
The Multifaceted Roles of Mast Cells in Immune Homeostasis, Infections and Cancers

 ,
,  ,
,  , , ,
, , ,  , ,
, ,  , and
, and
Abstract
1. Introduction
2. Mast Cell Pattern Recognition Receptors
3. Mast Cell Secretory Granules
4. Mast Cell Interaction with Other Mammalian Cells
5. Mast Cells in Tumor Microenvironments—From Basics to Clinics
6. The Role of Mast Cells in Tumor Progression and Regression—Preclinical Studies
7. Mast Cells in Allergic Diseases
8. Mast Cells as Key Players in Microbial Commensalism and Pathogenesis
9. Immunotherapy Targeting Mast Cells
10. Concluding Remarks
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krystel-Whittemore, M.; Dileepan, K.N.; Wood, J.G. Mast Cell: A Multi-Functional Master Cell. Front. Immunol. 2016, 6, 620. [Google Scholar] [CrossRef]
- Dahlin, J.; Hallgren, J. Mast cell progenitors: Origin, development and migration to tissues. Mol. Immunol. 2015, 63, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Voehringer, D. Protective and pathological roles of mast cells and basophils. Nat. Rev. Immunol. 2013, 13, 362–375. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, M.; Cervantes-García, D.; Córdova-Dávalos, L.E.; Pérez-Rodríguez, M.J.; Gonzalez-Espinosa, C.; Salinas, E. Responses of Mast Cells to Pathogens: Beneficial and Detrimental Roles. Front. Immunol. 2021, 12, 685865. [Google Scholar] [CrossRef]
- De Filippo, K.; Dudeck, A.; Hasenberg, M.; Nye, E.; Van Rooijen, N.; Hartmann, K.; Gunzer, M.; Roers, A.; Hogg, N. Mast cell and macrophage chemokines CXCL1/CXCL2 control the early stage of neutrophil recruitment during tissue inflammation. Blood 2013, 121, 4930–4937. [Google Scholar] [CrossRef] [PubMed]
- Palomino, D.C.T.; Marti, L.C. Chemokines and Immunity. Einstein (São Paulo) 2015, 13, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.E.; Hatfield, J.K.; Brown, M.A. New Insights into the Role of Mast Cells in Autoimmunity: Evidence for a Common Mechanism of Action? Biochim. Biophys. Acta 2012, 1822, 57–65. [Google Scholar] [CrossRef]
- Komi, D.E.A.; Shafaghat, F.; Kovanen, P.T.; Meri, S. Mast cells and complement system: Ancient interactions between components of innate immunity. Allergy 2020, 75, 2818–2828. [Google Scholar] [CrossRef]
- Gri, G.; Frossi, B.; D’Inca, F.; Danelli, L.; Betto, E.; Mion, F.; Sibilano, R.; Pucillo, C. Mast Cell: An Emerging Partner in Immune Interaction. Front. Immunol. 2012, 3, 120. [Google Scholar] [CrossRef] [PubMed]
- Pastwińska, J.; Żelechowska, P.; Walczak-Drzewiecka, A.; Brzezińska-Błaszczyk, E.; Dastych, J. The Art of Mast Cell Adhesion. Cells 2020, 9, 2664. [Google Scholar] [CrossRef]
- Thangam, E.B.; Jemima, E.A.; Singh, H.; Baig, M.S.; Khan, M.; Mathias, C.B.; Church, M.K.; Saluja, R. The Role of Histamine and Histamine Receptors in Mast Cell-Mediated Allergy and Inflammation: The Hunt for New Therapeutic Targets. Front. Immunol. 2018, 9, 1873. [Google Scholar] [CrossRef]
- Redegeld, F.A.; Yu, Y.; Kumari, S.; Charles, N.; Blank, U. Non-IgE mediated mast cell activation. Immunol. Rev. 2018, 282, 87–113. [Google Scholar] [CrossRef] [PubMed]
- Selander, C.; Engblom, C.; Nilsson, G.; Scheynius, A.; Andersson, C.L. TLR2/MyD88-Dependent and -Independent Activation of Mast Cell IgE Responses by the Skin Commensal Yeast Malassezia sympodialis. J. Immunol. 2009, 182, 4208–4216. [Google Scholar] [CrossRef] [PubMed]
- Ifrim, D.C.; Joosten, L.A.B.; Kullberg, B.-J.; Jacobs, L.; Jansen, T.; Williams, D.L.; Gow, N.A.R.; van der Meer, J.W.M.; Netea, M.G.; Quintin, J. Candida albicansPrimes TLR Cytokine Responses through a Dectin-1/Raf-1–Mediated Pathway. J. Immunol. 2013, 190, 4129–4135. [Google Scholar] [CrossRef]
