossRef]
- Lowe, A.; Harris, R.; Bhansali, P.; Cvekl, A.; Liu, W. Intercellular Adhesion-Dependent Cell Survival and ROCK-Regulated Actomyosin-Driven Forces Mediate Self-Formation of a Retinal Organoid. Stem Cell Rep. 2016, 6, 743–756. [Google Scholar] [CrossRef] [PubMed]
- Völkner, M.; Zschätzsch, M.; Rostovskaya, M.; Overall, R.W.; Busskamp, V.; Anastassiadis, K.; Karl, M.O. Retinal Organoids from Pluripotent Stem Cells Efficiently Recapitulate Retinogenesis. Stem Cell Rep. 2016, 6, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Hunt, N.C.; Hallam, D.; Karimi, A.; Mellough, C.B.; Chen, J.; Steel, D.H.W.; Lako, M. 3D culture of human pluripotent stem cells in RGD-alginate hydrogel improves retinal tissue development. Acta Biomater. 2017, 49, 329–343. [Google Scholar] [CrossRef] [PubMed]
- Capowski, E.E.; Samimi, K.; Mayerl, S.J.; Phillips, M.J.; Pinilla, I.; Howden, S.E.; Saha, J.; Jansen, A.D.; Edwards, K.L.; Jager, L.D.; et al. Reproducibility and staging of 3D human retinal organoids across multiple pluripotent stem cell lines. Development 2019, 146. [Google Scholar] [CrossRef]
- Kaya, K.D.; Chen, H.Y.; Brooks, M.J.; Kelley, R.A.; Shimada, H.; Nagashima, K.; de Val, N.; Drinnan, C.T.; Gieser, L.; Kruczek, K.; et al. Transcriptome-based molecular staging of human stem cell-derived retinal organoids uncovers accelerated photoreceptor differentiation by 9-cis retinal. Mol. Vis. 2019, 25, 663. [Google Scholar]
- Zerti, D.; Molina, M.M.; Dorgau, B.; Mearns, S.; Bauer, R.; Al-Aama, J.; Lako, M. IGFBPs mediate IGF-1’s functions in retinal lamination and photoreceptor development during pluripotent stem cell differentiation to retinal organoids. Stem Cells 2021, 39, 458–466. [Google Scholar] [CrossRef]
- Gonzalez-Cordero, A.; Kruczek, K.; Naeem, A.; Fernando, M.; Kloc, M.; Ribeiro, J.; Goh, D.; Duran, Y.; Blackford, S.J.I.; Abelleira-Hervas, L.; et al. Recapitulation of Human Retinal Development from Human Pluripotent Stem Cells Generates Transplantable Populations of Cone Photoreceptors. Stem Cell Rep. 2017, 9, 820–837. [Google Scholar] [CrossRef]
- Ohgushi, M.; Matsumura, M.; Eiraku, M.; Murakami, K.; Aramaki, T.; Nishiyama, A.; Muguruma, K.; Nakano, T.; Suga, H.; Ueno, M.; et al. Molecular pathway and cell state responsible for dissociation-induced apoptosis in human pluripotent stem cells. Cell Stem Cell 2010, 7, 225–239. [Google Scholar] [CrossRef]
- Chen, G.; Hou, Z.; Gulbranson, D.R.; Thomson, J.A. Actin-myosin contractility is responsible for the reduced viability of dissociated human embryonic stem cells. Cell Stem Cell 2010, 7, 240–248. [Google Scholar] [CrossRef]
- Shirai, H.; Mandai, M.; Matsushita, K.; Kuwahara, A.; Yonemura, S.; Nakano, T.; Assawachananont, J.; Kimura, T.; Saito, K.; Terasaki, H.; et al. Transplantation of human embryonic stem cell-derived retinal tissue in two primate models of retinal degeneration. Proc. Natl. Acad. Sci. USA 2016, 113, E81–E90. [Google Scholar] [CrossRef]
