
Targeted Analysis of Sphingolipids in Turkeys Fed Fusariotoxins: First Evidence of Key Changes That Could Help Explain Their Relative Resistance to Fumonisin Toxicity


Abstract
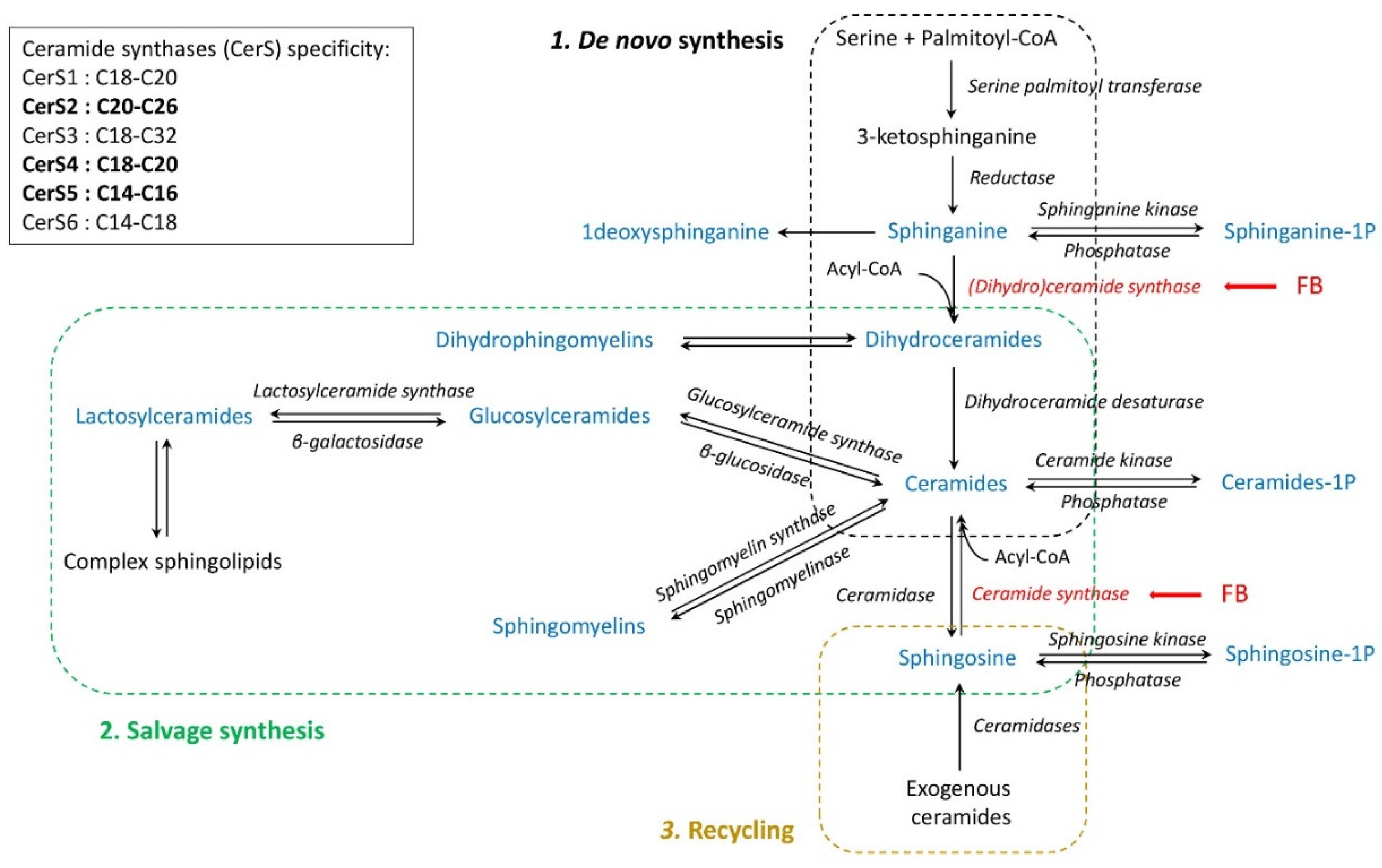
1. Introduction
2. Results
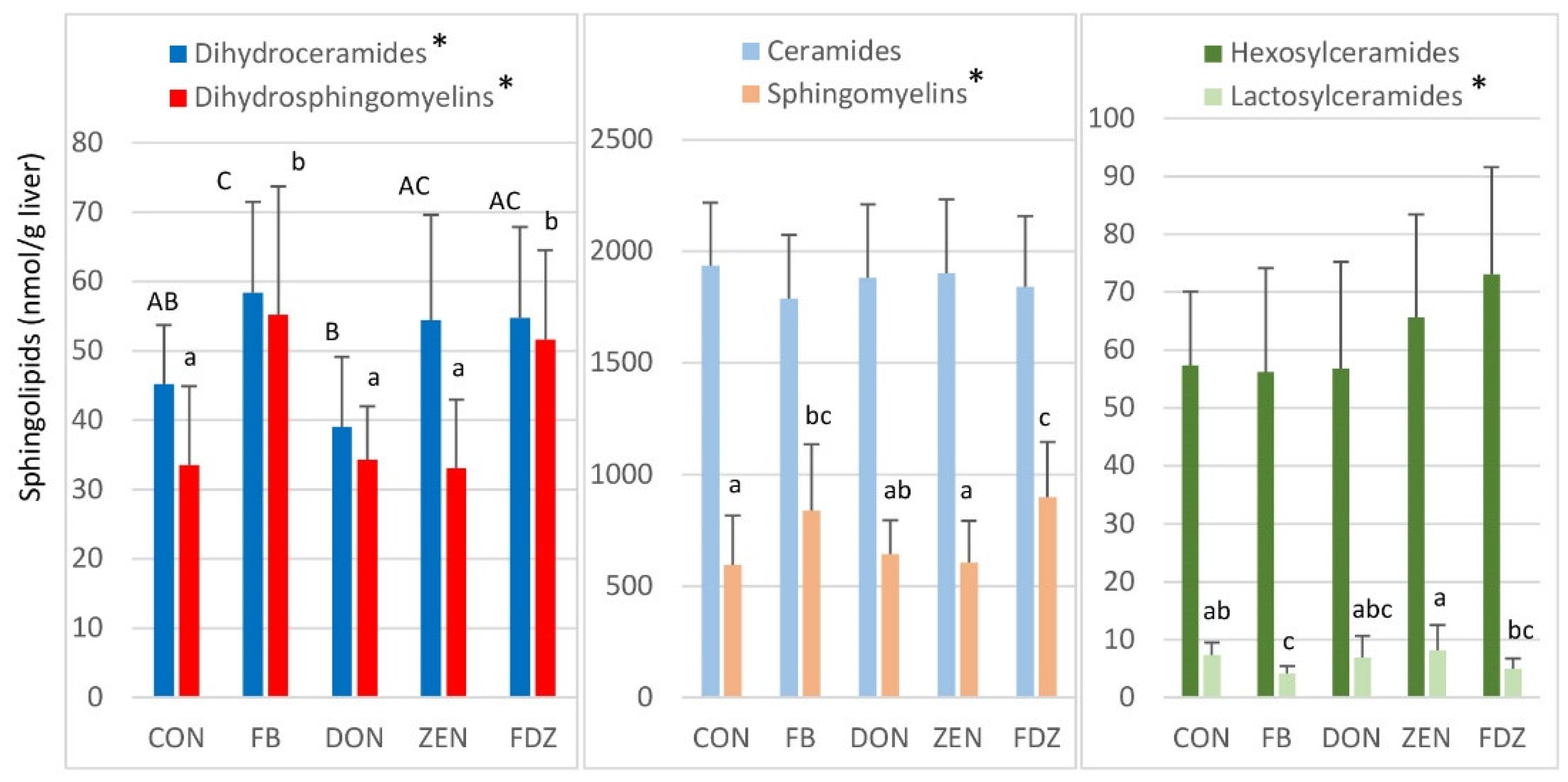
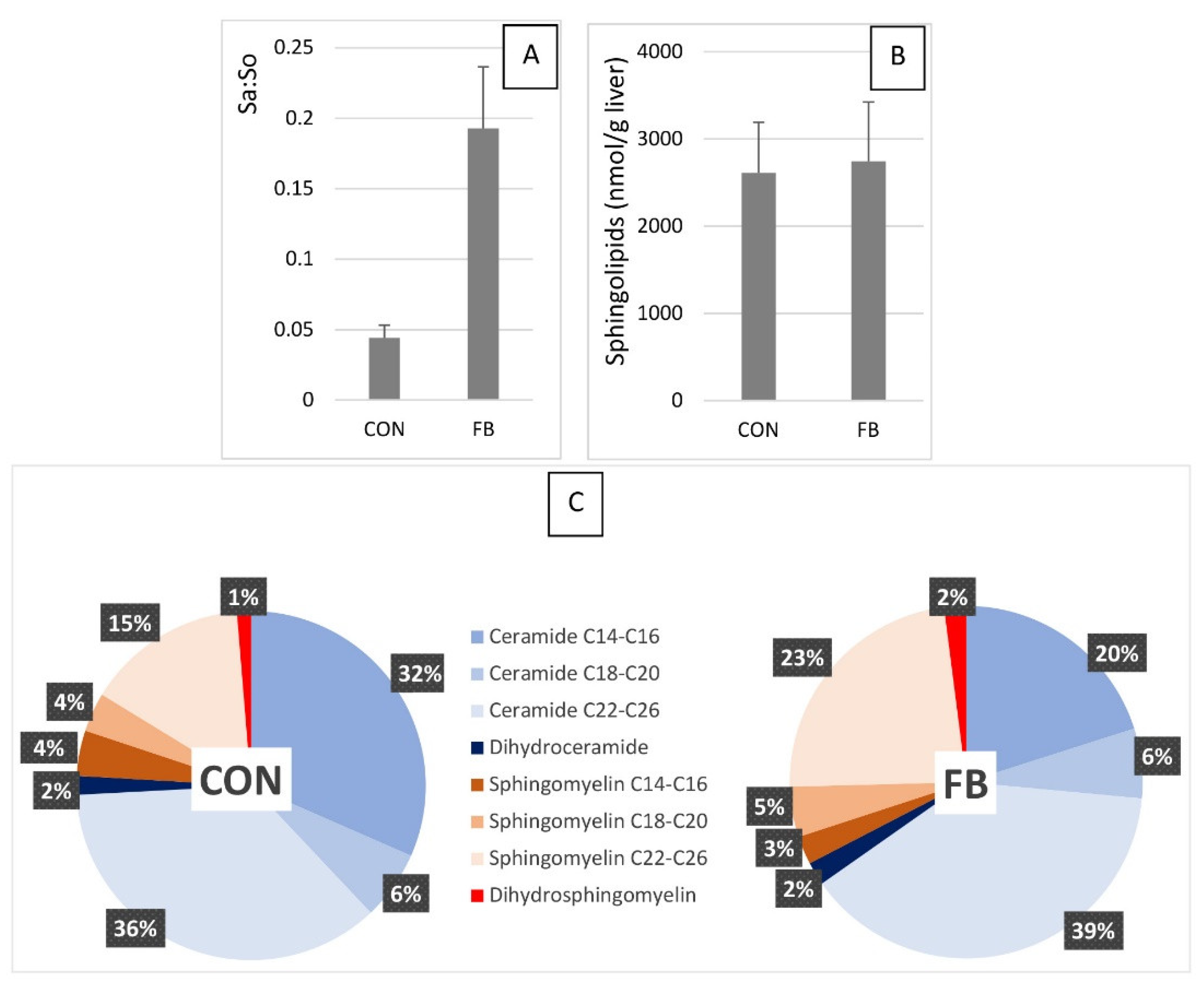
2.1. Sphingolipids in the Liver of Turkeys Fed Fumonisins
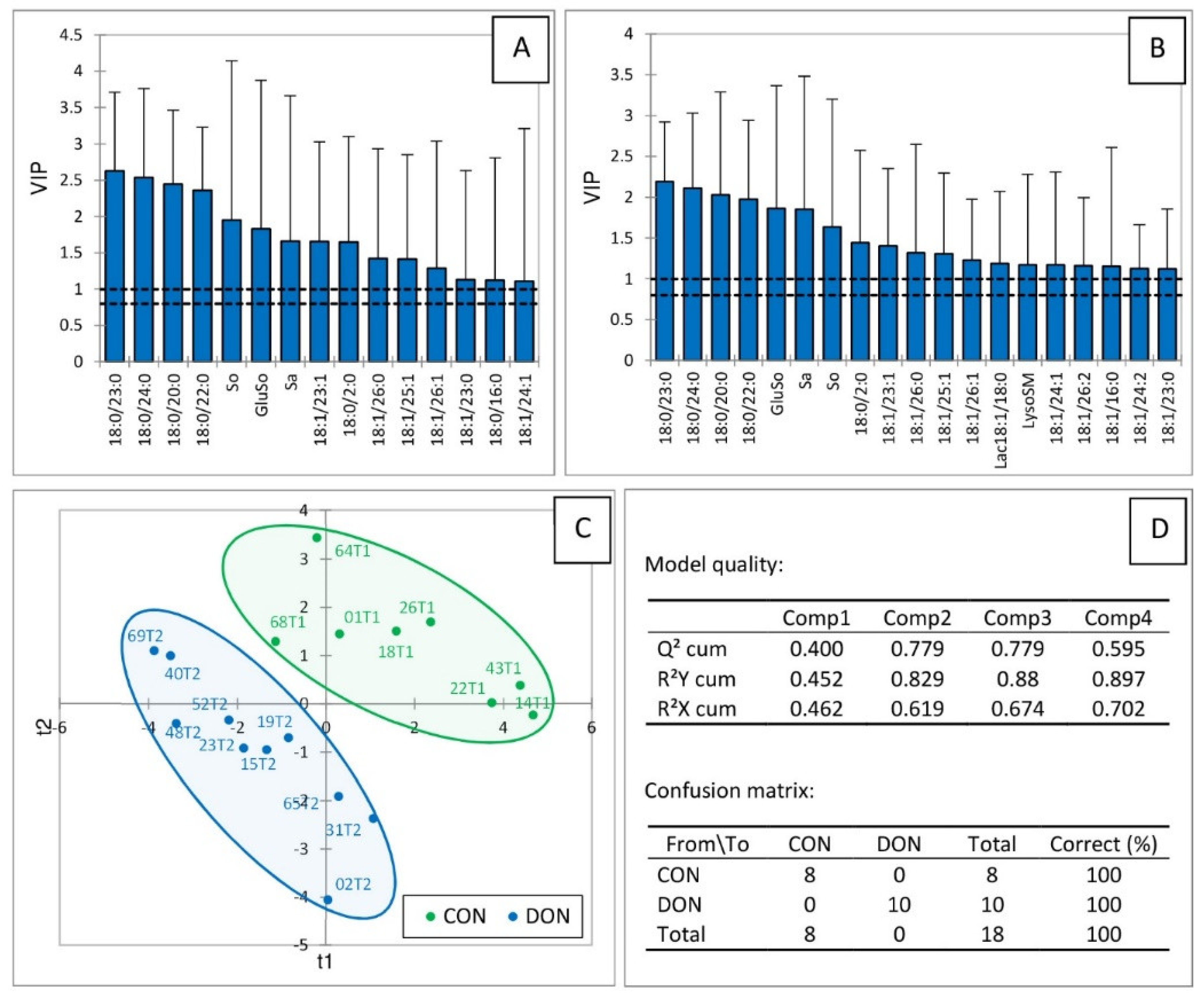
2.2. Sphingolipids in the Liver of Turkeys Fed Deoxynivalenol and Zearalenone
2.3. Sphingolipids in the Liver of Turkeys Fed A Combination of Fumonisins, Deoxynivalenol, and Zearalenone
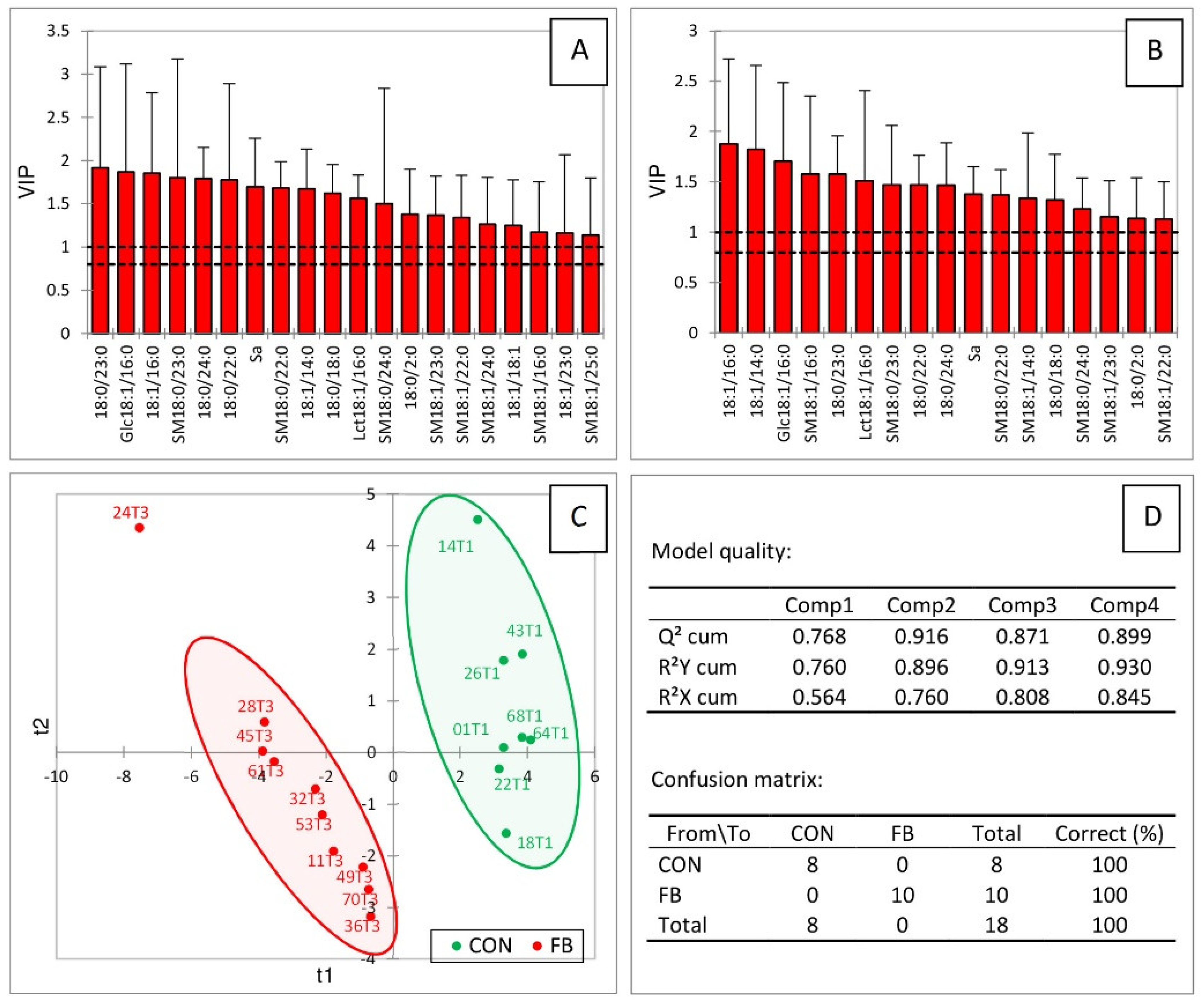
2.4. Correlations among Sphingolipids in the Liver of Turkeys Fed Fusariotoxins
2.5. Fumonisins in the Liver of Turkeys Fed the Different Diets
3. Discussion
3.1. Effects of Fumonisins
3.1.1. Sphingoid Bases
3.1.2. Dihydroceramides and Dihydrosphingomyelins
3.1.3. Ceramides
3.1.4. Glycosylceramides
3.1.5. Sphingomyelins
3.2. Effects of Deoxynivalenol and Zearalenone Alone and Combined with Fumonisins
4. Materials and Methods
4.1. Analytes and Reagents
4.2. Animals and Treatment
4.3. Chromatographic System
4.4. Sphingolipid Analysis
4.5. Measurement of Fumonisins in the Liver
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ochieng, P.E.; Scippo, M.-L.; Kemboi, D.C.; Croubels, S.; Okoth, S.; Kang’ethe, E.K.; Doupovec, B.; Gathumbi, J.K.; Lindahl, J.F.; Antonissen, G. Mycotoxins in Poultry Feed and Feed Ingredients from Sub-Saharan Africa and Their Impact on the Production of Broiler and Layer Chickens: A Review. Toxins 2021, 13, 633. [Google Scholar] [CrossRef]
- Agriopoulou, S.; Stamatelopoulou, E.; Varzakas, T. Advances in Occurrence, Importance, and Mycotoxin Control Strategies: Prevention and Detoxification in Foods. Foods 2020, 9, 137. [Google Scholar] [CrossRef] [PubMed]
- Tolosa, J.; Rodríguez-Carrasco, Y.; Ruiz, M.J.; Vila-Donat, P. Multi-mycotoxin occurrence in feed, metabolism and carry-over to animal-derived food products: A review. Food Chem. Toxicol. 2021, 158, 112661. [Google Scholar] [CrossRef] [PubMed]
- Riley, R.T.; Merrill, A.H. Ceramide synthase inhibition by fumonisins: A perfect storm of perturbed sphingolipid metabolism, signaling, and disease. J. Lipid. Res. 2019, 60, 1183–1189. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Contaminants in the Food Chain (CONTAM); Knutsen, H.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; et al. Risks for animal health related to the presence of fumonisins, their modified forms and hidden forms in feed. EFSA J. 2018, 16, e05242. [Google Scholar] [CrossRef]
- IARC. MOnograph 82, I.W.G. FUMONISIN B1; IARC Press: Lyon, France, 2002. [Google Scholar]
- Liu, X.; Fan, L.; Yin, S.; Chen, H.; Hu, H. Molecular mechanisms of fumonisin B1-induced toxicities and its applications in the mechanism-based interventions. Toxicon 2019, 167, 1–5. [Google Scholar] [CrossRef]
- Wangia-Dixon, R.N.; Nishimwe, K. Molecular toxicology and carcinogenesis of fumonisins: A review. J. Environ. Sci. Health C Toxicol. Carcinog. 2021, 39, 44–67. [Google Scholar] [CrossRef]
- Arumugam, T.; Ghazi, T.; Chuturgoon, A.A. Molecular and epigenetic modes of Fumonisin B1 mediated toxicity and carcinogenesis and detoxification strategies. Crit. Rev. Toxicol. 2021, 51, 76–94. [Google Scholar] [CrossRef]
- Chen, J.; Wei, Z.; Wang, Y.; Long, M.; Wu, W.; Kuca, K. Fumonisin B1: Mechanisms of toxicity and biological detoxification progress in animals. Food Chem. Toxicol. 2021, 149, 111977. [Google Scholar] [CrossRef]
- Wangia, R.N.; Githanga, D.P.; Xue, K.S.; Tang, L.; Anzala, O.A.; Wang, J.-S. Validation of urinary sphingolipid metabolites as biomarker of effect for fumonisins exposure in Kenyan children. Biomarkers 2019, 24, 379–388. [Google Scholar] [CrossRef]
