
Salidroside, 8(E)-Nuezhenide, and Ligustroside from Ligustrum japonicum Fructus Inhibit Expressions of MMP-2 and -9 in HT 1080 Fibrosarcoma


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
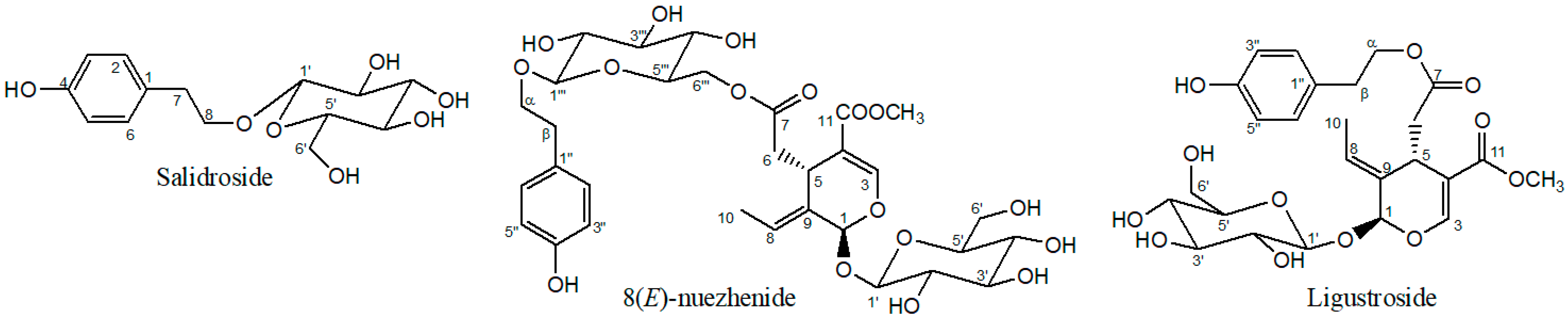
2.1. Structure Elucidation
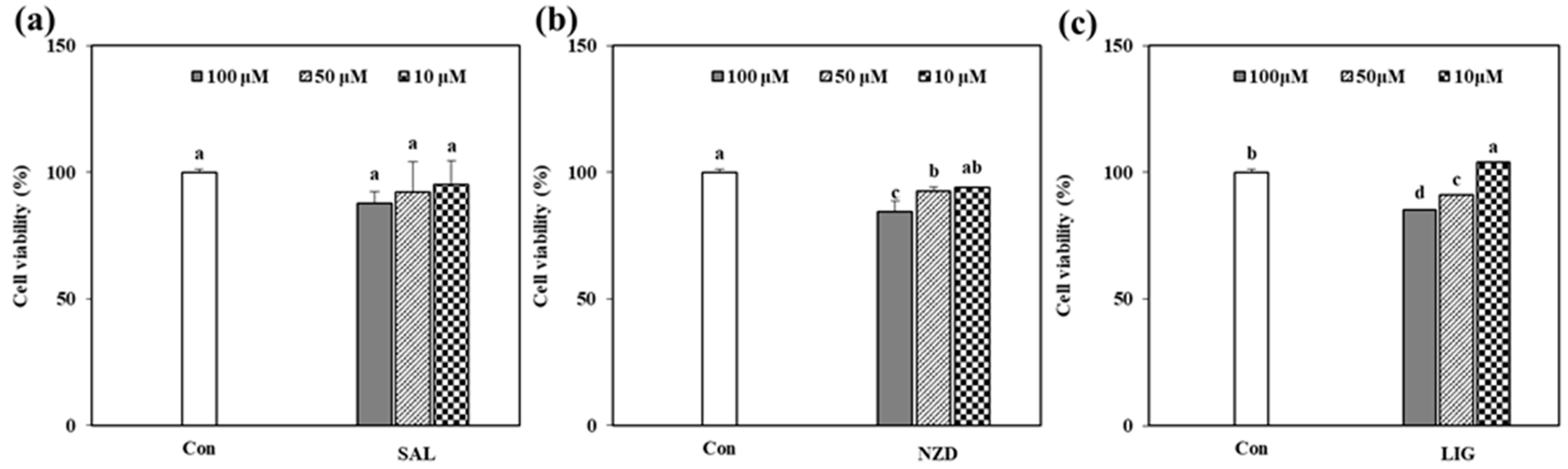
2.2. Cytotoxicity of the Isolated Compounds
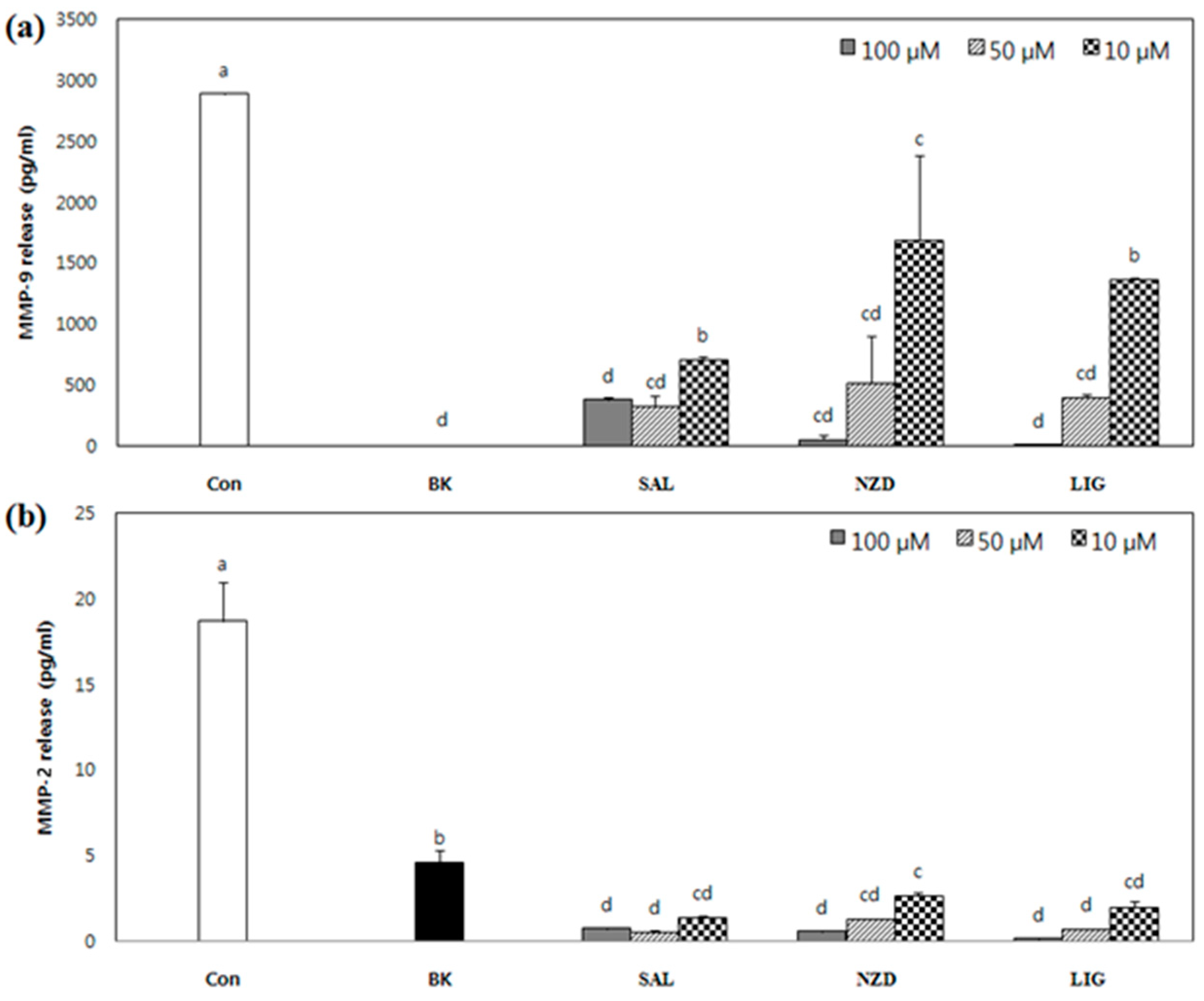
2.3. Effect of SAL, NZD and LIG on Enzymatic Activity of MMP-2 and MMP-9
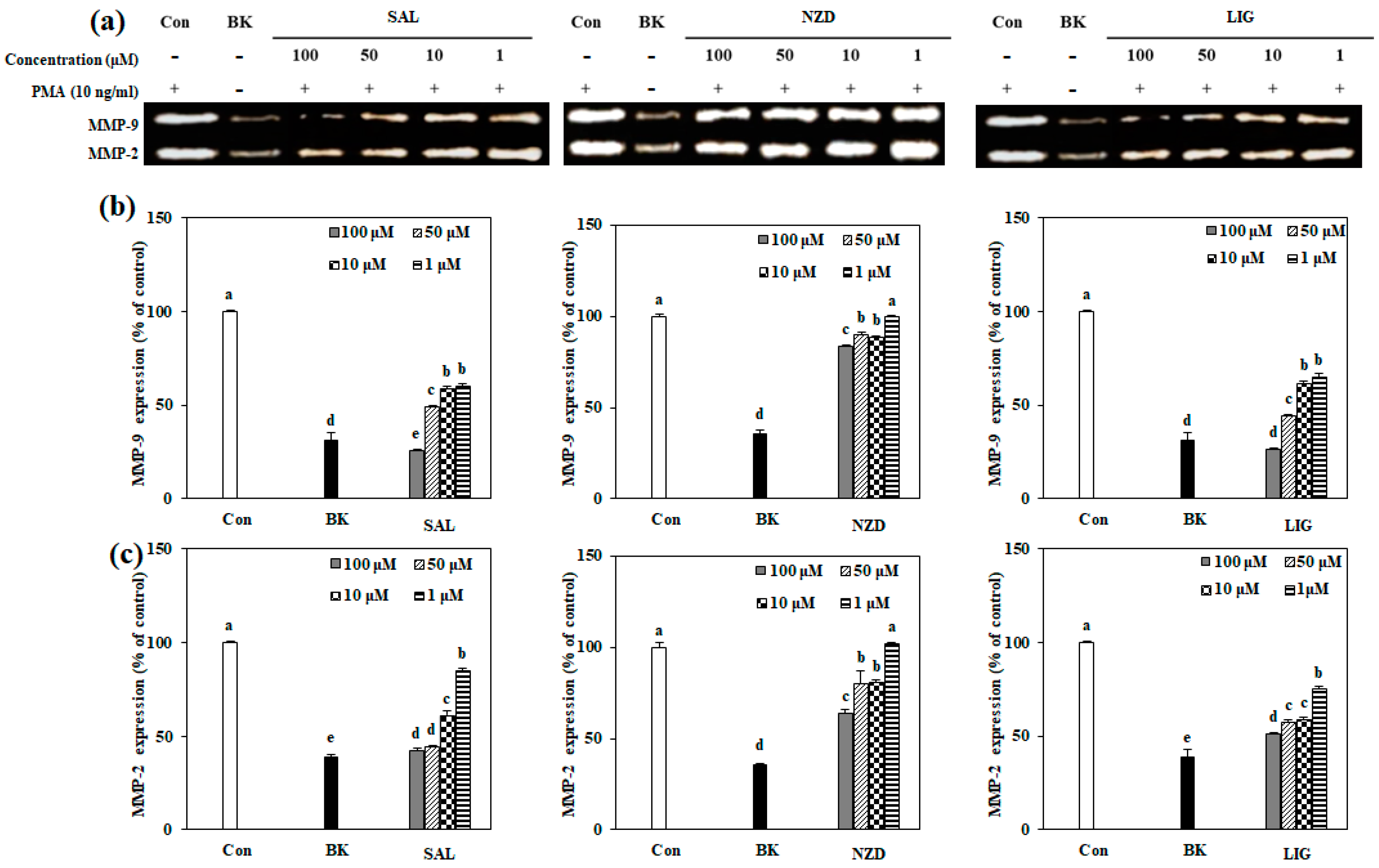
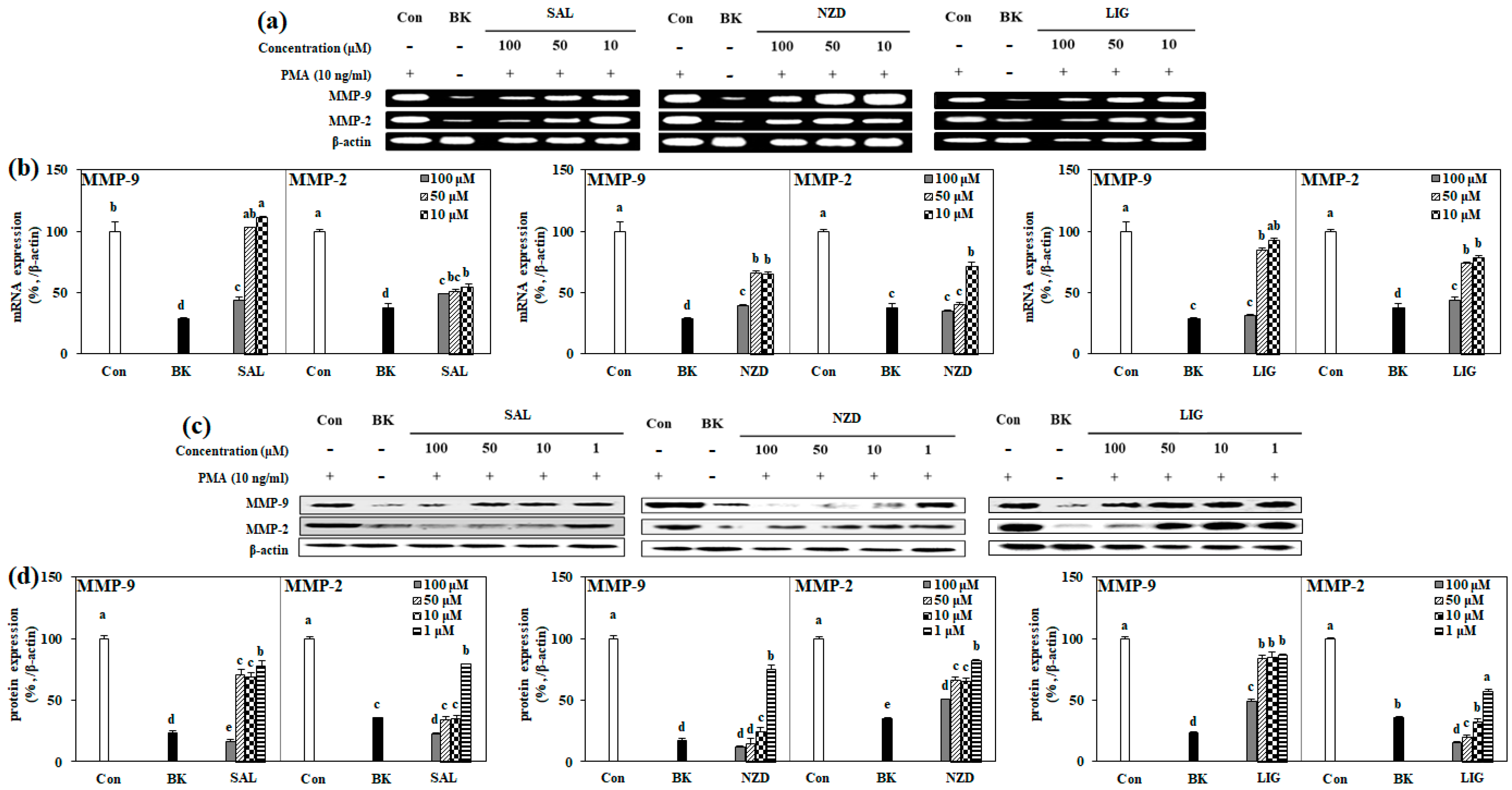
2.4. Effect of SAL, NZD and LIG on mRNA and Protein Expressions of MMP-2 and MMP-9
2.5. Effect of SAL, NZD and LIG on MAPK Signaling Pathway
3. Materials and Methods
3.1. Apparatus and Reagents
3.2. Extraction, Fractionation and Isolation
3.3. Cell Maintenance and Determination of Cytotoxicity
3.4. Determination of Active MMP-2 and MMP-9 Levels by Gelatin Zymography
3.5. Enzyme-Linked Immunosorbent Assay (ELISA) for the Detection of MMP-2 and MMP-9 Release
3.6. RNA Extraction and Reverse Transcription Polymerase Chain Reaction (RT-PCR) Analysis
3.7. Western Blot Analysis
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moss, L.A.S.; Jensen-Taubman, S.; Stetler-Stevenson, W.G. Matrix metalloproteinases: Changing roles in tumor progression and metastasis. Am. J. Pathol. 2012, 181, 1895–1899. [Google Scholar] [CrossRef] [PubMed]
