
Functional Characterization of Allatostatin C (PISCF/AST) and Juvenile Hormone Acid O-Methyltransferase in Dendroctonus armandi



Abstract
:1. Introduction
2. Results
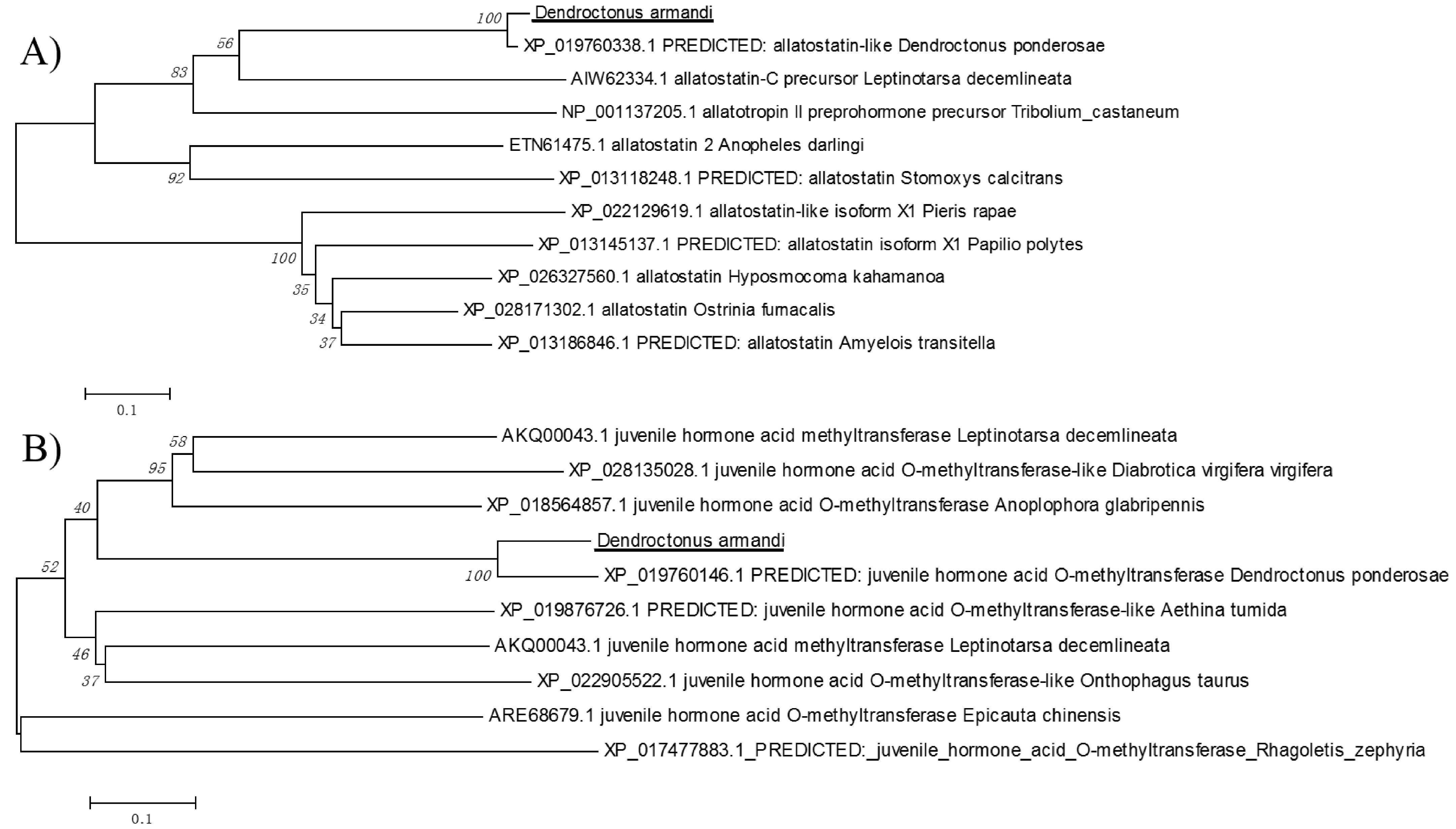
2.1. Identification of DaAST and DaJHAMT Genes
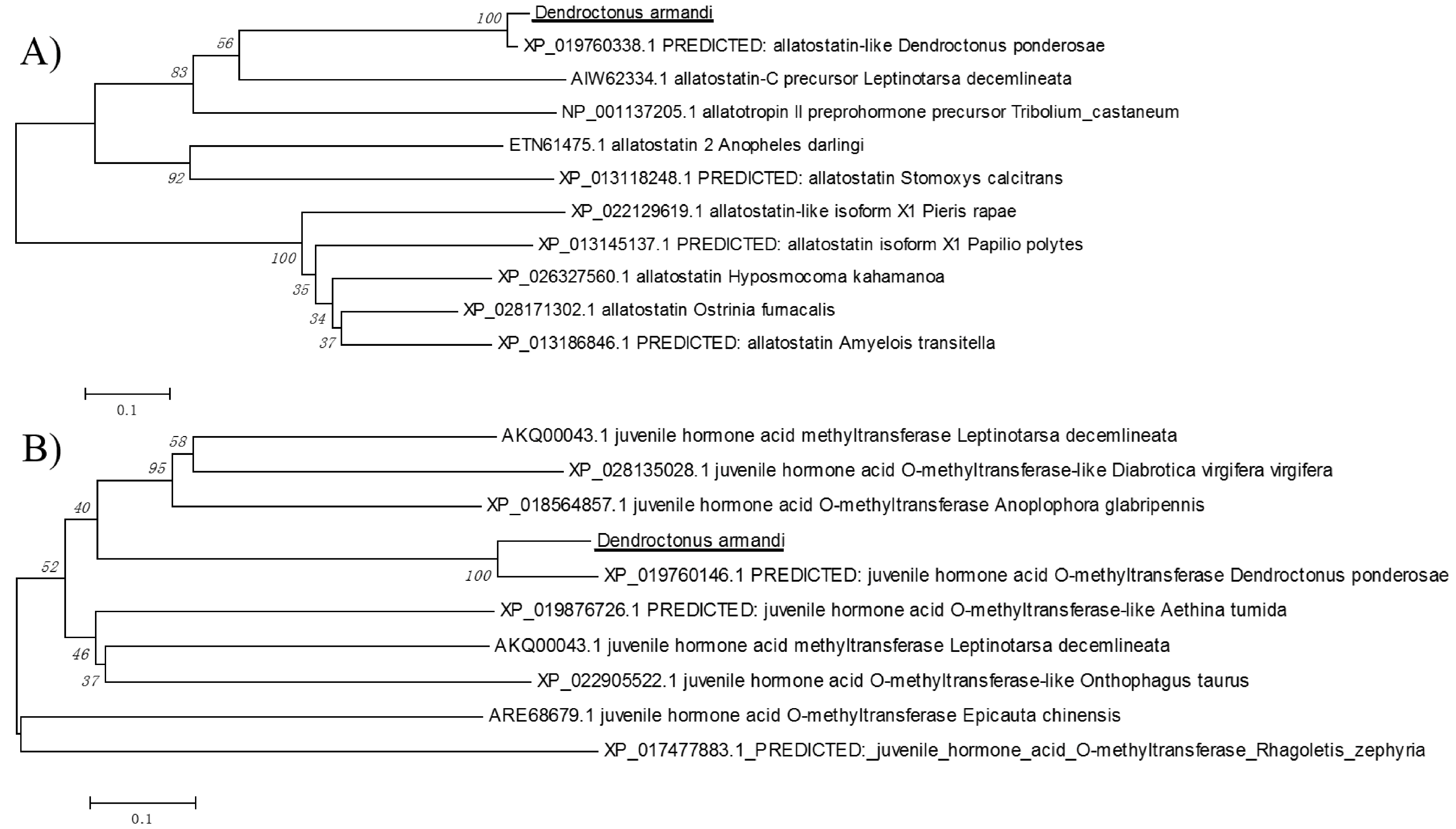
2.2. Physicochemical Properties and Bioinformatic Analysis