- Cingolani, F.; Futerman, A.H.; Casas, J. Ceramide synthases in biomedical research. Chem. Phys. Lipids 2016, 197, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, D.; Wegner, M.-S.; Wanger, R.A.; Ferreirós, N.; Schreiber, Y.; Lucks, J.; Schiffmann, S.; Geisslinger, G.; Grösch, S. The equilibrium between long and very long chain ceramides is important for the fate of the cell and can be influenced by co-expression of CerS. Int. J. Biochem. Cell Biol. 2013, 45, 1195–1203. [Google Scholar] [CrossRef] [PubMed]
- Pewzner-Jung, Y.; Park, H.; Laviad, E.L.; Silva, L.C.; Lahiri, S.; Stiban, J.; Erez-Roman, R.; Brügger, B.; Sachsenheimer, T.; Wieland, F.; et al. A critical role for ceramide synthase 2 in liver homeostasis: I. alterations in lipid metabolic pathways. J. Biol. Chem. 2010, 285, 10902–10910. [Google Scholar] [CrossRef] [PubMed]
- Pewzner-Jung, Y.; Brenner, O.; Braun, S.; Laviad, E.L.; Ben-Dor, S.; Feldmesser, E.; Horn-Saban, S.; Amann-Zalcenstein, D.; Raanan, C.; Berkutzki, T.; et al. A critical role for ceramide synthase 2 in liver homeostasis: II. insights into molecular changes leading to hepatopathy. J. Biol. Chem. 2010, 285, 10911–10923. [Google Scholar] [CrossRef] [PubMed]
- Lachkar, F.; Ferré, P.; Foufelle, F.; Papaioannou, A. Dihydroceramides: Their emerging physiological roles and functions in cancer and metabolic diseases. Am. J. Physiol. Endocrinol. Metab. 2021, 320, E122–E130. [Google Scholar] [CrossRef]
- Zitomer, N.C.; Mitchell, T.; Voss, K.A.; Bondy, G.S.; Pruett, S.T.; Garnier-Amblard, E.C.; Liebeskind, L.S.; Park, H.; Wang, E.; Sullards, M.C.; et al. Ceramide synthase inhibition by fumonisin B1 causes accumulation of 1-deoxysphinganine: A novel category of bioactive 1-deoxysphingoid bases and 1-deoxydihydroceramides biosynthesized by mammalian cell lines and animals. J. Biol. Chem. 2009, 284, 4786–4795. [Google Scholar] [CrossRef]
- He, Q.; Suzuki, H.; Sharma, N.; Sharma, R.P. Ceramide synthase inhibition by fumonisin B1 treatment activates sphingolipid-metabolizing systems in mouse liver. Toxicol. Sci. 2006, 94, 388–397. [Google Scholar] [CrossRef]
- Mignard, V.; Dubois, N.; Lanoé, D.; Joalland, M.-P.; Oliver, L.; Pecqueur, C.; Heymann, D.; Paris, F.; Vallette, F.M.; Lalier, L. Sphingolipid distribution at mitochondria-associated membranes (MAMs) upon induction of apoptosis. J. Lipid. Res. 2020, 61, 1025–1037. [Google Scholar] [CrossRef]
- Burger, H.-M.; Abel, S.; Gelderblom, W.C.A. Modulation of key lipid raft constituents in primary rat hepatocytes by fumonisin B1—Implications for cancer promotion in the liver. Food Chem. Toxicol. 2018, 115, 34–41. [Google Scholar] [CrossRef]
- Song, Y.; Liu, W.; Zhao, Y.; Zang, J.; Gao, H. Fumonisin B1 exposure induces apoptosis of human kidney tubular epithelial cells through regulating PTEN/PI3K/AKT signaling pathway via disrupting lipid raft formation. Toxicon 2021, 204, 31–36. [Google Scholar] [CrossRef]
- Riedel, S.; Abel, S.; Burger, H.-M.; van der Westhuizen, L.; Swanevelder, S.; Gelderblom, W.C.A. Differential modulation of the lipid metabolism as a model for cellular resistance to fumonisin B1–induced cytotoxic effects in vitro. Prostaglandins Leukot. Essent. Fat. Acids 2016, 109, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Travel, A.; Metayer, J.-P.; Mika, A.; Bailly, J.-D.; Cleva, D.; Boissieu, C.; Guennec, J.L.; Albaric, O.; Labrut, S.; Lepivert, G.; et al. Toxicity of Fumonisins, Deoxynivalenol, and Zearalenone Alone and in Combination in Turkeys Fed with the Maximum European Union–Tolerated Level. Avian Dis. 2019, 63, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Benlasher, E.; Geng, X.; Nguyen, N.T.X.; Tardieu, D.; Bailly, J.-D.; Auvergne, A.; Guerre, P. Comparative effects of fumonisins on sphingolipid metabolism and toxicity in ducks and turkeys. Avian Dis. 2012, 56, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Tardieu, D.; Bailly, J.-D.; Skiba, F.; Métayer, J.-P.; Grosjean, F.; Guerre, P. Chronic toxicity of fumonisins in turkeys. Poult. Sci. 2007, 86, 1887–1893. [Google Scholar] [CrossRef] [PubMed]
