
Complex Effects of Sertraline and Citalopram on In Vitro Murine Breast Cancer Proliferation and on In Vivo Progression and Anxiety Level



Abstract
:1. Introduction
- the comparative in vitro anti-proliferative effect of sertraline and citalopram on the murine breast cancer 4T1 cell-line, growth and survival,
- the in vivo interaction among SSRIs, stress and tumor progression in a mouse 4T1 breast cancer grafted model and ex-vivo splenocyte proliferation, and
- secretion of the pro-inflammatory cytokine TNF-α, in response to LPS stimulation, which may affect cancer progression.
2. Results
2.1. In Vitro—4T1 Cell-Line and Splenocytes
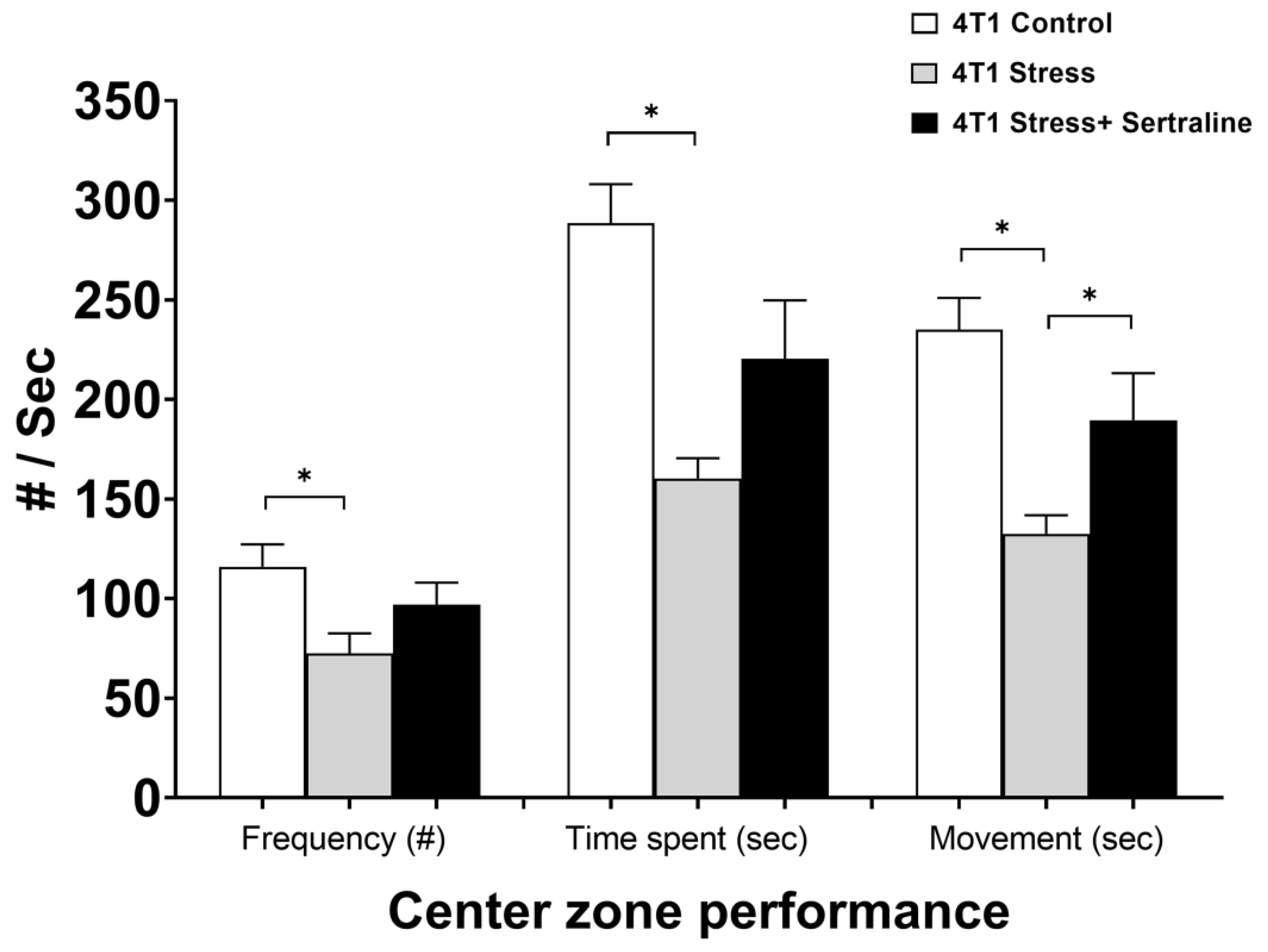
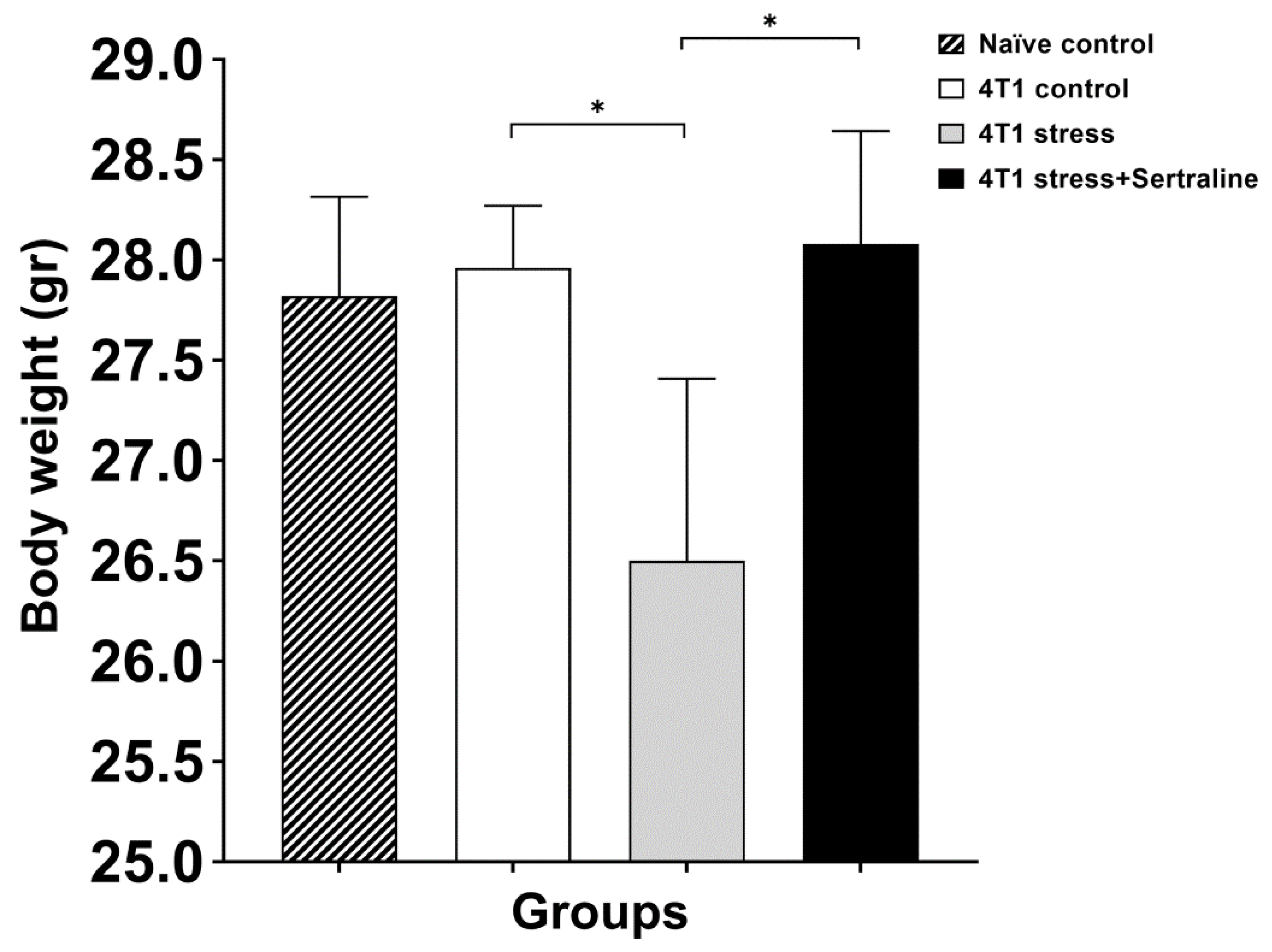
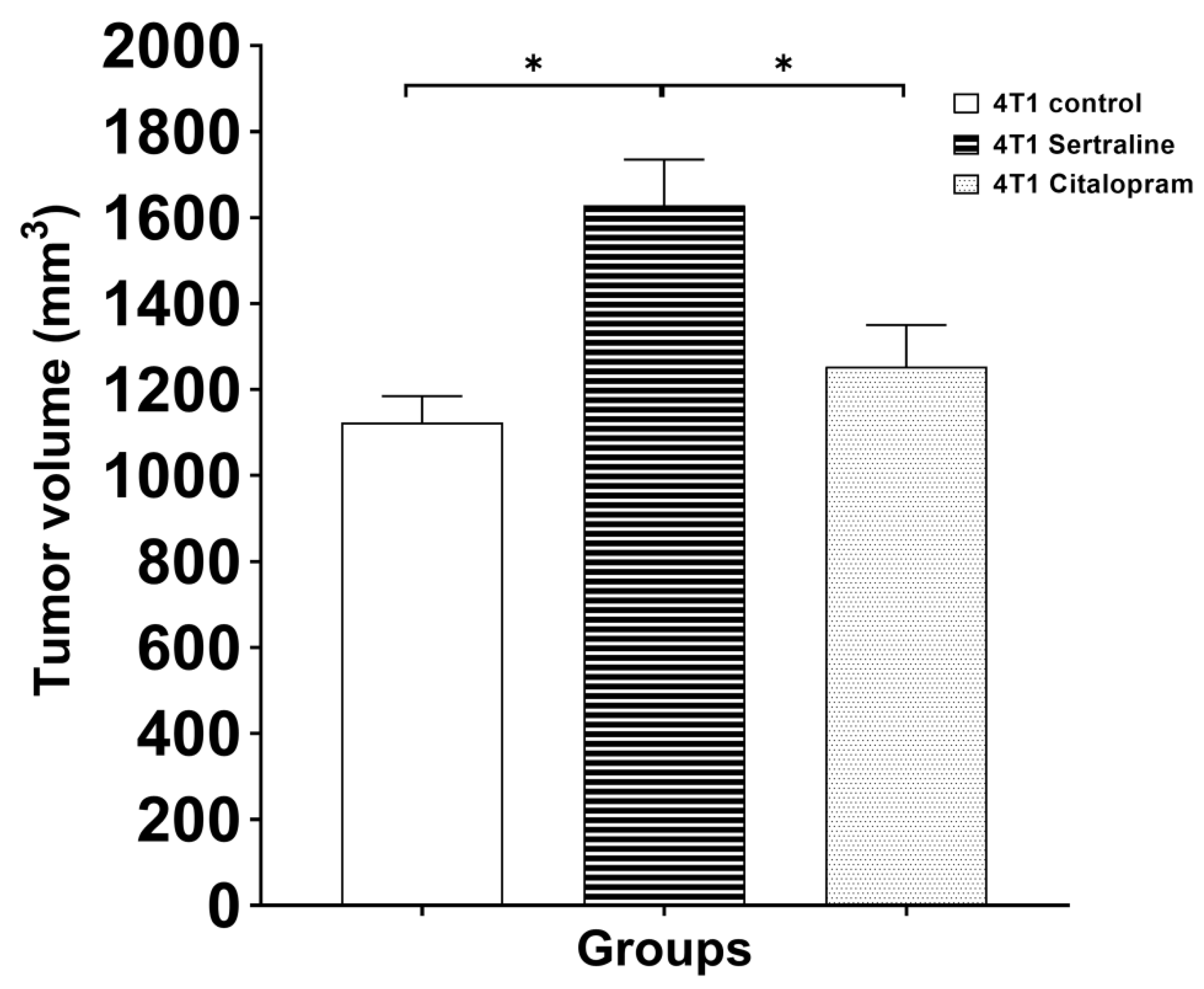
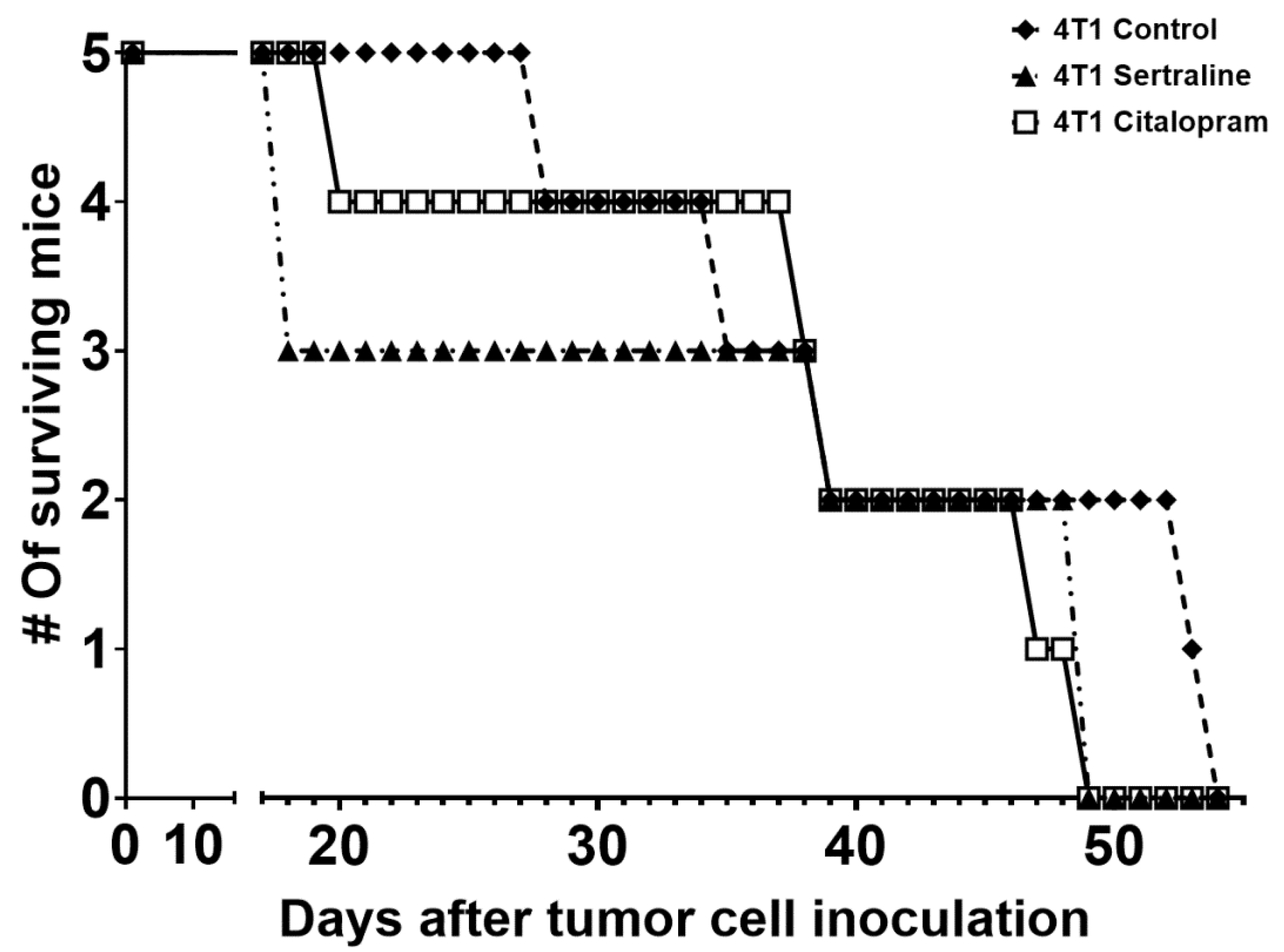
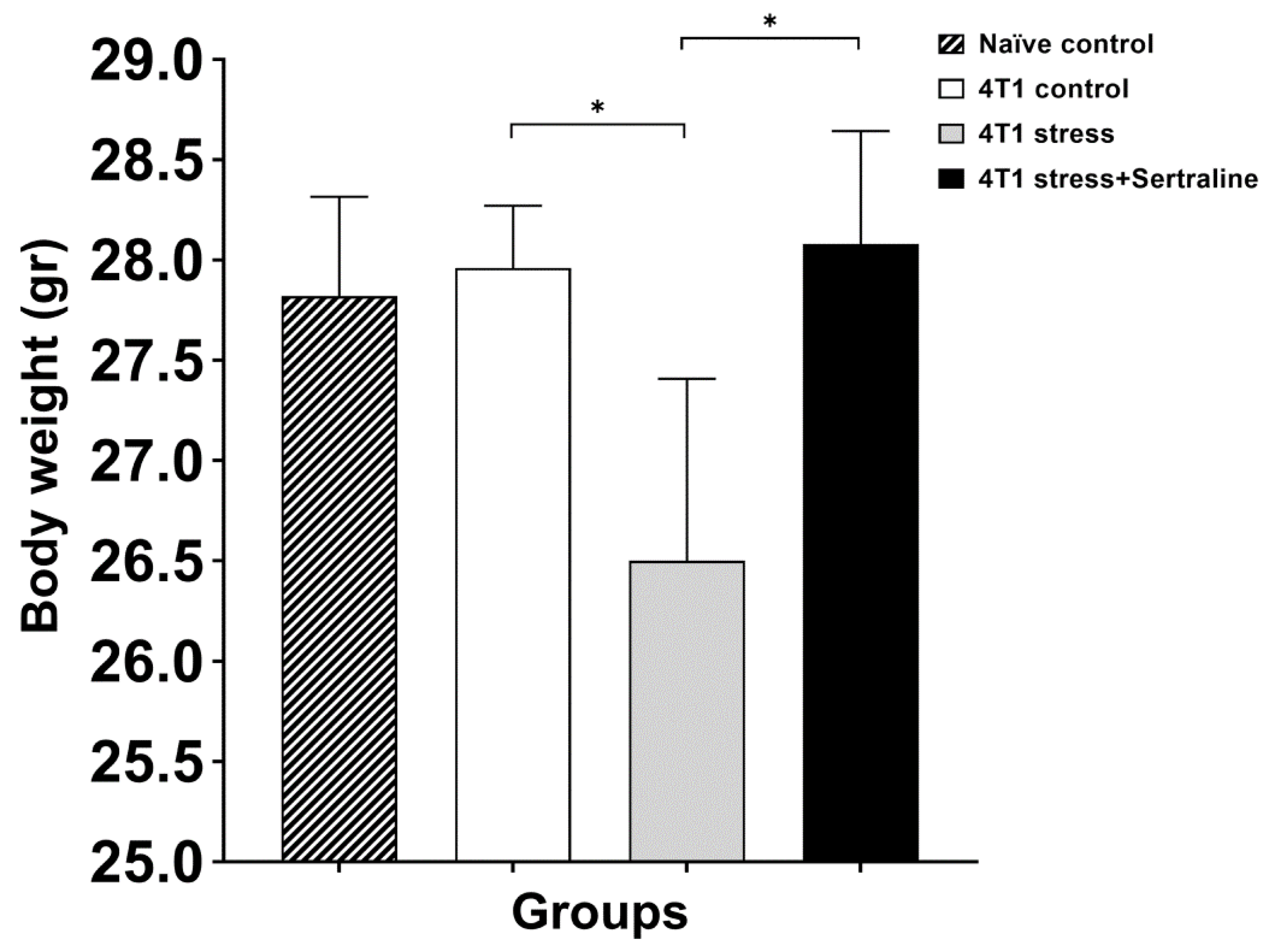
2.2. In Vivo—CMS
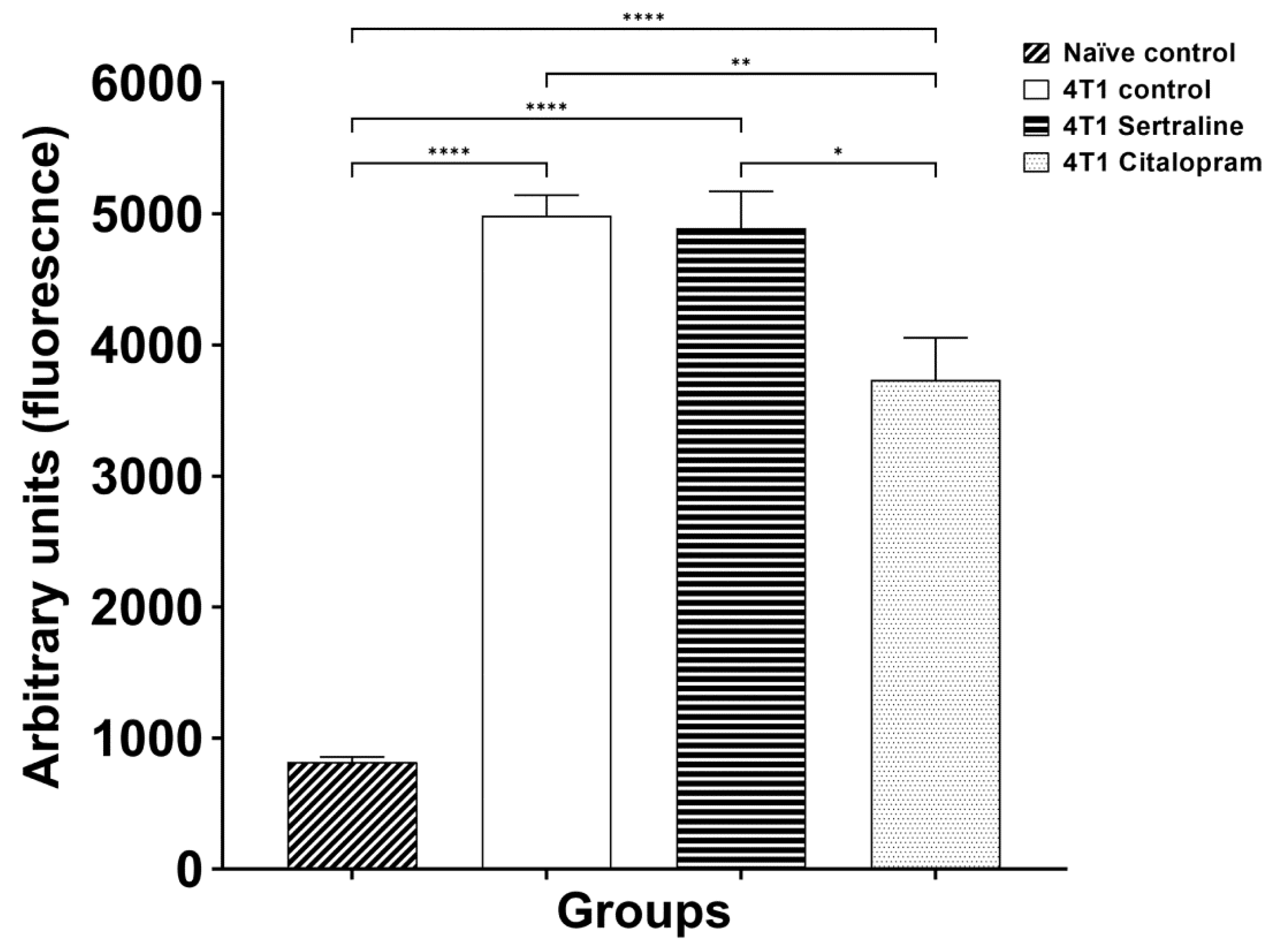
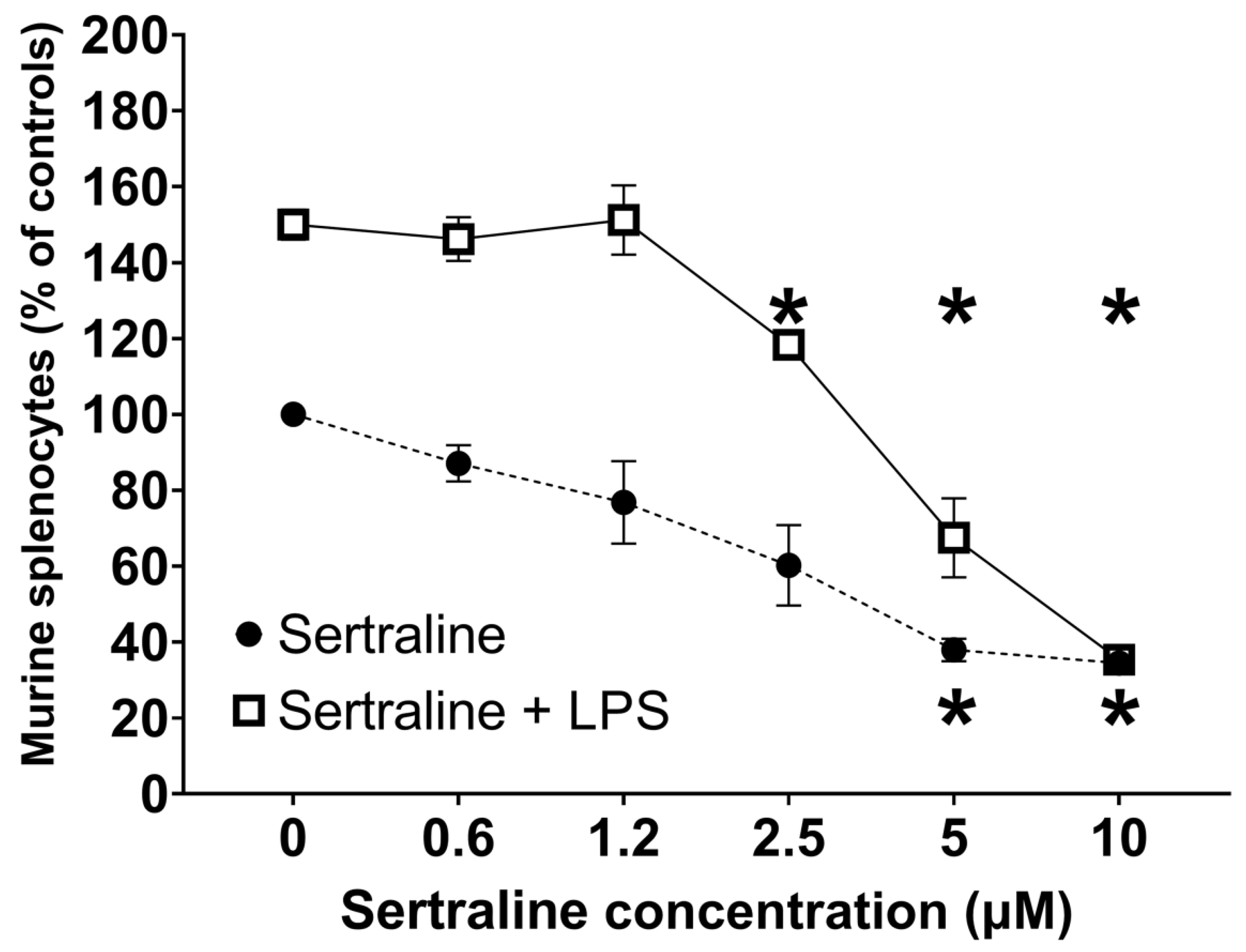
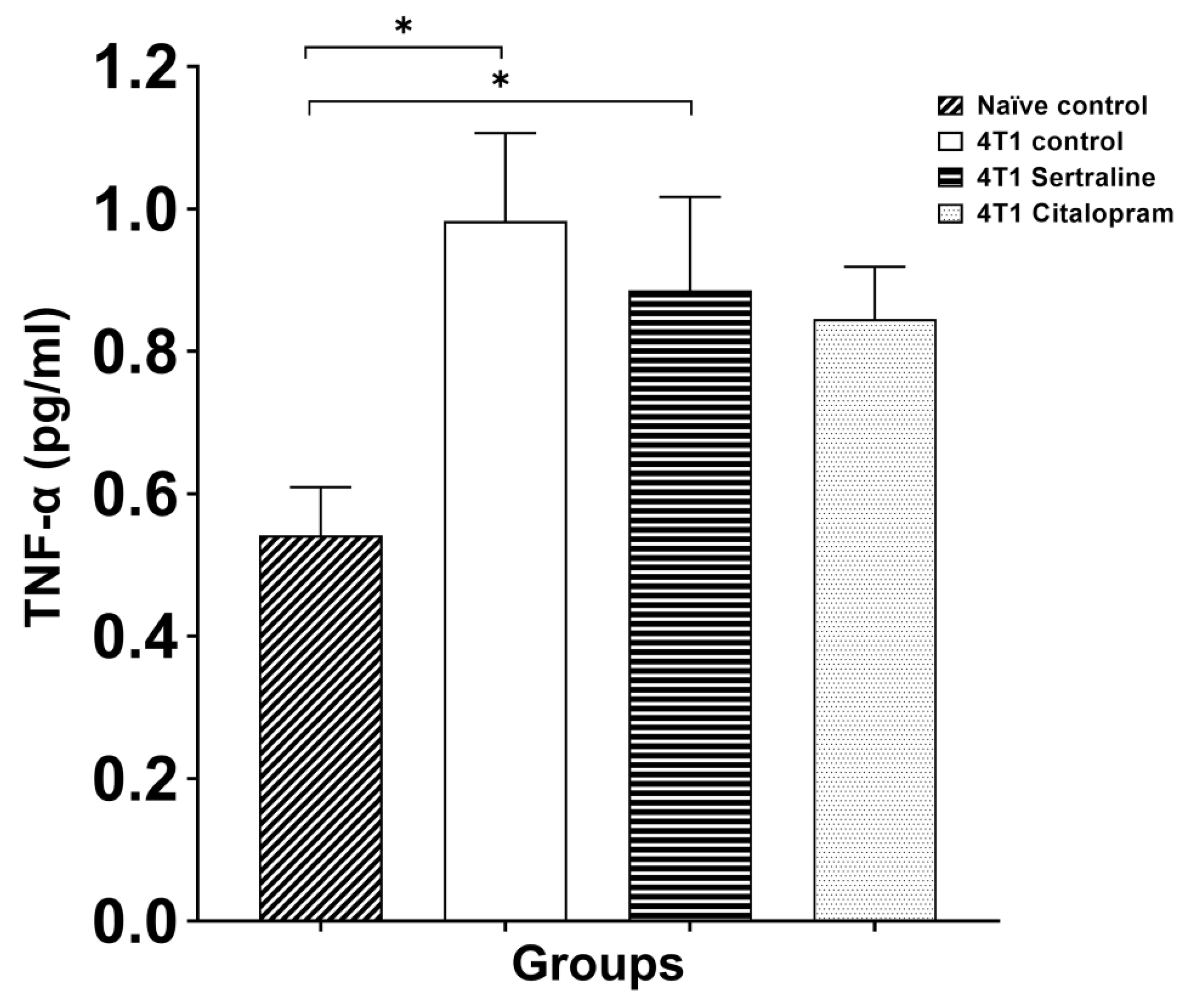
2.3. Ex Vivo—Splenocyte Proliferation
3. Discussion
4. Materials and Methods
4.1. Ethics
4.2. Chemicals and Reagents
4.3. Cell Line
4.4. Primary Splenocyte Cell Culture
4.5. Animals
4.5.1. Chronic Mild Stress
4.5.2. Behavioral Anxiety Model
T1 Murine Breast Cancer Model
4.6. Cell Viability Assay
4.7. In Vivo Experiments
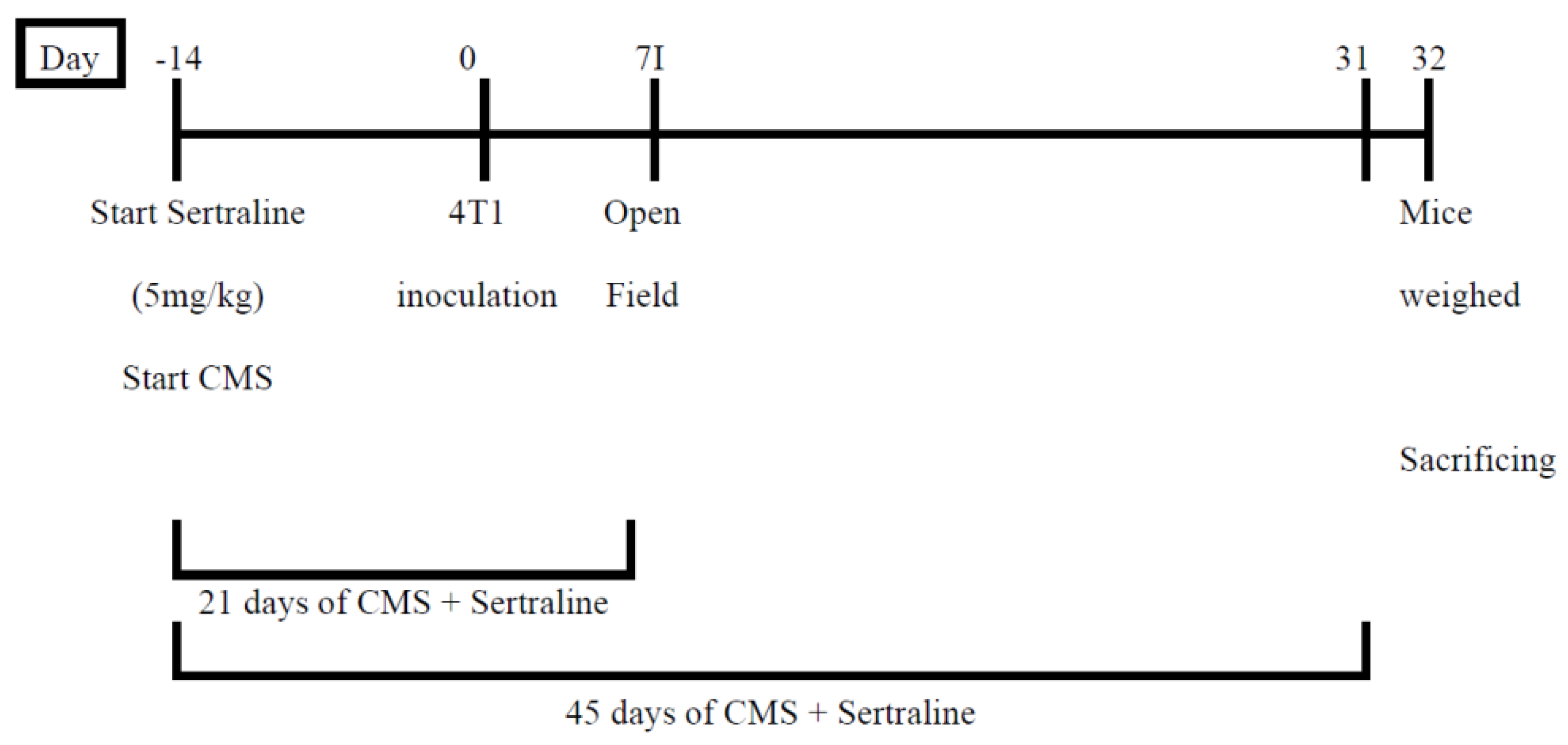
4.7.1. Experiment 1—Effects on Anxiety and Tumor Growth of Pre- (14 Days) and Short- (21 Days) or long- (45 Days) Term Treatment with Sertraline (5 mg/kg/d), in Mice Exposed to Chronic Mild Stress
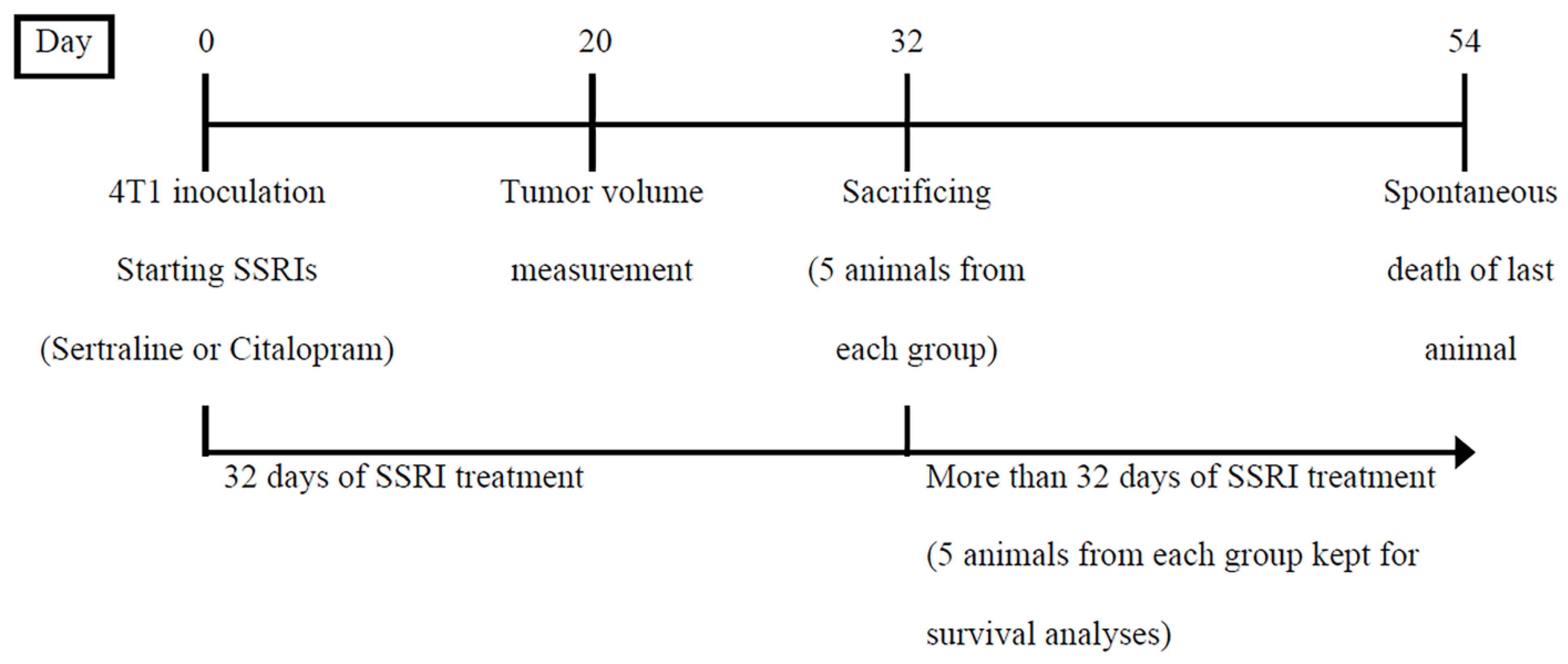
4.7.2. Experiment 2—Comparison between the Effects on Body Weight, Tumor Growth and Survival of Mice, of Sertraline and Citalopram (10 mg/kg/d Each) Administered after 4T1 Inoculation
4.8. Ex-Vivo TNF-α Cytokine Assay (ELISA)
4.9. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- McPherson, K.; Steel, C.M.; Dixon, J.M. ABC of breast diseases. Breast cancer-epidemiology, risk factors, and genetics. BMJ 2000, 321, 624–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostuzzi, G.; Matcham, F.; Dauchy, S.; Barbui, C.; Hotopf, M. Antidepressants for the treatment of depression in people with cancer. Cochrane Database Syst. Rev. 2015, CD011006. [Google Scholar] [CrossRef] [PubMed]
