
Ventx Family and Its Functional Similarities with Nanog: Involvement in Embryonic Development and Cancer Progression



Abstract
1. Introduction
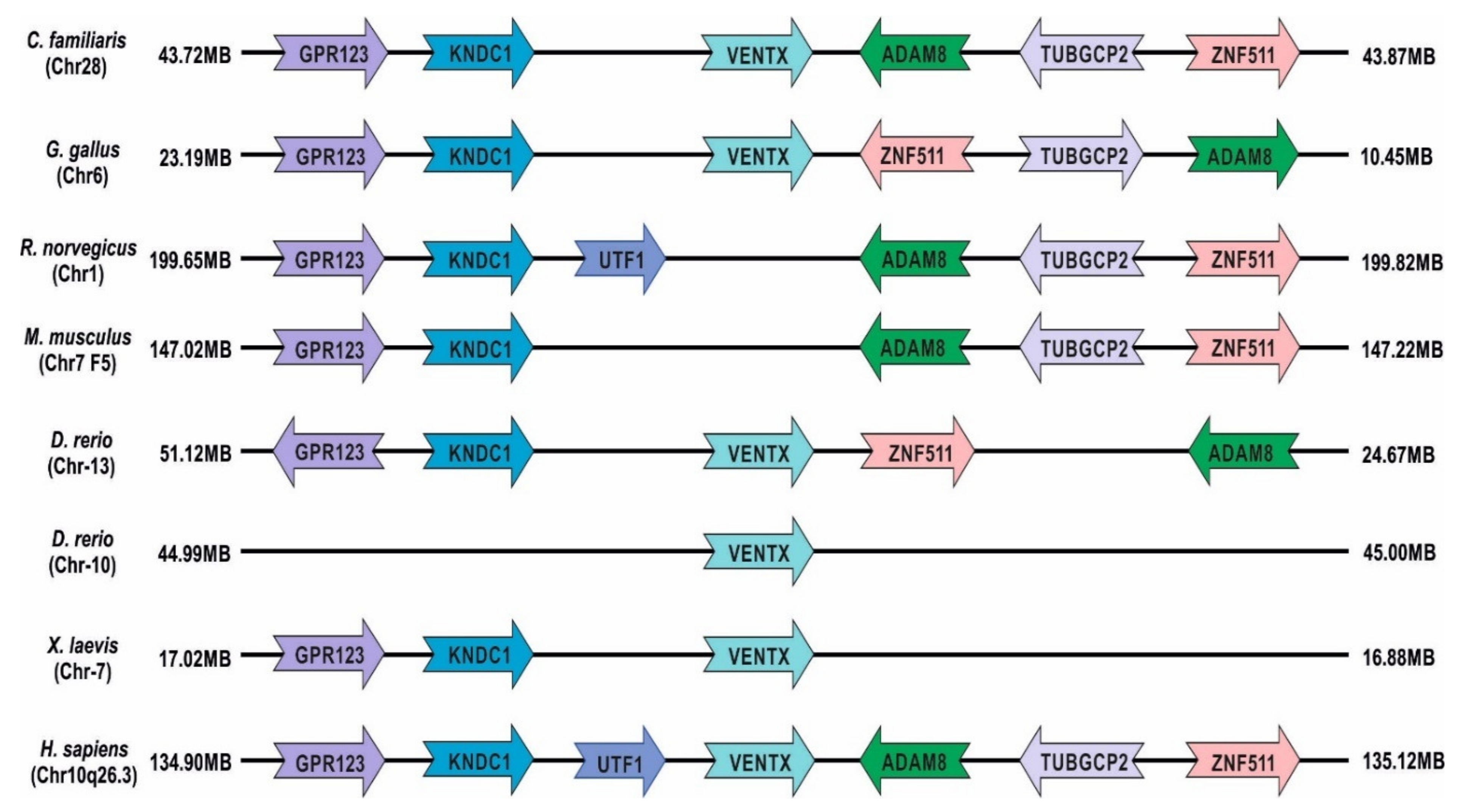
1.1. Xenopus laevis: Genome Evolution, Pseudo-Tetraploidy, and Gene Duplication
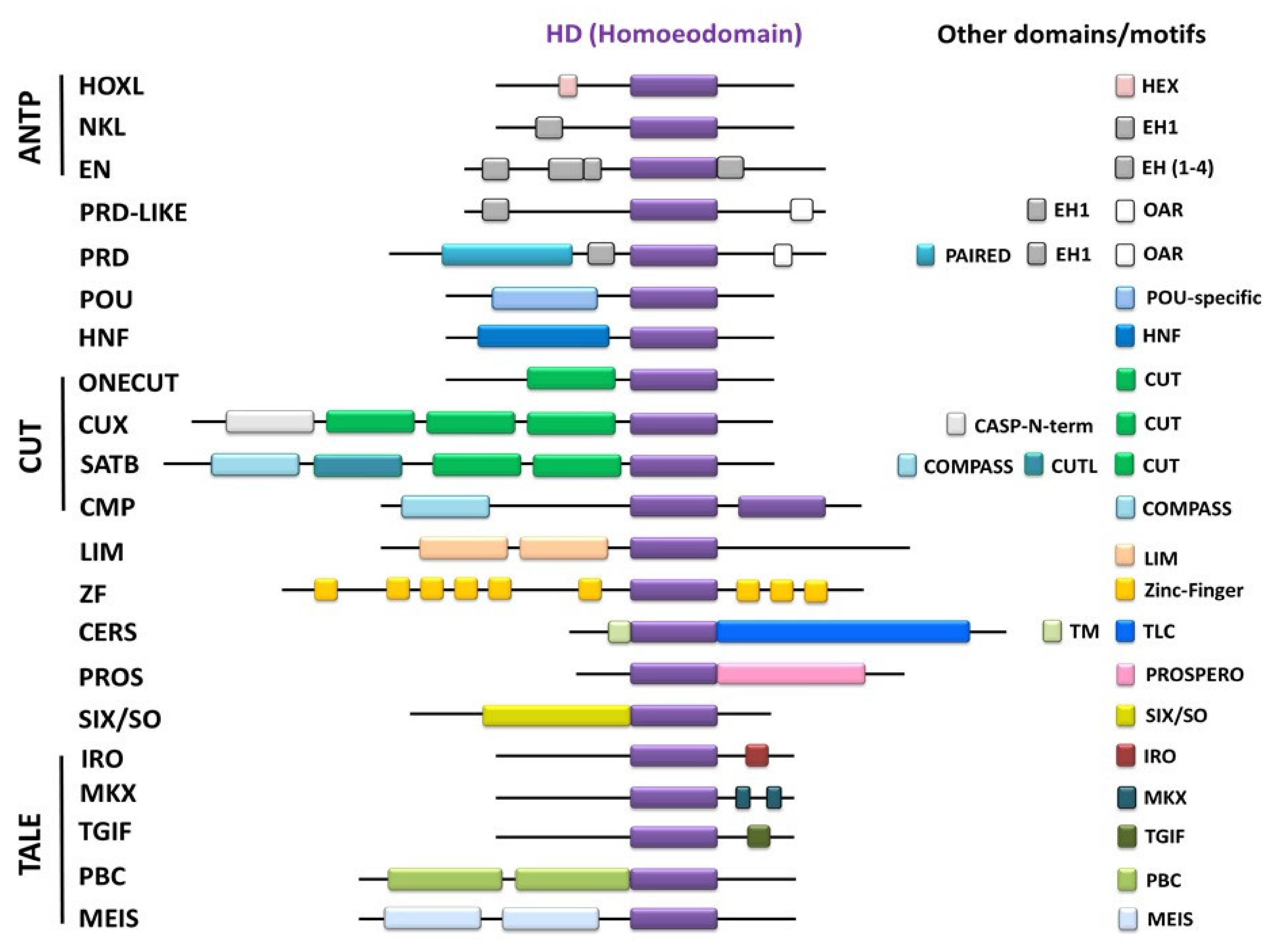
1.2. Homeobox Domain (HD)
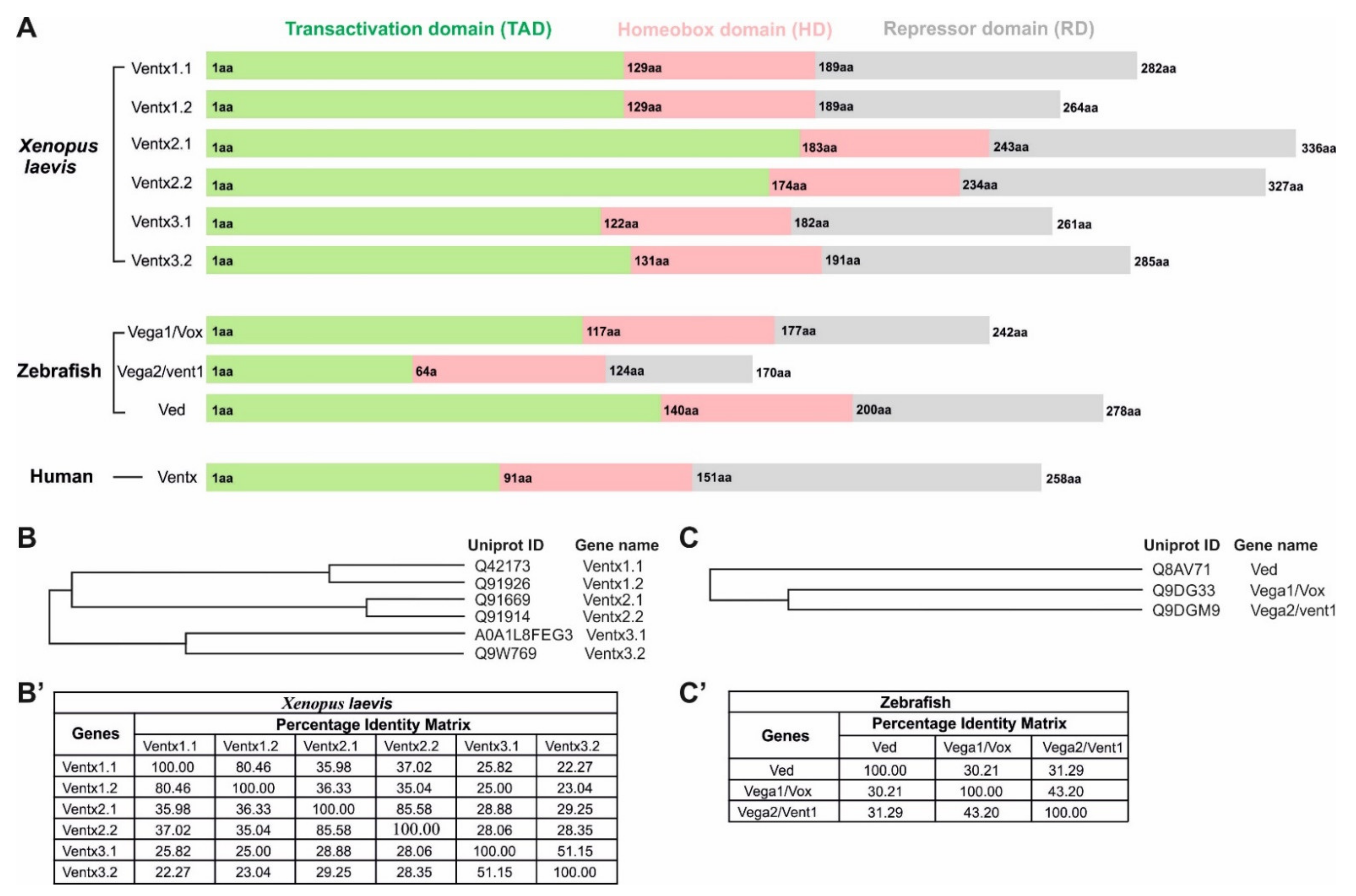
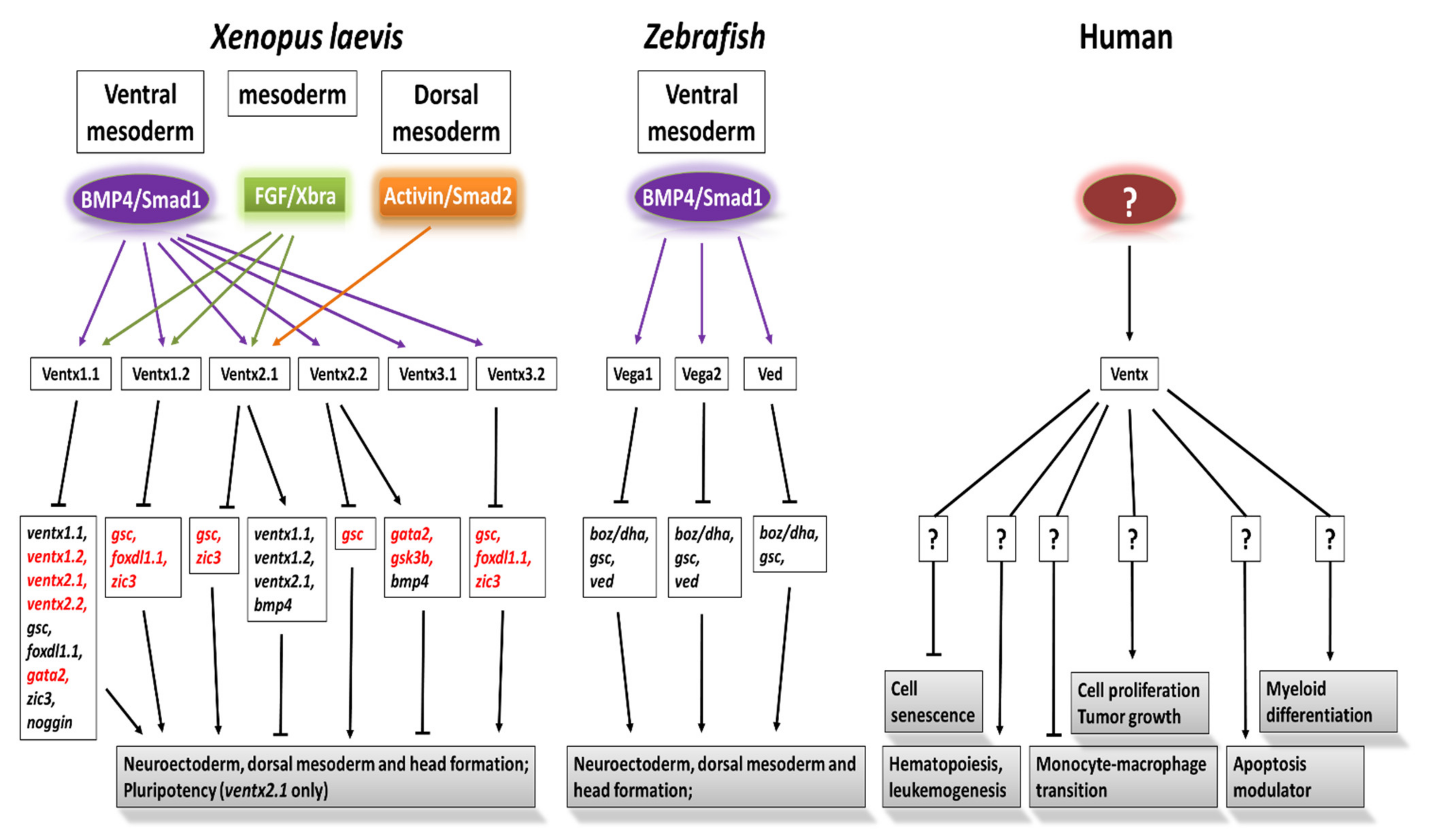
2. Ventx Family in Xenopus
2.1. Ventx1.1
2.2. Ventx1.2/Xvent1
2.3. Ventx2.1
2.4. Ventx2.2/Xom/Xbr-1
2.5. Ventx2.2 Maintains Cellular Pluripotency in Xenopus Similarly to Nanog in Other Animals
2.6. Ventx2.2 Leads to Ectomesenchyme Proliferation and Mandibular Cartilage Development in Xenopus as Barx1 Does in Rodents and Fruit Flies
2.7. Ventx3.1
2.8. Ventx3.2/Vex1
3. Vega Family in Zebrafish
3.1. Vega1/Vox
3.2. Vega2/Vent1/Vent
3.3. Ved
4. Human Ventx (VENT-like Homeobox Protein 2)
Nanog (Ventx-like) Is Considered an Oncogene in Humans
5. Discussion and Conclusions
6. Future Prospective for Targeting the GATA2- and Ventx-Associated Cancers
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Grainger, R.M. Xenopus tropicalis as a model organism for genetics and genomics: Past, present, and future. Methods Mol. Biol. 2012, 917, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Blum, M.; Ott, T. Xenopus: An Undervalued Model Organism to Study and Model Human Genetic Disease. Cells Tissues Organs 2018, 205, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Liu, X. Polyploidization and pseudogenization in allotetraploid frog Xenopus laevis promote the evolution of aquaporin family in higher vertebrates. BMC Genom. 2020, 21, 525. [Google Scholar] [CrossRef] [PubMed]
- Hellsten, U.; Khokha, M.K.; Grammer, T.C.; Harland, R.M.; Richardson, P.; Rokhsar, D.S. Accelerated gene evolution and subfunctionalization in the pseudotetraploid frog Xenopus laevis. BMC Biol. 2007, 5, 31. [Google Scholar] [CrossRef] [PubMed]
- Elurbe, D.M.; Paranjpe, S.S.; Georgiou, G.; van Kruijsbergen, I.; Bogdanovic, O.; Gibeaux, R.; Heald, R.; Lister, R.; Huynen, M.A.; van Heeringen, S.J.; et al. Regulatory remodeling in the allo-tetraploid frog Xenopus laevis. Genome Biol. 2017, 18, 198. [Google Scholar] [CrossRef]
- Session, A.M.; Uno, Y.; Kwon, T.; Chapman, J.A.; Toyoda, A.; Takahashi, S.; Fukui, A.; Hikosaka, A.; Suzuki, A.; Kondo, M.; et al. Genome evolution in the allotetraploid frog Xenopus laevis. Nature 2016, 538, 336–343. [Google Scholar] [CrossRef]
- Pollet, N.; Mazabraud, A. Insights from Xenopus genomes. Genome Dyn. 2006, 2, 138–153. [Google Scholar] [CrossRef]
- Gehring, W.J.; Affolter, M.; Burglin, T. Homeodomain proteins. Annu. Rev. Biochem. 1994, 63, 487–526. [Google Scholar] [CrossRef]
- Burglin, T.R.; Affolter, M. Homeodomain proteins: An update. Chromosoma 2016, 125, 497–521. [Google Scholar] [CrossRef]
- Burglin, T.R. Homeodomain subtypes and functional diversity. Subcell Biochem. 2011, 52, 95–122. [Google Scholar] [CrossRef]
- Holland, P.W.; Booth, H.A.; Bruford, E.A. Classification and nomenclature of all human homeobox genes. BMC Biol. 2007, 5, 47. [Google Scholar] [CrossRef] [PubMed]
- Henningfeld, K.A.; Friedle, H.; Rastegar, S.; Knochel, W. Autoregulation of Xvent-2B; direct interaction and functional cooperation of Xvent-2 and Smad1. J. Biol. Chem. 2002, 277, 2097–2103. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Umair, Z.; Yoon, J.; Lee, U.; Kim, S.C.; Park, J.B.; Lee, J.Y.; Kim, J. Xbra and Smad-1 cooperate to activate the transcription of neural repressor ventx1.1 in Xenopus embryos. Sci. Rep. 2018, 8, 11391. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Lee, S.Y.; Lee, H.; Hwang, Y.S.; Cha, S.W.; Park, S.; Lee, J.Y.; Park, J.B.; Kim, S.; Park, M.J.; et al. Direct response elements of BMP within the PV.1A promoter are essential for its transcriptional regulation during early Xenopus development. PLoS ONE 2011, 6, e22621. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Park, M.J.; Lee, S.Y.; Hwang, Y.S.; Lee, H.; Roh, D.H.; Kim, J.I.; Park, J.B.; Lee, J.Y.; Kung, H.F.; et al. Transcriptional regulation of Xbr-1a/Xvent-2 homeobox gene: Analysis of its promoter region. Biochem. Biophys. Res. Commun. 2002, 298, 815–823. [Google Scholar] [CrossRef]