- Masching, S.; Naehrer, K.; Schwartz-Zimmermann, H.-E.; Sărăndan, M.; Schaumberger, S.; Dohnal, I.; Nagl, V.; Schatzmayr, D. Gastrointestinal Degradation of Fumonisin B1 by Carboxylesterase FumD Prevents Fumonisin Induced Alteration of Sphingolipid Metabolism in Turkey and Swine. Toxins 2016, 8, 84. [Google Scholar] [CrossRef] [PubMed]
- Broomhead, J.N.; Ledoux, D.R.; Bermudez, A.J.; Rottinghaus, G.E. Chronic effects of fumonisin B1 in broilers and turkeys fed dietary treatments to market age. Poult. Sci. 2002, 81, 56–61. [Google Scholar] [CrossRef]
- Loiseau, N.; Polizzi, A.; Dupuy, A.; Therville, N.; Rakotonirainy, M.; Loy, J.; Viadere, J.-L.; Cossalter, A.-M.; Bailly, J.-D.; Puel, O.; et al. New insights into the organ-specific adverse effects of fumonisin B1: Comparison between lung and liver. Arch. Toxicol. 2015, 89, 1619–1629. [Google Scholar] [CrossRef]
- Terciolo, C.; Bracarense, A.P.; Souto, P.C.M.C.; Cossalter, A.-M.; Dopavogui, L.; Loiseau, N.; Oliveira, C.A.F.; Pinton, P.; Oswald, I.P. Fumonisins at Doses below EU Regulatory Limits Induce Histological Alterations in Piglets. Toxins 2019, 11, 548. [Google Scholar] [CrossRef]
- Ali, O.; Szabó-Fodor, J.; Fébel, H.; Mézes, M.; Balogh, K.; Glávits, R.; Kovács, M.; Zantomasi, A.; Szabó, A. Porcine Hepatic Response to Fumonisin B1 in a Short Exposure Period: Fatty Acid Profile and Clinical Investigations. Toxins 2019, 11, 655. [Google Scholar] [CrossRef]
- Guerre, P. Fusariotoxins in Avian Species: Toxicokinetics, Metabolism and Persistence in Tissues. Toxins 2015, 7, 2289–2305. [Google Scholar] [CrossRef]
- Tardieu, D.; Travel, A.; Le Bourhis, C.; Metayer, J.-P.; Mika, A.; Cleva, D.; Boissieu, C.; Guerre, P. Fumonisins and zearalenone fed at low levels can persist several days in the liver of turkeys and broiler chickens after exposure to the contaminated diet was stopped. Food Chem. Toxicol. 2021, 148, 111968. [Google Scholar] [CrossRef] [PubMed]
- Laurain, J.; Tardieu, D.; Matard-Mann, M.; Rodriguez, M.A.; Guerre, P. Fumonisin B1 Accumulates in Chicken Tissues over Time and This Accumulation Was Reduced by Feeding Algo-Clay. Toxins 2021, 13, 701. [Google Scholar] [CrossRef] [PubMed]
- Tardieu, D.; Tran, S.T.; Auvergne, A.; Babilé, R.; Benard, G.; Bailly, J.D.; Guerre, P. Effects of fumonisins on liver and kidney sphinganine and the sphinganine to sphingosine ratio during chronic exposure in ducks. Chem. Biol. Interact. 2006, 160, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Tardieu, D.; Matard-Mann, M.; Collén, P.N.; Guerre, P. Strong Alterations in the Sphingolipid Profile of Chickens Fed a Dose of Fumonisins Considered Safe. Toxins 2021, 13, 770. [Google Scholar] [CrossRef]
- Wang, J.-R.; Zhang, H.; Yau, L.F.; Mi, J.-N.; Lee, S.; Lee, K.C.; Hu, P.; Liu, L.; Jiang, Z.-H. Improved sphingolipidomic approach based on ultra-high performance liquid chromatography and multiple mass spectrometries with application to cellular neurotoxicity. Anal. Chem. 2014, 86, 5688–5696. [Google Scholar] [CrossRef]
- Voss, K.A.; Smith, G.W.; Haschek, W.M. Fumonisins: Toxicokinetics, mechanism of action and toxicity. Anim. Feed Sci. Technol. 2007, 137, 299–325. [Google Scholar] [CrossRef]
- Guerre, P. Worldwide Mycotoxins Exposure in Pig and Poultry Feed Formulations. Toxins 2016, 8, 350. [Google Scholar] [CrossRef]