- Compagnucci, C.; Barresi, S.; Petrini, S.; Billuart, P.; Piccini, G.; Chiurazzi, P.; Alfieri, P.; Bertini, E.; Zanni, G. Rho Kinase Inhibition Is Essential During In Vitro Neurogenesis and Promotes Phenotypic Rescue of Human Induced Pluripotent Stem Cell-Derived Neurons With Oligophrenin-1 Loss of Function. Stem Cells Transl. Med. 2016, 5, 860–869. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Haynes, T.; Gutierrez, C.; Aycinena, J.C.; Tsonis, P.A.; Del Rio-Tsonis, K. BMP signaling mediates stem/progenitor cell-induced retina regeneration. Proc. Natl. Acad. Sci. USA 2007, 104, 20380–20385. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Liu, Y.; Oltean, A.; Beebe, D.C. Bmp4 from the optic vesicle specifies murine retina formation. Dev. Biol. 2015, 402, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Kuwahara, A.; Ozone, C.; Nakano, T.; Saito, K.; Eiraku, M.; Sasai, Y. Generation of a ciliary margin-like stem cell niche from self-organizing human retinal tissue. Nat. Commun. 2015, 6, 6286. [Google Scholar] [CrossRef] [PubMed]
- Mellough, C.B.; Collin, J.; Khazim, M.; White, K.; Sernagor, E.; Steel, D.H.W.; Lako, M. IGF-1 Signaling Plays an Important Role in the Formation of Three-Dimensional Laminated Neural Retina and Other Ocular Structures from Human Embryonic Stem Cells. Stem Cells 2015, 33, 2416–2430. [Google Scholar] [CrossRef] [PubMed]
- Chichagova, V.; Hilgen, G.; Ghareeb, A.; Georgiou, M.; Carter, M.; Sernagor, E.; Lako, M.; Armstrong, L. Human iPSC differentiation to retinal organoids in response to IGF1 and BMP4 activation is line- and method-dependent. Stem Cells 2020, 38, 195–201. [Google Scholar] [CrossRef]
- Eldred, K.C.; Hadyniak, S.E.; Hussey, K.A.; Brenerman, B.; Zhang, P.W.; Chamling, X.; Sluch, V.M.; Welsbie, D.S.; Hattar, S.; Taylor, J.; et al. Thyroid hormone signaling specifies cone subtypes in human retinal organoids. Science 2018, 362. [Google Scholar] [CrossRef]
- Wagstaff, P.E.; ten Asbroek, A.L.M.A.; ten Brink, J.B.; Jansonius, N.M.; Bergen, A.A.B. An alternative approach to produce versatile retinal organoids with accelerated ganglion cell development. Sci. Rep. 2021, 11, 1101. [Google Scholar] [CrossRef]
- Cowan, C.S.; Renner, M.; De Gennaro, M.; Gross-Scherf, B.; Goldblum, D.; Hou, Y.; Munz, M.; Rodrigues, T.M.; Krol, J.; Szikra, T.; et al. Cell Types of the Human Retina and Its Organoids at Single-Cell Resolution. Cell 2020, 182, 1623–1640.e34. [Google Scholar] [CrossRef]
- Fligor, C.M.; Huang, K.C.; Lavekar, S.S.; VanderWall, K.B.; Meyer, J.S. Differentiation of retinal organoids from human pluripotent stem cells. In Methods in Cell Biology; Academic Press: Cambridge, MA, USA, 2020; Volume 159, pp. 279–302. ISBN 9780128215319. [Google Scholar]
- Corral-Serrano, J.C.; Lamers, I.J.C.; Van Reeuwijk, J.; Duijkers, L.; Hoogendoorn, A.D.M.; Yildirim, A.; Argyrou, N.; Ruigrok, R.A.A.; Letteboer, S.J.F.; Butcher, R.; et al. PCARE and WASF3 regulate ciliary F-actin assembly that is required for the initiation of photoreceptor outer segment disk formation. Proc. Natl. Acad. Sci. USA 2020, 117, 9922–9931. [Google Scholar] [CrossRef]
- Sinn, R.; Wittbrodt, J. An eye on eye development. Mech. Dev. 2013, 130, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Ang, L.C.; De Tilly, L.N.; Bidaisee, L.; Yücel, Y.H. Human glaucoma and neural degeneration in intracranial optic nerve, lateral geniculate nucleus, and visual cortex. Br. J. Ophthalmol. 2006, 90, 674–678. [Google Scholar] [CrossRef] [PubMed]
- Quadrato, G.; Nguyen, T.; Macosko, E.Z.; Sherwood, J.L.; Yang, S.M.; Berger, D.R.; Maria, N.; Scholvin, J.; Goldman, M.; Kinney, J.P.; et al. Cell diversity and network dynamics in photosensitive human brain organoids. Nature 2017, 545, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Fernando, M.; Lee, S.; Wark, J.R.; Xiao, D.; Kim, H.J.; Smith, G.C.; Wong, T.; Teber, E.T.; Ali, R.R.; Yang, P.; et al. Differentiation of cortical brain organoids and optic nerve-like structures from retinal confluent cultures of pluripotent stem cells. bioRxiv 2021, 7. [Google Scholar] [CrossRef]