- Faial, T.; Bernardo, A.S.; Mendjan, S.; Diamanti, E.; Ortmann, D.; Gentsch, G.E.; Mascetti, V.L.; Trotter, M.W.; Smith, J.C.; Pedersen, R.A. Brachyury and SMAD signalling collaboratively orchestrate distinct mesoderm and endoderm gene regulatory networks in differentiating human embryonic stem cells. Development 2015, 142, 2121–2135. [Google Scholar] [CrossRef]
- Moretti, P.A.; Davidson, A.J.; Baker, E.; Lilley, B.; Zon, L.I.; D’Andrea, R.J. Molecular cloning of a human Vent-like homeobox gene. Genomics 2001, 76, 21–29. [Google Scholar] [CrossRef]
- Rawat, V.P.; Arseni, N.; Ahmed, F.; Mulaw, M.A.; Thoene, S.; Heilmeier, B.; Sadlon, T.; D’Andrea, R.J.; Hiddemann, W.; Bohlander, S.K.; et al. The vent-like homeobox gene VENTX promotes human myeloid differentiation and is highly expressed in acute myeloid leukemia. Proc. Natl. Acad. Sci. USA 2010, 107, 16946–16951. [Google Scholar] [CrossRef]
- Genthe, J.R.; Min, J.; Farmer, D.M.; Shelat, A.A.; Grenet, J.A.; Lin, W.; Finkelstein, D.; Vrijens, K.; Chen, T.; Guy, R.K.; et al. Ventromorphins: A New Class of Small Molecule Activators of the Canonical BMP Signaling Pathway. ACS Chem. Biol. 2017, 12, 2436–2447. [Google Scholar] [CrossRef]
- Little, S.C.; Mullins, M.C. Extracellular modulation of BMP activity in patterning the dorsoventral axis. Birth Defects Res. C Embryo Today 2006, 78, 224–242. [Google Scholar] [CrossRef]
- Stickney, H.L.; Imai, Y.; Draper, B.; Moens, C.; Talbot, W.S. Zebrafish bmp4 functions during late gastrulation to specify ventroposterior cell fates. Dev. Biol. 2007, 310, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Friedle, H.; Knochel, W. Cooperative interaction of Xvent-2 and GATA-2 in the activation of the ventral homeobox gene Xvent-1B. J. Biol. Chem. 2002, 277, 23872–23881. [Google Scholar] [CrossRef] [PubMed]
- Messenger, N.J.; Kabitschke, C.; Andrews, R.; Grimmer, D.; Nunez Miguel, R.; Blundell, T.L.; Smith, J.C.; Wardle, F.C. Functional specificity of the Xenopus T-domain protein Brachyury is conferred by its ability to interact with Smad1. Dev. Cell 2005, 8, 599–610. [Google Scholar] [CrossRef] [PubMed]
- Onichtchouk, D.; Gawantka, V.; Dosch, R.; Delius, H.; Hirschfeld, K.; Blumenstock, C.; Niehrs, C. The Xvent-2 homeobox gene is part of the BMP-4 signalling pathway controlling [correction of controling] dorsoventral patterning of Xenopus mesoderm. Development 1996, 122, 3045–3053. [Google Scholar] [CrossRef] [PubMed]
- Schuler-Metz, A.; Knochel, S.; Kaufmann, E.; Knochel, W. The homeodomain transcription factor Xvent-2 mediates autocatalytic regulation of BMP-4 expression in Xenopus embryos. J. Biol. Chem. 2000, 275, 34365–34374. [Google Scholar] [CrossRef] [PubMed]
- Trindade, M.; Tada, M.; Smith, J.C. DNA-binding specificity and embryological function of Xom (Xvent-2). Dev. Biol. 1999, 216, 442–456. [Google Scholar] [CrossRef]
- Ladher, R.; Mohun, T.J.; Smith, J.C.; Snape, A.M. Xom: A Xenopus homeobox gene that mediates the early effects of BMP-4. Development 1996, 122, 2385–2394. [Google Scholar] [CrossRef]
- Papalopulu, N.; Kintner, C. A Xenopus gene, Xbr-1, defines a novel class of homeobox genes and is expressed in the dorsal ciliary margin of the eye. Dev. Biol. 1996, 174, 104–114. [Google Scholar] [CrossRef]
- Zhong, Y.F.; Holland, P.W. The dynamics of vertebrate homeobox gene evolution: Gain and loss of genes in mouse and human lineages. BMC Evol. Biol. 2011, 11, 169. [Google Scholar] [CrossRef]
- Ault, K.T.; Dirksen, M.L.; Jamrich, M. A novel homeobox gene PV.1 mediates induction of ventral mesoderm in Xenopus embryos. Proc. Natl. Acad. Sci. USA 1996, 93, 6415–6420. [Google Scholar] [CrossRef]
- Dale, L.; Wardle, F.C. A gradient of BMP activity specifies dorsal-ventral fates in early Xenopus embryos. Semin. Cell Dev. Biol. 1999, 10, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Gentner, E.; Vegi, N.M.; Mulaw, M.A.; Mandal, T.; Bamezai, S.; Claus, R.; Tasdogan, A.; Quintanilla-Martinez, L.; Grunenberg, A.; Dohner, K.; et al. VENTX induces expansion of primitive erythroid cells and contributes to the development of acute myeloid leukemia in mice. Oncotarget 2016, 7, 86889–86901. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.S.; Seo, J.J.; Cha, S.W.; Lee, H.S.; Lee, S.Y.; Roh, D.H.; Kung Hf, H.F.; Kim, J.; Ja Park, M. Antimorphic PV.1 causes secondary axis by inducing ectopic organizer. Biochem. Biophys. Res. Commun. 2002, 292, 1081–1086. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, A.; Wilm, T.; Solnica-Krezel, L.; Dawid, I.B. Antagonistic role of vega1 and bozozok/dharma homeobox genes in organizer formation. Proc. Natl. Acad. Sci. USA 2000, 97, 12121–12126. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Lim, S.K.; Cha, S.W.; Yoon, J.; Lee, S.H.; Lee, H.S.; Park, J.B.; Lee, J.Y.; Kim, S.C.; Kim, J. Inhibition of FGF signaling converts dorsal mesoderm to ventral mesoderm in early Xenopus embryos. Differentiation 2011, 82, 99–107. [Google Scholar] [CrossRef]