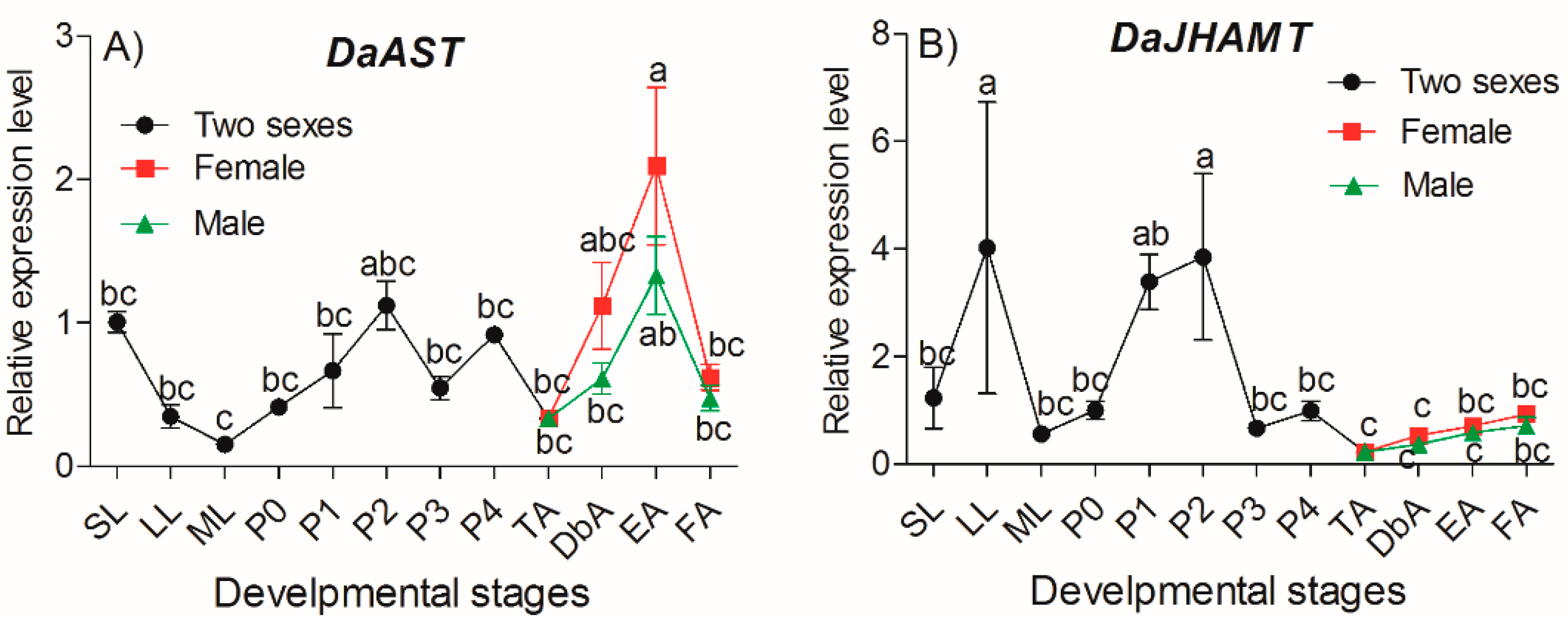
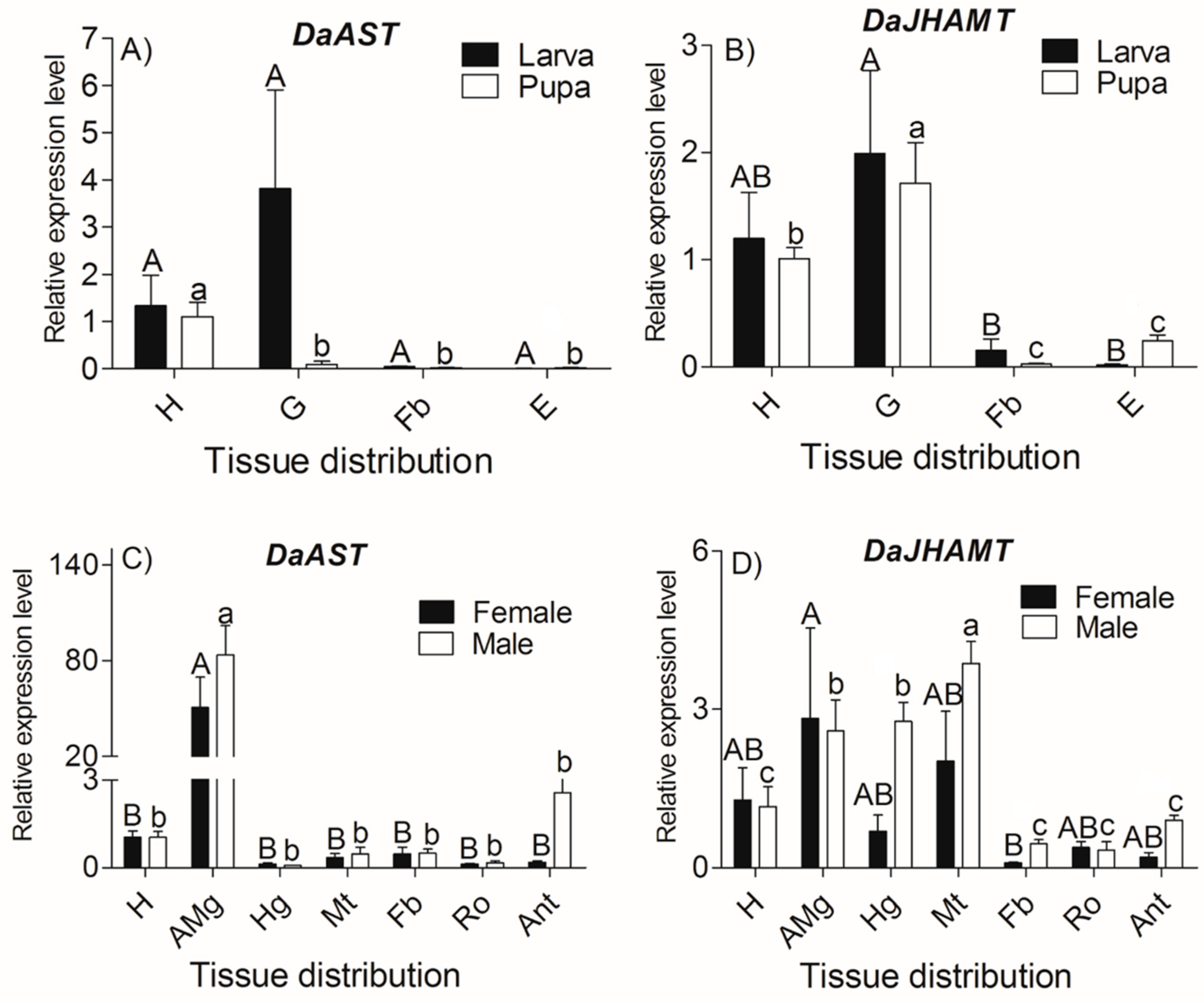
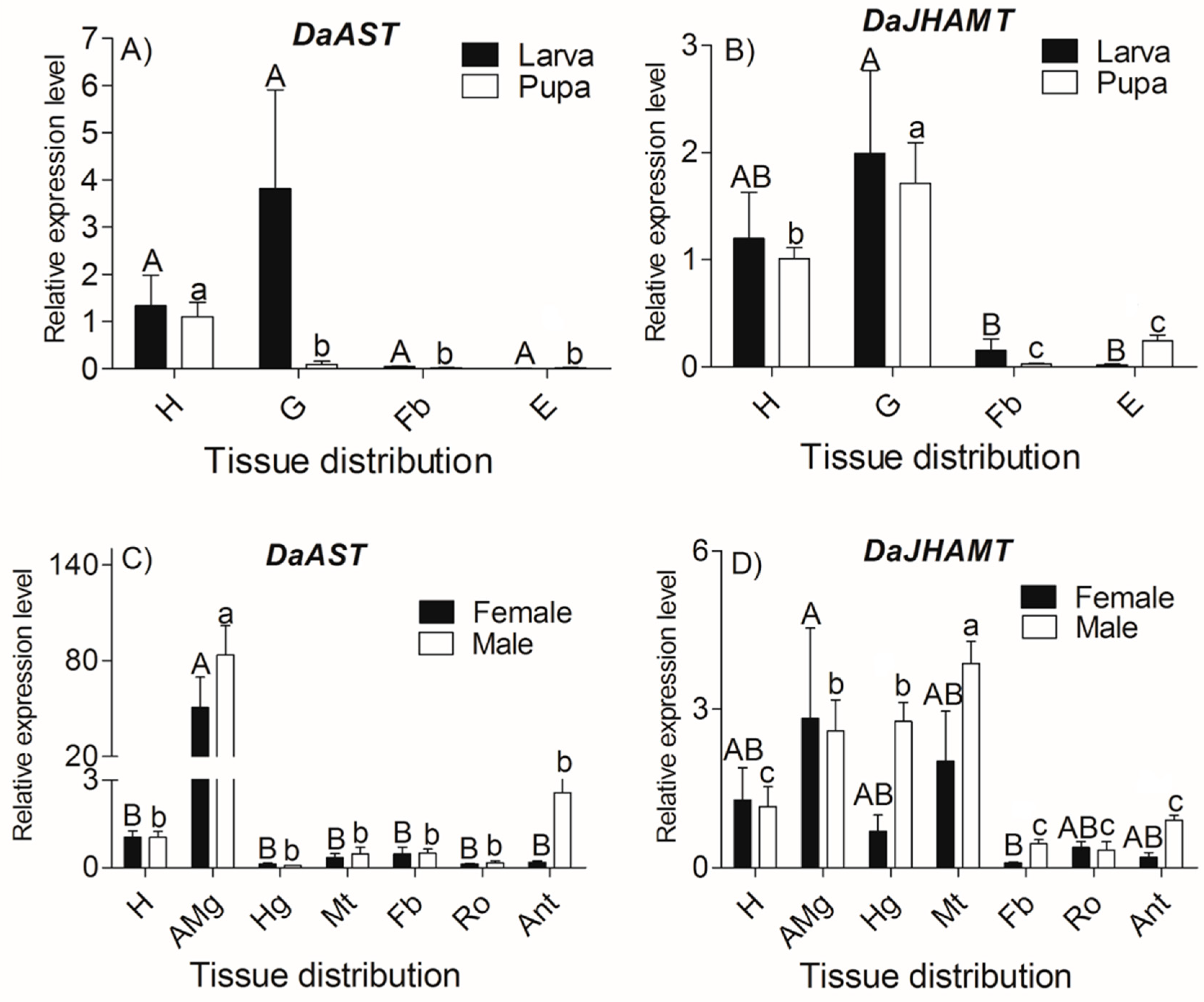
2.3. DaAST and DaJHAMT Transcript Levels in D. armandi at Different Life Stages and in Different Tissues
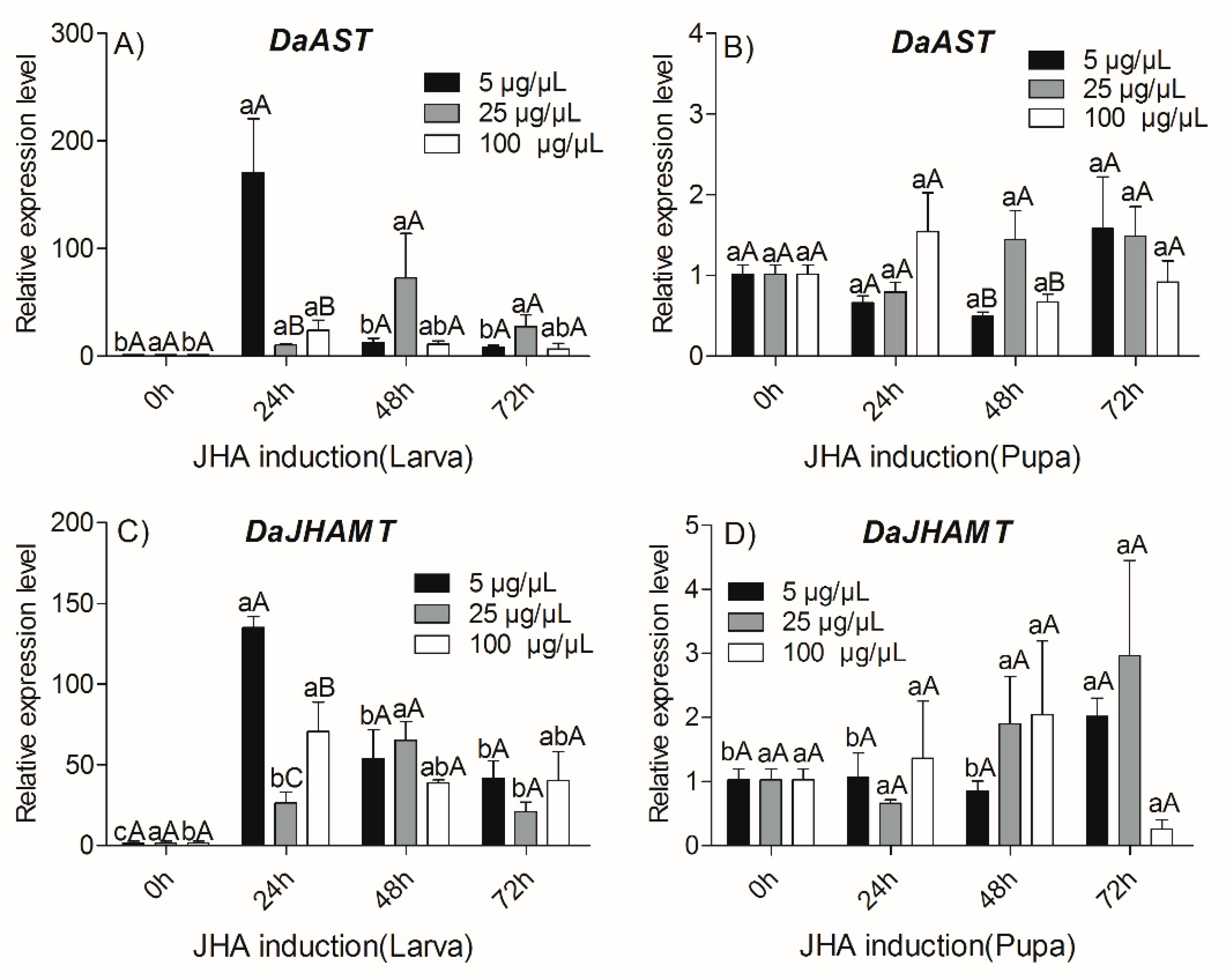
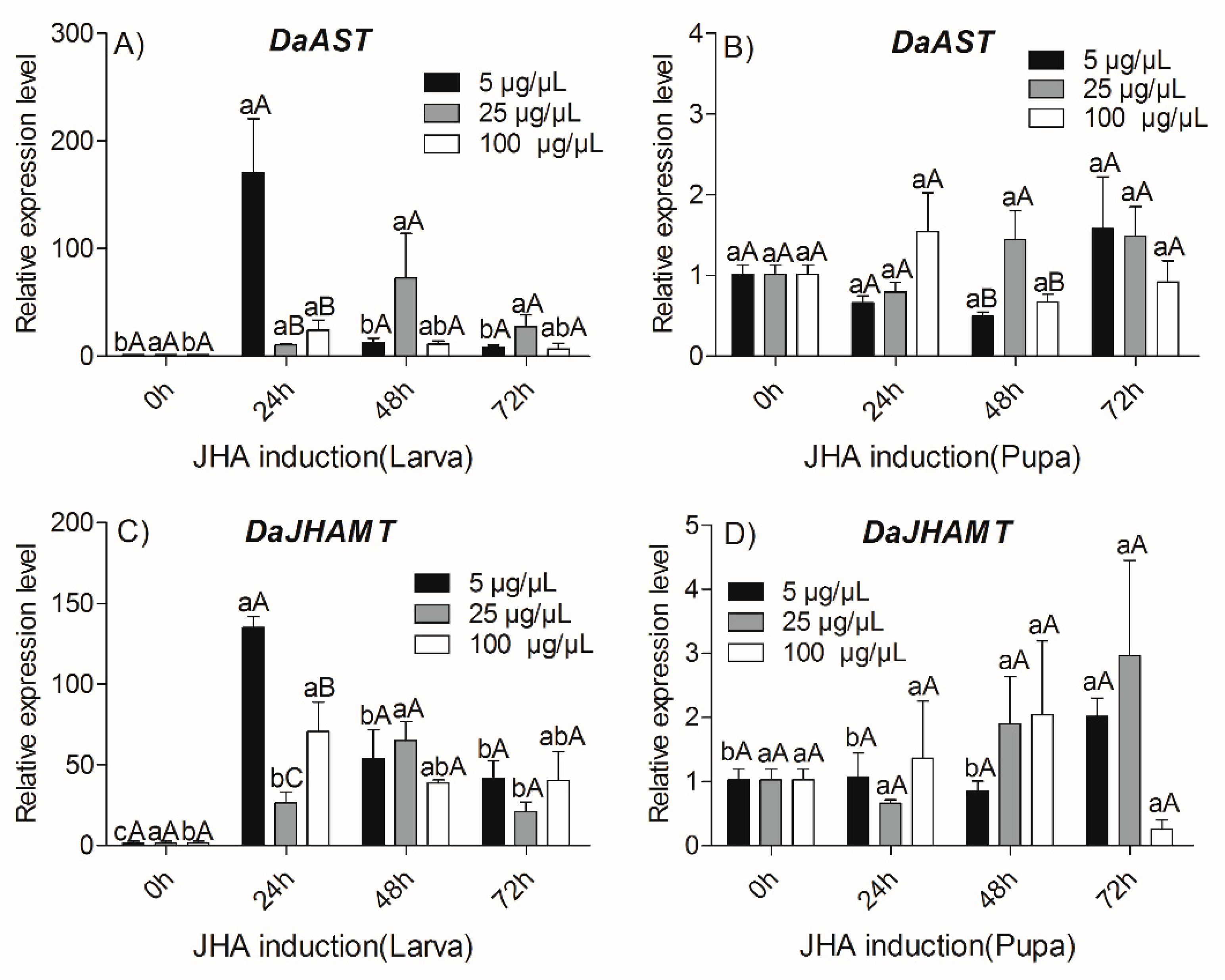
2.4. Effects of JHA Injection on Transcript Levels of DaAST and DaJHAMT