- Gabriel, E.; Albanna, W.; Pasquini, G.; Ramani, A.; Josipovic, N.; Mariappan, A.; Schinzel, F.; Karch, C.M.; Bao, G.; Gottardo, M.; et al. Human brain organoids assemble functionally integrated bilateral optic vesicles. Cell Stem Cell 2021, 28, 1740–1757.e8. [Google Scholar] [CrossRef]
- Hallam, D.; Hilgen, G.; Dorgau, B.; Zhu, L.; Yu, M.; Bojic, S.; Hewitt, P.; Schmitt, M.; Uteng, M.; Kustermann, S.; et al. Human-Induced Pluripotent Stem Cells Generate Light Responsive Retinal Organoids with Variable and Nutrient-Dependent Efficiency. Stem Cells 2018, 36, 1535–1551. [Google Scholar] [CrossRef]
- Hiler, D.; Chen, X.; Hazen, J.; Kupriyanov, S.; Carroll, P.A.; Qu, C.; Xu, B.; Johnson, D.; Griffiths, L.; Frase, S.; et al. Quantification of Retinogenesis in 3D Cultures Reveals Epigenetic Memory and Higher Efficiency in iPSCs Derived from Rod Photoreceptors. Cell Stem Cell 2015, 17, 101–115. [Google Scholar] [CrossRef]
- Achberger, K.; Probst, C.; Haderspeck, J.C.; Bolz, S.; Rogal, J.; Chuchuy, J.; Nikolova, M.; Cora, V.; Antkowiak, L.; Haq, W.; et al. Merging organoid and organ-on-a-chip technology to generate complex multi-layer tissue models in a human retina-on-a-chip platform. Elife 2019, 8, e46188. [Google Scholar] [CrossRef]
- Miller, J.D.; Ganat, Y.M.; Kishinevsky, S.; Bowman, R.L.; Liu, B.; Tu, E.Y.; Mandal, P.K.; Vera, E.; Shim, J.W.; Kriks, S.; et al. Human iPSC-based modeling of late-onset disease via progerin-induced aging. Cell Stem Cell 2013, 13, 691–705. [Google Scholar] [CrossRef]
- Kim, H.; Ham, S.; Jo, M.; Lee, G.H.; Lee, Y.; Shin, J.; Lee, Y. CRISPR-Cas9 Mediated Telomere Removal Leads to Mitochondrial Stress and Protein Aggregation. Int. J. Mol. Sci. 2017, 18, 2093. [Google Scholar] [CrossRef]
- Maeda, A.; Mandai, M.; Takahashi, M. Gene and Induced Pluripotent Stem Cell Therapy for Retinal Diseases. Annu. Rev. Genom. Hum. Genet. 2019, 20, 201–216. [Google Scholar] [CrossRef] [PubMed]
- Maya-Vetencourt, J.F.; Manfredi, G.; Mete, M.; Colombo, E.; Bramini, M.; Di Marco, S.; Shmal, D.; Mantero, G.; Dipalo, M.; Rocchi, A.; et al. Subretinally injected semiconducting polymer nanoparticles rescue vision in a rat model of retinal dystrophy. Nat. Nanotechnol. 2020, 15, 698–708. [Google Scholar] [CrossRef]
- Miraldi Utz, V.; Coussa, R.G.; Antaki, F.; Traboulsi, E.I. Gene therapy for RPE65-related retinal disease. Ophthalmic Genet. 2018, 39, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Ayton, L.N.; Barnes, N.; Dagnelie, G.; Fujikado, T.; Goetz, G.; Hornig, R.; Jones, B.W.; Muqit, M.M.K.; Rathbun, D.L.; Stingl, K.; et al. An update on retinal prostheses. Clin. Neurophysiol. 2020, 131, 1383–1398. [Google Scholar] [CrossRef] [PubMed]
- Humayun, M.S.; Dorn, J.D.; Da Cruz, L.; Dagnelie, G.; Sahel, J.A.; Stanga, P.E.; Cideciyan, A.V.; Duncan, J.L.; Eliott, D.; Filley, E.; et al. Interim results from the international trial of Second Sight’s visual prosthesis. Ophthalmology 2012, 119, 779–788. [Google Scholar] [CrossRef]
- Stingl, K.; Schippert, R.; Bartz-Schmidt, K.U.; Besch, D.; Cottriall, C.L.; Edwards, T.L.; Gekeler, F.; Greppmaier, U.; Kiel, K.; Koitschev, A.; et al. Interim Results of a Multicenter Trial with the New Electronic Subretinal Implant Alpha AMS in 15 Patients Blind from Inherited Retinal Degenerations. Front. Neurosci. 2017, 11, 445. [Google Scholar] [CrossRef]