- Lee, S.Y.; Yoon, J.; Lee, M.H.; Jung, S.K.; Kim, D.J.; Bode, A.M.; Kim, J.; Dong, Z. The role of heterodimeric AP-1 protein comprised of JunD and c-Fos proteins in hematopoiesis. J. Biol. Chem. 2012, 287, 31342–31348. [Google Scholar] [CrossRef]
- Shapira, E.; Marom, K.; Yelin, R.; Levy, A.; Fainsod, A. A role for the homeobox gene Xvex-1 as part of the BMP-4 ventral signaling pathway. Mech. Dev. 1999, 86, 99–111. [Google Scholar] [CrossRef]
- Shimizu, T.; Yamanaka, Y.; Nojima, H.; Yabe, T.; Hibi, M.; Hirano, T. A novel repressor-type homeobox gene, ved, is involved in dharma/bozozok-mediated dorsal organizer formation in zebrafish. Mech. Dev. 2002, 118, 125–138. [Google Scholar] [CrossRef]
- De Robertis, E.M.; Kuroda, H. Dorsal-ventral patterning and neural induction in Xenopus embryos. Annu. Rev. Cell Dev. Biol. 2004, 20, 285–308. [Google Scholar] [CrossRef]
- Hemmati-Brivanlou, A.; Thomsen, G.H. Ventral mesodermal patterning in Xenopus embryos: Expression patterns and activities of BMP-2 and BMP-4. Dev. Genet. 1995, 17, 78–89. [Google Scholar] [CrossRef]
- Maeno, M.; Mead, P.E.; Kelley, C.; Xu, R.H.; Kung, H.F.; Suzuki, A.; Ueno, N.; Zon, L.I. The role of BMP-4 and GATA-2 in the induction and differentiation of hematopoietic mesoderm in Xenopus laevis. Blood 1996, 88, 1965–1972. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.H.; Ault, K.T.; Kim, J.; Park, M.J.; Hwang, Y.S.; Peng, Y.; Sredni, D.; Kung, H. Opposite effects of FGF and BMP-4 on embryonic blood formation: Roles of PV.1 and GATA-2. Dev. Biol. 1999, 208, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Friedle, H.; Rastegar, S.; Paul, H.; Kaufmann, E.; Knochel, W. Xvent-1 mediates BMP-4-induced suppression of the dorsal-lip-specific early response gene XFD-1’ in Xenopus embryos. EMBO J. 1998, 17, 2298–2307. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Imai, Y.; Gates, M.A.; Melby, A.E.; Kimelman, D.; Schier, A.F.; Talbot, W.S. The homeobox genes vox and vent are redundant repressors of dorsal fates in zebrafish. Development 2001, 128, 2407–2420. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, A.; Wilm, T.; Solnica-Krezel, L.; Dawid, I.B. Functional interaction of vega2 and goosecoid homeobox genes in zebrafish. Genesis 2000, 28, 58–67. [Google Scholar] [CrossRef]
- Gawantka, V.; Delius, H.; Hirschfeld, K.; Blumenstock, C.; Niehrs, C. Antagonizing the Spemann organizer: Role of the homeobox gene Xvent-1. EMBO J. 1995, 14, 6268–6279. [Google Scholar] [CrossRef] [PubMed]
- Onichtchouk, D.; Glinka, A.; Niehrs, C. Requirement for Xvent-1 and Xvent-2 gene function in dorsoventral patterning of Xenopus mesoderm. Development 1998, 125, 1447–1456. [Google Scholar] [CrossRef]
- Scerbo, P.; Girardot, F.; Vivien, C.; Markov, G.V.; Luxardi, G.; Demeneix, B.; Kodjabachian, L.; Coen, L. Ventx factors function as Nanog-like guardians of developmental potential in Xenopus. PLoS ONE 2012, 7, e36855. [Google Scholar] [CrossRef]
- Scerbo, P.; Marchal, L.; Kodjabachian, L. Lineage commitment of embryonic cells involves MEK1-dependent clearance of pluripotency regulator Ventx2. eLife 2017, 6, e21526. [Google Scholar] [CrossRef]
- Wu, X.; Gao, H.; Bleday, R.; Zhu, Z. Homeobox transcription factor VentX regulates differentiation and maturation of human dendritic cells. J. Biol. Chem. 2014, 289, 14633–14643. [Google Scholar] [CrossRef]
- Wu, X.; Gao, H.; Ke, W.; Giese, R.W.; Zhu, Z. The homeobox transcription factor VentX controls human macrophage terminal differentiation and proinflammatory activation. J. Clin. Investig. 2011, 121, 2599–2613. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gartel, A.L.; Radhakrishnan, S.K. Lost in transcription: p21 repression, mechanisms, and consequences. Cancer Res. 2005, 65, 3980–3985. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Zhang, P.; Harper, J.W.; Elledge, S.J.; Leder, P. Mice lacking p21CIP1/WAF1 undergo normal development, but are defective in G1 checkpoint control. Cell 1995, 82, 675–684. [Google Scholar] [CrossRef]
