
Human CD4+CD45RA+ T Cells Behavior after In Vitro Activation: Modulatory Role of Vasoactive Intestinal Peptide

 , , , , , , ,
, , , , , , ,  and
and 
Abstract
1. Introduction
2. Results
2.1. CD4+CD45RA+ Cells Viability during Activation
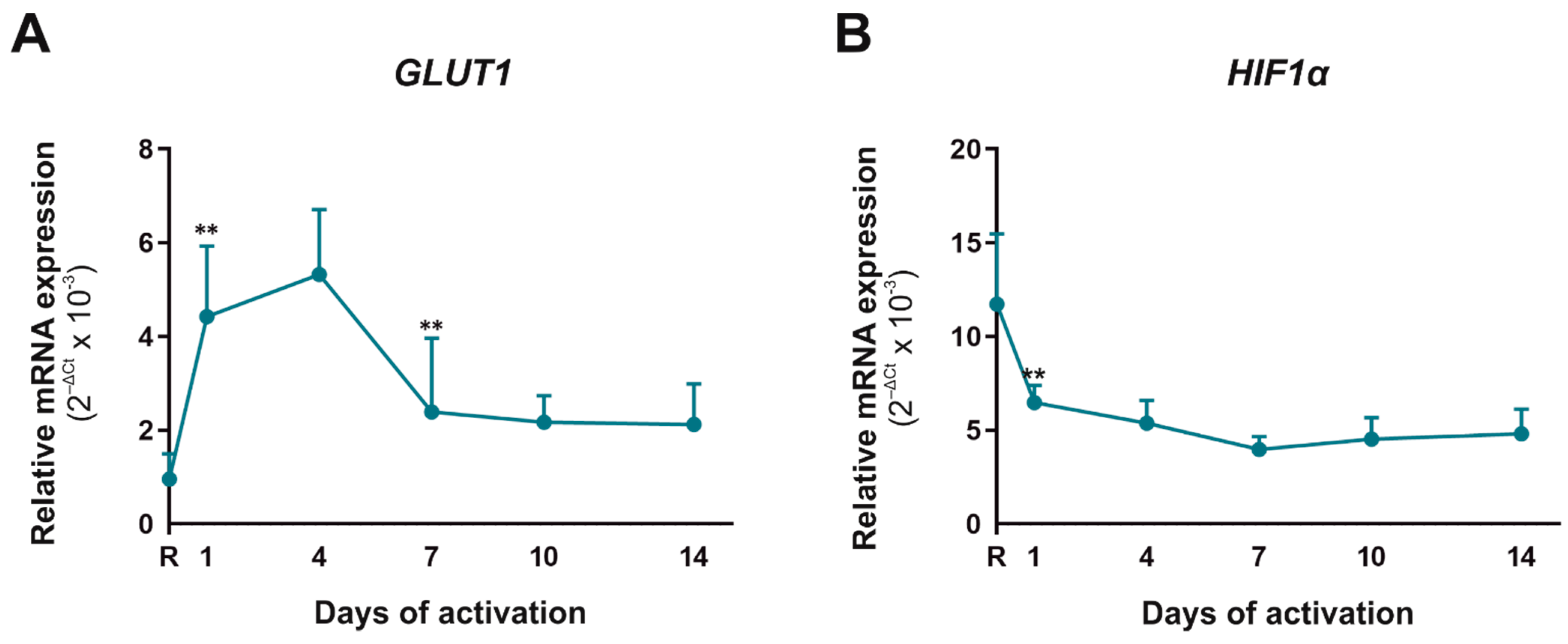
2.2. Metabolic Biomarkers in CD4+CD45RA+ Cells during Their Activation
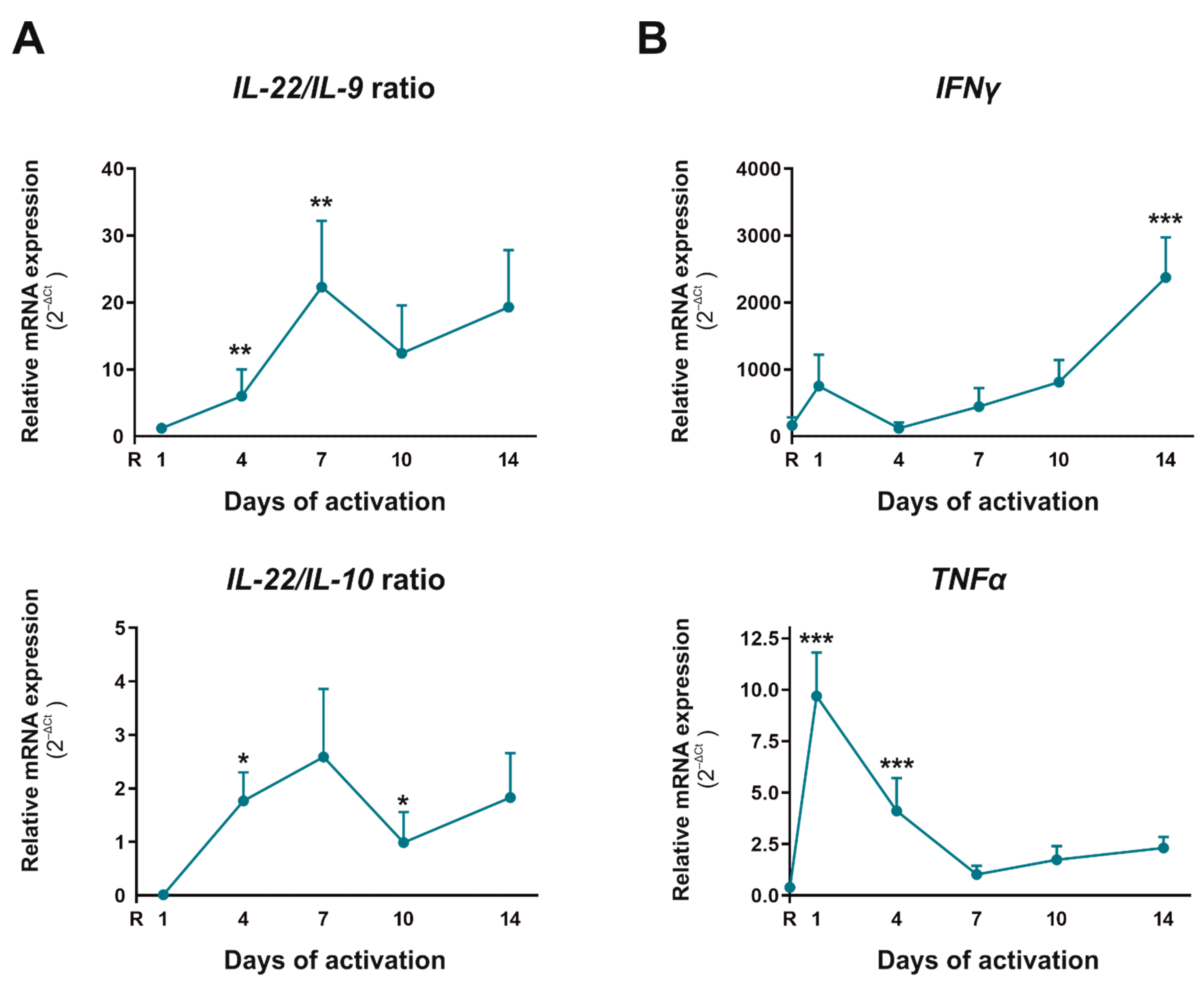
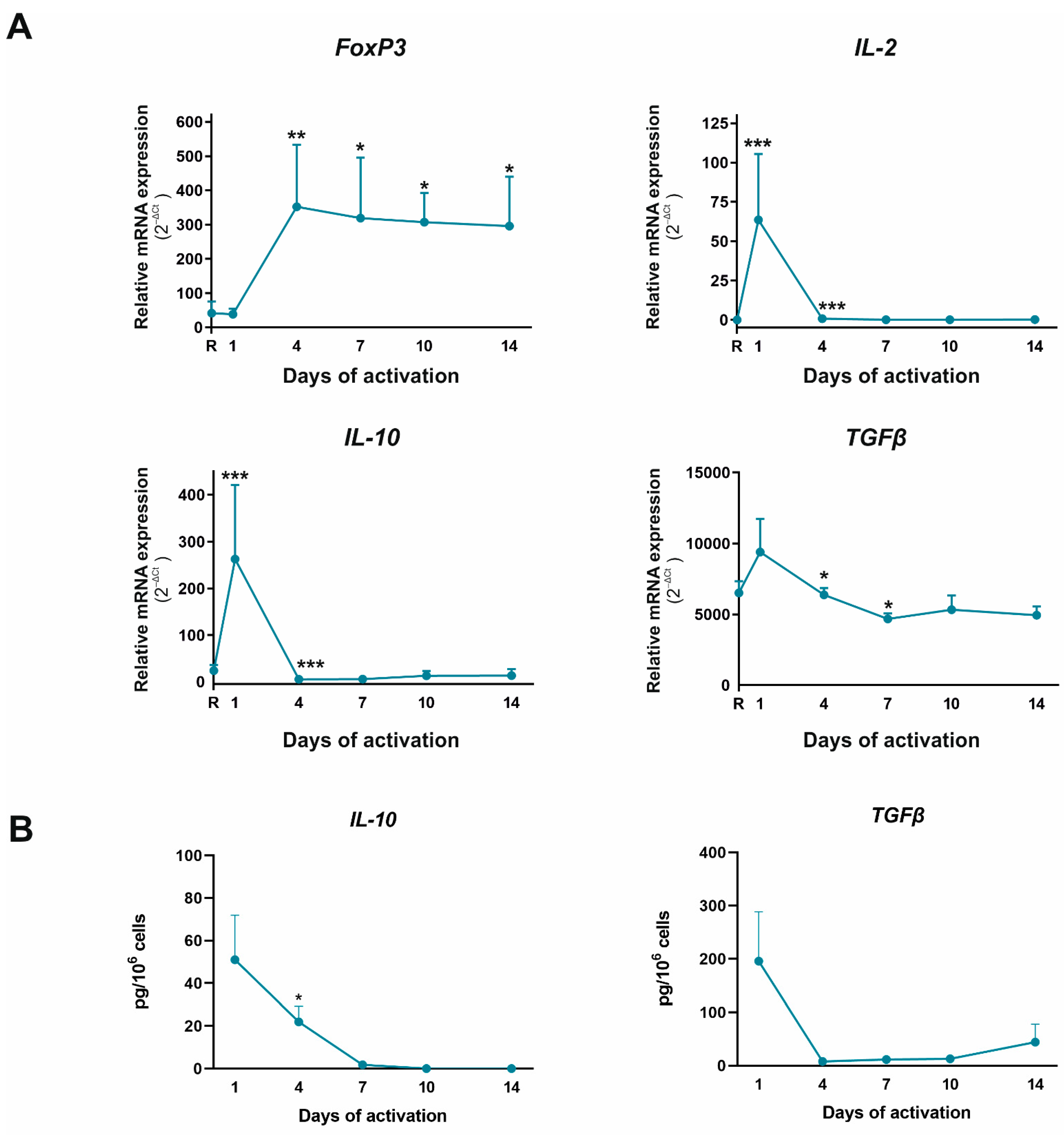
2.3. Functional Pathogenic Th17 Differentiation Profile Is the Main Subset That Appeared after Four Days of CD4+CD45RA+ Cells Activation
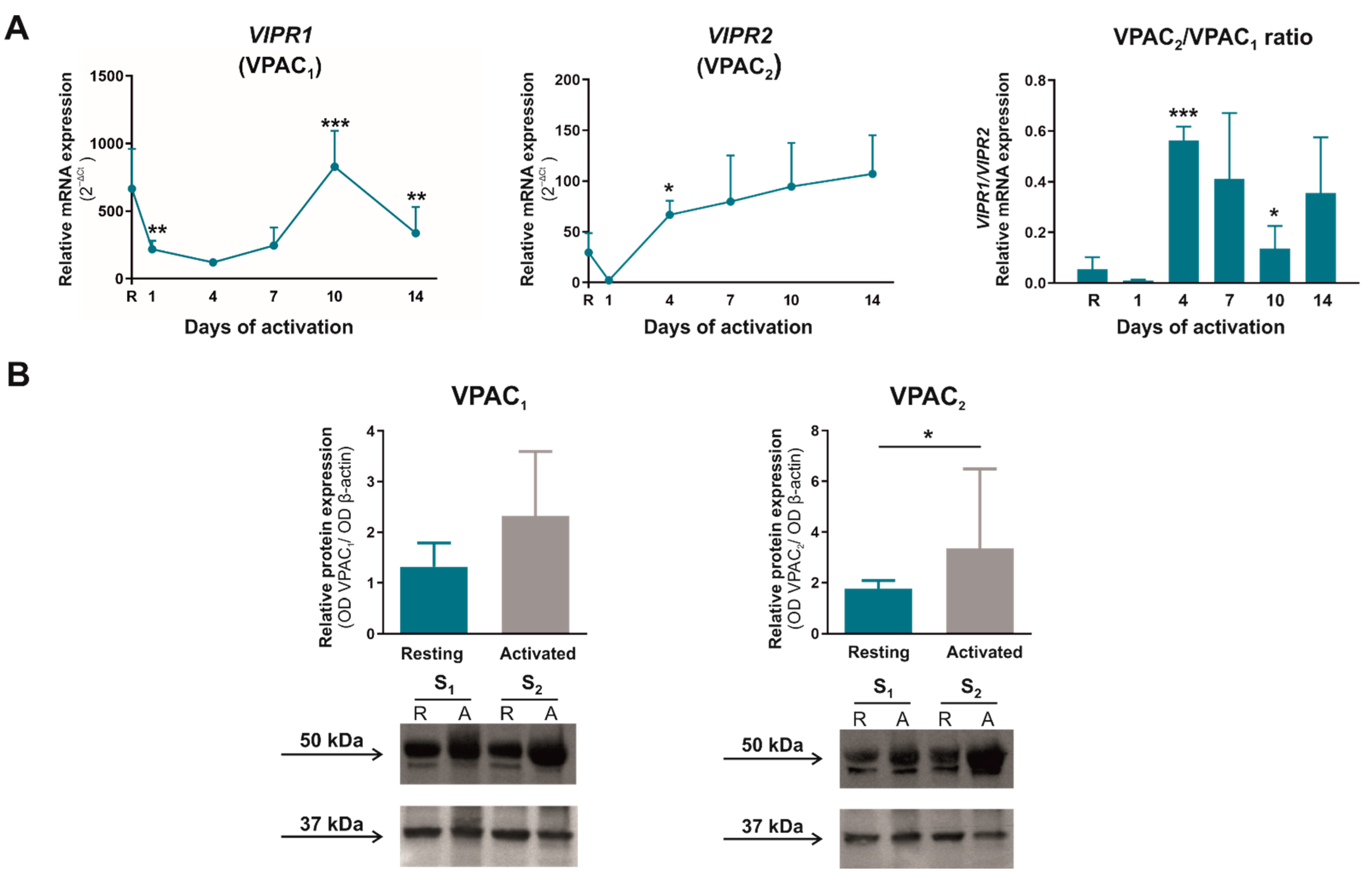
2.4. The Expression Pattern and Cellular Localization of VPAC Receptors Change in Human Activated CD4+CD45RA+ Cells
2.5. VIP Decreases Th17/1 Plasticity and Pathogenic Th17 Stabilization in Activated CD4+CD45RA+ Cells
3. Discussion
4. Materials and Methods
4.1. Healthy Donors
4.2. Isolation of Human Peripheral Blood CD4+CD45RA+ T Cells
4.3. In Vivo Activation of Human CD4+CD45RA+ T Cells
4.4. RNA Extraction and Semiquantitative Real-Time PCR
4.5. ELISA
4.6. Western Blot
4.7. Immunocytochemistry Staining
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Thome, J.J.; Yudanin, N.; Ohmura, Y.; Kubota, M.; Grinshpun, B.; Sathaliyawala, T.; Kato, T.; Lerner, H.; Shen, Y.; Farber, D.L. Spatial map of human T cell compartmentalization and maintenance over decades of life. Cell 2014, 159, 814–828. [Google Scholar] [CrossRef] [PubMed]
- van den Broek, T.; Borghans, J.A.M.; van Wijk, F. The full spectrum of human naive T cells. Nat. Rev. Immunol. 2018, 18, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Takada, K.; Jameson, S.C. Naive T cell homeostasis: From awareness of space to a sense of place. Nat. Rev. Immunol. 2009, 9, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.L.; Sousa, A.E. Establishment and Maintenance of the Human Naive CD4(+) T-Cell Compartment. Front. Pediatr. 2016, 4, 119. [Google Scholar] [CrossRef]
- Geltink, R.I.K.; Kyle, R.L.; Pearce, E.L. Unraveling the Complex Interplay between T Cell Metabolism and Function. Annu. Rev. Immunol. 2018, 36, 461–488. [Google Scholar] [CrossRef]
- Zhu, J. T Helper Cell Differentiation, Heterogeneity, and Plasticity. Cold Spring Harb. Perspect. Biol. 2018, 10, a030338. [Google Scholar] [CrossRef]
- Sallusto, F. Heterogeneity of Human CD4(+) T Cells Against Microbes. Annu. Rev. Immunol. 2016, 34, 317–334. [Google Scholar] [CrossRef]
- Zhou, L.; Chong, M.M.; Littman, D.R. Plasticity of CD4+ T cell lineage differentiation. Immunity 2009, 30, 646–655. [Google Scholar] [CrossRef]