- Thomas, N.V.; Kim, S.-K. Metalloproteinase inhibitors: Status and scope from marine organisms. Biochem. Res. Int. 2010, 2010, 845975. [Google Scholar] [CrossRef] [PubMed]
- Yoon, A.; Hurta, R.A. Insulin like growth factor-1 selectively regulates the expression of matrix metalloproteinase-2 in malignant H-ras transformed cells. Mol. Cell Biochem. 2001, 223, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.P.; Hansch, C. Matrix metalloproteinases (MMPs): Chemical-biological functions and (Q)SARs. Bioorganic Med. Chem. Lett. 2007, 15, 2223–2268. [Google Scholar] [CrossRef] [PubMed]
- Amar, S.; Fields, G.B. Potential clinical implications of recent matrix metalloproteinase inhibitor design strategies. Expert Rev. Proteomics. 2015, 12, 445–447. [Google Scholar]
- Zheng, H.; Takahashi, H.; Murai, Y.; Cui, Z.; Nomoto, K.; Niwa, H.; Tsuneyama, K.; Takano, Y. Expressions of MMP-2, MMP-9 and VEGF are closely linked to growth, invasion, metastasis and angiogenesis of gastric carcinoma. Anticancer Res. 2006, 26, 3579–3583. [Google Scholar]
- Daniele, A.; Zito, A.F.; Giannelli, G.; Divella, R.; Asselti, M.; Mazzocca, A.; Paradiso, A.; Quaranta, M. Expression of metalloproteinases MMP-2 and MMP-9 in sentinel lymph node and serum of patients with metastatic and non-metastatic breast cancer. Anticancer Res. 2010, 30, 3521–3527. [Google Scholar]
- Li, H.; Qiu, Z.; Li, F.; Wang, C. The relationship between MMP-2 and MMP-9 expression levels with breast cancer incidence and prognosis. Oncol. Lett. 2017, 14, 5865–5870. [Google Scholar] [CrossRef] [PubMed]