- Wu, X.; Gao, H.; Ke, W.; Hager, M.; Xiao, S.; Freeman, M.R.; Zhu, Z. VentX trans-activates p53 and p16ink4a to regulate cellular senescence. J. Biol. Chem. 2011, 286, 12693–12701. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M.; Evans, B.J.; Bogart, J.P. Polyploidy in Amphibia. Cytogenet. Genome Res. 2015, 145, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Becak, M.L. Polyploidy and epigenetic events in the evolution of Anura. Genet. Mol. Res. 2014, 13, 5995–6014. [Google Scholar] [CrossRef]
- Burgess, S. Genomics: A matched set of frog sequences. Nature 2016, 538, 320–321. [Google Scholar] [CrossRef][Green Version]
- Xie, J.; Nachabe, A.; Hathaway, L.J.; Farah, B.; Berbari, B.; Li, Y.; Brown, T.C.; Schmid, J.L.; Socola, F.; Saba, N.S.; et al. The prognostic implications of tetraploidy/near-Tetraploidy in acute myeloid leukemia: A case series and systematic review of the literature. Leuk Lymphoma 2021, 62, 203–210. [Google Scholar] [CrossRef]
- Singh, M.D.; Jensen, M.; Lasser, M.; Huber, E.; Yusuff, T.; Pizzo, L.; Lifschutz, B.; Desai, I.; Kubina, A.; Yennawar, S.; et al. NCBP2 modulates neurodevelopmental defects of the 3q29 deletion in Drosophila and Xenopus laevis models. PLoS Genet. 2020, 16, e1008590. [Google Scholar] [CrossRef]
- Yaguchi, K.; Yamamoto, T.; Matsui, R.; Shimada, M.; Shibanuma, A.; Kamimura, K.; Koda, T.; Uehara, R. Tetraploidy-associated centrosome overduplication in mouse early embryos. Commun. Integr. Biol. 2018, 11, e1526605. [Google Scholar] [CrossRef]
- Tanaka, K.; Goto, H.; Nishimura, Y.; Kasahara, K.; Mizoguchi, A.; Inagaki, M. Tetraploidy in cancer and its possible link to aging. Cancer Sci. 2018, 109, 2632–2640. [Google Scholar] [CrossRef] [PubMed]
- Frade, J.M.; Lopez-Sanchez, N. Neuronal tetraploidy in Alzheimer and aging. Aging 2017, 9, 2014–2015. [Google Scholar] [CrossRef] [PubMed]
- Tandon, P.; Conlon, F.; Furlow, J.D.; Horb, M.E. Expanding the genetic toolkit in Xenopus: Approaches and opportunities for human disease modeling. Dev. Biol. 2017, 426, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Jonsdottir, A.B.; Stefansson, O.A.; Bjornsson, J.; Jonasson, J.G.; Ogmundsdottir, H.M.; Eyfjord, J.E. Tetraploidy in BRCA2 breast tumours. Eur. J. Cancer 2012, 48, 305–310. [Google Scholar] [CrossRef]
- Takeuchi, M.; Takeuchi, K.; Ozawa, Y.; Kohara, A.; Mizusawa, H. Aneuploidy in immortalized human mesenchymal stem cells with non-random loss of chromosome 13 in culture. Vitr. Cell Dev. Biol. Anim. 2009, 45, 290–299. [Google Scholar] [CrossRef]
- Nguyen, H.G.; Ravid, K. Tetraploidy/aneuploidy and stem cells in cancer promotion: The role of chromosome passenger proteins. J. Cell Physiol. 2006, 208, 12–22. [Google Scholar] [CrossRef]
- Burglin, T.R. The PBC domain contains a MEINOX domain: Coevolution of Hox and TALE homeobox genes? Dev. Genes. Evol. 1998, 208, 113–116. [Google Scholar] [CrossRef]
- Yoon, J.; Kim, J.H.; Lee, S.Y.; Kim, S.; Park, J.B.; Lee, J.Y.; Kim, J. PV.1 induced by FGF-Xbra functions as a repressor of neurogenesis in Xenopus embryos. BMB Rep. 2014, 47, 673. [Google Scholar] [CrossRef]
- Zhong, Y.F.; Holland, P.W. HomeoDB2: Functional expansion of a comparative homeobox gene database for evolutionary developmental biology. Evol. Dev. 2011, 13, 567–568. [Google Scholar] [CrossRef]
- Zhong, Y.F.; Butts, T.; Holland, P.W. HomeoDB: A database of homeobox gene diversity. Evol. Dev. 2008, 10, 516–518. [Google Scholar] [CrossRef]
- Ault, K.T.; Xu, R.H.; Kung, H.F.; Jamrich, M. The homeobox gene PV.1 mediates specification of the prospective neural ectoderm in Xenopus embryos. Dev. Biol. 1997, 192, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.S.; Lee, H.S.; Roh, D.H.; Cha, S.; Lee, S.Y.; Seo, J.J.; Kim, J.; Park, M.J. Active repression of organizer genes by C-terminal domain of PV.1. Biochem. Biophys. Res. Commun. 2003, 308, 79–86. [Google Scholar] [CrossRef]
- Kumar, V.; Park, S.; Lee, U.; Kim, J. The Organizer and Its Signaling in Embryonic Development. J. Dev. Biol. 2021, 9, 47. [Google Scholar] [CrossRef]