- Suzuki, T.; Iwahashi, Y. Acetylated Deoxynivalenol Generates Differences of Gene Expression that Discriminate Trichothecene Toxicity. Toxins 2016, 8, 42. [Google Scholar] [CrossRef]
- Johnson, E.L.; Heaver, S.L.; Waters, J.L.; Kim, B.I.; Bretin, A.; Goodman, A.L.; Gewirtz, A.T.; Worgall, T.S.; Ley, R.E. Sphingolipids produced by gut bacteria enter host metabolic pathways impacting ceramide levels. Nat. Commun. 2020, 11, 2471. [Google Scholar] [CrossRef]
- Guerre, P. Mycotoxin and Gut Microbiota Interactions. Toxins 2020, 12, 769. [Google Scholar] [CrossRef]
- Szabó, A.; Szabó-Fodor, J.; Fébel, H.; Mézes, M.; Balogh, K.; Bázár, G.; Kocsó, D.; Ali, O.; Kovács, M. Individual and Combined Effects of Fumonisin B1, Deoxynivalenol and Zearalenone on the Hepatic and Renal Membrane Lipid. Integrity of Rats. Toxins 2017, 10, 4. [Google Scholar] [CrossRef]
- Ali, O.; Mézes, M.; Balogh, K.; Kovács, M.; Szabó, A. The Effects of Mixed Fusarium Mycotoxins at EU-Permitted Feed Levels on Weaned Piglets’ Tissue Lipids. Toxins 2021, 13, 444. [Google Scholar] [CrossRef] [PubMed]
- Panel, E.C. Opinion of the Scientific Panel on Contaminants in Food Chain on a Request from the Commission Related to Fumonisins as Undesirable Substances in Animal Feed. EFSA J. 2015, 235, 1–32. [Google Scholar]
- EFSA Panel on Contaminants in the Food Chain (CONTAM); Knutsen, H.-K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; et al. Risks for animal health related to the presence of zearalenone and its modified forms in feed. EFSA J. 2017, 15, e04851. [Google Scholar] [CrossRef]
- EFSA. Risks to Human and Animal Health Related to the Presence of Deoxynivalenol and Its Acetylated and Modified Forms in Food and Feed. Available online: https://www.efsa.europa.eu/en/efsajournal/pub/4718 (accessed on 16 December 2021).
- Ledoux, D.R.; Bermudez, A.J.; Rottinghaus, G.E. Effects of feeding Fusarium moniliforme culture material, containing known levels of fumonisin B1, in the young turkey poult. Poult. Sci. 1996, 75, 1472–1478. [Google Scholar] [CrossRef] [PubMed]
- Weibking, T.; Ledoux, D.R.; Bermudez, A.J.; Turk, J.R.; Rottinghaus, G.E. Effects on Turkey Poults of Feeding Fusarium moniliforme M-1325 Culture Material Grown under Different Environmental Conditions. Avian Dis. 1995, 39, 32–38. [Google Scholar] [CrossRef]
- Weibking, T.S.; Ledoux, D.R.; Bermudez, A.J.; Rotttinghaus, G.E. Individual and Combined Effects of Feeding Fusarium moniliforme Culture Material, Containing Known Levels of Fumonisin B1 and Aflatoxin B1 in the Young Turkey Poult. Poult. Sci. 1994, 73, 1517–1525. [Google Scholar] [CrossRef]
- Duan, J.; Merrill, A.H. 1-Deoxysphingolipids Encountered Exogenously and Made de Novo: Dangerous Mysteries inside an Enigma. J. Biol. Chem. 2015, 290, 15380–15389. [Google Scholar] [CrossRef]
- He, Q.; Riley, R.T.; Sharma, R.P. Myriocin prevents fumonisin B1-induced sphingoid base accumulation in mice liver without ameliorating hepatotoxicity. Food Chem. Toxicol. 2005, 43, 969–979. [Google Scholar] [CrossRef]
- Wang, H.; Maurer, B.J.; Liu, Y.-Y.; Wang, E.; Allegood, J.C.; Kelly, S.; Symolon, H.; Liu, Y.; Merrill, A.H.; Gouazé-Andersson, V.; et al. N-(4-Hydroxyphenyl)retinamide increases dihydroceramide and synergizes with dimethylsphingosine to enhance cancer cell killing. Mol. Cancer 2008, 7, 2967–2976. [Google Scholar] [CrossRef]
- Signorelli, P.; Munoz-Olaya, J.M.; Gagliostro, V.; Casas, J.; Ghidoni, R.; Fabriàs, G. Dihydroceramide intracellular increase in response to resveratrol treatment mediates autophagy in gastric cancer cells. Cancer Lett. 2009, 282, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Ghandour, B.; Pisano, C.; Darwiche, N.; Dbaibo, G. Restoration of ceramide de novo synthesis by the synthetic retinoid ST1926 as it induces adult T-cell leukemia cell death. Biosci. Rep. 2020, 40, BSR20200050. [Google Scholar] [CrossRef] [PubMed]