- Torta, R.; Siri, I.; Caldera, P. Sertraline effectiveness and safety in depressed oncological patients. Support. Care Cancer 2008, 16, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, R.; Koyama, A. Effective Therapy Against Severe Anxiety Caused by Cancer: A Case Report and Review of the Literature. Cureus 2020, 12, e8414. [Google Scholar] [CrossRef] [PubMed]
- Gil-Ad, I.; Zolokov, A.; Lomnitski, L.; Taler, M.; Bar, M.; Luria, D.; Ram, E.; Weizman, A. Evaluation of the potential anti-cancer activity of the antidepressant sertraline in human colon cancer cell lines and in colorectal cancer-xenografted mice. Int. J. Oncol. 2008, 33, 277–286. [Google Scholar] [CrossRef] [Green Version]
- Amit, B.H.; Gil-Ad, I.; Taler, M.; Bar, M.; Zolokov, A.; Weizman, A. Proapoptotic and chemosensitizing effects of selective serotonin reuptake inhibitors on T cell lymphoma/leukemia (Jurkat) In Vitro. Eur. Neuropsychopharmacol. 2009, 19, 726–734. [Google Scholar] [CrossRef]
- Tzadok, S.; Beery, E.; Israeli, M.; Uziel, O.; Lahav, M.; Fenig, E.; Gil-Ad, I.; Weizman, A.; Nordenberg, J. In Vitro novel combinations of psychotropics and anti-cancer modalities in U87 human glioblastoma cells. Int. J. Oncol. 2010, 37, 1043–1051. [Google Scholar] [CrossRef] [Green Version]
- Taler, M.; Bar, M.; Korob, I.; Lomnitski, L.; Baharav, E.; Grunbaum-Novak, N.; Weizman, A.; Gil-Ad, I. Evidence for an inhibitory immunomodulatory effect of selected antidepressants on rat splenocytes: Possible relevance to depression and hyperactive-immune disorders. Int. Immunopharmacol. 2008, 8, 526–533. [Google Scholar] [CrossRef]
- Edgar, V.A.; Genaro, A.M.; Cremaschi, G.; Sterin-Borda, L. Fluoxetine action on murine T-lymphocyte proliferation: Participation of PKC activation and calcium mobilisation. Cell. Signal. 1998, 10, 721–726. [Google Scholar] [CrossRef]
- Taler, M.; Miron, O.; Gil-Ad, I.; Weizman, A. Neuroprotective and procognitive effects of sertraline: In Vitro and In Vivo studies. Neurosci. Lett. 2013, 550, 93–97. [Google Scholar] [CrossRef]