- Kumar, V.; Goutam, R.S.; Park, S.; Lee, U.; Kim, J. Functional Roles of FGF Signaling in Early Development of Vertebrate Embryos. Cells 2021, 10, 2148. [Google Scholar] [CrossRef]
- Kumar, V.; Umair, Z.; Kumar, S.; Lee, U.; Kim, J. Smad2 and Smad3 differentially modulate chordin transcription via direct binding on the distal elements in gastrula Xenopus embryos. Biochem. Biophys. Res. Commun. 2021, 559, 168–175. [Google Scholar] [CrossRef]
- Umair, Z.; Kumar, S.; Kim, D.H.; Rafiq, K.; Kumar, V.; Kim, S.; Park, J.B.; Lee, J.Y.; Lee, U.; Kim, J. Ventx1.1 as a Direct Repressor of Early Neural Gene zic3 in Xenopus laevis. Mol. Cells 2018, 41, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.; Kim, J.H.; Kim, S.C.; Park, J.B.; Lee, J.Y.; Kim, J. PV.1 suppresses the expression of FoxD5b during neural induction in Xenopus embryos. Mol. Cells 2014, 37, 220–225. [Google Scholar] [CrossRef]
- Kumar, V.; Goutam, R.S.; Umair, Z.; Park, S.; Lee, U.; Kim, J. Foxd4l1.1 Negatively Regulates Chordin Transcription in Neuroectoderm of Xenopus Gastrula. Cells 2021, 10, 2779. [Google Scholar] [CrossRef]
- Kumar, S.; Umair, Z.; Kumar, V.; Kumar, S.; Lee, U.; Kim, J. Foxd4l1.1 negatively regulates transcription of neural repressor ventx1.1 during neuroectoderm formation in Xenopus embryos. Sci. Rep. 2020, 10, 16780. [Google Scholar] [CrossRef]
- Kumar, S.; Umair, Z.; Kumar, V.; Lee, U.; Choi, S.C.; Kim, J. Ventx1.1 competes with a transcriptional activator Xcad2 to regulate negatively its own expression. BMB Rep. 2019, 52, 403–408. [Google Scholar] [CrossRef]
- Pillemer, G.; Yelin, R.; Epstein, M.; Gont, L.; Frumkin, Y.; Yisraeli, J.K.; Steinbeisser, H.; Fainsod, A. The Xcad-2 gene can provide a ventral signal independent of BMP-4. Mech. Dev. 1998, 74, 133–143. [Google Scholar] [CrossRef]
- Kumano, G.; Belluzzi, L.; Smith, W.C. Spatial and temporal properties of ventral blood island induction in Xenopus laevis. Development 1999, 126, 5327–5337. [Google Scholar] [CrossRef] [PubMed]
- Sander, V.; Reversade, B.; De Robertis, E.M. The opposing homeobox genes Goosecoid and Vent1/2 self-regulate Xenopus patterning. EMBO J. 2007, 26, 2955–2965. [Google Scholar] [CrossRef] [PubMed]
- Rogers, C.D.; Archer, T.C.; Cunningham, D.D.; Grammer, T.C.; Casey, E.M. Sox3 expression is maintained by FGF signaling and restricted to the neural plate by Vent proteins in the Xenopus embryo. Dev. Biol. 2008, 313, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, A.; Ishii, K.; Yamashita, S.; Nejigane, S.; Matsukawa, S.; Ito, Y.; Onuma, Y.; Asashima, M.; Michiue, T. mNanog possesses dorsal mesoderm-inducing ability by modulating both BMP and Activin/nodal signaling in Xenopus ectodermal cells. PLoS ONE 2012, 7, e46630. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Kirschner, M. Regulated proteolysis of Xom mediates dorsoventral pattern formation during early Xenopus development. Dev. Cell 2002, 3, 557–568. [Google Scholar] [CrossRef]
- Gao, H.; Wu, B.; Giese, R.; Zhu, Z. Xom interacts with and stimulates transcriptional activity of LEF1/TCFs: Implications for ventral cell fate determination during vertebrate embryogenesis. Cell Res. 2007, 17, 345–356. [Google Scholar] [CrossRef]
- Wroble, B.N.; Finkielstein, C.V.; Sible, J.C. Wee1 kinase alters cyclin E/Cdk2 and promotes apoptosis during the early embryonic development of Xenopus laevis. BMC Dev. Biol. 2007, 7, 119. [Google Scholar] [CrossRef]
- Trindade, M.; Messenger, N.; Papin, C.; Grimmer, D.; Fairclough, L.; Tada, M.; Smith, J.C. Regulation of apoptosis in the Xenopus embryo by Bix3. Development 2003, 130, 4611–4622. [Google Scholar] [CrossRef]
- Wu, B.; Gao, H.; Le, Y.; Wu, X.; Zhu, Z. Xom induces proteolysis of beta-catenin through GSK3beta-mediated pathway. FEBS Lett. 2018, 592, 299–309. [Google Scholar] [CrossRef]
- Schuff, M.; Siegel, D.; Philipp, M.; Bundschu, K.; Heymann, N.; Donow, C.; Knochel, W. Characterization of Danio rerio Nanog and functional comparison to Xenopus Vents. Stem. Cells Dev. 2012, 21, 1225–1238. [Google Scholar] [CrossRef] [PubMed]
- Chambers, I.; Silva, J.; Colby, D.; Nichols, J.; Nijmeijer, B.; Robertson, M.; Vrana, J.; Jones, K.; Grotewold, L.; Smith, A. Nanog safeguards pluripotency and mediates germline development. Nature 2007, 450, 1230–1234. [Google Scholar] [CrossRef] [PubMed]