- Baorto, D.M.; Gao, Z.; Malaviya, R.; Dustin, M.; Van Der Merwe, A.; Lublin, D.M.; Abraham, S.N. Survival of FimH-expressing enterobacteria in macrophages relies on glycolipid traffic. Nature 1997, 389, 636–639. [Google Scholar] [CrossRef]
- De Zuani, M.; Paolicelli, G.; Zelante, T.; Renga, G.; Romani, L.; Arzese, A.; Pucillo, C.E.M.; Frossi, B. Mast Cells Respond to Candida albicans Infections and Modulate Macrophages Phagocytosis of the Fungus. Front. Immunol. 2018, 9, 2829. [Google Scholar] [CrossRef]
- Nieto-Patlán, A.; Campillo-Navarro, M.; Rodríguez-Cortés, O.; Muñoz-Cruz, S.; Wong-Baeza, I.; Estrada-Parra, S.; Estrada-García, I.; Serafín-López, J.; Chacón-Salinas, R. Recognition of Candida albicans by Dectin-1 induces mast cell activation. Immunobiology 2015, 220, 1093–1100. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, Y.; Kitaura, J.; Hata, D.; Yao, L.; Kawakami, T. Functions of Bruton’s tyrosine kinase in mast and B cells. J. Leukoc. Biol. 1999, 65, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Horwood, N.J.; Page, T.H.; McDaid, J.P.; Palmer, C.D.; Campbell, J.; Mahon, T.; Brennan, F.M.; Webster, D.; Foxwell, B.M.J. Bruton’s Tyrosine Kinase Is Required for TLR2 and TLR4-Induced TNF, but Not IL-6, Production. J. Immunol. 2006, 176, 3635–3641. [Google Scholar] [CrossRef]
- Krupa, A.; Fudala, R.; Florence, J.M.; Tucker, T.; Allen, T.C.; Standiford, T.J.; Luchowski, R.; Fol, M.; Rahman, M.; Gryczynski, Z.; et al. Bruton’s Tyrosine Kinase Mediates FcγRIIa/Toll-Like Receptor–4 Receptor Crosstalk in Human Neutrophils. Am. J. Respir. Cell Mol. Biol. 2013, 48, 240–249. [Google Scholar] [CrossRef]
- Takeda, K.; Akira, S. TLR signaling pathways. Semin. Immunol. 2004, 16, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Kempuraj, D.; Madhappan, B.; Christodoulou, S.; Boucher, W.; Cao, J.; Papadopoulou, N.; Cetrulo, C.L.; Theoharides, T.C. Flavonols inhibit proinflammatory mediator release, intracellular calcium ion levels and protein kinase C theta phosphorylation in human mast cells. J. Cereb. Blood Flow Metab. 2005, 145, 934–944. [Google Scholar] [CrossRef] [PubMed]
- Wernersson, S.; Pejler, G. Mast cell secretory granules: Armed for battle. Nat. Rev. Immunol. 2014, 14, 478–494. [Google Scholar] [CrossRef] [PubMed]
- Ball, D.H.; Al-Riyami, L.; Harnett, W.; Harnett, M.M. IL-33/ST2 signalling and crosstalk with FcεRI and TLR4 is targeted by the parasitic worm product, ES-62. Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef]
- Brodsky, I.E.; Monack, D. NLR-mediated control of inflammasome assembly in the host response against bacterial pathogens. Semin. Immunol. 2009, 21, 199–207. [Google Scholar] [CrossRef]
- Xie, Q.; Shen, W.-W.; Zhong, J.; Huang, C.; Zhang, L.; Li, J. Lipopolysaccharide/adenosine triphosphate induces IL-1β and IL-18 secretion through the NLRP3 inflammasome in RAW264.7 murine macrophage cells. Int. J. Mol. Med. 2014, 34, 341–349. [Google Scholar] [CrossRef]
- Piliponsky, A.M.; Acharya, M.; Shubin, N.J. Mast Cells in Viral, Bacterial, and Fungal Infection Immunity. Int. J. Mol. Sci. 2019, 20, 2851. [Google Scholar] [CrossRef]
- Honjoh, C.; Chihara, K.; Yoshiki, H.; Yamauchi, S.; Takeuchi, K.; Kato, Y.; Hida, Y.; Ishizuka, T.; Sada, K. Association of C-Type Lectin Mincle with FcεRIβγ Subunits Leads to Functional Activation of RBL-2H3 Cells through Syk. Sci. Rep. 2017, 7, 46064. [Google Scholar] [CrossRef]