- Allen, P.J. Retinal prostheses: Where to from here? Clin. Experiment. Ophthalmol. 2021, 49, 418–429. [Google Scholar] [CrossRef]
- Hallum, L.E.; Dakin, S.C. Retinal Implantation of Electronic Vision Prostheses to Treat Retinitis Pigmentosa: A Systematic Review. Transl. Vis. Sci. Technol. 2021, 10, 8. [Google Scholar] [CrossRef]
- Fujikado, T.; Kamei, M.; Sakaguchi, H.; Kanda, H.; Endo, T.; Hirota, M.; Morimoto, T.; Nishida, K.; Kishima, H.; Terasawa, Y.; et al. One-Year Outcome of 49-Channel Suprachoroidal-Transretinal Stimulation Prosthesis in Patients with Advanced Retinitis Pigmentosa. Investig. Ophthalmol. Vis. Sci. 2016, 57, 6147–6157. [Google Scholar] [CrossRef]
- Zrenner, E.; Bartz-Schmidt, K.U.; Benav, H.; Besch, D.; Bruckmann, A.; Gabel, V.P.; Gekeler, F.; Greppmaier, U.; Harscher, A.; Kibbel, S.; et al. Subretinal electronic chips allow blind patients to read letters and combine them to words. Proc. Biol. Sci. 2011, 278, 1489–1497. [Google Scholar] [CrossRef]
- Lorach, H.; Goetz, G.; Smith, R.; Lei, X.; Mandel, Y.; Kamins, T.; Mathieson, K.; Huie, P.; Harris, J.; Sher, A.; et al. Photovoltaic restoration of sight with high visual acuity. Nat. Med. 2015, 21, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.C.; Chang, H.M.; Hsu, C.C.; Hung, K.H.; Chen, Y.T.; Chen, S.Y.; Chen, S.J. Retinal prostheses in degenerative retinal diseases. J. Chin. Med. Assoc. 2015, 78, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Da Cruz, L.; Coley, B.F.; Dorn, J.; Merlini, F.; Filley, E.; Christopher, P.; Chen, F.K.; Wuyyuru, V.; Sahel, J.; Stanga, P.; et al. The Argus II epiretinal prosthesis system allows letter and word reading and long-term function in patients with profound vision loss. Br. J. Ophthalmol. 2013, 97, 632–636. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, A.K.; Behrend, M.R.; Kuroda, M.; Humayun, M.S.; Weiland, J.D. An in vitro model of a retinal prosthesis. IEEE Trans. Biomed. Eng. 2008, 55, 1744–1753. [Google Scholar] [CrossRef] [PubMed]
- Stiles, N.R.B.; Patel, V.R.; Weiland, J.D. Multisensory perception in Argus II retinal prosthesis patients: Leveraging auditory-visual mappings to enhance prosthesis outcomes. Vis. Res. 2021, 182, 58–68. [Google Scholar] [CrossRef]
- Corna, A.; Herrmann, T.; Zeck, G. Electrode-size dependent thresholds in subretinal neuroprosthetic stimulation. J. Neural Eng. 2018, 15, 045003. [Google Scholar] [CrossRef]
- Abbott, C.J.; Nayagam, D.A.X.; Luu, C.D.; Epp, S.B.; Williams, R.A.; Salinas-LaRosa, C.M.; Villalobos, J.; McGowan, C.; Shivdasani, M.N.; Burns, O.; et al. Safety Studies for a 44-Channel Suprachoroidal Retinal Prosthesis: A Chronic Passive Study. Investig. Ophthalmol. Vis. Sci. 2018, 59, 1410–1424. [Google Scholar] [CrossRef]
- Aurelia, N.; Chenais, L.; Jole, M.; Leccardi, I.A.; Ghezzi, D. Photovoltaic retinal prosthesis restores high-resolution responses to single-pixel stimulation in blind retinas. Commun. Mater. 2021, 2, 1–16. [Google Scholar] [CrossRef]
- Nguyen, D.; Valet, M.; Dégardin, J.; Boucherit, L.; Illa, X.; de la Cruz, J.; del Corro, E.; Bousquet, J.; Garrido, J.A.; Hébert, C.; et al. Novel Graphene Electrode for Retinal Implants: An in vivo Biocompatibility Study. Front. Neurosci. 2021, 15. [Google Scholar] [CrossRef]