2.5. RNAi Effect of DaAST and DaJHAMT
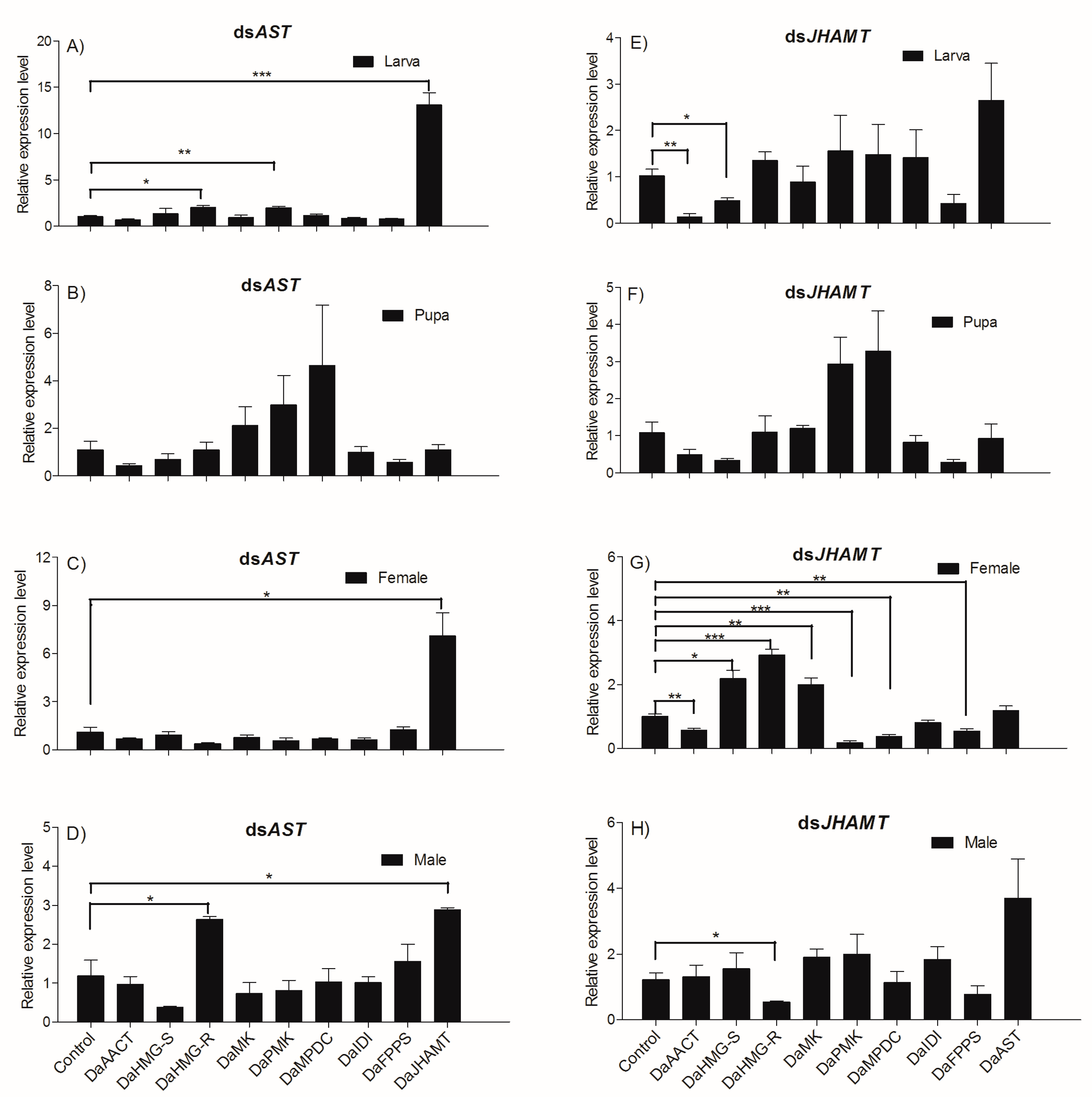
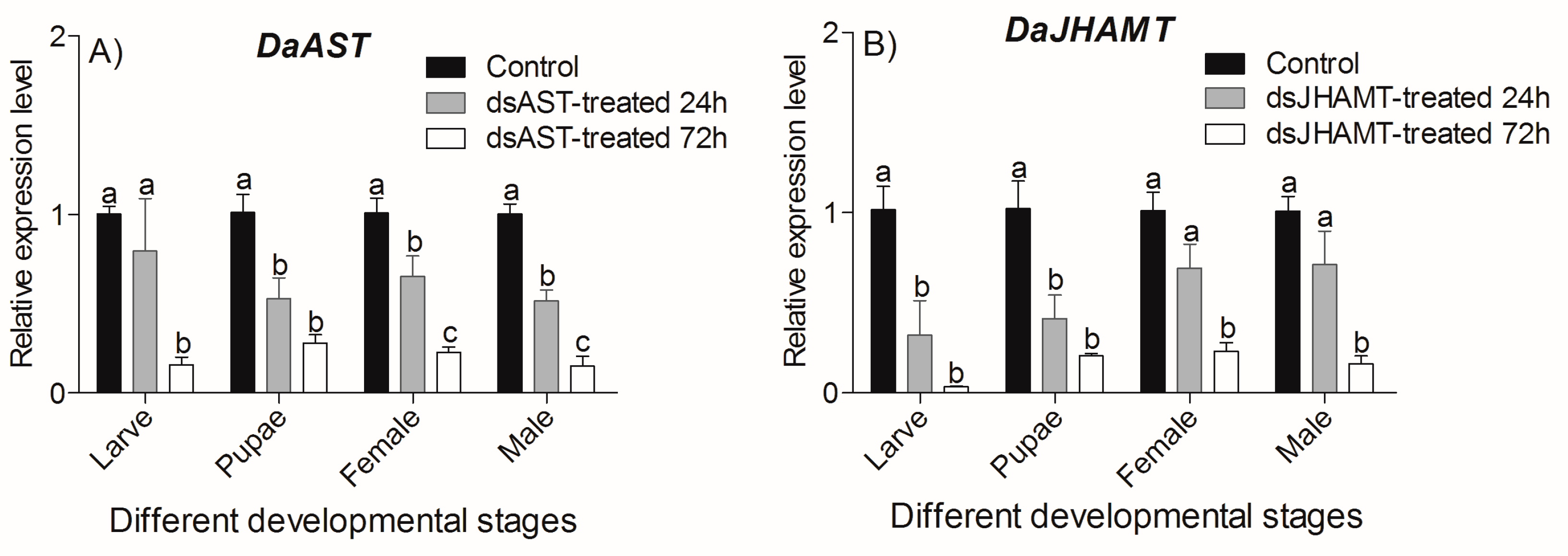
2.5.1. Determination of DaAST and DaJHAMT Silencing by qRT-PCR
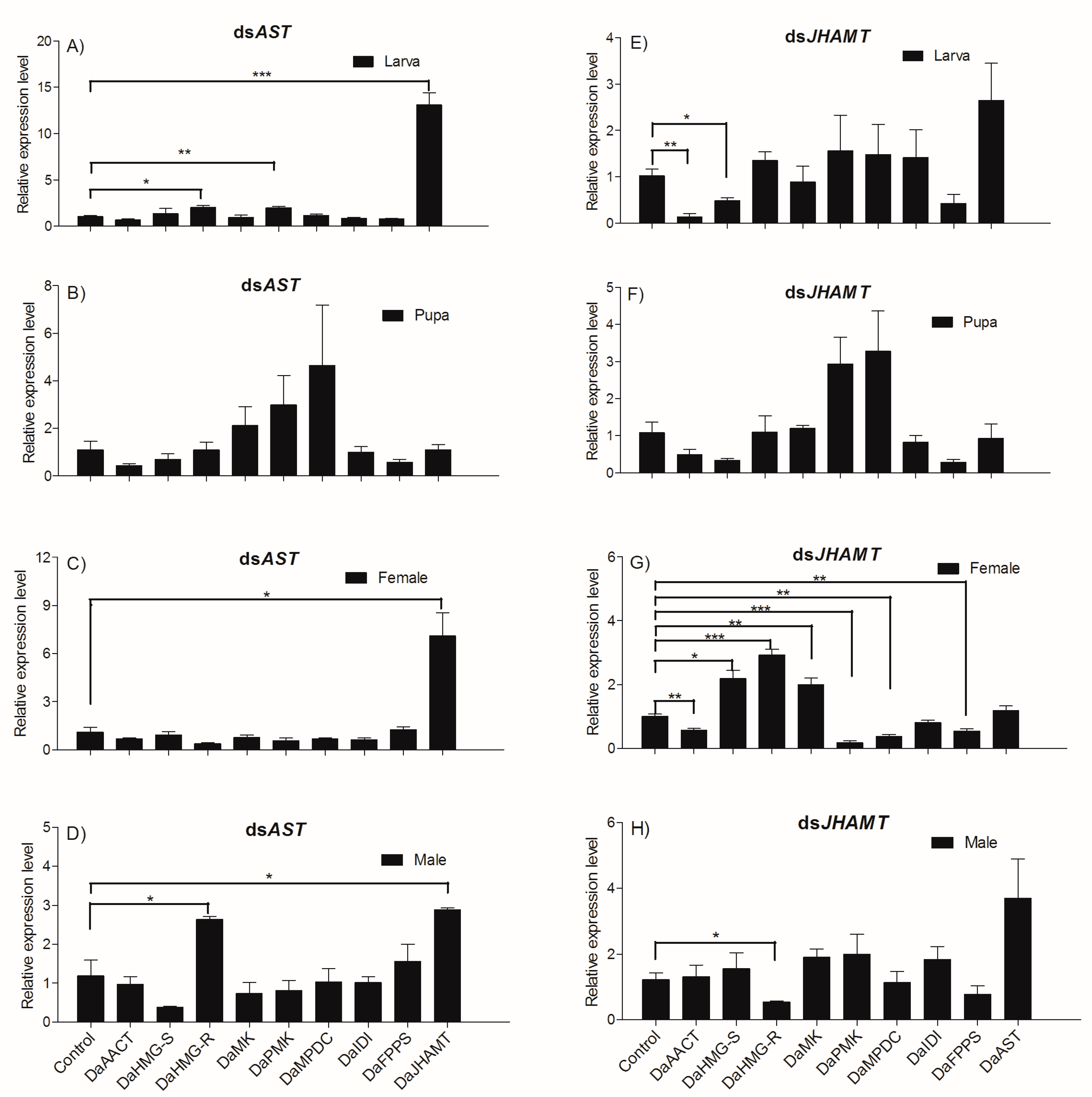
2.5.2. Knockdown Effect of Injecting dsAST and dsJHAMT Separately
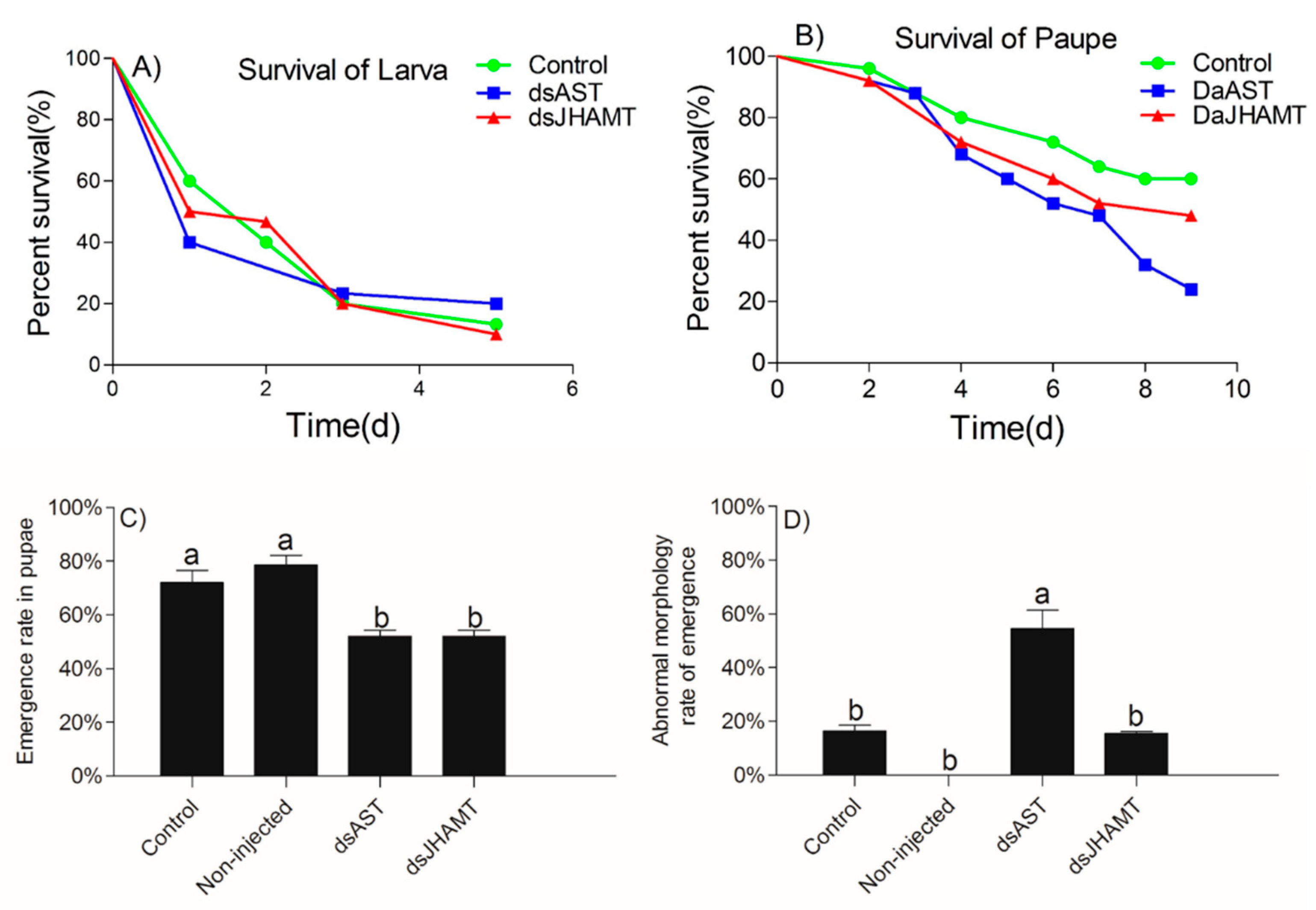
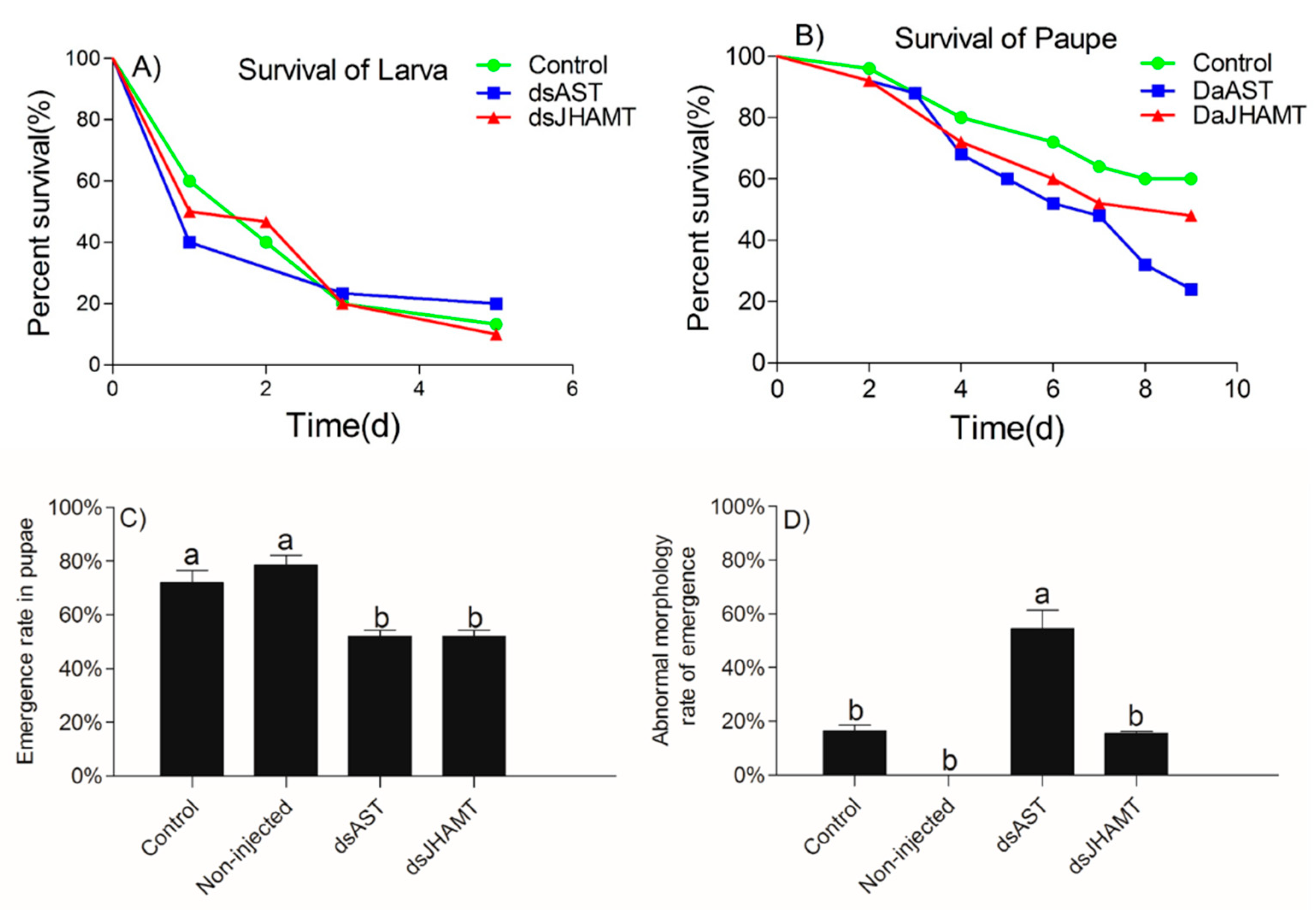