- Huang, M.; Wang, X.; Xing, B.; Yang, H.; Sa, Z.; Zhang, D.; Yao, W.; Yin, N.; Xia, Y.; Ding, G. Critical roles of TRPV2 channels, histamine H1 and adenosine A1 receptors in the initiation of acupoint signals for acupuncture analgesia. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef]
- Matsushima, H.; Yamada, N.; Matsue, H.; Shimada, S. The effects of endothelin-1 on degranulation, cytokine, and growth factor production by skin-derived mast cells. Eur. J. Immunol. 2004, 34, 1910–1919. [Google Scholar] [CrossRef]
- Lee, H.; Whitfeld, P.L.; Mackay, C.R. Receptors for complement C5a. The importance of C5aR and the enigmatic role of C5L2. Immunol. Cell Biol. 2008, 86, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Gallwitz, M.; Reimer, J.M.; Hellman, L. Expansion of the mast cell chymase locus over the past 200 million years of mammalian evolution. Immunogenetics 2006, 58, 655–669. [Google Scholar] [CrossRef] [PubMed]
- Siiskonen, H.; Poukka, M.; Bykachev, A.; Tyynelä-Korhonen, K.; Sironen, R.; Pasonen-Seppänen, S.; Harvima, I.T. Low numbers of tryptase+ and chymase+ mast cells associated with reduced survival and advanced tumor stage in melanoma. Melanoma Res. 2015, 25, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Arvan, P.; Castle, D. Sorting and storage during secretory granule biogenesis: Looking backward and looking forward. Biochem. J. 1998, 332, 593–610. [Google Scholar] [CrossRef]
- Elieh Ali Komi, D.; Wöhrl, S.; Bielory, L. Mast Cell Biology at Molecular Level: A Comprehensive Review. Clin. Rev. Allergy Immunol. 2020, 58, 342–365. [Google Scholar] [CrossRef]
- Knight, P.A.; Wright, S.H.; Lawrence, C.E.; Paterson, Y.Y.; Miller, H.R. Delayed Expulsion of the Nematode Trichinella spiralisIn Mice Lacking the Mucosal Mast Cell–Specific Granule Chymase, Mouse Mast Cell Protease-1. J. Exp. Med. 2000, 192, 1849–1856. [Google Scholar] [CrossRef]
- Caughey, G.H.; Schaumberg, T.H.; Zerweck, E.H.; Butterfield, J.H.; Hanson, R.D.; Silverman, G.A.; Ley, T.J. The Human Mast Cell Chymase Gene (CMA1): Mapping to the Cathepsin G/Granzyme Gene Cluster and Lineage-Restricted Expression. Genomics 1993, 15, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Wong, G.W.; Yasuda, S.; Morokawa, N.; Li, L.; Stevens, R.L.; Yoneyama, T.; Shiozawa, M.; Nakamura, M.; Suzuki, T.; Sagane, Y.; et al. Mouse Chromosome 17A3.3 Contains 13 Genes That Encode Functional Tryptic-like Serine Proteases with Distinct Tissue and Cell Expression Patterns. J. Biol. Chem. 2004, 279, 2438–2452. [Google Scholar] [CrossRef]
- Bagher, M.; Larsson-Callerfelt, A.-K.; Rosmark, O.; Hallgren, O.; Bjermer, L.; Westergren-Thorsson, G. Mast cells and mast cell tryptase enhance migration of human lung fibroblasts through protease-activated receptor 2. Cell Commun. Signal. 2018, 16, 59. [Google Scholar] [CrossRef]
- Bulfone-Paus, S.; Bahri, R. Mast Cells as Regulators of T Cell Responses. Front. Immunol. 2015, 6, 394. [Google Scholar] [CrossRef]
- McNeil, H.P.; Adachi, R.; Stevens, R.L. Mast Cell-restricted Tryptases: Structure and Function in Inflammation and Pathogen Defense. J. Biol. Chem. 2007, 282, 20785–20789. [Google Scholar] [CrossRef]
- Tani, K.; Ogushi, F.; Kido, H.; Kawano, T.; Kunori, Y.; Kamimura, T.; Cui, P.; Sone, S. Chymase is a potent chemoattractant for human monocytes and neutrophils. J. Leukoc. Biol. 2000, 67, 585–589. [Google Scholar] [CrossRef] [PubMed]
- Caughey, G.H. MAST CELL PROTEASES AS PROTECTIVE AND INFLAMMATORY MEDIATORS. In Madame Curie Bio-Science Database [Internet]; Landes Bioscience: Austin, TX, USA, 2013; pp. 212–234. [Google Scholar]