- Khin, P.P.; Po, W.W.; Thein, W.; Sohn, U.D. Apoptotic effect of fluoxetine through the endoplasmic reticulum stress pathway in the human gastric cancer cell line AGS. Naunyn-Schmiedebergs Arch. Pharmacol. 2020, 393, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Xu, H.; Tang, Q.; Bi, F. The selective serotonin reuptake inhibitors enhance the cytotoxicity of sorafenib in hepatocellular carcinoma cells. Anticancer Drugs 2021, 32, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Hsu, L.C.; Tu, H.F.; Hsu, F.T.; Yueh, P.F.; Chiang, I.T. Beneficial effect of fluoxetine on anti-tumor progression on hepatocellular carcinoma and non-small cell lung cancer bearing animal model. Biomed. Pharmacother. 2020, 126, 110054. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.W.; Kim, E.J.; Nyiramana, M.M.; Shin, E.J.; Jin, H.; Ryu, J.H.; Kang, K.R.; Lee, G.W.; Kim, H.J.; Han, J.; et al. Paroxetine Induces Apoptosis of Human Breast Cancer MCF-7 Cells through Ca(2+)-and p38 MAP Kinase-Dependent ROS Generation. Cancers 2019, 11, 64. [Google Scholar] [CrossRef] [Green Version]
- Reiche, E.M.; Morimoto, H.K.; Nunes, S.M. Stress and depression-induced immune dysfunction: Implications for the development and progression of cancer. Int. Rev. Psychiatry 2005, 17, 515–527. [Google Scholar] [CrossRef]
- Barden, N.; Reul, J.M.; Holsboer, F. Do antidepressants stabilize mood through actions on the hypothalamic-pituitary-adrenocortical system? Trends Neurosci. 1995, 18, 6–11. [Google Scholar] [CrossRef]
- Miguel, C.; Albuquerque, E. Drug interaction in psycho-oncology: Antidepressants and antineoplastics. Pharmacology 2011, 88, 333–339. [Google Scholar] [CrossRef]
- Li, X.J.; Dai, Z.Y.; Zhu, B.Y.; Zhen, J.P.; Yang, W.F.; Li, D.Q. Effects of sertraline on executive function and quality of life in patients with advanced cancer. Med. Sci. Monit. 2014, 20, 1267–1273. [Google Scholar] [CrossRef] [Green Version]