2.5.3. The Effects of RNAi of dsAST and dsJHAMT on the Development of D. armandi
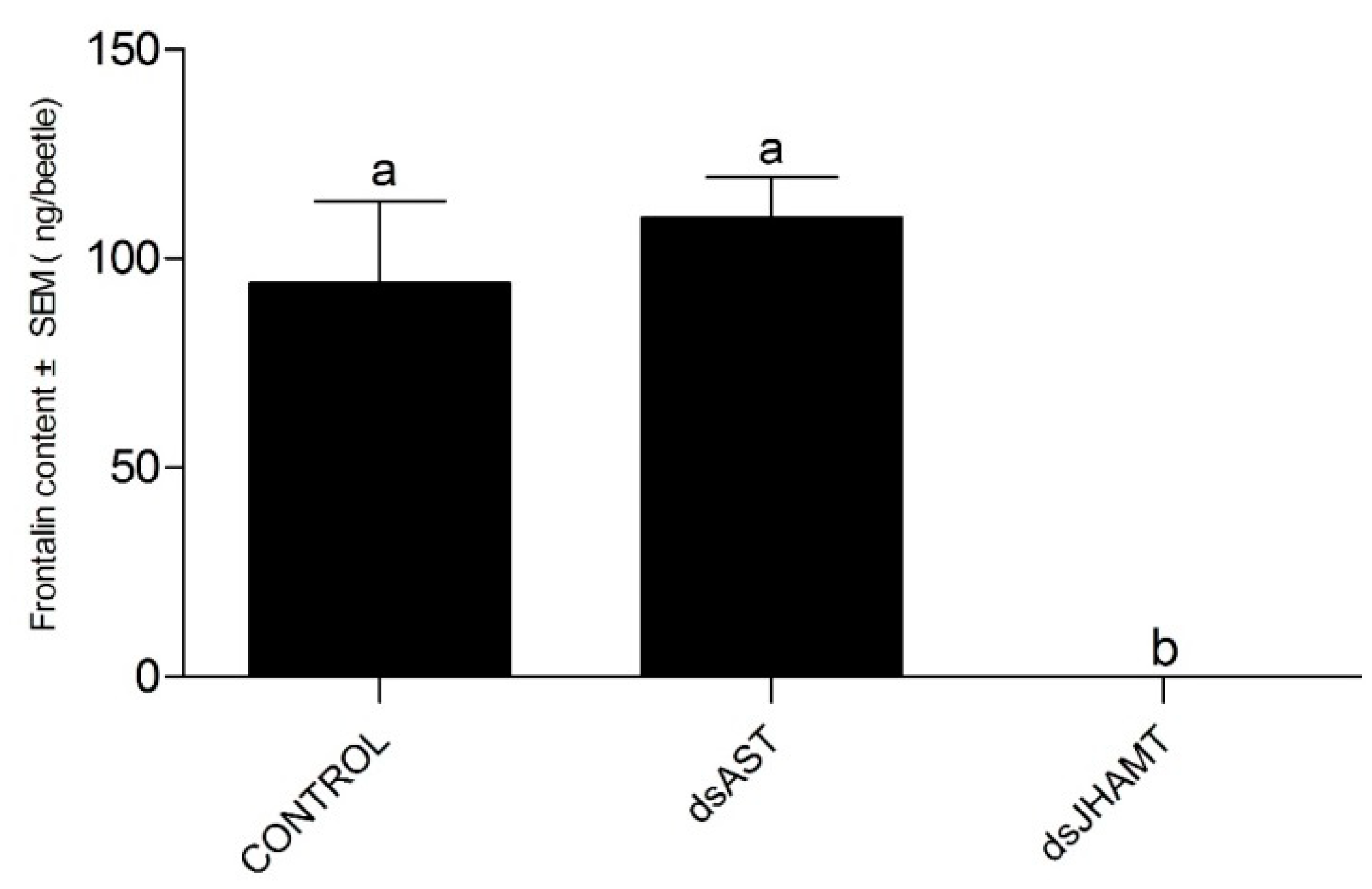
2.5.4. Adult Development of the dsRNA Phenotypes
3. Discussion
4. Materials and Methods
4.1. Insects
4.2. RNA Isolation and cDNA Synthesis
4.3. Amplification of Genes, Cloning and Sequence Analyses
4.4. Sequence Analyses of the Genes
4.5. Analysis of the DaAST and DaJHAMT Genes Transcript Levels (Real Time-qPCR)
4.5.1. Expression Patterns of Different Life Stages and Tissues
4.5.2. Effects of JHA Injection on Transcript Levels of DaAST and DaJHAMT
4.6. dsRNA Synthesis
4.6.1. Target Genes
4.6.2. Vector Construction and Expression
- 1.
- Construction of transformed E. coli expressing dsRNA
- 2.