- Ghoreschi, K.; Laurence, A.; Yang, X.P.; Hirahara, K.; O’Shea, J.J. T helper 17 cell heterogeneity and pathogenicity in autoimmune disease. Trends Immunol. 2011, 32, 395–401. [Google Scholar] [CrossRef]
- Stockinger, B.; Omenetti, S. The dichotomous nature of T helper 17 cells. Nat. Rev. Immunol. 2017, 17, 535–544. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, Z.; Qin, L.; Wang, T.; Bai, O. Insights into the mechanisms of Th17 differentiation and the Yin-Yang of Th17 cells in human diseases. Mol. Immunol. 2021, 134, 109–117. [Google Scholar] [CrossRef]
- Hirahara, K.; Vahedi, G.; Ghoreschi, K.; Yang, X.P.; Nakayamada, S.; Kanno, Y.; O’Shea, J.J.; Laurence, A. Helper T-cell differentiation and plasticity: Insights from epigenetics. Immunology 2011, 134, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Shen-Orr, S.S.; Furman, D.; Kidd, B.A.; Hadad, F.; Lovelace, P.; Huang, Y.W.; Rosenberg-Hasson, Y.; Mackey, S.; Grisar, F.A.; Pickman, Y.; et al. Defective Signaling in the JAK-STAT Pathway Tracks with Chronic Inflammation and Cardiovascular Risk in Aging Humans. Cell Syst. 2016, 3, 374–384.e4. [Google Scholar] [CrossRef] [PubMed]
- Moskowitz, D.M.; Zhang, D.W.; Hu, B.; Le Saux, S.; Yanes, R.E.; Ye, Z.; Buenrostro, J.D.; Weyand, C.M.; Greenleaf, W.J.; Goronzy, J.J. Epigenomics of human CD8 T cell differentiation and aging. Sci. Immunol. 2017, 2, eaag0192. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Yu, M.; Lee, W.W.; Tsang, M.; Krishnan, E.; Weyand, C.M.; Goronzy, J.J. Decline in miR-181a expression with age impairs T cell receptor sensitivity by increasing DUSP6 activity. Nat. Med. 2012, 18, 1518–1524. [Google Scholar] [CrossRef]
- Gomariz, R.P.; Juarranz, Y.; Abad, C.; Arranz, A.; Leceta, J.; Martinez, C. VIP-PACAP system in immunity: New insights for multitarget therapy. Ann. N. Y. Acad. Sci. 2006, 1070, 51–74. [Google Scholar] [CrossRef]
- Gomariz, R.P.; Juarranz, Y.; Carrion, M.; Perez-Garcia, S.; Villanueva-Romero, R.; Gonzalez-Alvaro, I.; Gutierrez-Canas, I.; Lamana, A.; Martinez, C. An Overview of VPAC Receptors in Rheumatoid Arthritis: Biological Role and Clinical Significance. Front. Endocrinol. 2019, 10, 729. [Google Scholar] [CrossRef]
- Villanueva-Romero, R.; Gutierrez-Canas, I.; Carrion, M.; Perez-Garcia, S.; Seoane, I.V.; Martinez, C.; Gomariz, R.P.; Juarranz, Y. The Anti-Inflammatory Mediator, Vasoactive Intestinal Peptide, Modulates the Differentiation and Function of Th Subsets in Rheumatoid Arthritis. J. Immunol. Res. 2018, 2018, 6043710. [Google Scholar] [CrossRef]