- Funasaka, T.; Hu, H.; Higan, V.; Raz, A. Down-regulation of phosphoglucose Isomerase/autocrine motility factor expression sensitizes human fibrosarcoma cells to oxidative stress leading to cellular senescence. J. Biol. Chem. 2007, 282, 36362–36369. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Joh, T.H. Matrix metalloproteinases, new insights into the understanding of neurodegenerativedisorders. Biomol. Ther. 2012, 20, 133–143. [Google Scholar] [CrossRef]
- Dufour, A.; Overall, C.M. Missing the target: Matrix metalloproteinase antitargets in inflammation and cancer. Trends Pharmacol. Sci. 2013, 34, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Vandenbroucke, R.E.; Libert, C. Is there new hope for therapeutic matrix metalloproteinase inhibition? Nat. Rev. Drug Discov. 2014, 13, 904–927. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, X.; Zhang, S.; Lu, W.; Liao, S.; Liu, X.; Shan, L.; Shen, X.; Jiang, H.; Zhang, W.; et al. Natural products as a gold mine for selective matrix metalloproteinases inhibitors. Bioorg Med. Chem. 2012, 20, 4164–4171. [Google Scholar] [CrossRef]
- Fields, G.B. The rebirth of matrix metalloproteinase inhibitors: Moving beyond the dogma. Cells 2019, 8, 984. [Google Scholar] [CrossRef]