- Isolation of dsRNA using conventional method
4.6.3. RNAi Experiment
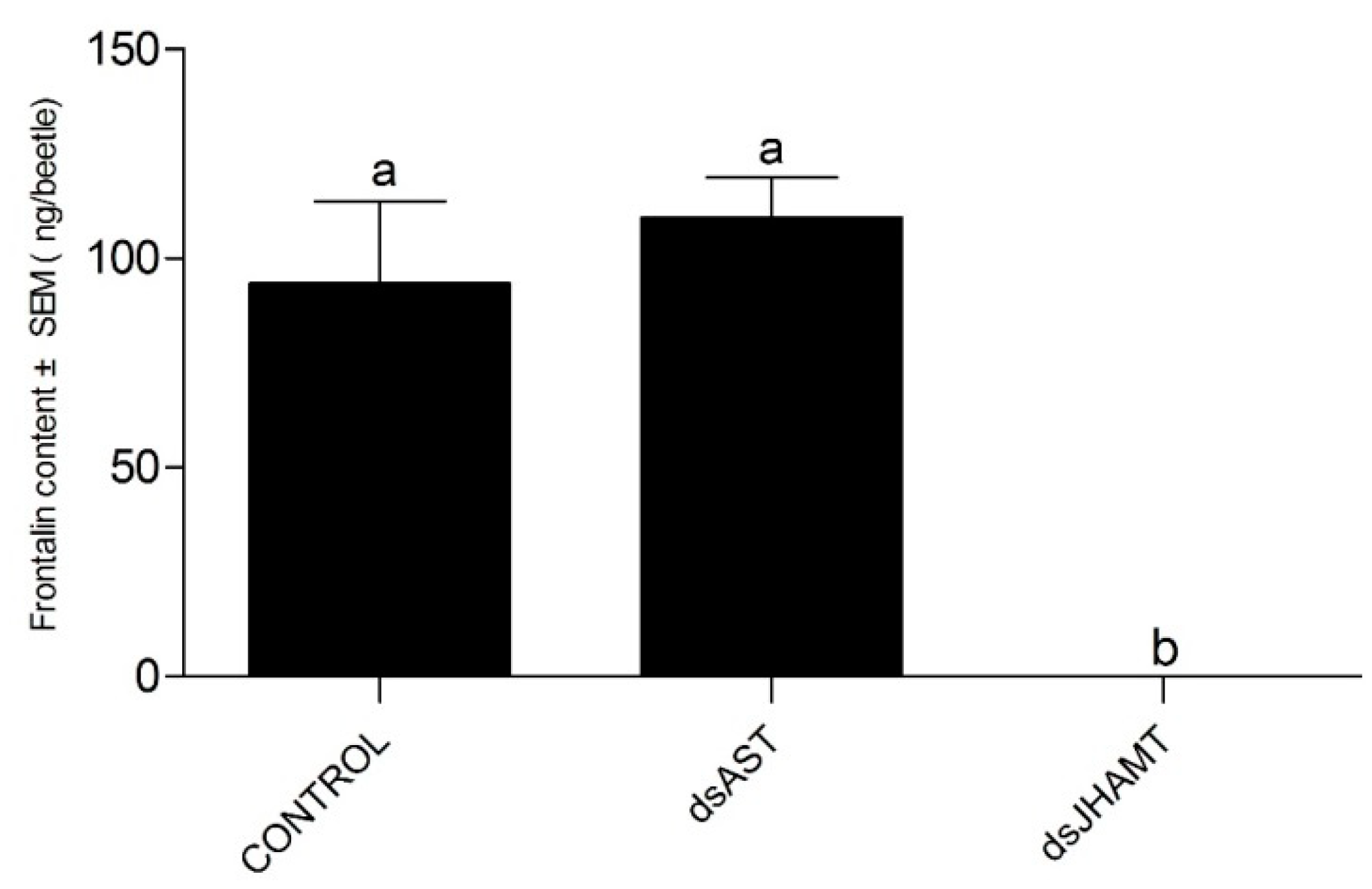
4.6.4. RNA Interference and Quantification of Frontalin
4.7. Real-Time PCR
4.8. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Wilde, J.; de Boer, J.A. Humoral and nervous pathways in photoperiodic induction of diapause in Leptinotarsa decemlineata. J. Insect Physiol. 1969, 15, 661–675. [Google Scholar] [CrossRef]
- De Loof, A.; de Wilde, J. Hormonal control of synthesis of vitellogenic female protein in the Colorado beetle, Leptinotarsa decemlineata. J. Insect Physiol. 1970, 16, 1455–1466. [Google Scholar] [CrossRef]
- Schooneveld, H.; Otazo Sanchez, A.; de Wilde, J. Juvenile hormone-induced break and termination of diapause in the Colorado potato beetle. J. Insect Physiol. 1977, 23, 689–696. [Google Scholar] [CrossRef]
- Kramer, S.J. Age-dependent changes in corpus allatum activity in vitro in the adult Colorado potato beetle, Leptinotarsa decemlineata. J. Insect Physiol. 1978, 24, 461–464. [Google Scholar] [CrossRef]
- Dortland, J.F. The hormonal control of vitellogenin synthesis in the fat body of the female Colorado potato beetle. Gen. Comp. Endocrinol. 1979, 38, 332–344. [Google Scholar] [CrossRef]
- Vermunt, A.M.W.; Koopmanschap, A.B.; Vlak, J.M.; de Kort, C.A.D. Expression of the juvenile hormone esterase gene in the Colorado potato beetle, Leptinotarsa decemlineata: Photoperiodic and juvenile hormone analog response. J. Insect Physiol. 1999, 45, 135–142. [Google Scholar] [CrossRef]
- Dominguez, C.V.; Maestro, J.L. Expression of juvenile hormone acid O-methyltransferase and juvenile hormone synthesis in Blattella germanica. Insect Sci. 2018, 25, 787–796. [Google Scholar] [CrossRef] [Green Version]
- Goodman, W.G.; Granger, N.A. The Juvenile Hormones. Compr. Mol. Insect Sci. 2005, 3, 319–408. [Google Scholar]
- Noriega, F.G. Juvenile hormone biosynthesis in insects: What is new, what do we know, and what questions remain? Int. Sch. Res. Not. 2014, 2014, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Belles, X.; Martin, D.; Piulachs, M.D. The mevalonate pathway and the synthesis of juvenile hormone in insects. Annu. Rev. Entomol. 2005, 50, 181–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz, M.E.; Mayoral, J.G.; Priestap, H.; Nouzova, M.; Rivera-Perez, C.; Noriega, F.G. Characterization of an isopentenyl diphosphate isomerase involved in the juvenile hormone pathway in Aedes aegypti. Insect Biochem. Mol. Biol. 2012, 42, 751–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Dai, L.; Zhang, R.; Li, Z.; Pham, T.; Chen, H. Isolation and expression of HMG-CoA synthase and HMG-CoA reductase genes in different development stages, tissues and treatments of the Chinese white pine beetle, Dendroctonus armandi (Curculionidae: Scolytinae). Comp. Biochem. Physiol. Part B 2015, 187, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Dai, L.; Kang, X.; Fu, D.; Gao, H.; Chen, H. Isolation and expression of five genes in the mevalonate pathway of the Chinese white pine beetle, Dendroctonus armandi (Curculionidae: Scolytinae). Arch. Insect Biochem. Physiol. 2020, 106, e21760. [Google Scholar] [CrossRef]
- Goodman, W.G.; Cusson, M. The Juvenile Hormones. In Insect Endocrinology; Gilbert, L.I., Ed.; Elsevier: San Diego, CA, USA, 2012; pp. 310–365. [Google Scholar]
- Wen, D.; Rivera-Perez, C.; Abdou, M.; Jia, Q.; He, Q.; Liu, X.; Zyaan, O.; Xu, J.; Bendena, W.G.; Tobe, S.S.; et al. Methyl Farnesoate plays a dual role in regulating Drosophila metamorphosis. PLoS Genet. 2015, 11, e1005038. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wen, D.; Li, E.Y.; Palli, S.R.; Li, S.; Wang, J.; Liu, S. MicroRNA miR-8 promotes cell growth of corpus allatum and juvenile hormone biosynthesis independent of insulin/IGF signaling in Drosophila melanogaster. Insect Biochem. Mol. Biol. 2021, 136, 103611. [Google Scholar] [CrossRef]
- Shinoda, T.; Itoyama, K. Juvenile hormone acid methyltransferase: A key regulatory enzyme for insect metamorphosis. Proc. Natl. Acad. Sci. USA 2003, 100, 11986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daimon, T.; Kozaki, T.; Niwa, R.; Kobayashi, I.; Furuta, K.; Namiki, T.; Uchino, K.; Banno, Y.; Katsuma, S.; Tamura, T.; et al. Precocious metamorphosis in the juvenile hormone–deficient mutant of the Silkworm, Bombyx mori. PLoS Genet. 2012, 8, e1002486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minakuchi, C.; Namiki, T.; Yoshiyama, M.; Shinoda, T. RNAi-mediated knockdown of juvenile hormone acid O-methyltransferase gene causes precocious metamorphosis in the red flour beetle Tribolium castaneum. FEBS J. 2008, 275, 2919–2931. [Google Scholar] [CrossRef] [PubMed]
- Kinjoh, T.; Kaneko, Y.; Itoyama, K.; Mita, K.; Hiruma, K.; Shinoda, T. Control of juvenile hormone biosynthesis in Bombyx mori: Cloning of the enzymes in the mevalonate pathway and assessment of their developmental expression in the corpora allata. Insect Biochem. Mol. Biol. 2007, 37, 808–818. [Google Scholar] [CrossRef] [PubMed]
- Minakuchi, C.; Ishii, F.; Washidu, Y.; Ichikawa, A.; Tanaka, T.; Miura, K.; Shinoda, T. Expressional and functional analysis of CYP15A1, a juvenile hormone epoxidase, in the red flour beetle Tribolium castaneum. J. Insect Physiol. 2015, 80, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Niwa, R.; Niimi, T.; Honda, N.; Yoshiyama, M.; Itoyama, K.; Kataoka, H.; Shinoda, T. Juvenile hormone acid O-methyltransferase in Drosophila melanogaster. Insect Biochem. Mol. Biol. 2008, 38, 714–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nouzova, M.; Edwards, M.J.; Mayoral, J.G.; Noriega, F.G. A coordinated expression of biosynthetic enzymes controls the flux of juvenile hormone precursors in the corpora allata of mosquitoes. Insect Biochem. Mol. Biol. 2011, 41, 660–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homma, Y.; Mita, K.; Nakamura, Y.; Namiki, T.; Noda, H.; Shinoda, T.; Togawa, T. Identification of novel genes expressed highly and selectively in the corpora allata of the silkworm, Bombyx mori (Lepidoptera: Bombycidae). Appl. Entomol. Zool. 2019, 55, 45–54. [Google Scholar] [CrossRef]
- Bellés, X.; Maestro, J.-L.; Piulachs, M.-D.; Johnsen, A.H.; Duve, H.; Thorpe, A. Allatostatic neuropeptides from the cockroach Blattella germanica (L.) (Dictyoptera, Blattellidae). Identification, immunolocalization and activity. Regul. Pept. 1994, 53, 237–247. [Google Scholar] [CrossRef]
- Verlinden, H.; Gijbels, M.; Lismont, E.; Lenaerts, C.; Vanden Broeck, J.; Marchal, E. The pleiotropic allatoregulatory neuropeptides and their receptors: A mini-review. J. Insect Physiol. 2015, 80, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Gruntenko, N.E.; Rauschenbach, I.Y. Interplay of JH, 20E and biogenic amines under normal and stress conditions and its effect on reproduction. J. Insect Physiol. 2008, 54, 902–908. [Google Scholar] [CrossRef]
- Borras-Castells, F.; Nieva, C.; Maestro, J.L.; Maestro, O.; Belles, X.; Martín, D. Juvenile hormone biosynthesis in adult Blattella germanica requires nuclear receptors Seven-up and FTZ-F1. Sci. Rep. 2017, 7, e40234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, S.J.; Toschi, A.; Miller, C.A.; Kataoka, H.; Quistad, G.B.; Li, J.P.; Carney, R.L.; Schooley, D.A. Identification of an allatostatin from the tobacco hornworm Manduca sexta. Proc. Natl. Acad. Sci. USA 1991, 88, 9458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackburn, M.B.; Wagner, R.M.; Kochansky, J.P.; Harrison, D.J.; Thomas-Laemont, P.; Raina, A.K. The identification of two myoinhibitory peptides, with sequence similarities to the galanins, isolated from the ventral nerve cord of Manduca sexta. Regul. Pept. 1995, 57, 213–219. [Google Scholar] [CrossRef]
- Blackburn, M.B.; Jaffe, H.; Kochansky, J.; Raina, A.K. Identification of four additional myoinhibitory peptides (MIPs) from the ventral nerve cord of Manduca sexta. Arch. Insect Biochem. Physiol. 2001, 48, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, K.H.; Meyering-Vos, M.; Lorenz, M.W. Allatostatins and allatotropins: Is the regulation of corpora allata activity their primary function? Eur. J. Entomol. 1999, 96, 255–266. [Google Scholar]