- Stockler, M.R.; O’Connell, R.; Nowak, A.K.; Goldstein, D.; Turner, J.; Wilcken, N.R.; Wyld, D.; Abdi, E.A.; Glasgow, A.; Beale, P.J.; et al. Effect of sertraline on symptoms and survival in patients with advanced cancer, but without major depression: A placebo-controlled double-blind randomised trial. Lancet Oncol. 2007, 8, 603–612. [Google Scholar] [CrossRef]
- Zhang, N.; Sundquist, J.; Sundquist, K.; Zhang, Z.G.; Ji, J. Combined Use of Aspirin and Selective Serotonin Reuptake Inhibitors Is Associated With Lower Risk of Colorectal Cancer: A Nested Case-Control Study. Am. J. Gastroenterol. 2021, 116, 1313–1321. [Google Scholar] [CrossRef]
- Chen, V.C.; Lee, M.J.; Yang, Y.H.; Lu, M.L.; Chiu, W.C.; Dewey, M.E. Selective serotonin reuptake inhibitors use and hepatocellular carcinoma in patients with alcohol use disorder. Drug Alcohol Depend. 2021, 219, 108495. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Huang, C.W.; Chen, Y.L.; Yang, Y.H.; Chen, V.C. Association between selective serotonin reuptake inhibitors and kidney cancer risk: A nationwide population-based cohort study. Int. J. Cancer 2021, 148, 1331–1337. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.C.; Chen, V.C.; Lu, M.L.; Lee, M.J.; McIntyre, R.S.; Majeed, A.; Lee, Y.; Chen, Y.L. The Association between Selective Serotonin Reuptake Inhibitors (SSRIs) Use and the Risk of Bladder Cancer: A Nationwide Population-Based Cohort Study. Cancers 2020, 12, 1184. [Google Scholar] [CrossRef]
- Levkovitz, Y.; Gil-Ad, I.; Zeldich, E.; Dayag, M.; Weizman, A. Differential induction of apoptosis by antidepressants in glioma and neuroblastoma cell lines: Evidence for p-c-Jun, cytochrome c, and caspase-3 involvement. J. Mol. Neurosci. 2005, 27, 29–42. [Google Scholar] [CrossRef]
- Istifli, E.S.; Celik, R.; Husunet, M.T.; Cetinel, N.; Demirhan, O.; Ila, H.B. In Vitro cytogenotoxic evaluation of sertraline. Interdiscip. Toxicol. 2018, 11, 181–188. [Google Scholar] [CrossRef] [Green Version]
- Taler, M.; Gil-Ad, I.; Korob, I.; Weizman, A. The immunomodulatory effect of the antidepressant sertraline in an experimental autoimmune encephalomyelitis mouse model of multiple sclerosis. Neuroimmunomodulation 2011, 18, 117–122. [Google Scholar] [CrossRef]