- Giannetti, M.P.; Akin, C.; Hufdhi, R.; Hamilton, M.J.; Weller, E.; van Anrooij, B.; Lyons, J.J.; Hornick, J.L.; Pinkus, G.; Castells, M.; et al. Patients with mast cell activation symptoms and elevated baseline serum tryptase level have unique bone marrow morphology. J. Allergy Clin. Immunol. 2021, 147, 1497–1501.e1. [Google Scholar] [CrossRef]
- Moon, T.C.; Befus, A.D.; Kulka, M. Mast Cell Mediators: Their Differential Release and the Secretory Pathways Involved. Front. Immunol. 2014, 5, 569. [Google Scholar] [CrossRef] [PubMed]
- Pettersson, H.; Zarnegar, B.; Westin, A.; Persson, V.; Peuckert, C.; Jonsson, J.; Hallgren, J.; Kullander, K. SLC10A4 regulates IgE-mediated mast cell degranulation in vitro and mast cell-mediated reactions in vivo. Sci. Rep. 2017, 7, 1085. [Google Scholar] [CrossRef] [PubMed]
- Fukuishi, N.; Murakami, S.; Ohno, A.; Yamanaka, N.; Matsui, N.; Fukutsuji, K.; Yamada, S.; Itoh, K.; Akagi, M. Does β-Hexosaminidase Function Only as a Degranulation Indicator in Mast Cells? The Primary Role of β-Hexosaminidase in Mast Cell Granules. J. Immunol. 2014, 193, 1886–1894. [Google Scholar] [CrossRef]
- Nakamura, Y.; Ishimaru, K.; Shibata, S.; Nakao, A. Regulation of plasma histamine levels by the mast cell clock and its modulation by stress. Sci. Rep. 2017, 7, 39934. [Google Scholar] [CrossRef]
- Bachelet, I.; Levi-Schaffer, F. Mast cells as effector cells: A co-stimulating question. Trends Immunol. 2007, 28, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Abraham, S.N.; John, A.L.S. Mast cell-orchestrated immunity to pathogens. Nat. Rev. Immunol. 2010, 10, 440–452. [Google Scholar] [CrossRef]
- Kurashima, Y.; Kiyono, H.; Kunisawa, J. Pathophysiological Role of Extracellular Purinergic Mediators in the Control of Intestinal Inflammation. Mediators Inflamm. 2015, 2015, 427125. [Google Scholar] [CrossRef]
- Rudich, N.; Ravid, K.; Sagi-Eisenberg, R. Mast Cell Adenosine Receptors Function: A Focus on the A3 Adenosine Receptor and Inflammation. Front. Immunol. 2012, 3, 134. [Google Scholar] [CrossRef] [PubMed]
- Sumpter, T.L.; Balmert, S.C.; Kaplan, D.H. Cutaneous immune responses mediated by dendritic cells and mast cells. JCI Insight 2019, 4. [Google Scholar] [CrossRef]
- Varricchi, G.; Rossi, F.W.; Galdiero, M.R.; Granata, F.; Criscuolo, G.; Spadaro, G.; De Paulis, A.; Marone, G. Physiological Roles of Mast Cells: Collegium Internationale Allergologicum Update 2019. Int. Arch. Allergy Immunol. 2019, 179, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Varricchi, G.; Galdiero, M.R.; Loffredo, S.; Marone, G.; Iannone, R.; Marone, G.; Granata, F. Are Mast Cells MASTers in Cancer? Front. Immunol. 2017, 8, 00424. [Google Scholar] [CrossRef]
- Galdiero, M.R.; Varricchi, G.; Seaf, M.; Marone, G.G.; Levi-Schaffer, F.; Marone, G.G. Bidirectional Mast Cell-Eosinophil Interactions in Inflammatory Disorders and Cancer. Front. Med. 2017, 4, 103. [Google Scholar] [CrossRef]
- Frossi, B.; Mion, F.; Tripodo, C.; Colombo, M.P.; Pucillo, C.E. Rheostatic Functions of Mast Cells in the Control of Innate and Adaptive Immune Responses. Trends Immunol. 2017, 38, 648–656. [Google Scholar] [CrossRef]
- Jimi, S.; Takebayashi, S.; Kawamoto, N. Cutaneous malignant melanoma: Correlation between neovascularization and peritumor accumulation of mast cells overexpressing vascular endothelial growth factor. Hum. Pathol. 2000, 31, 955–960. [Google Scholar] [CrossRef]