- Kumar, G.B.; Nair, B.G.; Perry, J.J.P.; Martin, D.B.C. Recent insights into natural product inhibitors of matrix metalloproteinases. Med. Chem. Commun. 2019, 10, 2024–2037. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kong, C.-S.; Seo, Y. Inhibitory activity of Ligustrum japonicum fructus on MMP-2 and MMP-9. KSBB J. 2017, 32, 328–334. [Google Scholar] [CrossRef]
- Kim, H.; Karadeniz, F.; Kong, C.-S.; Seo, Y. Evaluation of MMP inhibitors isolated from Ligustrum japonicum fructus. Molecules 2019, 24, 604. [Google Scholar] [CrossRef]
- Kim, S.J.; Kim, K.S.; Hwang, S.J.; Chon, S.U.; Kim, Y.H.; Ahn, J.C.; Hwang, B. Identification of salidroside from Rhodiola sachalinensis A. Bor. and its production through cell suspension culture. Korean J. Medicinal Crop Sci. 2004, 12, 203–208. [Google Scholar]
- Lee, K.J.; Song, N.-Y.; Ma, J.Y. Isolation and bioactivity analysis of salidroside from Acer tegmentosum using on-line screening HPLC-ABTS+ assay. KSBB 2014, 29, 124–130. [Google Scholar] [CrossRef][Green Version]
- He, Z.D.; Dong, H.; Xu, H.-X.; Ye, W.-C.; Sun, H.-D.; But, P.-P. Secoiridoid constituents from the fruits of Ligustrum lucidum. Phytochemistry 2001, 56, 327–330. [Google Scholar] [CrossRef]
- Machida, K.; Kaneko, A.; Hosogai, T.; Kakuda, R.; Yaoita, Y.; Kikuchi, M. Studies on the constituents of Syringa species. X. Five new iridoid glycosides from the leaves of Syringa reticulata (Blume) Hara. Chem. Pharm. Bull. 2002, 50, 493–497. [Google Scholar] [CrossRef] [PubMed]