- Huang, T.W.; Kamins, T.I.; Chen, Z.C.; Wang, B.Y.; Bhuckory, M.; Galambos, L.; Ho, E.; Ling, T.; Afshar, S.; Shin, A.; et al. Vertical-junction photodiodes for smaller pixels in retinal prostheses. J. Neural Eng. 2021, 18, 036015. [Google Scholar] [CrossRef]
- Palanker, D.; Le Mer, Y.; Mohand-Said, S.; Sahel, J.A. Simultaneous perception of prosthetic and natural vision in AMD patients. Nat. Commun. 2022, 13, 513. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Lorach, H.; Huie, P.; Palanker, D. Implantation of Modular Photovoltaic Subretinal Prosthesis. Ophthalmic Surg. Lasers Imaging Retin. 2016, 47, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Cha, K.; Horch, K.W.; Normann, R.A. Mobility performance with a pixelized vision system. Vis. Res. 1992, 32, 1367–1372. [Google Scholar] [CrossRef]
- Faber, H.; Besch, D.; Bartz-Schmidt, K.U.; Eisenstein, H.; Roider, J.; Sachs, H.; Gekeler, F.; Zrenner, E.; Stingl, K. Restriction of eye motility in patients with RETINA IMPLANT Alpha AMS. Acta Ophthalmol. 2020, 98, e998–e1003. [Google Scholar] [CrossRef]
- Bloch, E.; da Cruz, L. The Argus II Retinal Prosthesis System. In Prosthesis; Intech Open: London, UK, 2019. [Google Scholar] [CrossRef]
- Palanker, D.; Le Mer, Y.; Mohand-Said, S.; Muqit, M.; Sahel, J.A. Photovoltaic Restoration of Central Vision in Atrophic Age-Related Macular Degeneration. Ophthalmology 2020, 127, 1097–1104. [Google Scholar] [CrossRef]
- Jole, M.; Leccardi, I.A.; Aurelia, N.; Chenais, L.; Ferlauto, L.; Kawecki, M.; Geneviève Zollinger, E.; Ghezzi, D. Photovoltaic organic interface for neuronal stimulation in the near-infrared. Commun. Mater. 2020, 1, 1–13. [Google Scholar] [CrossRef]
- Medagoda, D.I.; Ghezzi, D. Organic semiconductors for light-mediated neuromodulation. Commun. Mater. 2021, 2, 111. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, Q.; Wang, H.; Wang, M. Multiscale engineering of functional organic polymer interfaces for neuronal stimulation and recording. Mater. Chem. Front. 2020, 4, 3444–3471. [Google Scholar] [CrossRef]
- Antognazza, M.R.; Di Paolo, M.; Ghezzi, D.; Mete, M.; Di Marco, S.; Maya-Vetencourt, J.F.; Maccarone, R.; Desii, A.; Di Fonzo, F.; Bramini, M.; et al. Characterization of a Polymer-Based, Fully Organic Prosthesis for Implantation into the Subretinal Space of the Rat. Adv. Healthc. Mater. 2016, 5, 2271–2282. [Google Scholar] [CrossRef]
- Hong, J.-W.; Yoon, C.; Jo, K.; Won, J.H.; Park, S. Recent advances in recording and modulation technologies for next-generation neural interfaces. iScience 2021, 24, 103550. [Google Scholar] [CrossRef]
- Maya-Vetencourt, J.F.; Ghezzi, D.; Antognazza, M.R.; Colombo, E.; Mete, M.; Feyen, P.; Desii, A.; Buschiazzo, A.; Di Paolo, M.; Di Marco, S.; et al. A fully organic retinal prosthesis restores vision in a rat model of degenerative blindness. Nat. Mater. 2017, 16, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Ghezzi, D.; Antognazza, M.R.; Dal Maschio, M.; Lanzarini, E.; Benfenati, F.; Lanzani, G. A hybrid bioorganic interface for neuronal photoactivation. Nat. Commun. 2011, 2, 166. [Google Scholar] [CrossRef] [PubMed]
- Ferlauto, L.; Airaghi Leccardi, M.J.I.; Chenais, N.A.L.; Gilliéron, S.C.A.; Vagni, P.; Bevilacqua, M.; Wolfensberger, T.J.; Sivula, K.; Ghezzi, D. Design and validation of a foldable and photovoltaic wide-field epiretinal prosthesis. Nat. Commun. 2018, 9, 1–15. [Google Scholar] [CrossRef]