- Martinez, C.; Juarranz, Y.; Gutierrez-Canas, I.; Carrion, M.; Perez-Garcia, S.; Villanueva-Romero, R.; Castro, D.; Lamana, A.; Mellado, M.; Gonzalez-Alvaro, I.; et al. A Clinical Approach for the Use of VIP Axis in Inflammatory and Autoimmune Diseases. Int. J. Mol. Sci. 2019, 21, 65. [Google Scholar] [CrossRef]
- Juarranz, Y.; Gutierrez-Canas, I.; Santiago, B.; Carrion, M.; Pablos, J.L.; Gomariz, R.P. Differential expression of vasoactive intestinal peptide and its functional receptors in human osteoarthritic and rheumatoid synovial fibroblasts. Arthritis Rheum. 2008, 58, 1086–1095. [Google Scholar] [CrossRef]
- Carrion, M.; Perez-Garcia, S.; Martinez, C.; Juarranz, Y.; Estrada-Capetillo, L.; Puig-Kroger, A.; Gomariz, R.P.; Gutierrez-Canas, I. VIP impairs acquisition of the macrophage proinflammatory polarization profile. J. Leukoc. Biol. 2016, 100, 1385–1393. [Google Scholar] [CrossRef] [PubMed]
- Villanueva-Romero, R.; Lamana, A.; Flores-Santamaria, M.; Carrion, M.; Perez-Garcia, S.; Triguero-Martinez, A.; Tomero, E.; Criado, G.; Pablos, J.L.; Gonzalez-Alvaro, I.; et al. Comparative Study of Senescent Th Biomarkers in Healthy Donors and Early Arthritis Patients. Analysis of VPAC Receptors and Their Influence. Cells 2020, 9, 2592. [Google Scholar] [CrossRef] [PubMed]
- Delgado, M.; Martinez, C.; Leceta, J.; Garrido, E.; Gomariz, R.P. Differential VIP and VIP1 receptor gene expression in rat thymocyte subsets. Peptides 1996, 17, 803–807. [Google Scholar] [CrossRef]
- Lara-Marquez, M.; O’Dorisio, M.; O’Dorisio, T.; Shah, M.; Karacay, B. Selective gene expression and activation-dependent regulation of vasoactive intestinal peptide receptor type 1 and type 2 in human T cells. J. Immunol. 2001, 166, 2522–2530. [Google Scholar] [CrossRef]
- Vomhof-DeKrey, E.E.; Hermann, R.J.; Palmer, M.F.; Benton, K.D.; Sandy, A.R.; Dorsam, S.T.; Dorsam, G.P. TCR signaling and environment affect vasoactive intestinal peptide receptor-1 (VPAC-1) expression in primary mouse CD4 T cells. Brain Behav. Immun. 2008, 22, 1032–1040. [Google Scholar] [CrossRef] [PubMed]
- Martinez, C.; Abad, C.; Delgado, M.; Arranz, A.; Juarranz, M.G.; Rodriguez-Henche, N.; Brabet, P.; Leceta, J.; Gomariz, R.P. Anti-inflammatory role in septic shock of pituitary adenylate cyclase-activating polypeptide receptor. Proc. Natl. Acad. Sci. USA 2002, 99, 1053–1058. [Google Scholar] [CrossRef]
- Khodabakhsh, P.; Asgari Taei, A.; Mohseni, M.; Bahrami Zanjanbar, D.; Khalili, H.; Masoumi, K.; Haji Abbas Shirazi, A.; Dargahi, L. Vasoactive Peptides: Their Role in COVID-19 Pathogenesis and Potential Use as Biomarkers and Therapeutic Targets. Arch Med. Res 2021, 52, 777–787. [Google Scholar] [CrossRef]
- ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT04311697 (accessed on 30 November 2021).
- MacIver, N.J.; Michalek, R.D.; Rathmell, J.C. Metabolic regulation of T lymphocytes. Annu. Rev. Immunol. 2013, 31, 259–283. [Google Scholar] [CrossRef]
- Ikejiri, A.; Nagai, S.; Goda, N.; Kurebayashi, Y.; Osada-Oka, M.; Takubo, K.; Suda, T.; Koyasu, S. Dynamic regulation of Th17 differentiation by oxygen concentrations. Int. Immunol. 2012, 24, 137–146. [Google Scholar] [CrossRef]
- Jacobs, S.R.; Herman, C.E.; Maciver, N.J.; Wofford, J.A.; Wieman, H.L.; Hammen, J.J.; Rathmell, J.C. Glucose uptake is limiting in T cell activation and requires CD28-mediated Akt-dependent and independent pathways. J. Immunol. 2008, 180, 4476–4486. [Google Scholar] [CrossRef]
- Hebel, K.; Weinert, S.; Kuropka, B.; Knolle, J.; Kosak, B.; Jorch, G.; Arens, C.; Krause, E.; Braun-Dullaeus, R.C.; Brunner-Weinzierl, M.C. CD4+ T cells from human neonates and infants are poised spontaneously to run a nonclassical IL-4 program. J Immunol. 2014, 192, 5160–5170. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.K.; Gracias, D.T.; Croft, M. TNF activity and T cells. Cytokine 2018, 101, 14–18. [Google Scholar] [CrossRef]
- Ross, S.H.; Cantrell, D.A. Signaling and Function of Interleukin-2 in T Lymphocytes. Annu. Rev. Immunol. 2018, 36, 411–433. [Google Scholar] [CrossRef]
- Zhu, J.; Paul, W.E. Peripheral CD4+ T-cell differentiation regulated by networks of cytokines and transcription factors. Immunol. Rev. 2010, 238, 247–262. [Google Scholar] [CrossRef] [PubMed]
- Trinchieri, G. Interleukin-10 production by effector T cells: Th1 cells show self control. J. Exp. Med. 2007, 204, 239–243. [Google Scholar] [CrossRef]
- Saraiva, M.; Christensen, J.R.; Veldhoen, M.; Murphy, T.L.; Murphy, K.M.; O’Garra, A. Interleukin-10 production by Th1 cells requires interleukin-12-induced STAT4 transcription factor and ERK MAP kinase activation by high antigen dose. Immunity 2009, 31, 209–219. [Google Scholar] [CrossRef]
- Nistala, K.; Adams, S.; Cambrook, H.; Ursu, S.; Olivito, B.; de Jager, W.; Evans, J.G.; Cimaz, R.; Bajaj-Elliott, M.; Wedderburn, L.R. Th17 plasticity in human autoimmune arthritis is driven by the inflammatory environment. Proc. Natl. Acad. Sci. USA 2010, 107, 14751–14756. [Google Scholar] [CrossRef] [PubMed]