- Dang, H.; Chen, Y.; Liu, X.; Wang, Q.; Wang, L.; Jia, W.; Wang, Y. Antidepressant effects of ginseng total saponins in the forced swimming test and chronic mild stress models of depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 2009, 33, 1417–1424. [Google Scholar] [CrossRef]
- Brandes, L.J.; Arron, R.J.; Bogdanovic, R.P.; Tong, J.; Zaborniak, C.L.; Hogg, G.R.; Warrington, R.C.; Fang, W.; LaBella, F.S. Stimulation of malignant growth in rodents by antidepressant drugs at clinically relevant doses. Cancer Res. 1992, 52, 3796–3800. [Google Scholar]
- Christensen, D.K.; Armaiz-Pena, G.N.; Ramirez, E.; Matsuo, K.; Zimmerman, B.; Zand, B.; Shinn, E.; Goodheart, M.J.; Bender, D.; Thaker, P.H.; et al. SSRI use and clinical outcomes in epithelial ovarian cancer. Oncotarget 2016, 7, 33179–33191. [Google Scholar] [CrossRef]
- Haque, R.; Shi, J.; Schottinger, J.E.; Ahmed, S.A.; Cheetham, T.C.; Chung, J.; Avila, C.; Kleinman, K.; Habel, L.A.; Fletcher, S.W.; et al. Tamoxifen and Antidepressant Drug Interaction in a Cohort of 16,887 Breast Cancer Survivors. J. Natl. Cancer Inst. 2016, 108, djv337. [Google Scholar] [CrossRef]
- Coogan, P.F.; Strom, B.L.; Rosenberg, L. Antidepressant use and colorectal cancer risk. Pharmacoepidemiol. Drug Saf. 2009, 18, 1111–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Couch, L.; Higuchi, M.; Fang, J.L.; Guo, L. Mitochondrial dysfunction induced by sertraline, an antidepressant agent. Toxicol. Sci. 2012, 127, 582–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giampieri, R.; Cantini, L.; Giglio, E.; Bittoni, A.; Lanese, A.; Crocetti, S.; Pecci, F.; Copparoni, C.; Meletani, T.; Lenci, E.; et al. Impact of Polypharmacy for Chronic Ailments in Colon Cancer Patients: A Review Focused on Drug Repurposing. Cancers 2020, 12, 2724. [Google Scholar] [CrossRef] [PubMed]
- Abdelaleem, M.; Ezzat, H.; Osama, M.; Megahed, A.; Alaa, W.; Gaber, A.; Shafei, A.; Refaat, A. Prospects for repurposing CNS drugs for cancer treatment. Oncol. Rev. 2019, 13, 411. [Google Scholar] [CrossRef]