- Gelderblom, W.C.A.; Moritz, W.; Swanevelder, S.; Smuts, C.M.; Abel, S. Lipids and delta6-desaturase activity alterations in rat liver microsomal membranes induced by fumonisin B1. Lipids 2002, 37, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Burger, H.-M.; Abel, S.; Snijman, P.W.; Swanevelder, S.; Gelderblom, W.C.A. Altered lipid parameters in hepatic subcellular membrane fractions induced by fumonisin B1. Lipids 2007, 42, 249–261. [Google Scholar] [CrossRef]
- Riebeling, C.; Allegood, J.C.; Wang, E.; Merrill, A.H.; Futerman, A.H. Two Mammalian Longevity Assurance Gene (LAG1) Family Members, trh1 and trh4, Regulate Dihydroceramide Synthesis Using Different Fatty Acyl-CoA Donors. J. Biol. Chem. 2003, 278, 43452–43459. [Google Scholar] [CrossRef]
- Blitzer, J.T.; Wang, L.; Summers, S.A. DES1: A Key Driver of Lipotoxicity in Metabolic Disease. DNA Cell Biol. 2020, 39, 733–737. [Google Scholar] [CrossRef]
- Javed, T.; Bunte, R.M.; Dombrink-Kurtzman, M.A.; Richard, J.L.; Bennett, G.A.; Côté, L.M.; Buck, W.B. Comparative pathologic changes in broiler chicks on feed amended with Fusarium proliferatum culture material or purified fumonisin B1 and moniliformin. Mycopathologia 2005, 159, 553–564. [Google Scholar] [CrossRef]
- Tardieu, D.; Bailly, J.D.; Benard, G.; Tran, T.S.; Guerre, P. Toxicity of maize containing known levels of fumonisin B1 during force-feeding of ducks. Poult. Sci. 2004, 83, 1287–1293. [Google Scholar] [CrossRef]
- Savira, F.; Magaye, R.; Scullino, C.V.; Flynn, B.L.; Pitson, S.M.; Anderson, D.; Creek, D.J.; Hua, Y.; Xiong, X.; Huang, L.; et al. Sphingolipid imbalance and inflammatory effects induced by uremic toxins in heart and kidney cells are reversed by dihydroceramide desaturase 1 inhibition. Toxicol. Lett. 2021, 350, 133–142. [Google Scholar] [CrossRef]
- Wang, E.; Norred, W.P.; Bacon, C.W.; Riley, R.T.; Merrill, A.H. Inhibition of sphingolipid biosynthesis by fumonisins. Implications for diseases associated with Fusarium moniliforme. J. Biol. Chem. 1991, 266, 14486–14490. [Google Scholar] [CrossRef]
- Lumsangkul, C.; Tso, K.-H.; Fan, Y.-K.; Chiang, H.-I.; Ju, J.-C. Mycotoxin Fumonisin B1 Interferes Sphingolipid Metabolisms and Neural Tube Closure during Early Embryogenesis in Brown Tsaiya Ducks. Toxins 2021, 13, 743. [Google Scholar] [CrossRef]
- Mesicek, J.; Lee, H.; Feldman, T.; Jiang, X.; Skobeleva, A.; Berdyshev, E.V.; Haimovitz-Friedman, A.; Fuks, Z.; Kolesnick, R. Ceramide synthases 2, 5, and 6 confer distinct roles in radiation-induced apoptosis in HeLa cells. Cell Signal. 2010, 22, 1300–1307. [Google Scholar] [CrossRef]
- Stiban, J.; Perera, M. Very long chain ceramides interfere with C16-ceramide-induced channel formation: A plausible mechanism for regulating the initiation of intrinsic apoptosis. Biochim. Biophys. Acta 2015, 1848, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Mullen, T.D.; Spassieva, S.; Jenkins, R.W.; Kitatani, K.; Bielawski, J.; Hannun, Y.A.; Obeid, L.M. Selective knockdown of ceramide synthases reveals complex interregulation of sphingolipid metabolism. J. Lipid. Res. 2011, 52, 68–77. [Google Scholar] [CrossRef]
- Merrill, A.H.; van Echten, G.; Wang, E.; Sandhoff, K. Fumonisin B1 inhibits sphingosine (sphinganine) N-acyltransferase and de novo sphingolipid biosynthesis in cultured neurons in situ. J. Biol. Chem. 1993, 268, 27299–27306. [Google Scholar] [CrossRef]
- He, Q.; Riley, R.T.; Sharma, R.P. Pharmacological antagonism of fumonisin B1 cytotoxicity in porcine renal epithelial cells (LLC-PK1): A model for reducing fumonisin-induced nephrotoxicity in vivo. Pharm. Toxicol. 2002, 90, 268–277. [Google Scholar] [CrossRef]