- Jimeno, R.; Gomariz, R.P.; Garin, M.; Gutierrez-Canas, I.; Gonzalez-Alvaro, I.; Carrion, M.; Galindo, M.; Leceta, J.; Juarranz, Y. The pathogenic Th profile of human activated memory Th cells in early rheumatoid arthritis can be modulated by VIP. J. Mol. Med. (Berl) 2015, 93, 457–467. [Google Scholar] [CrossRef]
- Martin-Orozco, N.; Chung, Y.; Chang, S.H.; Wang, Y.H.; Dong, C. Th17 cells promote pancreatic inflammation but only induce diabetes efficiently in lymphopenic hosts after conversion into Th1 cells. Eur. J. Immunol. 2009, 39, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Zhang, S.; Huang, Y.; Wu, Y.; Lu, Y.; Zhong, S.; Liu, X.; Wang, Y.; Li, Y.; Sun, J.; et al. Evidence for Associations Between Th1/Th17 “Hybrid” Phenotype and Altered Lipometabolism in Very Severe Graves Orbitopathy. J. Clin. Endocrinol. Metab. 2020, 105, 1851–1867. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.G.; Wang, J.H.; Gray, J.D.; Soucier, H.; Horwitz, D.A. Natural and induced CD4+CD25+ cells educate CD4+CD25- cells to develop suppressive activity: The role of IL-2, TGF-beta, and IL-10. J. Immunol. 2004, 172, 5213–5221. [Google Scholar] [CrossRef]
- Zheng, S.G.; Wang, J.; Wang, P.; Gray, J.D.; Horwitz, D.A. IL-2 is essential for TGF-beta to convert naive CD4+CD25- cells to CD25+Foxp3+ regulatory T cells and for expansion of these cells. J. Immunol. 2007, 178, 2018–2027. [Google Scholar] [CrossRef]
- Ali, A.J.; Makings, J.; Ley, K. Regulatory T Cell Stability and Plasticity in Atherosclerosis. Cells 2020, 9, 2665. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Perez, R.; Lopez-Santalla, M.; Sanchez-Dominguez, R.; Alberquilla, O.; Gutierrez-Canas, I.; Juarranz, Y.; Bueren, J.A.; Garin, M.I. Enhanced Susceptibility of Galectin-1 Deficient Mice to Experimental Colitis. Front. Immunol. 2021, 12, 687443. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Lee, W.W.; Kim, S.H.; Kang, Y.; Lee, N.; Shin, M.S.; Kang, S.W.; Kang, I. Age-associated alteration in naive and memory Th17 cell response in humans. Clin. Immunol. 2011, 140, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Li, G.; Ye, Z.; Gustafson, C.E.; Tian, L.; Weyand, C.M.; Goronzy, J.J. Transcription factor networks in aged naive CD4 T cells bias lineage differentiation. Aging Cell 2019, 18, e12957. [Google Scholar] [CrossRef] [PubMed]
- Davignon, J.L.; Rauwel, B.; Degboe, Y.; Constantin, A.; Boyer, J.F.; Kruglov, A.; Cantagrel, A. Modulation of T-cell responses by anti-tumor necrosis factor treatments in rheumatoid arthritis: A review. Arthritis Res. Ther. 2018, 20, 229. [Google Scholar] [CrossRef]