- Duarte, D.; Cardoso, A.; Vale, N. Synergistic Growth Inhibition of HT-29 Colon and MCF-7 Breast Cancer Cells with Simultaneous and Sequential Combinations of Antineoplastics and CNS Drugs. Int. J. Mol. Sci. 2021, 22, 7408. [Google Scholar] [CrossRef]
- Jang, W.J.; Jung, S.K.; Vo, T.T.L.; Jeong, C.H. Anticancer activity of paroxetine in human colon cancer cells: Involvement of MET and ERBB3. J. Cell. Mol. Med. 2019, 23, 1106–1115. [Google Scholar] [CrossRef] [Green Version]
- Haukka, J.; Sankila, R.; Klaukka, T.; Lonnqvist, J.; Niskanen, L.; Tanskanen, A.; Wahlbeck, K.; Tiihonen, J. Incidence of cancer and antidepressant medication: Record linkage study. Int. J. Cancer 2010, 126, 285–296. [Google Scholar] [CrossRef]
- Boursi, B.; Lurie, I.; Mamtani, R.; Haynes, K.; Yang, Y.X. Anti-depressant therapy and cancer risk: A nested case-control study. Eur. Neuropsychopharmacol. 2015, 25, 1147–1157. [Google Scholar] [CrossRef]
- Willner, P. Validity, reliability and utility of the chronic mild stress model of depression: A 10-year review and evaluation. Psychopharmacology 1997, 134, 319–329. [Google Scholar] [CrossRef]
- First, M.; Gil-Ad, I.; Taler, M.; Tarasenko, I.; Novak, N.; Weizman, A. The effects of reboxetine treatment on depression-like behavior, brain neurotrophins, and ERK expression in rats exposed to chronic mild stress. J. Mol. Neurosci. 2013, 50, 88–97. [Google Scholar] [CrossRef]
- Ducottet, C.; Griebel, G.; Belzung, C. Effects of the selective nonpeptide corticotropin-releasing factor receptor 1 antagonist antalarmin in the chronic mild stress model of depression in mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2003, 27, 625–631. [Google Scholar] [CrossRef]
- Prut, L.; Belzung, C. The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: A review. Eur. J. Pharmacol. 2003, 463, 3–33. [Google Scholar] [CrossRef]
- de Souza, C.M.; Araujo e Silva, A.C.; de Jesus Ferraciolli, C.; Moreira, G.V.; Campos, L.C.; dos Reis, D.C.; Lopes, M.T.; Ferreira, M.A.; Andrade, S.P.; Cassali, G.D. Combination therapy with carboplatin and thalidomide suppresses tumor growth and metastasis in 4T1 murine breast cancer model. Biomed. Pharmacother. 