- Siebenhaar, F.; Redegeld, F.A.; Bischoff, S.C.; Gibbs, B.F.; Maurer, M. Mast Cells as Drivers of Disease and Therapeutic Targets. Trends Immunol. 2017, 39, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Giustizieri, M.L.; Albanesi, C.; Fluhr, J.; Gisondi, P.; Norgauer, J.; Girolomoni, G. H1 histamine receptor mediates inflammatory responses in human keratinocytes. J. Allergy Clin. Immunol. 2004, 114, 1176–1182. [Google Scholar] [CrossRef]
- Eissmann, M.F.; Dijkstra, C.; Jarnicki, A.; Phesse, T.; Brunnberg, J.; Poh, A.; Etemadi, N.; Tsantikos, E.; Thiem, S.; Huntington, N.; et al. IL-33-mediated mast cell activation promotes gastric cancer through macrophage mobilization. Nat. Commun. 2019, 10, 1–16. [Google Scholar] [CrossRef]
- Porta, C.; Kumar, B.S.; Larghi, P.; Rubino, L.; Mancino, A.; Sica, A. Tumor Promotion by Tumor-Associated Macrophages. Adv. Mol. Oncol. 2007, 604, 67–86. [Google Scholar] [CrossRef]
- Ribatti, D. Mast cells and macrophages exert beneficial and detrimental effects on tumor progression and angiogenesis. Immunol. Lett. 2013, 152, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Komi, D.E.A.; Redegeld, F.A. Role of Mast Cells in Shaping the Tumor Microenvironment. Clin. Rev. Allergy Immunol. 2019, 58, 313–325. [Google Scholar] [CrossRef]
- Iliev, I.D.; Leonardi, I. Fungal dysbiosis: Immunity and interactions at mucosal barriers. Nat. Rev. Immunol. 2017, 17, 635–646. [Google Scholar] [CrossRef]
- Ma, Y.; Hwang, R.F.; Logsdon, C.D.; Ullrich, S.E. Dynamic mast cell-stromal cell interactions promote growth of pancreatic cancer. Cancer Res. 2011, 23, 3927–3937. [Google Scholar] [CrossRef]
- Cardamone, C.; Parente, R.; De Feo, G.; Triggiani, M. Mast cells as effector cells of innate immunity and regulators of adaptive immunity. Immunol. Lett. 2016, 178, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Mirchandani, A.S.; Salmond, R.J.; Liew, F.Y. Interleukin-33 and the function of innate lymphoid cells. Trends Immunol. 2012, 33, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Feoktistov, I.; Ryzhov, S.; Goldstein, A.E.; Biaggioni, I. Mast Cell–Mediated Stimulation of Angiogenesis. Circ. Res. 2003, 92, 485–492. [Google Scholar] [CrossRef]
- Chang, D.Z.; Ma, Y.; Ji, B.; Wang, H.; Deng, D.; Liu, Y.; Abbruzzese, J.L.; Liu, Y.-J.; Logsdon, C.D.; Hwu, P. Mast Cells in Tumor Microenvironment Promotes the In Vivo Growth of Pancreatic Ductal Adenocarcinoma. Clin. Cancer Res. 2011, 17, 7015–7023. [Google Scholar] [CrossRef]
- Blair, R.J.; Meng, H.; Marchese, M.J.; Ren, S.; Schwartz, L.B.; Tonnesen, M.G.; Gruber, B.L. Human mast cells stimulate vascular tube formation. Tryptase is a novel, potent angiogenic factor. J. Clin. Investig. 1997, 99, 2691–2700. [Google Scholar] [CrossRef] [PubMed]
- Derakhshan, T.; Bhowmick, R.; Meinkoth, J.H.; Ritchey, J.W.; Gappa-Fahlenkamp, H. Human Mast Cell Development from Hematopoietic Stem Cells in a Connective Tissue-Equivalent Model. Tissue Eng. Part A 2019, 25, 1564–1574. [Google Scholar] [CrossRef] [PubMed]
- Kunder, C.A.; John, A.L.S.; Li, G.; Leong, K.W.; Berwin, B.; Staats, H.F.; Abraham, S.N. Mast cell–derived particles deliver peripheral signals to remote lymph nodes. J. Exp. Med. 2009, 206, 2455–2467. [Google Scholar] [CrossRef] [PubMed]