- Jimeno, R.; Leceta, J.; Garin, M.; Ortiz, A.M.; Mellado, M.; Rodriguez-Frade, J.M.; Martinez, C.; Perez-Garcia, S.; Gomariz, R.P.; Juarranz, Y. Th17 polarization of memory Th cells in early arthritis: The vasoactive intestinal peptide effect. J. Leukoc. Biol. 2015, 98, 257–269. [Google Scholar] [CrossRef]
- Gomariz, R.P.; Arranz, A.; Abad, C.; Torroba, M.; Martinez, C.; Rosignoli, F.; Garcia-Gomez, M.; Leceta, J.; Juarranz, Y. Time-course expression of Toll-like receptors 2 and 4 in inflammatory bowel disease and homeostatic effect of VIP. J. Leukoc. Biol. 2005, 78, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Jimeno, R.; Leceta, J.; Martinez, C.; Gutierrez-Canas, I.; Carrion, M.; Perez-Garcia, S.; Garin, M.; Mellado, M.; Gomariz, R.P.; Juarranz, Y. Vasoactive intestinal peptide maintains the nonpathogenic profile of human th17-polarized cells. J. Mol. Neurosci. 2014, 54, 512–525. [Google Scholar] [CrossRef]
- Villanueva-Romero, R.; Gutierrez-Canas, I.; Carrion, M.; Gonzalez-Alvaro, I.; Rodriguez-Frade, J.M.; Mellado, M.; Martinez, C.; Gomariz, R.P.; Juarranz, Y. Activation of Th lymphocytes alters pattern expression and cellular location of VIP receptors in healthy donors and early arthritis patients. Sci. Rep. 2019, 9, 7383. [Google Scholar] [CrossRef] [PubMed]
- Carrion, M.; Ramos-Levi, A.M.; Seoane, I.V.; Martinez-Hernandez, R.; Serrano-Somavilla, A.; Castro, D.; Juarranz, Y.; Gonzalez-Alvaro, I.; Gomariz, R.P.; Marazuela, M. Vasoactive intestinal peptide axis is dysfunctional in patients with Graves’ disease. Sci. Rep. 2020, 10, 13018. [Google Scholar] [CrossRef] [PubMed]
- Bhosle, V.K.; Rivera, J.C.; Chemtob, S. New insights into mechanisms of nuclear translocation of G-protein coupled receptors. Small GTPases 2019, 10, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Magalhaes, A.C.; Dunn, H.; Ferguson, S.S. Regulation of GPCR activity, trafficking and localization by GPCR-interacting proteins. Br. J. Pharmacol. 2012, 165, 1717–1736. [Google Scholar] [CrossRef]
- Valdehita, A.; Bajo, A.M.; Fernandez-Martinez, A.B.; Arenas, M.I.; Vacas, E.; Valenzuela, P.; Ruiz-Villaespesa, A.; Prieto, J.C.; Carmena, M.J. Nuclear localization of vasoactive intestinal peptide (VIP) receptors in human breast cancer. Peptides 2010, 31, 2035–2045. [Google Scholar] [CrossRef]
- Yu, R.; Liu, H.; Peng, X.; Cui, Y.; Song, S.; Wang, L.; Zhang, H.; Hong, A.; Zhou, T. The palmitoylation of the N-terminal extracellular Cys37 mediates the nuclear translocation of VPAC1 contributing to its anti-apoptotic activity. Oncotarget 2017, 8, 42728–42741. [Google Scholar] [CrossRef]