- Lorenz, M.W.; Kellner, R.; Hoffmann, K.H. A family of neuropeptides that inhibit juvenile hormone biosynthesis in the Cricket, Gryllus bimaculatus. J. Biol. Chem. 1995, 270, 21103–21108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenz, M.W.; Kellner, R.; Hoffmann, K.H.; Gäde, G. Identification of multiple peptides homologous to cockroach and cricket allatostatins in the stick insect Carausius morosus. Insect Biochem. Mol. Biol. 2000, 30, 711–718. [Google Scholar] [CrossRef]
- Lubawy, J.; Marciniak, P.; Kuczer, M.; Rosiński, G. Myotropic activity of allatostatins in tenebrionid beetles. Neuropeptides 2018, 70, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Weaver, R.J.; Audsley, N. Neuropeptides of the beetle, Tenebrio molitor identified using MALDI-TOF mass spectrometry and deduced sequences from the Tribolium castaneum genome. Peptides 2008, 29, 168–178. [Google Scholar] [CrossRef]
- Woodhead, A.P.; Stay, B.; Seidel, S.L.; Khan, M.A.; Tobe, S.S. Primary structure of four allatostatins: Neuropeptide inhibitors of juvenile hormone synthesis. Proc. Natl. Acad. Sci. USA 1989, 86, 5997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kataoka, H.; Toschi, A.; Li Jorge, P.; Carney Robert, L.; Schooley David, A.; Kramer Steven, J. Identification of an allatotropin from adult Manduca Sexta. Science 1989, 243, 1481–1483. [Google Scholar] [CrossRef] [PubMed]
- Janzen, W.P.; Menold, M.; Granger, N.A. Effects of endogenous esterases and an allatostatin on the products of Manduca sexta larval corpora allata in vitro. Physiol. Entomol. 1991, 16, 283–293. [Google Scholar] [CrossRef]
- Coast, G.M.; Schooley, D.A. Toward a consensus nomenclature for insect neuropeptides and peptide hormones. Peptides 2011, 32, 620–631. [Google Scholar] [CrossRef] [PubMed]
- Abdel-latief, M.; Hoffmann, K.H. Neuropeptide regulators of the juvenile hormone biosynthesis (in vitro) in the beetle, Tenebrio molitor (Coleoptera, Tenebrionidae). Arch. Insect Biochem. Physiol. 2010, 74, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Abdel-latief, M.; Hoffmann, K.H. Functional activity of allatotropin and allatostatin in the pupal stage of a holometablous insect, Tribolium castaneum (Coleoptera, Tenebrionidae). Peptides 2014, 53, 172–184. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.-W.; Liu, X.-P.; Lü, F.-G.; Fu, K.-Y.; Guo, W.-C.; Li, G.-Q. Involvement of a putative allatostatin in regulation of juvenile hormone titer and the larval development in Leptinotarsa decemlineata (Say). Gene 2015, 554, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Zamore, P.D. RNA interference: Listening to the sound of silence. Nat. Struct. Biol. 2001, 8, 746–750. [Google Scholar] [CrossRef] [PubMed]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Adams Mark, D.; Celniker Susan, E.; Holt Robert, A.; Evans Cheryl, A.; Gocayne Jeannine, D.; Amanatides Peter, G.; Scherer Steven, E.; Li Peter, W.; Hoskins Roger, A.; Galle Richard, F.; et al. The genome sequence of Drosophila melanogaster. Science 2000, 287, 2185–2195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baum, J.A.; Bogaert, T.; Clinton, W.; Heck, G.R.; Feldmann, P.; Ilagan, O.; Johnson, S.; Plaetinck, G.; Munyikwa, T.; Pleau, M.; et al. Control of coleopteran insect pests through RNA interference. Nat. Biotechnol. 2007, 25, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Price, D.R.G.; Gatehouse, J.A. RNAi-mediated crop protection against insects. Trends Biotechnol. 2008, 26, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Huvenne, H.; Smagghe, G. Mechanisms of dsRNA uptake in insects and potential of RNAi for pest control: A review. J. Insect Physiol. 2010, 56, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Schlüns, H.; Crozier, R.H. Relish regulates expression of antimicrobial peptide genes in the honeybee, Apis mellifera, shown by RNA interference. Insect Mol. Biol. 2007, 16, 753–759. [Google Scholar] [CrossRef]
- Marco Antonio, D.S.; Guidugli-Lazzarini, K.R.; do Nascimento, A.M.; Paulino Simoes, Z.L.; Hartfelder, K. RNAi-mediated silencing of vitellogenin gene function turns honeybee (Apis mellifera) workers into extremely precocious foragers. Naturwissenschaften 2008, 95, 953–961. [Google Scholar] [CrossRef]
- Lum, L.; Yao, S.; Mozer, B.; Rovescalli, A.; Von Kessler, D.; Nirenberg, M.; Beachy, P.A. Identification of hedgehog pathway components by RNAi in Drosophila cultured cells. Science 2003, 299, 2039–2045. [Google Scholar] [CrossRef] [PubMed]
- Dietzl, G.; Chen, D.; Schnorrer, F.; Su, K.-C.; Barinova, Y.; Fellner, M.; Gasser, B.; Kinsey, K.; Oppel, S.; Scheiblauer, S.; et al. A genome-wide transgenic RNAi library for conditional gene inactivation in Drosophila. Nature 2007, 448, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Arakane, Y.; Hogenkamp, D.G.; Zhu, Y.C.; Kramer, K.J.; Specht, C.A.; Beeman, R.W.; Kanost, M.R.; Muthukrishnan, S. Characterization of two chitin synthase genes of the red flour beetle, Tribolium castaneum, and alternate exon usage in one of the genes during development. Insect Biochem. Mol. Biol. 2004, 34, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Truman, J.W.; Riddiford, L.M. The role of Broad in the development of Tribolium castaneum: Implications for the evolution of the holometabolous insect pupa. Development 2008, 135, 569–577. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Tian, H.; Zou, L.; Tang, B.; Hu, J.; Zhang, W. Disruption of Spodoptera exigua larval development by silencing chitin synthase gene A with RNA interference. Bull. Entomol. Res. 2008, 98, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Peng, H.; Yao, Q.; Chen, H.; Xie, Q.; Tang, B.; Zhang, W. Developmental control of a Lepidopteran pest Spodoptera exigua by ingestion of bacteria expressing dsRNA of a non-midgut gene. PLoS ONE 2009, 4, e6225. [Google Scholar] [CrossRef] [PubMed]
- Terenius, O.; Papanicolaou, A.; Garbutt, J.S.; Eleftherianos, I.; Huvenne, H.; Kanginakudru, S.; Albrechtsen, M.; An, C.; Aymeric, J.-L.; Barthel, A.; et al. RNA interference in Lepidoptera: An overview of successful and unsuccessful studies and implications for experimental design. J. Insect Physiol. 2011, 57, 231–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhargava, A.; Clifton, M.S.; Mhsake, P.; Liao, M.; Pothoulakis, C.; Leeman, S.E.; Grady, E.F. Local injection of dsRNA targeting calcitonin receptor-like receptor (CLR) ameliorates Clostridium difficile toxin A-induced ileitis. Proc. Natl. Acad. Sci. USA 2013, 110, 731–736. [Google Scholar] [CrossRef] [Green Version]
- Yu, N.; Christiaens, O.; Liu, J.; Niu, J.; Cappelle, K.; Caccia, S.; Huvenne, H.; Smagghe, G. Delivery of dsRNA for RNAi in insects: An overview and future directions. Insect Sci. 2012, 20, 4–14. [Google Scholar] [CrossRef]
- Mao, Y.-B.; Cai, W.-J.; Wang, J.-W.; Hong, G.-J.; Tao, X.-Y.; Wang, L.-J.; Huang, Y.-P.; Chen, X.-Y. Silencing a cotton bollworm P450 monooxygenase gene by plant-mediated RNAi impairs larval tolerance of gossypol. Nat. Biotechnol. 2007, 25, 1307–1313. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Gasteiger, J.; Hristozov, D. Coding and decoding chemical structure information. Abstr. Pap. Am. Chem. Soc. 2005, 229, 761. [Google Scholar]
- Emanuelsson, O.; Nielsen, H.; Brunak, S.; von Heijne, G. Predicting subcellular localization of proteins based on their N-terminal amino acid sequence. J. Mol. Biol. 2000, 300, 1005–1016. [Google Scholar] [CrossRef] [Green Version]
- Veenstra, J.A. Mono- and dibasic proteolytic cleavage sites in insect neuroendocrine peptide precursors. Arch. Insect Biochem. Physiol. 2000, 43, 49–63. [Google Scholar] [CrossRef]
- Lubawy, J.; Marciniak, P.; Rosiński, G. Identification, localization in the central nervous system and novel myostimulatory effect of allatostatins in Tenebrio molitor beetle. Int. J. Mol. Sci. 2020, 21, 3510. [Google Scholar] [CrossRef]