- Aponte-López, A.; Fuentes-Pananá, E.M.; Cortes-Muñoz, D.; Muñoz-Cruz, S. Mast Cell, the Neglected Member of the Tumor Microenvironment: Role in Breast Cancer. J. Immunol. Res. 2018, 2018, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, K.; Piao, H.M.; Nakadate, T.; Shikanai, T.; Nakamura, Y.; Niisato, M.; Ito, H.; Mouri, T.; Kobayashi, H.; Maesawa, C.; et al. Progress in Allergy Signal Research on Mast Cells: The Role of Histamine in Goblet Cell Hyperplasia in Allergic Airway Inflammation – A Study Using the Hdc Knockout Mouse. J. Pharmacol. Sci. 2008, 106, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Kokenyesi, R. Ovarian carcinoma cells synthesize both chondroitin sulfate and heparan sulfate cell surface proteoglycans that mediate cell adhesion to interstitial matrix. J. Cell. Biochem. 2001, 83, 259–270. [Google Scholar] [CrossRef]
- Johnson, A.R.; Hugli, E.T.; Müller-Eberhard, H.J. Release of histamine from rat mast cells by the complement peptides C3a and C5a. Immunology 1975, 28, 1067–1080. [Google Scholar] [PubMed]
- Marichal, T.; Tsai, M.; Galli, S.J. Mast Cells: Potential Positive and Negative Roles in Tumor Biology. Cancer Immunol. Res. 2013, 1, 269–279. [Google Scholar] [CrossRef]
- Toth, T.; Tóth-Jakatics, R.; Jimi, S.; Takebayashi, S. Increased Density of Interstitial Mast Cells in Amyloid A Renal Amyloidosis. Mod. Pathol. 2000, 13, 1020–1028. [Google Scholar] [CrossRef]
- Galli, S.J.; Tsai, M. IgE and mast cells in allergic disease. Nat. Med. 2012, 18, 693–704. [Google Scholar] [CrossRef]
- Ieni, A.; Barresi, V.; Branca, G.; Caruso, R.A.; Tuccari, G. Mast Cell Interaction with Neutrophils in Human Gastric Carcinomas: Ultrastructural Observations. Anal. Cell. Pathol. 2016, 2016, 1–5. [Google Scholar] [CrossRef]
- Esposito, P.; Chandler, N.; Kandere, K.; Basu, S.; Jacobson, S.; Connolly, R.; Tutor, D.; Theoharides, T.C. Corticotropin-Releasing Hormone and Brain Mast Cells Regulate Blood-Brain-Barrier Permeability Induced by Acute Stress. J. Pharmacol. Exp. Ther. 2002, 303, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Marech, I.; Leporini, C.; Ammendola, M.; Porcelli, M.; Gadaleta, C.D.; Russo, E.; De Sarro, G.; Ranieri, G. Classical and non-classical proangiogenic factors as a target of antiangiogenic therapy in tumor microenvironment. Cancer Lett. 2016, 380, 216–226. [Google Scholar] [CrossRef]
- Pejler, G.; Rönnberg, E.; Waern, I.; Wernersson, S. Mast cell proteases: Multifaceted regulators of inflammatory disease. Blood 2010, 115, 4981–4990. [Google Scholar] [CrossRef]
- Weller, C.L.; Collington, S.J.; Williams, T.; Lamb, J.R. Mast cells in health and disease. Clin. Sci. 2011, 120, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Sagi-Eisenberg, R. The mast cell: Where endocytosis and regulated exocytosis meet. Immunol. Rev. 2007, 217, 292–303. [Google Scholar] [CrossRef] [PubMed]
- Enriquez, E.M.; Hallgren, J. Mast Cells and Their Progenitors in Allergic Asthma. Front. Immunol. 2019, 10, 821. [Google Scholar] [CrossRef] [PubMed]
- Merickel, A.; Edward, R.H. Transport of Histamine by Vesicular Monoamine Transporter-2. Neuropharmacology 1995, 34, 1543–1547. [Google Scholar] [CrossRef]
- Gaudenzio, N.; Espagnolle, N.; Mars, L.T.; Liblau, R.; Valitutti, S.; Espinosa, E. Cell-cell cooperation at the T helper cell/mast cell immunological synapse. Blood 2009, 114, 4979–4988. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.; Grimbaldeston, M.; Galli, S.J. Mast Cells and Immunoregulation/Immunomodulation. In Madame Curie Bioscience Database [Internet]; Landes Bioscience: Austin, TX, USA, 2013. [Google Scholar]
- Brill, A.; Baram, D.; Sela, U.; Salamon, P.; Mekori, Y.A.; Hershkoviz, R. Induction of mast cell interactions with blood vessel wall components by direct contact with intact T cells or T cell membranes in vitro. Clin. Exp. Allergy 2004, 34, 1725–1731. [Google Scholar] [CrossRef] [PubMed]