- Luttrell, L.M.; Gesty-Palmer, D. Beyond desensitization: Physiological relevance of arrestin-dependent signaling. Pharmacol. Rev. 2010, 62, 305–330. [Google Scholar] [CrossRef]
- Calebiro, D.; Godbole, A. Internalization of G-protein-coupled receptors: Implication in receptor function, physiology and diseases. Best Pract. Res. Clin. Endocrinol. Metab. 2018, 32, 83–91. [Google Scholar] [CrossRef]
- Goetzl, E.J. Hypothesis: VPAC G protein-coupled receptors for vasoactive intestinal peptide constitute a dynamic system for signaling T cells from plasma membrane and nuclear membrane complexes. Regul. Pept. 2006, 137, 75–78. [Google Scholar] [CrossRef]
- Jong, Y.I.; Harmon, S.K.; O’Malley, K.L. Location and Cell-Type-Specific Bias of Metabotropic Glutamate Receptor, mGlu5, Negative Allosteric Modulators. ACS Chem. Neurosci. 2019, 10, 4558–4570. [Google Scholar] [CrossRef]
- Calebiro, D.; Nikolaev, V.O.; Lohse, M.J. Imaging of persistent cAMP signaling by internalized G protein-coupled receptors. J Mol. Endocrinol. 2010, 45, 1–8. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
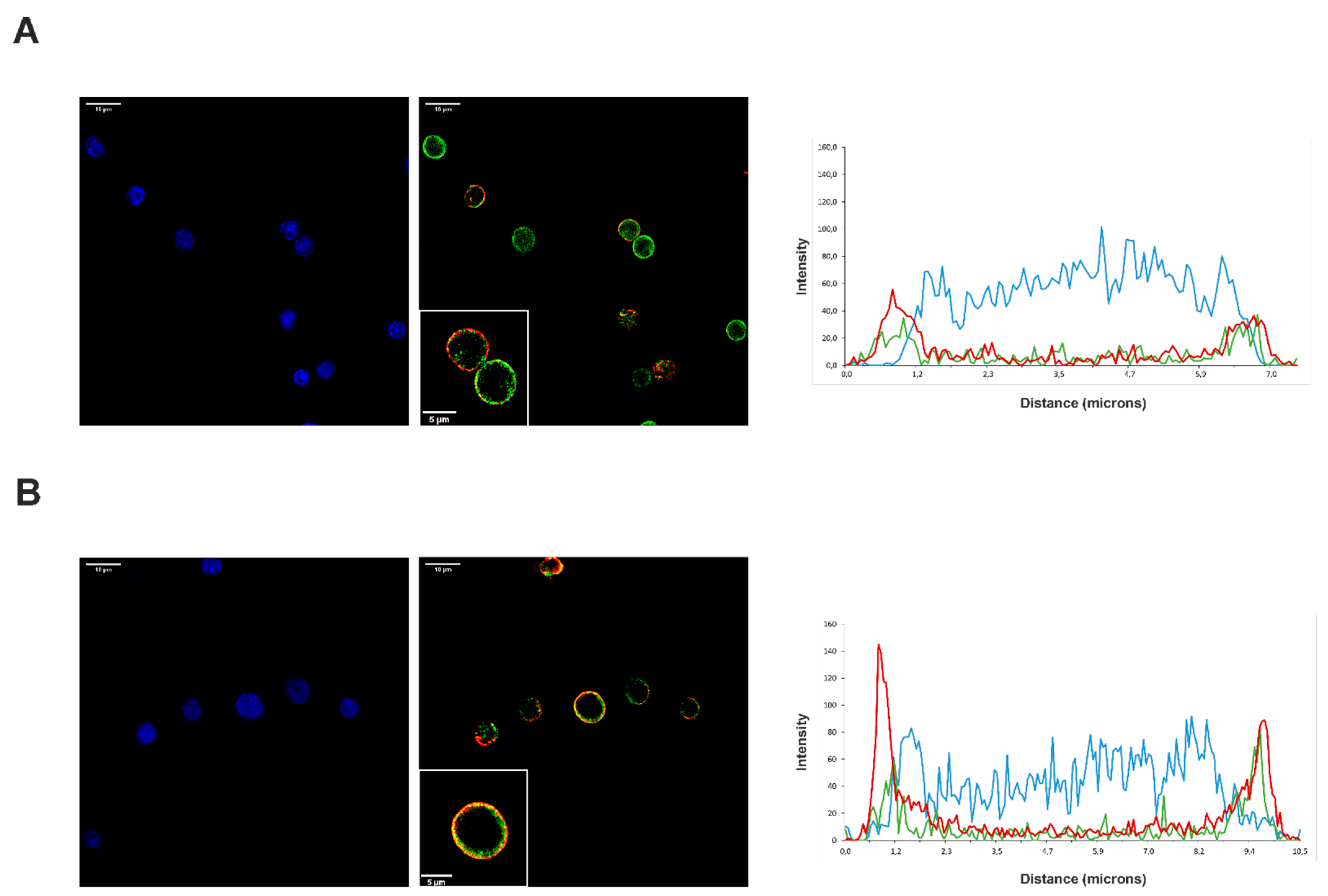{kind=link}
| Concentration Cells/mL × 106 | Viability % | |
|---|---|---|
| Day 1 | 0.19 ± 0.04 | 64.3 ± 11.2 |
| Day 4 | 1.2 ± 0.12 | 81.3 ± 3.8 |
| Day 7 | 2.53 ± 0.28 | 74.3 ± 7.8 |
| Day 10 | 1.75 ± 0.26 | 50.8 ± 9.8 |
| Day 14 | 1.15 ± 0.45 | 28.3 ± 9.4 |
| IFNγ | Activated | Activated + VIP | Activated + VPAC1 Agonist | Activated + VPAC2 Agonist |
|---|---|---|---|---|
| Resting | 166 ± 67.7 | |||
| Day 1 | 750 ± 236 | 209 ± 58.8 ** | 420 ± 98.3 | 489 ± 143 |
| Day 4 | 122 ± 43.9 | 33.6 ± 10.5 * | 64.4 ± 14.3 | 60.7 ± 8.0 |
| Day 7 | 446 ± 124 | 123 ± 21.9 ** | 203 ± 15.5 * | 175 ± 59.9 * |
| Day 10 | 810 ± 148 | 185 ± 51.9 ** | 369 ± 102 * | 388 ± 102 * |
| Day 14 | 2375 ± 244 | 735 ± 118 ** | 1097 ± 98.9 * | 425 ± 211 * |
| IL-23R | Activated | Activated + VIP | Activated + VPAC1 Agonist | Activated+ VPAC2 Agonist |
| Resting | 0.011 ± 0.002 | |||
| Day 1 | 0.154 ± 0.079 | 0.123 ± 0.065 | 0.074 ± 0.013 | 0.144 ± 0.076 |
| Day 4 | 0.111 ± 0.024 | 0.064 ± 0.009 ** | 0.075 ± 0.001 | 0.063 ± 0.016 |
| Day 7 | 0.136 ± 0.054 | 0.054 ± 0.008 ** | 0.100 ± 0.017 * | 0.090 ± 0.017 ** |
| Day 10 | 0.178 ± 0.016 | 0.115 ± 0.017 ** | 0.122 ± 0.019 | 0.119 ± 0.015 ** |
| Day 14 | 0.157 ± 0.018 | 0.171 ± 0.014 | 0.156 ± 0.015 | 0.205 ± 0.014 |
| Gene 1 | GeneBank Accession No. 2 | Sequence Position/Assay Location (TaqMan®) 3 | Sequence/Assay ID (TaqMan®) 4 |
|---|---|---|---|
| Foxp3 | NM_001114377.1 | 899 | Hs01085834_m1 |