- Lei, Z.; Chen, H.; Zhang, H.; Wang, Y.; Meng, X.; Wang, Z. Evaluation of matrix metalloproteinase inhibition by peptide microarray-based fluorescence assay on polymer brush substrate and in vivo assessment. Appl. Mater. Interfaces 2017, 9, 44241–44250. [Google Scholar] [CrossRef] [PubMed]
- Krueger, J.S.; Keshamouni, V.G.; Atanaskova, N.; Reddy, K.B. Temporal and quantitative regulation of mitogen-activated protein kinase (MAPK) modulates cell motility and invasion. Oncogene 2001, 20, 4209–4218. [Google Scholar] [CrossRef]
- Vijayababu, M.R.; Arunkumar, A.; Kanagaraj, P.; Venkataraman, P.; Krishnamoorthy, G.; Arunakaran, J. Quercetin downregulates matrix metalloproteinases 2 and 9 proteins expression in prostate cancer cells (PC-3). Mol. Cell Biochem. 2006, 287, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Davidson, B.; Givant-Horwitz, V.; Lazarovici, P.; Risberg, B.; Nesland, J.M.; Trope, C.G.; Schaefer, E.; Reich, R. Matrix metalloproteinases (MMP), EMMPRIN (extracellular matrix metalloproteinase inducer) and mitogen-activated protein kinases (MAPK): Co-expression in metastatic serous ovarian carcinoma. Clin. Exp. Metastasis 2003, 20, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.B.; Krueger, J.S.; Kondapaka, S.B.; Diglio, C.A. Mitogen-activated protein kinase (MAPK) regulates the expression of progelatinase B (MMP-9) in breast epithelial cells. Int. J. Cancer 1999, 82, 268–273. [Google Scholar] [CrossRef]
- Gialeli, C.; Theocharis, A.D.; Karamanos, N.K. Roles of matrix metalloproteinases in cancer progression and their pharmacological targeting. FEBS J. 2011, 278, 16–27. [Google Scholar] [CrossRef]