- Irimia-Vladu, M.; Kanbur, Y.; Camaioni, F.; Coppola, M.E.; Yumusak, C.; Irimia, C.V.; Vlad, A.; Operamolla, A.; Farinola, G.M.; Suranna, G.P.; et al. Stability of Selected Hydrogen Bonded Semiconductors in Organic Electronic Devices. Chem. Mater. 2019, 31, 6315–6346. [Google Scholar] [CrossRef]
- Sytnyk, M.; Jakešová, M.; Litviňuková, M.; Mashkov, O.; Kriegner, D.; Stangl, J.; Nebesářová, J.; Fecher, F.W.; Schöfberger, W.; Sariciftci, N.S.; et al. Cellular interfaces with hydrogen-bonded organic semiconductor hierarchical nanocrystals. Nat. Commun. 2017, 8, 91. [Google Scholar] [CrossRef]
- Sytnyk, M.; Głowacki, E.D.; Yakunin, S.; Voss, G.; Schöfberger, W.; Kriegner, D.; Stangl, J.; Trotta, R.; Gollner, C.; Tollabimazraehno, S.; et al. Hydrogen-bonded organic semiconductor micro- and nanocrystals: From colloidal syntheses to (opto-)electronic devices. J. Am. Chem. Soc. 2014, 136, 16522–16532. [Google Scholar] [CrossRef]
- Gregory, P. Industrial applications of phthalocyanines. J. Porphyr. Phthalocyanines 2000, 4, 432–437. [Google Scholar] [CrossRef]
- Gautam, V.; Rand, D.; Hanein, Y.; Narayan, K.S. A Polymer Optoelectronic Interface Provides Visual Cues to a Blind Retina. Adv. Mater. 2014, 26, 1751–1756. [Google Scholar] [CrossRef]
- Antognazza, M.R.; Ghezzi, D.; Musitelli, D.; Garbugli, M.; Lanzani, G. A hybrid solid-liquid polymer photodiode for the bioenvironment. Appl. Phys. Lett. 2009, 94, 243501. [Google Scholar] [CrossRef]
- Simone, G.; Di Carlo Rasi, D.; de Vries, X.; Heintges, G.H.; Meskers, S.C.; Janssen, R.A.; Gelinck, G.H. Near-Infrared Tandem Organic Photodiodes for Future Application in Artificial Retinal Implants. Adv. Mater. 2018, 30, 1804678. [Google Scholar] [CrossRef]
- Stern, J.H.; Temple, S. Stem cells for retinal replacement therapy. Neurotherapeutics 2011, 8, 736–743. [Google Scholar] [CrossRef] [PubMed]
- Cooke, J.A.; Meyer, J.S. Human Pluripotent Stem Cell-Derived Retinal Ganglion Cells: Applications for the Study and Treatment of Optic Neuropathies. Curr. Ophthalmol. Rep. 2015, 3, 200–206. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Garcia, J.M.; Mendonça, L.; Brant, R.; Abud, M.; Regatieri, C.; Diniz, B. Stem cell therapy for retinal diseases. World J. Stem Cells 2015, 7, 160. [Google Scholar] [CrossRef] [PubMed]
- Jayakody, S.A.; Gonzalez-Cordero, A.; Ali, R.R.; Pearson, R.A. Cellular strategies for retinal repair by photoreceptor replacement. Prog. Retin. Eye Res. 2015, 46, 31–66. [Google Scholar] [CrossRef] [PubMed]
- Seiler, M.J.; Aramant, R.B. Cell replacement and visual restoration by retinal sheet transplants. Prog. Retin. Eye Res. 2012, 31, 661–687. [Google Scholar] [CrossRef]
- Pearson, R.A.; Hippert, C.; Graca, A.B.; Barber, A.C. Photoreceptor replacement therapy: Challenges presented by the diseased recipient retinal environment. Vis. Neurosci. 2014, 31, 333–344. [Google Scholar] [CrossRef]
- Jones, B.W.; Marc, R.E. Retinal remodeling during retinal degeneration. Exp. Eye Res. 2005, 81, 123–137. [Google Scholar] [CrossRef]
- Madrakhimov, S.B.; Yang, J.Y.; Ahn, D.H.; Han, J.W.; Ha, T.H.; Park, T.K. Peripapillary Intravitreal Injection Improves AAV-Mediated Retinal Transduction. Mol. Ther. Methods Clin. Dev. 2020, 17, 647–656. [Google Scholar] [CrossRef]
- Vázquez-Domínguez, I.; Garanto, A.; Collin, R.W.J. Molecular Therapies for Inherited Retinal Diseases-Current Standing, Opportunities and Challenges. Genes 2019, 10, 654. [Google Scholar] [CrossRef]