| HIF-1α | NM_001243084.1 | 757 | Hs00153153_m1 |
| IFNG (IFNγ) | NM_000619.2 | 495 | Hs00989291_m1 |
| IL-2 | NM_000586.3 | 267 | Hs00174114_m1 |
| IL-9 | NM_000590.1 | 325 | Hs00914237_m1 |
| IL-10 | NM_000572.2 | 510 | Hs00961622_m1 |
| IL-22 | NM_020525.4 | 445 | Hs01574154_m1 |
| IL-23R | NM_144701.2 | 1037 | Hs00332759_m1 |
| RORA (RORα) | NM_002943.3 | 1173 | Hs00536545_m1 |
| RORC (RORγt) | NM_005060.3 | 182 | Hs00172860_m1 |
| SDHA | NM_001294332.1 | 757 | Hs00188166_m1 |
| SLC2A1 (GLUT1) | NM_006516.2 | Hs00892681_m1 | |
| STAT1 | NM_007315.3 | 1832 | Hs01013996_m1 |
| STAT3 | NM_003150.3 | 488 | Hs01047580_m1 |
| Tbx21 (T-bet) | NM_013351.1 | 707 | Hs00203436_m1 |
| TGFβ | NM_000660.5 | 1905 | Hs00998133_m1 |
| TNFA (TNFα) | NM_000594.3 | 457 | Hs00174128_m1 |
| VIPR1 (VPAC1) | NM_004624.3 | 306 | Hs00270351_m1 |
| VIPR1 (VPAC2) | NM_003382.4 | 644 | Hs00173643_m1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villanueva-Romero, R.; Cabrera-Martín, A.; Álvarez-Corrales, E.; Carrión, M.; Pérez-García, S.; Lamana, A.; Castro-Vázquez, D.; Martínez, C.; Gomariz, R.P.; Gutiérrez-Cañas, I.; et al. Human CD4+CD45RA+ T Cells Behavior after In Vitro Activation: Modulatory Role of Vasoactive Intestinal Peptide. Int. J. Mol. Sci. 2022, 23, 2346. https://doi.org/10.3390/ijms23042346
Villanueva-Romero R, Cabrera-Martín A, Álvarez-Corrales E, Carrión M, Pérez-García S, Lamana A, Castro-Vázquez D, Martínez C, Gomariz RP, Gutiérrez-Cañas I, et al. Human CD4+CD45RA+ T Cells Behavior after In Vitro Activation: Modulatory Role of Vasoactive Intestinal Peptide. International Journal of Molecular Sciences. 2022; 23(4):2346. https://doi.org/10.3390/ijms23042346
Chicago/Turabian StyleVillanueva-Romero, Raúl, Alicia Cabrera-Martín, Emigdio Álvarez-Corrales, Mar Carrión, Selene Pérez-García, Amalia Lamana, David Castro-Vázquez, Carmen Martínez, Rosa P. Gomariz, Irene Gutiérrez-Cañas, and et al. 2022. "Human CD4+CD45RA+ T Cells Behavior after In Vitro Activation: Modulatory Role of Vasoactive Intestinal Peptide" International Journal of Molecular Sciences 23, no. 4: 2346. https://doi.org/10.3390/ijms23042346
APA StyleVillanueva-Romero, R., Cabrera-Martín, A., Álvarez-Corrales, E., Carrión, M., Pérez-García, S., Lamana, A., Castro-Vázquez, D., Martínez, C., Gomariz, R. P., Gutiérrez-Cañas, I., & Juarranz, Y. (2022). Human CD4+CD45RA+ T Cells Behavior after In Vitro Activation: Modulatory Role of Vasoactive Intestinal Peptide. International Journal of Molecular Sciences, 23(4), 2346. https://doi.org/10.3390/ijms23042346