- Bauvois, B. New facets of matrix metalloproteinases MMP-2 and MMP-9 as cell surface transducers: Outside-in signaling and relationship to tumor progression. Biochim. Biophys. Acta 2012, 1825, 29–36. [Google Scholar] [CrossRef]
- Kang, H.; Jang, S.-W. Roles of matrix metalloproteinase-9 in cancer metastasis. Biomed. Sci. Lett. 2014, 20, 103–110. [Google Scholar] [CrossRef][Green Version]
- Li, T.; Xu, K.; Liu, Y. Anticancer effect of salidroside reduces viability through autophagy/PI3K/Akt and MMP-9 signaling pathways in human bladder cancer cells. Oncol. Lett. 2018, 16, 3162–3168. [Google Scholar] [CrossRef]
- Ren, M.; Xu, W.; Xu, T. Salidroside represses proliferation, migration and invasion of human lung cancer cells through AKT and MEK/ERK signal pathway. Artif. Cells Nanomed. Biotechnol. 2019, 47, 1014–1021. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Wang, Z.; Zheng, Q.; Zhang, H. Salidroside inhibits migration and invasion of human fibrosarcoma HT1080 cells. Phytomed. 2012, 19, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Dinda, B.; Debnath, S.; Banik, R. Naturally occurring iridoids and secoiridoids. An updated review, part 4. Chem. Pharm. Bull. 2011, 59, 803–833. [Google Scholar] [CrossRef] [PubMed]