- Grenier, B.; Dohnal, I.; Shanmugasundaram, R.; Eicher, S.D.; Selvaraj, R.K.; Schatzmayr, G.; Applegate, T.J. Susceptibility of Broiler Chickens to Coccidiosis When Fed Subclinical Doses of Deoxynivalenol and Fumonisins-Special Emphasis on the Immunological Response and the Mycotoxin Interaction. Toxins 2016, 8, 231. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-J.; Zhang, R.-Q.; Zhai, Q.-Y.; Liu, J.-C.; Li, N.; Liu, W.-X.; Li, L.; Shen, W. Metagenomic analysis of gut microbiota alteration in a mouse model exposed to mycotoxin deoxynivalenol. Toxicol. Appl. Pharmacol. 2019, 372, 47–56. [Google Scholar] [CrossRef]
- Vignal, C.; Djouina, M.; Pichavant, M.; Caboche, S.; Waxin, C.; Beury, D.; Hot, D.; Gower-Rousseau, C.; Body-Malapel, M. Chronic ingestion of deoxynivalenol at human dietary levels impairs intestinal homeostasis and gut microbiota in mice. Arch. Toxicol. 2018, 92, 2327–2338. [Google Scholar] [CrossRef]
- Lee, M.-T.; Le, H.H.; Johnson, E.L. Dietary sphinganine is selectively assimilated by members of the mammalian gut microbiome. J. Lipid. Res. 2021, 62, 100034. [Google Scholar] [CrossRef]
- Norred, W.P.; Plattner, R.D.; Dombrink-Kurtzman, M.A.; Meredith, F.I.; Riley, R.T. Mycotoxin-induced elevation of free sphingoid bases in precision-cut rat liver slices: Specificity of the response and structure-activity relationships. Toxicol. Appl. Pharm. 1997, 147, 63–70. [Google Scholar] [CrossRef][Green Version]
- Metayer, J.-P.; Travel, A.; Mika, A.; Bailly, J.-D.; Cleva, D.; Boissieu, C.; Guennec, J.L.; Froment, P.; Albaric, O.; Labrut, S.; et al. Lack of Toxic Interaction Between Fusariotoxins in Broiler Chickens Fed throughout Their Life at the Highest Level Tolerated in the European Union. Toxins 2019, 11, 455. [Google Scholar] [CrossRef] [PubMed]
- Peillod, C.; Laborde, M.; Travel, A.; Mika, A.; Bailly, J.D.; Cleva, D.; Boissieu, C.; Le Guennec, J.; Albaric, O.; Labrut, S.; et al. Toxic Effects of Fumonisins, Deoxynivalenol and Zearalenone Alone and in Combination in Ducks Fed the Maximum EUTolerated Level. Toxins 2021, 13, 152. [Google Scholar] [CrossRef] [PubMed]
- Tardieu, D.; Travel, A.; Metayer, J.-P.; Le Bourhis, C.; Guerre, P. Fumonisin B1, B2 and B3 in Muscle and Liver of Broiler Chickens and Turkey Poults Fed with Diets Containing Fusariotoxins at the EU Maximum Tolerable Level. Toxins 2019, 11, 590. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CON | FB | DON | ZEN | FDZ | |
|---|---|---|---|---|---|
| Sphingoid bases and derivates 2 | |||||
| dSa | 24 ± 6 | 25 ± 8 | 21 ± 7 | 24 ± 5 | 24 ± 6 |
| d18:1 (So) | 49,651 ± 13,923 | 50,648 ± 13,053 | 37,057 ± 9774 | 60,817 ± 34,705 | 51,751 ± 13,794 |
| d18:0 (Sa) * | 2313 ± 524 A | 10,160 ± 4784 B | 3149 ± 1191 A | 2944 ± 1075 A | 9412 ± 2650 B |
| GluSo * | 321 ± 92 AB | 301 ± 61 AB | 432 ± 127 C | 250 ± 131 A | 372 ± 82 BC |
| LysoSM | 123 ± 25 | 119 ± 20 | 137 ± 26 | 100 ± 44 | 135 ± 31 |
| Ceramides 3 | |||||
| 18:1/2:0 | 126 ± 25 | 138 ± 42 | 122 ± 24 | 134 ± 39 | 168 ± 48 |
| 18:1/14:0 * | 2404 ± 491 A | 1472 ± 396 B | 2314 ± 660 A | 2402 ± 419 A | 1536 ± 405 B |
| 18:1/16:0 * | 821,967 ± 125,521 A | 549,109 ± 70,814 B | 819,315 ± 151,516 A | 864,008 ± 109,172 A | 621,920 ± 116,635 B |
| 18:1/18:1 * | 568 ± 163 A | 762 ± 144 A | 578 ± 190 A | 711 ± 311 A | 1017 ± 317 B |
| 18:1/18:0 | 89,899 ± 16,997 | 95,785 ± 16,495 | 95,023 ± 20,222 | 96,228 ± 19,789 | 110,841 ± 30,488 |
| 18:1/20:0 | 76,599 ± 15,945 | 78,651 ± 21,306 | 74,291 ± 21,561 | 66,223 ± 17,414 | 75,575 ± 16,013 |
| 18:1/22:2 | 6468 ± 1327 | 7146 ± 2463 | 5883 ± 1546 | 6164 ± 1752 | 5694 ± 1260 |
| 18:1/22:1 | 8944 ± 2055 | 8681 ± 2202 | 8000 ± 1884 | 7466 ± 1618 | 7928 ± 1651 |