2014, 68, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Ansermot, N.; Brawand-Amey, M.; Eap, C.B. Simultaneous quantification of selective serotonin reuptake inhibitors and metabolites in human plasma by liquid chromatography-electrospray mass spectrometry for therapeutic drug monitoring. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2012, 885, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Repetto, G.; del Peso, A.; Zurita, J.L. Neutral red uptake assay for the estimation of cell viability/cytotoxicity. Nat. Protoc. 2008, 3, 1125–1131. [Google Scholar] [CrossRef]
- Ahmed, S.A.; Gogal, R.M., Jr.; Walsh, J.E. A new rapid and simple non-radioactive assay to monitor and determine the proliferation of lymphocytes: An alternative to [3H]thymidine incorporation assay. J. Immunol. Methods 1994, 170, 211–224. [Google Scholar] [CrossRef]
- Karlsson, L.; Carlsson, B.; Hiemke, C.; Ahlner, J.; Bengtsson, F.; Schmitt, U.; Kugelberg, F.C. Altered brain concentrations of citalopram and escitalopram in P-glycoprotein deficient mice after acute and chronic treatment. Eur. Neuropsychopharmacol. 2013, 23, 1636–1644. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Start of Stress Protocol | Day 1 | Day 2 | Day 3 | Day 4 | Day 5 | Day 6 | Day 7 | |
|---|---|---|---|---|---|---|---|---|
| Week 1 | 8:00 a.m. | Confinement in a small tube (1 h) | Confinement in a small tube (1 h) | Food restriction (3 h) | Confinement in a small tube (1 h) | Confinement in a small tube (1 h) | Access to an empty bottle (2 h) | Inversion of the light/dark cycle |
| 2:00 p.m. | Forced bath in water at 32 °C (30 min) | Confinement in a small tube (1 h) | Confinement in a small tube (1 h) | Paired housing in damp sawdust (10 min) | Dark (2 h) | Housing in mildly damp sawdust (24 h) | ||
| 5:00 p.m. | Water and food deprivation (14 h) | Water deprivation (14 h) | ||||||
| Week 2 | 8:00 a.m. | Confinement in a small tube (1 h) | Confinement in a small tube (1 h) | Confinement in a small tube (1 h) | Paired housing with food restriction (10 min) | Inversion of light/dark cycle | Inversion of light/dark cycle | Inversion of light/dark cycle |