- Sherif, I.O.; Al-Gayyar, M.M.H. Oleuropein potentiates anti-tumor activity of cisplatin against HepG2 through affecting proNGF/NGF balance. Life Sci. 2018, 198, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Wongrattanakamon, P.; Nimmanpipug, P.; Sirithunyalug, B.; Chaiyana, W.; Jiranusornkul, S. Molecular modeling of non-covalent binding of Ligustrum lucidum secoiridoid glucosides to AP-1/matrix metalloproteinase pathway components. J. Bioenerg. Biomembr. 2018, 50, 315–327. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.; Kong, C.-S.; Seo, Y. Salidroside, 8(E)-Nuezhenide, and Ligustroside from Ligustrum japonicum Fructus Inhibit Expressions of MMP-2 and -9 in HT 1080 Fibrosarcoma. Int. J. Mol. Sci. 2022, 23, 2660. https://doi.org/10.3390/ijms23052660
Kim H, Kong C-S, Seo Y. Salidroside, 8(E)-Nuezhenide, and Ligustroside from Ligustrum japonicum Fructus Inhibit Expressions of MMP-2 and -9 in HT 1080 Fibrosarcoma. International Journal of Molecular Sciences. 2022; 23(5):2660. https://doi.org/10.3390/ijms23052660
Chicago/Turabian StyleKim, Hojun, Chang-Suk Kong, and Youngwan Seo. 2022. "Salidroside, 8(E)-Nuezhenide, and Ligustroside from Ligustrum japonicum Fructus Inhibit Expressions of MMP-2 and -9 in HT 1080 Fibrosarcoma" International Journal of Molecular Sciences 23, no. 5: 2660. https://doi.org/10.3390/ijms23052660
APA StyleKim, H., Kong, C.-S., & Seo, Y. (2022). Salidroside, 8(E)-Nuezhenide, and Ligustroside from Ligustrum japonicum Fructus Inhibit Expressions of MMP-2 and -9 in HT 1080 Fibrosarcoma. International Journal of Molecular Sciences, 23(5), 2660. https://doi.org/10.3390/ijms23052660