- Collin, R.W.J.; Garanto, A. Applications of antisense oligonucleotides for the treatment of inherited retinal diseases. Curr. Opin. Ophthalmol. 2017, 28, 260–266. [Google Scholar] [CrossRef]
- Smith, C.; Abalde-Atristain, L.; He, C.; Brodsky, B.R.; Braunstein, E.M.; Chaudhari, P.; Jang, Y.Y.; Cheng, L.; Ye, Z. Efficient and allele-specific genome editing of disease loci in human iPSCs. Mol. Ther. 2015, 23, 570–577. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; González, F.; Huangfu, D. The iCRISPR platform for rapid genome editing in human pluripotent stem cells. Methods Enzymol. 2014, 546, 215–250. [Google Scholar] [CrossRef] [PubMed]
- Pearson, R.A.; Barber, A.C.; Rizzi, M.; Hippert, C.; Xue, T.; West, E.L.; Duran, Y.; Smith, A.J.; Chuang, J.Z.; Azam, S.A.; et al. Restoration of vision after transplantation of photoreceptors. Nature 2012, 485, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Cameron, D.E.; Bashor, C.J.; Collins, J.J. A brief history of synthetic biology. Nat. Rev. Microbiol. 2014, 12, 381–390. [Google Scholar] [CrossRef]
- Konig, H.; Frank, D.; Heil, R.; Coenen, C. Synthetic genomics and synthetic biology applications between hopes and concerns. Curr. Genom. 2013, 14, 11–24. [Google Scholar] [CrossRef][Green Version]
- Sridhar, A.; Hoshino, A.; Finkbeiner, C.R.; Chitsazan, A.; Dai, L.; Haugan, A.K.; Eschenbacher, K.M.; Jackson, D.L.; Trapnell, C.; Bermingham-McDonogh, O.; et al. Single-Cell Transcriptomic Comparison of Human Fetal Retina, hPSC-Derived Retinal Organoids, and Long-Term Retinal Cultures. Cell Rep. 2020, 30, 1644–1659.e4. [Google Scholar] [CrossRef]
- Lane, A.; Jovanovic, K.; Shortall, C.; Ottaviani, D.; Panes, A.B.; Schwarz, N.; Guarascio, R.; Hayes, M.J.; Palfi, A.; Chadderton, N.; et al. Modeling and Rescue of RP2 Retinitis Pigmentosa Using iPSC-Derived Retinal Organoids. Stem Cell Rep. 2020, 15, 67–79. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, W.; Jin, Z.B. Retinal organoids as models for development and diseases. Cell Regen. 2021, 10, 33. [Google Scholar] [CrossRef]


| Disease | Suitable Species | Rodent Models |
|---|---|---|
| Diabetic Retinopathy | Mice | Ins2Akita nonobese diabetic (NOD) ob/ob or Db/Db (Leprdb) Kimba; Akimba |
| Age-related Macular Degeneration | Mice, Rats, Rabbits; Pigs; Non-human primates | Complements H, C3a and C5a KO C3 overexpression Chemokines Ccl2/Cx3cr1 double KO Nrf-2 or SOD1 KO ApoE4 KI |
| Glaucoma | Mice | TDBA/2J |
| Retinitis Pigmentosa | Mice; Rats; Rabbits; Pigs; Zebrafish; Non-human primates | rd1, rd4, rd8, rd10 VPP (V20G, P27L, P23H mutations) RCS rat model |
| Study | Cell Source | Culture Initiation | Culture Diff. | Prot. Length | Tissues Produced | Notes |
|---|---|---|---|---|---|---|
| Lamba et al., 2006 [62] | hiPSCs | Matrigel-coated dishes. noggin, DKK1, IGF1 for 3 w. | Cells cultured in N2/B27 medium | 60 d | NR by d2; OC by d25; Rods PRs and OF by 6 w | First 2D retinal cells from ESCs |
| Nakano et al., 2012 [66] | hESCs | Matrigel KSR medium + IWR-1e, ROCKi for 12 d. | FBS, SAG for 6 d, DMEM/F12+N2 medium Chir99021 d15 to d18 NR isolated on d18 in suspension culture. | 126 d | Bi-layered OC of NR and RPE; PRs d126 | 3D method improvement |
| Phillips et al., 2012 [71] | Blood-derived hiPSCs | Cell aggregates in KSR for 4 d. N2 + heparin for 2 d. | Aggregates on laminin 10 d. d16, neural clusters in B27 medium. d20, OVs maintained in adherent culture. | 50 d | OVs by d20; NR or RPE d40; NR rosettes d50 | |