| 18:1/22:0 | 276,112 ± 56,500 | 322,875 ± 74,191 | 274,742 ± 65,610 | 256,810 ± 72,396 | 324,251 ± 65,039 |
| 18:1/:23:1 | 7601 ± 1404 | 7372 ± 1550 | 6337 ± 1494 | 6680 ± 1166 | 6972 ± 1494 |
| 18:1/23:0 * | 149,491 ± 15,447 AB | 175,097 ± 26,545 BC | 136,275 ± 28,192 A | 144,874 ± 31,886 A | 184,736 ± 3559 C |
| 18:1/24:2 | 270,944 ± 56,818 | 296,481 ± 79,986 | 250,626 ± 55,473 | 242,898 ± 55,874 | 255,533 ± 53,678 |
| 18:1/24:1 | 110,845 ± 21,795 | 113,820 ± 23,573 | 99,647 ± 19,098 | 96,379 ± 20,166 | 106,094 ± 17,652 |
| 18:1/24:0 * | 106,041 ± 14,237 AB | 121,664 ± 20,682 BC | 102,467 ± 15,256 A | 103,407 ± 22,040 A | 129,104 ± 18,446 C |
| 18:1/25:1 * | 4810 ± 787 AB | 5554 ± 1021 A | 4178 ± 939 B | 4465 ± 829 B | 5506 ± 914 A |
| 18:1/26:2 | 1675 ± 512 | 1793 ± 405 | 1491 ± 288 | 1645 ± 314 | 1685 ± 375 |
| 18:1/26:1 | 1024 ± 321 | 1048 ± 249 | 861 ± 179 | 991 ± 213 | 1093 ± 212 |
| 18:1/26:0 | 530 ± 50 | 517 ± 53 | 498 ± 39 | 546 ± 77 | 536 ± 20 |
| Dihydroceramides | |||||
| 18:0/2:0 * | 28 ± 5 A | 70 ± 37 B | 23 ± 6 A | 27 ± 3 A | 55 ± 16 B |
| 18:0/16:0 * | 39,330 ± 7838 AB | 49,331 ± 12,089 A | 34,423 ± 9441 B | 48,595 ± 13,933 A | 45,982 ± 11,738 A |
| 18:0/18:0 * | 1408 ± 275 A | 2189 ± 468 B | 1252 ± 328 A | 1564 ± 406 A | 2268 ± 605 B |
| 18:0/20:0 * | 779 ± 131 A | 867 ± 204 A | 569 ± 149 B | 704 ± 264 AB | 781 ± 199 A |
| 18:0/22:0 * | 1544 ± 298 A | 2515 ± 457 B | 1166 ± 246 A | 1427 ± 425 A | 2338 ± 531 B |
| 18:0/23:0 * | 211 ± 36 AB | 477 ± 110 C | 146 ± 41 B | 222 ± 63 A | 447 ± 94 C |
| 18:0/24:0 * | 1919 ± 355 A | 2991 ± 477 B | 1473 ± 212 C | 1938 ± 457 A | 2924 ± 446 B |
| Glycosylceramides 4 | |||||
| Glu18:1/16:0 * | 15,119 ± 3924 A | 5972 ± 2564 B | 14,059 ± 4659 A | 16,730 ± 4615 A | 7526 ± 2620 B |
| Hex18:1/18:0 | 2339 ± 627 | 2347 ± 841 | 2431 ± 948 | 2881 ± 1055 | 3094 ± 1137 |
| Hex18:1/22:0 | 15,832 ± 2737 A | 17,094 ± 5220 A | 16,269 ± 3605 A | 17,363 ± 4219 A | 21,613 ± 5417 B |
| Glu18:1/24:1 | 3146 ± 736 | 3146 ± 1232 | 2819 ± 939 | 3282 ± 896 | 3629 ± 1002 |
| Hex18:1/24:0 * | 20,973 ± 5608 A | 27,628 ± 9389 A | 21,214 ± 9553 A | 25,339 ± 9306 A | 37,192 ± 10,065 B |
| Lac18:1/16:0 * | 6602 ± 2207 AB | 3483 ± 1155 C | 6169 ± 3735 ABC | 7637 ± 4290 A | 4166 ± 1665 BC |
| Lac18:1/18:0 | 695 ± 210 | 693 ± 198 | 734 ±213 | 581 ± 160 | 841 ± 282 |
| Sphingomyelins | |||||
| SM18:1/14:0 | 212 ± 133 | 133 ± 105 | 193 ± 97 | 147 ± 66 | 128 ± 53 |
| SM18:1/16:0 * | 109,485 ± 40,754 A | 70,543 ± 26,689 C | 106,364 ± 29,354 AB | 100,271 ± 31,924 AB | 77,609 ± 23,466 BC |
| SM18:1/18:0 | 69,213 ± 35,767 | 90,806 ± 35,616 | 78,152 ± 13,186 | 66,340 ± 24,894 | 102,553 ± 45,202 |
| SM18:1/20:0 * | 25,422 ± 10,726 A | 36,254 ± 11,172 BC | 28,016 ± 4890 AB | 25,372 ± 7535 A | 40,712 ± 15,095 C |
| SM18:1/22:2 | 476 ± 174 | 558 ± 223 | 496 ± 187 | 486 ± 178 | 548 ± 226 |
| SM18:1/22:1 | 697 ± 224 | 772 ± 316 | 681 ± 185 | 647 ± 183 | 790 ± 234 |
| SM18:1/22:0 * | 264,502 ± 94,272 A | 435,724 ± 138,805 B | 295,101 ± 65,768 A | 281,556 ± 87,361 A | 460,163 ± 117,685 B |
| SM18:1/23:1 | 522 ± 291 | 701 ± 403 | 479 ± 129 | 594 ± 223 | 655 ± 307 |
| SM18:1/23:0 * | 43,430 ± 14,488 A | 80,456 ± 31,499 B | 46,794 ± 13,648 A | 46,942 ± 16,591 A | 83,112 ± 19,614 B |
| SM18:1/24:3 | 722 ± 465 | 648 ± 318 | 655 ± 293 | 734 ± 554 | 601 ± 220 |
| SM18:1/24:2 | 13,038 ± 6339 | 17,281 ± 10,583 | 13,709 ± 7828 | 12,631 ± 7168 | 17,711 ± 8021 |
| SM18:1/24:1 | 20,481 ± 8119 | 24,271 ± 14,149 | 19,771 ± 8159 | 18,890 ± 8496 | 25,992 ± 9622 |
| SM18:1/24:0 * | 42,082 ± 13,575 A | 73,438 ± 30,144 B | 47,161 ± 17,575 A | 45,702 ± 15,073 A | 79,637 ± 19,888 B |
| SM18:1/25:2 | 522 ± 144 | 676 ± 235 | 558 ± 154 | 528 ± 140 | 646 ± 137 |