| 2:00 p.m. | Forced bath in water at 32 °C (30 min) | Paired housing in damp sawdust (10 min) | Confinement in a small tube (1 h) | Inversion of light/dark cycle | ||||
| 5:00 p.m. | Water and food deprivation (14 h) | |||||||
| Week 3 | 8:00 a.m. | Confinement in a small tube (1 h) | Food restriction (3 h) | Paired housing in damp sawdust (10 min) | Forced bath in water at 32 °C (30 min) | Access to an empty bottle (2 h) | Inversion of light/dark cycle | |
| 2:00 p.m. | Confinement in a small tube (1 h) | Confinement in a small tube (1 h) | Confinement in a small tube (1 h) | Confinement in a small tube (1 h) | Confinement in a small tube (1 h) | Inversion of light/dark cycle | ||
| 5:00 p.m. | Water and food deprivation (14 h) | Water deprivation (14 h) | Housing in mildly damp sawdust (24 h) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taler, M.; Gil-Ad, I.; Brener, I.; Hornfeld, S.H.; Weizman, A. Complex Effects of Sertraline and Citalopram on In Vitro Murine Breast Cancer Proliferation and on In Vivo Progression and Anxiety Level. Int. J. Mol. Sci. 2022, 23, 2711. https://doi.org/10.3390/ijms23052711
Taler M, Gil-Ad I, Brener I, Hornfeld SH, Weizman A. Complex Effects of Sertraline and Citalopram on In Vitro Murine Breast Cancer Proliferation and on In Vivo Progression and Anxiety Level. International Journal of Molecular Sciences. 2022; 23(5):2711. https://doi.org/10.3390/ijms23052711
Chicago/Turabian StyleTaler, Michal, Irit Gil-Ad, Iris Brener, Shay Henry Hornfeld, and Abraham Weizman. 2022. "Complex Effects of Sertraline and Citalopram on In Vitro Murine Breast Cancer Proliferation and on In Vivo Progression and Anxiety Level" International Journal of Molecular Sciences 23, no. 5: 2711. https://doi.org/10.3390/ijms23052711
APA StyleTaler, M., Gil-Ad, I., Brener, I., Hornfeld, S. H., & Weizman, A. (2022). Complex Effects of Sertraline and Citalopram on In Vitro Murine Breast Cancer Proliferation and on In Vivo Progression and Anxiety Level. International Journal of Molecular Sciences, 23(5), 2711. https://doi.org/10.3390/ijms23052711