- Defelipe, L.A.; Dolghih, E.; Roitberg, A.E.; Nouzova, M.; Mayoral, J.G.; Noriega, F.G.; Turjanski, A.G. Juvenile hormone synthesis: “esterify then epoxidize” or “epoxidize then esterify”? Insights from the structural characterization of juvenile hormone acid methyltransferase. Insect Biochem. Mol. Biol. 2011, 41, 228–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiesari, L.; Kyriacou, C.P.; Costa, R. The hormonal and circadian basis for insect photoperiodic timing. FEBS Lett. 2011, 585, 1450–1460. [Google Scholar] [CrossRef]
- Nouzova, M.; Michalkova, V.; Ramirez, C.E.; Fernandez-Lima, F.; Noriega, F.G. Inhibition of juvenile hormone synthesis in mosquitoes by the methylation inhibitor 3-deazaneplanocin A (DZNep). Insect Biochem. Mol. Biol. 2019, 113, 103183. [Google Scholar] [CrossRef]
- Ma, H.-Y.; Li, Y.-Y.; Li, L.; Tan, Y.; Pang, B.-P. Regulation of juvenile hormone on summer diapause of Geleruca daurica and its pathway analysis. Insects 2021, 12, 237. [Google Scholar] [CrossRef]
- Liu, W.; Li, Y.; Zhu, L.; Zhu, F.; Lei, C.-L.; Wang, X.-P. Juvenile hormone facilitates the antagonism between adult reproduction and diapause through the methoprene-tolerant gene in the female Colaphellus bowringi. Insect Biochem. Mol. Biol. 2016, 74, 50–60. [Google Scholar] [CrossRef]
- Tobe, S.S.; Stay, B. Modulation of juvenile hormone synthesis by an analogue in the cockroach. Nature 1979, 281, 481–482. [Google Scholar] [CrossRef]
- Rüegg, R.P.; Lococo, D.J.; Tobe, S.S. Control of corpus allatum activity in Diploptera punctata: Roles of the pars intercerebralis and pars lateralis. Experientia 1983, 39, 1329–1334. [Google Scholar] [CrossRef]
- Stay, B.; Sereg Bachmann, J.A.; Stoltzman, C.A.; Fairbairn, S.E.; Yu, C.G.; Tobe, S.S. Factors affecting allatostatin release in a cockroach (Diploptera punctata): Nerve section, juvenile hormone analog and ovary. J. Insect Physiol. 1994, 40, 365–372. [Google Scholar] [CrossRef]
- Khan, M.A.; Koopmanschap, A.B.; de Kort, C.A.D. The effects of juvenile hormone, 20-hydroxyecdysone and precocene II on activity of corpora allata and the mode of negative-feedback regulation of these glands in the adult Colorado potato beetle. J. Insect Physiol. 1982, 28, 995–1001. [Google Scholar] [CrossRef]
- Khan, M.A.; Koopmanschap, A.B.; Privee, H.; de Kort, C.A.D. The mode of regulation of the corpus allatum activity during starvation in adult females of the Colorado potato beetle, Leptinotarsa decemlineata (Say). J. Insect Physiol. 1982, 28, 791–796. [Google Scholar] [CrossRef]
- Veenstra, J.A. Allatostatin C and its paralog allatostatin double C: The arthropod somatostatins. Insect Biochem. Mol. Biol. 2009, 39, 161–170. [Google Scholar] [CrossRef]
- Dong, Y.-Q.; Wang, Z.-Y.; Jing, T.-Z. Functional characterization of ASTC (allatostatin C) and ASTCC (allatostatin double C) in Clostera anastomosis (Lepidoptera: Notodontidae). Gene 2017, 598, 1–8. [Google Scholar] [CrossRef]
- Griebler, M.; Westerlund, S.A.; Hoffmann, K.H.; Meyering-Vos, M. RNA interference with the allatoregulating neuropeptide genes from the fall armyworm Spodoptera frugiperda and its effects on the JH titer in the hemolymph. J. Insect Physiol. 2008, 54, 997–1007. [Google Scholar] [CrossRef]
- Konopova, B.; Jindra, M. Broad-Complex acts downstream of Met in juvenile hormone signaling to coordinate primitive holometabolan metamorphosis. Development 2008, 135, 559–568. [Google Scholar] [CrossRef] [Green Version]
- Weaver, R.J.; Audsley, N. Neuropeptide regulators of juvenile hormone synthesis. Ann. N. Y. Acad. Sci. 2009, 1163, 316–329. [Google Scholar] [CrossRef]
- Stay, B. A review of the role of neurosecretion in the control of juvenile hormone synthesis: A tribute to Berta Scharrer. Insect Biochem. Mol. Biol. 2000, 30, 653–662. [Google Scholar] [CrossRef]
- Dai, L.; Wang, C.; Zhang, X.; Yu, J.; Zhang, R.; Chen, H. Two CYP4 genes of the Chinese white pine beetle, Dendroctonus armandi (Curculionidae: Scolytinae), and their transcript levels under different development stages and treatments. Insect Mol. Biol. 2014, 23, 598–610. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Dai, L.; Fu, D.; Gao, J.; Chen, H. Electrophysiological and behavioral responses of Dendroctonus armandi (Coleoptera: Curculionidae: Scolytinae) to two candidate pheromone components: Frontalin and exo-brevicomin. Chemoecology 2017, 27, 91–99. [Google Scholar] [CrossRef]
- Le, S.Q.; Gascuel, O. An improved general amino acid replacement matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.Y.; Lin, S.; Ahn, K. Methoprene does not affect juvenile hormone titers in honey bee (Apis mellifera) workers. Insect Sci. 2016, 25, 235–240. [Google Scholar] [CrossRef]
- Sun, Y.; Fu, D.; Kang, X.; Liu, B.; Ning, H.; Chen, H. Function of mevalonate pathway genes in the synthesis of frontalin in Chinese white pine beetle, Dendroctonus armandi (curculionidae: Scolytinae). Arch. Insect Biochem. Physiol. 2021, 107, e21828. [Google Scholar] [CrossRef]
- Keeling, C.I.; Chiu, C.C.; Aw, T.; Li, M.; Henderson, H.; Tittiger, C.; Weng, H.B.; Blomquist, G.J.; Bohlmann, J. Frontalin pheromone biosynthesis in the mountain pine beetle, Dendroctonus ponderosae, and the role of isoprenyl diphosphate synthases. Proc. Natl. Acad. Sci. USA 2013, 110, 18838–18843. [Google Scholar] [CrossRef] [Green Version]
- Dai, L.; Ma, M.; Wang, C.; Shi, Q.; Zhang, R.; Chen, H. Cytochrome P450s from the Chinese white pine beetle, Dendroctonus armandi (Curculionidae: Scolytinae): Expression profiles of different stages and responses to host allelochemicals. Insect Biochem. Mol. Biol. 2015, 65, 35–46. [Google Scholar] [CrossRef]
- Gillespie, M.J.; Fisher, L. Confidence bands for the kaplan-meier survival curve estimate. Ann. Stat. 1979, 7, 920–924. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | BLAST Matches in Genbank | Identity 1 | ||
|---|---|---|---|---|
| Species | Gene | Accession No. | BlastP | |
| DaAST | Dendroctonus ponderosae | AST | XP_019760338.1 | 69% |
| Tribolium castaneum | AST | NP_001137205.1 | 67% | |
| Leptinotarsa decemlineata | AST | AIW62334.1 | 64% | |
| DaJHAMT | Dendroctonus ponderosae | JHAMT | XP_019760146.1 | 53% |
| Nicrophorus vespilloides | JHAMT | XP_017772157.1 | 36.8% | |
| Aethina tumida | JHAMT | XP_019876726.1 | 36.5% | |
| Gene Name | Accession No. | Full Length (bp) 1 | ORF Size (aa/bp) 1 | Mw (kDa) 1 | I.P. 1 | Signal Peptide Prediction 2 |
|---|---|---|---|---|---|---|
| DaAST | MW645339 | 471 | 114/345 | 13.28 | 8.66 | SP 0.766 mTP 0.043 other 0.194 |
| DaJHAMT | MW645341 | 1137 | 275/828 | 15.11 | 9.04 | SP 0.069 mTP 0.062 other 0.932 |
| Groups | Emergence (%) | Adult Deformity (%) | Death of Deformed Adults (%) | Days of Death (d) |
|---|---|---|---|---|
| Control | 72.00 | 16.39 | 66.67 | 4 |
| Non-injected | 78.67 | 1.33 | 100 | 3 |
| dsAST | 52.00 | 54.46 | 71.42 | 1 |
| dsJHAMT | 52.00 | 15.45 | 100 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Fu, D.; Liu, B.; Wang, L.; Chen, H. Functional Characterization of Allatostatin C (PISCF/AST) and Juvenile Hormone Acid O-Methyltransferase in Dendroctonus armandi. Int. J. Mol. Sci. 2022, 23, 2749. https://doi.org/10.3390/ijms23052749
Sun Y, Fu D, Liu B, Wang L, Chen H. Functional Characterization of Allatostatin C (PISCF/AST) and Juvenile Hormone Acid O-Methyltransferase in Dendroctonus armandi. International Journal of Molecular Sciences. 2022; 23(5):2749. https://doi.org/10.3390/ijms23052749
Chicago/Turabian StyleSun, Yaya, Danyang Fu, Bin Liu, Linjun Wang, and Hui Chen. 2022. "Functional Characterization of Allatostatin C (PISCF/AST) and Juvenile Hormone Acid O-Methyltransferase in Dendroctonus armandi" International Journal of Molecular Sciences 23, no. 5: 2749. https://doi.org/10.3390/ijms23052749
APA StyleSun, Y., Fu, D., Liu, B., Wang, L., & Chen, H. (2022). Functional Characterization of Allatostatin C (PISCF/AST) and Juvenile Hormone Acid O-Methyltransferase in Dendroctonus armandi. International Journal of Molecular Sciences, 23(5), 2749. https://doi.org/10.3390/ijms23052749