- Rodewald, H.R.; Feyerabend, T.B. Widespread Immunological Functions of Mast Cells: Fact or Fiction? Immunity 2012, 37, 13–24. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Conti, P. Mast cells: The JEKYLL and HYDE of tumor growth. Trends Immunol. 2004, 25, 235–241. [Google Scholar] [CrossRef]
- Ellerbroek, S.M.; Hudson, L.G.; Stack, M.S. Proteinase requirements of epidermal growth factor–induced ovarian cancer cell invasion. Int. J. Cancer 1998, 78, 331–337. [Google Scholar] [CrossRef]
- Tan, S.-Y.; Fan, Y.; Luo, H.-S.; Shen, Z.-X.; Guo, Y.; Zhao, L.-J. Prognostic Significance of Cell Infiltrations of Immunosurveillance in Colorectal Cancer. World J. Gastroenterol. 2005, 11, 1210–1214. [Google Scholar] [CrossRef] [PubMed]
- Visciano, C.; Prevete, N.; Liotti, F.; Marone, G. Tumor-Associated Mast Cells in Thyroid Cancer. Int. J. Endocrinol. 2015, 2015, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chichlowski, M.; Westwood, G.S.; Abraham, S.N.; Hale, L.P. Role of Mast Cells in Inflammatory Bowel Disease and Inflammation-Associated Colorectal Neoplasia in IL-10-Deficient Mice. PLoS ONE 2010, 5, e12220. [Google Scholar] [CrossRef] [PubMed]
- Gulubova, M.; Vlaykova, T. Prognostic significance of mast cell number and microvascular density for the survival of patients with primary colorectal cancer. J. Gastroenterol. Hepatol. 2009, 24, 1265–1275. [Google Scholar] [CrossRef]
- Samoszuk, M.; Corwin, M.A. Mast cell inhibitor cromolyn increases blood clotting and hypoxia in murine breast cancer. Int. J. Cancer 2003, 107, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Fleischmann, A.; Schlomm, T.; Köllermann, J.; Sekulic, N.; Huland, H.; Mirlacher, M.; Sauter, G.; Simon, R.; Erbersdobler, A. Immunological microenvironment in prostate cancer: High mast cell densities are associated with favorable tumor characteristics and good prognosis. Prostate 2009, 69, 976–981. [Google Scholar] [CrossRef]
- Xiong, L.; Zhen, S.; Yu, Q.; Gong, Z. HCV-E2 inhibits hepatocellular carcinoma metastasis by stimulating mast cells to secrete exosomal shuttle microRNAs. Oncol. Lett. 2017, 14, 2141–2146. [Google Scholar] [CrossRef]
- Conti, P.; Castellani, M.L.; Kempuraj, D.; Salini, V.; Vecchiet, J.; Tetè, S.; Mastrangelo, F.; Perrella, A.; De Lutiis, M.A.; Tagen, M.; et al. Role of Mast Cells in Tumor Growth. Ann. Clin. Lab. Sci. 2007, 37, 315–322. [Google Scholar]
- Dubreuil, P.; Letard, S.; Ciufolini, M.; Gros, L.; Humbert, M.; Castéran, N.; Borge, L.; Hajem, B.; Lermet, A.; Sippl, W.; et al. Masitinib (AB1010), a Potent and Selective Tyrosine Kinase Inhibitor Targeting KIT. PLoS ONE 2009, 4, e7258. [Google Scholar] [CrossRef]
- Wedemeyer, J.; Galli, S.J. Decreased susceptibility of mast cell-deficient KitW/KitW-v mice to the development of 1, 2-dimethylhydrazine-induced intestinal tumors. Lab. Investig. 2005, 85, 388–396. [Google Scholar] [CrossRef]
- Rigoni, A.; Colombo, M.P.; Pucillo, C. The Role of Mast Cells in Molding the Tumor Microenvironment. Cancer Microenviron. 2014, 8, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Nonomura, N.; Takayama, H.; Nishimura, K.; Oka, D.; Nakai, Y.; Shiba, M.; Tsujimura, A.; Nakayama, M.; Aozasa, K.; Okuyama, A. Decreased number of mast cells infiltrating into needle biopsy specimens leads to a better prognosis of prostate cancer. Br. J. Cancer 2007, 97, 952–956. [Google Scholar] [CrossRef]