- Scerbo, P.; Monsoro-Burq, A.H. The vertebrate-specific VENTX/NANOG gene empowers neural crest with ectomesenchyme potential. Sci. Adv. 2020, 6, eaaz1469. [Google Scholar] [CrossRef] [PubMed]
- Shakhova, O.; Sommer, L. Neural crest-derived stem cells. In StemBook; Harvard Stem Cell Institute: Cambridge, MA, USA, 2008. [Google Scholar] [CrossRef]
- Fraser, G.J.; Hulsey, C.D.; Bloomquist, R.F.; Uyesugi, K.; Manley, N.R.; Streelman, J.T. An ancient gene network is co-opted for teeth on old and new jaws. PLoS Biol. 2009, 7, e31. [Google Scholar] [CrossRef]
- Miletich, I.; Yu, W.Y.; Zhang, R.; Yang, K.; Caixeta de Andrade, S.; Pereira, S.F.; Ohazama, A.; Mock, O.B.; Buchner, G.; Sealby, J.; et al. Developmental stalling and organ-autonomous regulation of morphogenesis. Proc. Natl. Acad. Sci. USA 2011, 108, 19270–19275. [Google Scholar] [CrossRef]
- Nichols, J.T.; Pan, L.; Moens, C.B.; Kimmel, C.B. barx1 represses joints and promotes cartilage in the craniofacial skeleton. Development 2013, 140, 2765–2775. [Google Scholar] [CrossRef]
- Sperber, S.M.; Dawid, I.B. barx1 is necessary for ectomesenchyme proliferation and osteochondroprogenitor condensation in the zebrafish pharyngeal arches. Dev. Biol. 2008, 321, 101–110. [Google Scholar] [CrossRef]
- Gao, H.; Le, Y.; Wu, X.; Silberstein, L.E.; Giese, R.W.; Zhu, Z. VentX, a novel lymphoid-enhancing factor/T-cell factor-associated transcription repressor, is a putative tumor suppressor. Cancer Res. 2010, 70, 202–211. [Google Scholar] [CrossRef][Green Version]
- Kim, B.M.; Buchner, G.; Miletich, I.; Sharpe, P.T.; Shivdasani, R.A. The stomach mesenchymal transcription factor Barx1 specifies gastric epithelial identity through inhibition of transient Wnt signaling. Dev. Cell 2005, 8, 611–622. [Google Scholar] [CrossRef]
- Kim, B.M.; Miletich, I.; Mao, J.; McMahon, A.P.; Sharpe, P.A.; Shivdasani, R.A. Independent functions and mechanisms for homeobox gene Barx1 in patterning mouse stomach and spleen. Development 2007, 134, 3603–3613. [Google Scholar] [CrossRef]
- Shapira, E.; Marom, K.; Levy, V.; Yelin, R.; Fainsod, A. The Xvex-1 antimorph reveals the temporal competence for organizer formation and an early role for ventral homeobox genes. Mech. Dev. 2000, 90, 77–87. [Google Scholar] [CrossRef]
- Gilardelli, C.N.; Pozzoli, O.; Sordino, P.; Matassi, G.; Cotelli, F. Functional and hierarchical interactions among zebrafish vox/vent homeobox genes. Dev. Dyn. 2004, 230, 494–508. [Google Scholar] [CrossRef] [PubMed]
- Melby, A.E.; Clements, W.K.; Kimelman, D. Regulation of dorsal gene expression in Xenopus by the ventralizing homeodomain gene Vox. Dev. Biol. 1999, 211, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Liu, C. Xom, a ventralizing factor, regulates beta-catenin levels and cell fate. FEBS Lett. 2018, 592, 297–298. [Google Scholar] [CrossRef]
- Leibovich, A.; Kot-Leibovich, H.; Ben-Zvi, D.; Fainsod, A. ADMP controls the size of Spemann’s organizer through a network of self-regulating expansion-restriction signals. BMC Biol. 2018, 16, 13. [Google Scholar] [CrossRef]
- Watanabe, M.; Yasuoka, Y.; Mawaribuchi, S.; Kuretani, A.; Ito, M.; Kondo, M.; Ochi, H.; Ogino, H.; Fukui, A.; Taira, M.; et al. Conservatism and variability of gene expression profiles among homeologous transcription factors in Xenopus laevis. Dev. Biol. 2017, 426, 301–324. [Google Scholar] [CrossRef]
- Trompouki, E.; Bowman, T.V.; Lawton, L.N.; Fan, Z.P.; Wu, D.C.; DiBiase, A.; Martin, C.S.; Cech, J.N.; Sessa, A.K.; Leblanc, J.L.; et al. Lineage regulators direct BMP and Wnt pathways to cell-specific programs during differentiation and regeneration. Cell 2011, 147, 577–589. [Google Scholar] [CrossRef]
- Le, Y.; Gao, H.; Bleday, R.; Zhu, Z. The homeobox protein VentX reverts immune suppression in the tumor microenvironment. Nat. Commun. 2018, 9, 2175. [Google Scholar] [CrossRef]
- Le, Y.; Gao, H.; Richards, W.; Zhao, L.; Bleday, R.; Clancy, T.; Zhu, Z. VentX expression in tumor-associated macrophages promotes phagocytosis and immunity against pancreatic cancers. JCI Insight 2020, 5, e137088. [Google Scholar] [CrossRef]
- Ino, Y.; Yamazaki-Itoh, R.; Shimada, K.; Iwasaki, M.; Kosuge, T.; Kanai, Y.; Hiraoka, N. Immune cell infiltration as an indicator of the immune microenvironment of pancreatic cancer. Br. J. Cancer 2013, 108, 914–923. [Google Scholar] [CrossRef]