| SM18:1/25:1 | 702 ± 419 | 1052 ± 800 | 680 ± 325 | 856 ± 384 | 973 ± 539 |
| SM18:1/25:0 * | 1755 ± 511 A | 2976 ± 1395 B | 1946 ± 791 A | 1955 ± 620 A | 3063 ± 782 B |
| SM18:1/26:3 | 296 ± 87 | 276 ± 31 | 274 ± 24 | 264 ± 26 | 265 ± 18 |
| SM18:1/26:2 | 532 ± 125 | 650 ± 206 | 566 ± 122 | 534 ± 126 | 626 ± 133 |
| SM18:1/26:1 | 501 ± 105 | 578 ± 196 | 506 ± 108 | 496 ± 97 | 588 ± 122 |
| SM18:1/26:0 | 663 ± 162 | 863 ± 286 | 738 ± 200 | 715 ± 155 | 925 ± 222 |
| Dihydrosphingomyelins | |||||
| SM18:0/16:0 * | 23,217 ± 7631 AB | 33,726 ± 11,917 BC | 24,714 ± 5839 AB | 21,586 ± 6263 A | 30,914 ± 7038 C |
| SM18:0/18:0 | 796 ± 858 | 1693 ± 1700 | 912 ± 982 | 1217 ± 1251 | 1302 ± 1971 |
| SM18:0/20:0 * | 2655 ± 2024 AB | 4816 ± 3387 BC | 2052 ± 1469 A | 3174 ± 1731 ABC | 5238 ± 3062 C |
| SM18:0/22:0 * | 3633 ± 1124 A | 8095 ± 2592 B | 3531 ± 783 A | 3715 ± 989 A | 7563 ± 1847 B |
| SM18:0/23:0 * | 1314 ± 296 A | 3317 ± 1014 B | 1271 ± 316 A | 1416 ± 393 A | 3035 ± 632 B |
| SM18:0/24:1 * | 553 ± 117 A | 853 ± 403 B | 528 ± 127 A | 526 ± 106 A | 788 ± 171 B |
| SM18:0/24:0 * | 1368 ± 374 A | 2719 ± 1007 B | 1283 ± 400 A | 1447 ± 396 A | 2765 ± 736 B |
| Variables | 18:1/14:0 | 18:1/16:0 | 18:1/18:0 | 18:1/20:0 | 18:1/22:0 | 18:1/23:0 | 18:1/24:0 |
|---|---|---|---|---|---|---|---|
| 18:1/14:0 | 1 | 0.827 | 0.315 | 0.076 | −0.021 | −0.035 | 0.011 |
| 18:1/16:0 | 0.827 | 1 | 0.353 | 0.145 | 0.065 | 0.054 | 0.063 |
| 18:1/18:0 | 0.315 | 0.353 | 1 | 0.526 | 0.655 | 0.668 | 0.705 |
| 18:1/20:0 | 0.076 | 0.145 | 0.526 | 1 | 0.913 | 0.630 | 0.647 |
| 18:1/22:0 | −0.021 | 0.065 | 0.655 | 0.913 | 1 | 0.856 | 0.869 |
| 18:1/23:0 | −0.035 | 0.054 | 0.668 | 0.630 | 0.856 | 1 | 0.926 |
| 18:1/24:0 | 0.011 | 0.063 | 0.705 | 0.647 | 0.869 | 0.926 | 1 |
| Variables | 18:0/16:0 | 18:0/18:0 | 18:0/20:0 | 18:0/22:0 | 18:0/23:0 | 18:0/24:0 |
|---|---|---|---|---|---|---|
| 18:0/16:0 | 1 | 0.742 | 0.480 | 0.565 | 0.614 | 0.608 |
| 18:0/18:0 | 0.742 | 1 | 0.589 | 0.869 | 0.899 | 0.902 |
| 18:0/20:0 | 0.480 | 0.589 | 1 | 0.775 | 0.538 | 0.580 |
| 18:0/22:0 | 0.565 | 0.869 | 0.775 | 1 | 0.924 | 0.920 |
| 18:0/23:0 | 0.614 | 0.899 | 0.538 | 0.924 | 1 | 0.961 |
| 18:0/24:0 | 0.608 | 0.902 | 0.580 | 0.920 | 0.961 | 1 |
| CON | FB | DON | ZEN | FDZ | |
|---|---|---|---|---|---|
| FB1 2 | <LOQ | 60.58 ± 18.18 | <LOQ | <LOQ | 60.89 ± 21.23 |
| FB2 2 | <LOQ | 5.81 ± 3.71 | <LOQ | <LOQ | 5.59 ± 2.18 |
| FB3 2 | <LOQ | 2.29 ± 0.86 | <LOQ | <LOQ | 2.56 ± 0.93 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerre, P.; Travel, A.; Tardieu, D. Targeted Analysis of Sphingolipids in Turkeys Fed Fusariotoxins: First Evidence of Key Changes That Could Help Explain Their Relative Resistance to Fumonisin Toxicity. Int. J. Mol. Sci. 2022, 23, 2512. https://doi.org/10.3390/ijms23052512
Guerre P, Travel A, Tardieu D. Targeted Analysis of Sphingolipids in Turkeys Fed Fusariotoxins: First Evidence of Key Changes That Could Help Explain Their Relative Resistance to Fumonisin Toxicity. International Journal of Molecular Sciences. 2022; 23(5):2512. https://doi.org/10.3390/ijms23052512
Chicago/Turabian StyleGuerre, Philippe, Angelique Travel, and Didier Tardieu. 2022. "Targeted Analysis of Sphingolipids in Turkeys Fed Fusariotoxins: First Evidence of Key Changes That Could Help Explain Their Relative Resistance to Fumonisin Toxicity" International Journal of Molecular Sciences 23, no. 5: 2512. https://doi.org/10.3390/ijms23052512
APA StyleGuerre, P., Travel, A., & Tardieu, D. (2022). Targeted Analysis of Sphingolipids in Turkeys Fed Fusariotoxins: First Evidence of Key Changes That Could Help Explain Their Relative Resistance to Fumonisin Toxicity. International Journal of Molecular Sciences, 23(5), 2512. https://doi.org/10.3390/ijms23052512