| Zhong et al., 2014 [67] | hiPSCs | Cell aggregates in mTeSR1 medium with blebbistatin. Medium gradually transitioned into N2 + heparin. | d16, B27 medium. w4, aggregates detached. d42, medium with FBS, taurine. Addition of RA for PR maturation. | 21 w | NR; 3D retinal cups on d21–28; rhodopsin+ PRs by w21. | First 3D/2D method to describe mature and light-responding PRs |
| Reichman et al., 2014 [72] | hiPSCs | Confluent culture without FGF2 for 2 d, Medium transitioned into N2. | d14, neural clusters floating in N2 + FGF2; pigmented patches isolated on gelatin. FGF2 removed at d21. | 30 d | Rapid diff. of NR and RPE; NR rosettes d42 | First 2D/3D method; NRV excision |
| Zhou et al., 2015 [70] | hESCs hiPSCs | EBs cultured in KSR + B27, noggin, DKK1, IGF1 for 3 d. | Adherent culture in N2/B27 + noggin, DKK1, IGF1, COCO, FGF2 4 w. | 5 w | Cones PR d35; polarized cone PRs + CC + OS d60 | |
| Singh et al., 2015 [73] | hESCs (H9) | Dense colonies in mTeSR1 + FGF2. Medium changed to FGF2- free Neurobasal medium + noggin. d3, N2/B27 added. | 2 w + FGF2, 4 w DKK1+ IGF1 for 1 w. Neurobasal medium + noggin, FGF2, FGF9 for 12 w. | 12 w | Four retina layers: RPE, early PRs, INL and RGCs | |
| Lowe et al., 2016 [74] | hESCs hiPSCs | Cell gelling for 30 min Floating clusters in N2/B27 medium 5 d. | d12–17, detach adherent cultures; floating aggregates in B27; 2 w + FBS + taurine | 25–30 d | NR, ciliary margin, and RPE. | Spontaneous formationof NR |
| Völkner et al., 2016 [75] | hESCs | ROs in KSR + ROCKi + IWR1e 12 d, + Matrigel + FBS + SAG | ROs cut into 5 parts in N2 + FBS + EC23 41 d. | 41 d | cone or rod PRs. | |
| Hunt et al., 2017 [76] | hESCs hiPSCs | EBs in mTeSR1 + ROCKi. | d3, KSR + IGF1 + B27; d5–9, + FBS d12, EBs encapsulated in hydrogel 45 d. | 45 d | NR and RPE | |
| Capowski et al., 2019 [77] | hESCs hiPSCs | hPSCs in mTeSR1 + Matrigel. EBs lifted and weaned in N2 + Heparin 4 d. | d6, + BMP4, d16, B27 + FBS + taurine + RA; d100. RA removed | 175 d | Highly developed ONL, OPL, INL | staging system of ROs. BMP4 increases NRV |
| Kaya et al., 2019 [78] | hESCs hiPSCs | Cells in E8 + Matrigel. EBs lifted and weaned in N2 + Heparin 16 d. | d16, B27 + FBS; d42, + taurine; d63, + 9-cis retinal; d92, half conc. + N2. | 200 d | NR and cone, rod PRs | 9-cis retinal enhance rod PRs. |
| Zerti et al., 2021 [79] | hESCs | Cells in mTeSR1 + Matrigel + ROCKi; KOSR + B27 + IGF-1 18 d. | d18, +RA + IGF-1 + T3 + Taurine; d37, N2/B27 + IGF-1 | 90 d | NR, RPE, lensand cornea; PRs by d90 | IGF-1 increases the formation of laminated NRVs. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellapianta, A.; Cetkovic, A.; Bolz, M.; Salti, A. Retinal Organoids and Retinal Prostheses: An Overview. Int. J. Mol. Sci. 2022, 23, 2922. https://doi.org/10.3390/ijms23062922
Bellapianta A, Cetkovic A, Bolz M, Salti A. Retinal Organoids and Retinal Prostheses: An Overview. International Journal of Molecular Sciences. 2022; 23(6):2922. https://doi.org/10.3390/ijms23062922
Chicago/Turabian StyleBellapianta, Alessandro, Ana Cetkovic, Matthias Bolz, and Ahmad Salti. 2022. "Retinal Organoids and Retinal Prostheses: An Overview" International Journal of Molecular Sciences 23, no. 6: 2922. https://doi.org/10.3390/ijms23062922
APA StyleBellapianta, A., Cetkovic, A., Bolz, M., & Salti, A. (2022). Retinal Organoids and Retinal Prostheses: An Overview. International Journal of Molecular Sciences, 23(6), 2922. https://doi.org/10.3390/ijms23062922