- Gao, H.; Wu, X.; Sun, Y.; Zhou, S.; Silberstein, L.E.; Zhu, Z. Suppression of homeobox transcription factor VentX promotes expansion of human hematopoietic stem/multipotent progenitor cells. J. Biol. Chem. 2012, 287, 29979–29987. [Google Scholar] [CrossRef] [PubMed]
- Elagib, K.E.; Goldfarb, A.N. Oncogenic pathways of AML1-ETO in acute myeloid leukemia: Multifaceted manipulation of marrow maturation. Cancer Lett. 2007, 251, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Vargas, A.C.; Gray, L.A.; White, C.L.; Maclean, F.M.; Grimison, P.; Ardakani, N.M.; Bonar, F.; Algar, E.M.; Cheah, A.L.; Russell, P.; et al. Genome wide methylation profiling of selected matched soft tissue sarcomas identifies methylation changes in metastatic and recurrent disease. Sci. Rep. 2021, 11, 667. [Google Scholar] [CrossRef]
- Gao, H.; Wu, B.; Le, Y.; Zhu, Z. Homeobox protein VentX induces p53-independent apoptosis in cancer cells. Oncotarget 2016, 7, 39719–39729. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Grubelnik, G.; Bostjancic, E.; Pavlic, A.; Kos, M.; Zidar, N. NANOG expression in human development and cancerogenesis. Exp Biol. Med. 2020, 245, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Nagel, S.; Scherr, M.; MacLeod, R.A.F.; Pommerenke, C.; Koeppel, M.; Meyer, C.; Kaufmann, M.; Dallmann, I.; Drexler, H.G. NKL homeobox gene activities in normal and malignant myeloid cells. PLoS ONE 2019, 14, e0226212. [Google Scholar] [CrossRef] [PubMed]
- Brandner, S. Nanog, Gli, and p53: A new network of stemness in development and cancer. EMBO J. 2010, 29, 2475–2476. [Google Scholar] [CrossRef]
- Zbinden, M.; Duquet, A.; Lorente-Trigos, A.; Ngwabyt, S.N.; Borges, I.; Ruiz i Altaba, A. NANOG regulates glioma stem cells and is essential in vivo acting in a cross-functional network with GLI1 and p53. EMBO J. 2010, 29, 2659–2674. [Google Scholar] [CrossRef]
- Wang, S.; Fan, H.; Xu, J.; Zhao, E. Prognostic implication of NOTCH1 in early stage oral squamous cell cancer with occult metastases. Clin. Oral Investig. 2018, 22, 1131–1138. [Google Scholar] [CrossRef]
- Schier, A.F. Axis formation and patterning in zebrafish. Curr. Opin. Genet. Dev. 2001, 11, 393–404. [Google Scholar] [CrossRef]
- Mimoto, M.S.; Kwon, S.; Green, Y.S.; Goldman, D.; Christian, J.L. GATA2 regulates Wnt signaling to promote primitive red blood cell fate. Dev. Biol. 2015, 407, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kelley, C.; Yee, K.; Harland, R.; Zon, L.I. Ventral expression of GATA-1 and GATA-2 in the Xenopus embryo defines induction of hematopoietic mesoderm. Dev. Biol. 1994, 165, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Sun, H.; Cao, Y.; Xuan, B.; Fan, Y.; Sheng, H.; Zhuang, W. GATA2 Inhibition Sensitizes Acute Myeloid Leukemia Cells to Chemotherapy. PLoS ONE 2017, 12, e0170630. [Google Scholar] [CrossRef]
- Saida, S.; Zhen, T.; Kim, E.; Yu, K.; Lopez, G.; McReynolds, L.J.; Liu, P.P. Gata2 deficiency delays leukemogenesis while contributing to aggressive leukemia phenotype in Cbfb-MYH11 knockin mice. Leukemia 2020, 34, 759–770. [Google Scholar] [CrossRef] [PubMed]
- Luesink, M.; Hollink, I.H.; van der Velden, V.H.; Knops, R.H.; Boezeman, J.B.; de Haas, V.; Trka, J.; Baruchel, A.; Reinhardt, D.; van der Reijden, B.A.; et al. High GATA2 expression is a poor prognostic marker in pediatric acute myeloid leukemia. Blood 2012, 120, 2064–2075. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.S.; Chae, J.P.; Kim, D.S.; Park, K.M.; Bae, Y.C.; Park, M.J. Screening of Interacting Proteins with PV. 1 as Downstream Factors of BMP Signal. Korean J. Anat. 2007, 40, 203–210. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Superclass | Class | Subclass | AA Position in HD | AA in HD | |||
|---|---|---|---|---|---|---|---|
| ANTP | HoxL | N = 5 | N + 1 | X = 50 | X + 1 | ||
| Arginine | - | Glutamine | Lysine | 60 | |||
| NKL | Arginine | Glutamine | Glutamine | - | 60 | ||
| PRD | Pax | Arginine | - | Serine | - | 60 | |
| PRD-like | PaxL | Arginine | - | Glutamine | (X + 3) Lysine | 60 | |
| LIM | Arginine | - | Glutamine | - | 60 | ||
| ZF | Arginine | - | Glutamine | Arginine | 60 | ||
| POU | Arginine | - | Cysteine | - | 60 | ||
| HNF | Arginine | - | Alanine | - | 60 | ||
| CUT | Cux | Arginine | - | Histidine | - | 60 | |
| ONECUT | Arginine | - | Methionine | - | 60 | ||
| CMP | Arginine | - | Lysine | 60 | |||
| SATB | Arginine | - | Glutamine | - | 60 | ||
| PROS | Serine | - | Serine | - | 60 | ||
| CERS | Serine | - | Arginine | 60 | |||
| SIX/SO | Serine, threonine, valine | - | lysine | - | 60 | ||
| TALE | PBC | Arginine | - | Glycine | - | 63 | |
| MEIS | Arginine | - | Isoleucine | - | 63 | ||
| TGIF | Arginine | - | Isoleucine | - | 63 | ||
| IRO | Arginine | - | Alanine | - | 63 | ||
| MKX | Lysine | - | Alanine | - | 63 | ||
| Homeobox Class | Subclass | Number of Families of Different Classes | Number of Genes | References | ||||
|---|---|---|---|---|---|---|---|---|
| Human | Xenopus | Zebrafish | Human | Xenopus | Zebrafish | [68,69] http://homeodb.zoo.ox.ac.uk/ (accessed on 27 December 2021) | ||
| ANTP | HoxL | 14 | 14 | 14 | 52 | 53 | 68 | |
| NKL | 23 | 22 | 22 | 67 | 60 | 64 | ||
| PRD | Pax | 3 | 3 | 3 | 7 | 5 | 9 | |
| PRD-like | PaxL | 28 | 18 | 18 | 91 | 32 | 44 | |
| LIM | 6 | 6 | 6 | 12 | 12 | 20 | ||
| POU | 7 | 6 | 6 | 24 | 19 | 19 | ||
| HNF | 2 | 2 | 2 | 3 | 3 | 6 | ||
| SINE | 3 | 3 | 3 | 6 | 6 | 13 | ||
| TALE | 6 | 6 | 6 | 30 | 16 | 29 | ||
| CUT | 3 | 3 | 3 | 10 | 7 | 9 | ||
| PROS | 1 | 1 | 1 | 2 | 2 | 3 | ||
| ZF | 5 | 5 | 5 | 15 | 14 | 17 | ||
| CERS | 1 | 1 | 1 | 5 | 2 | 3 | ||
| Others | 3 | 3 | 1 | 9 | 19 | 18 | ||
| Total | 105 | 93 | 91 | 333 | 250 | 322 | ||
| Names | Chromosomal Localization | Similarity with Human Ventx (%) | Developmental Roles | References |
|---|---|---|---|---|
| Xenopus laevis | ||||
| Venxt1.1/PV.1/Ventx1.1a/Ventx1.1b | Chr7L:21,842,291–21,845,156 Chr7S:16,920,567–16,923,476 | 59.02 | Neural repressor, Spemann-organizer inhibitor, embryonic ventralization, hematopoiesis, axial patterning | [30,33,72] |
| Ventx1.2/Xvent1/Vent1/Vent-1/Xvent-1 | Chr7L:21,802,386–21,807,247 Chr7S:16,883,662–16,887,492 | 44.95 | Neural repressor, Spemann-organizer inhibitor, embryonic ventralization, axial patterning | [46,47,83] |
| Ventx2.1a/Ventx2.1b, Vox1, Vox, Xvent2 | Chr7L:21,859,318–21,861,104 Chr7S:16,932,131–16,933,936 | 66.67 | Neural repressor, Spemann-organizer inhibitor, embryonic ventralization, Ventx1.2 and ventx1.1 activator, hematopoiesis, axial patterning | [12,15,104] |
| Ventx2.2/Xom */Xvent-2/Xbr-1 | Chr7L:21,830,246–21,832,823 Chr7S:16,908,607–16,911,052 | 66.67 | Neural repressor, VMZ activator, induce GSK3β expression, axial patterning, pluripotency marker, neural crest migration | [27,28,87,90,105] |
| Ventx3.1/vent3 | Chr7L-NA Chr7S:16,889,505–16,895,419 | 39.1 | NA | [106,107] |
| Ventx3.2/Vex1 | Chr7L:21,784,572–21,787,885 Chr7S:16,870,294–16,873,640 | 51.32 | Neural repressor, VMZ activator, axial patterning | [37,102] |
| Zebrafish | ||||
| Vega1/Vox | Chr13:50,621,929–50,624,154 | 55 | Neural repressor, VMZ, activator, axial patterning | [34] |
| Vega2/Vent1 | Chr13:50,609,716–50,614,718 | 55 | Neural repressor, VMZ activator, axial patterning | [45] |
| Ved | Chr10:44,997,062–45,002,196 | 49.12 | Neural repressor, VMZ activator, axial patterning | [38] |
| Human | ||||
| Ventx | Chr10:133,237,855–133,241,928 | 100 | Acute myeloid leukemia, cellular senescence, myeloid and macrophage differentiation, dendritic cell differentiation, immunosuppressor, apoptotic activator | [17,18] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, S.; Kumar, V.; Li, W.; Kim, J. Ventx Family and Its Functional Similarities with Nanog: Involvement in Embryonic Development and Cancer Progression. Int. J. Mol. Sci. 2022, 23, 2741. https://doi.org/10.3390/ijms23052741
Kumar S, Kumar V, Li W, Kim J. Ventx Family and Its Functional Similarities with Nanog: Involvement in Embryonic Development and Cancer Progression. International Journal of Molecular Sciences. 2022; 23(5):2741. https://doi.org/10.3390/ijms23052741
Chicago/Turabian StyleKumar, Shiv, Vijay Kumar, Wenchang Li, and Jaebong Kim. 2022. "Ventx Family and Its Functional Similarities with Nanog: Involvement in Embryonic Development and Cancer Progression" International Journal of Molecular Sciences 23, no. 5: 2741. https://doi.org/10.3390/ijms23052741
APA StyleKumar, S., Kumar, V., Li, W., & Kim, J. (2022). Ventx Family and Its Functional Similarities with Nanog: Involvement in Embryonic Development and Cancer Progression. International Journal of Molecular Sciences, 23(5), 2741. https://doi.org/10.3390/ijms23052741