- Melillo, R.M.; Guarino, V.; Avilla, E.; Galdiero, M.R.; Liotti, F.; Prevete, N.; Rossi, F.W.; Basolo, F.; Ugolini, C.; de Paulis, A.; et al. Mast cells have a protumorigenic role in human thyroid cancer. Oncogene 2010, 29, 6203–6215. [Google Scholar] [CrossRef] [PubMed]
- Saleem, S.J.; Martin, R.K.; Morales, J.K.; Sturgill, J.L.; Gibb, D.R.; Graham, L.; Bear, H.D.; Manjili, M.H.; Ryan, J.J.; Conrad, D.H. Cutting Edge: Mast Cells Critically Augment Myeloid-Derived Suppressor Cell Activity. J. Immunol. 2012, 189, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Alì, G.; Boldrini, L.; Lucchi, M.; Mussi, A.; Corsi, V.; Fontanini, G. Tryptase Mast Cells in Malignant Pleural Mesothelioma as an Independent Favorable Prognostic Factor. J. Thorac. Oncol. 2009, 4, 348–354. [Google Scholar] [CrossRef]
- Galanis, E.; Hartmann, L.C.; Cliby, W.A.; Long, H.J.; Peethambaram, P.P.; Barrette, B.A.; Kaur, J.S.; Haluska, P.J., Jr.; Aderca, I.; Zollman, P.J.; et al. Phase I Trial of Intraperitoneal Administration of an Oncolytic Measles Virus Strain Engineered to Express Carcinoembryonic Antigen for Recurrent Ovarian Cancer. Cancer Res. 2010, 70, 875–882. [Google Scholar] [CrossRef]
- Chan, J.K.; Magistris, A.; Loizzi, V.; Lin, F.; Rutgers, J.; Osann, K.; DiSaia, P.J.; Samoszuk, M. Mast cell density, angiogenesis, blood clotting, and prognosis in women with advanced ovarian cancer. Gynecol. Oncol. 2005, 99, 20–25. [Google Scholar] [CrossRef]
- Elpek, G.; Gelen, T.; Aksoy, N.H.; Erdogan, A.; Dertsiz, L.; Demircan, A.; Keles, N. The prognostic relevance of angiogenesis and mast cells in squamous cell carcinoma of the oesophagus. J. Clin. Pathol. 2001, 54, 940–944. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, N.; Pagliaro, L. Sequential pathogenesis of metastatic VHL mutant clear cell renal cell carcinoma: Putting it together with a translational perspective. Ann. Oncol. 2016, 27, 1685–1695. [Google Scholar] [CrossRef]
- Wang, X.; Lopez, R.; Luchtel, R.A.; Hafizi, S.; Gartrell, B.; Shenoy, N. Immune evasion in renal cell carcinoma: Biology, clinical translation, future directions. Kidney Int. 2020, 99, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Brugarolas, J. Molecular Genetics of Clear-Cell Renal Cell Carcinoma. J. Clin. Oncol. 2014, 32, 1968–1976. [Google Scholar] [CrossRef]
- Sanchez, D.J.; Simon, M.C. Genetic and metabolic hallmarks of clear cell renal cell carcinoma. Biochim. Biophys. Acta 2018, 1870, 23–31. [Google Scholar] [CrossRef]
- Huber, M.; Cato, A.C.; Ainooson, G.; Freichel, M.; Tsvilovskyy, V.; Jessberger, R.; Riedlinger, E.; Sommerhoff, C.P.; Bischoff, S.C. Regulation of the pleiotropic effects of tissue-resident mast cells. J. Allergy Clin. Immunol. 2019, 144, S31–S45. [Google Scholar] [CrossRef] [PubMed]
- Caughey, G.H. Mast cell tryptases and chymases in inflammation and host defense. Immunol. Rev. 2007, 217, 141–154. [Google Scholar] [CrossRef]
- Rubinchik, E.; Levi-Schaffer, F. Mast Cells and Fibroblasts: Two Interacting Cells. Int. J. Clin. Lab. Res. 1994, 24, 139–142. [Google Scholar] [CrossRef]
- Mukai, K.; Tsai, M.; Saito, H.; Galli, S.J. Mast cells as sources of cytokines, chemokines, and growth factors. Immunol. Rev. 2018, 282, 121–150. [Google Scholar] [CrossRef]
- Frungieri, M.B.; Weidinger, S.; Meineke, V.; Köhn, F.M.; Mayerhofer, A. Proliferative action of mast-cell tryptase is mediated by PAR2, COX2, prostaglandins, and PPAR: Possible relevance to human fibrotic disorders. Proc. Natl. Acad. Sci. USA 2002, 99, 15072–15077. [Google Scholar] [CrossRef] [PubMed]
- Wulff, B.C.; Wilgus, T.A. Mast Cell Activity in the Healing Wound: More than Meets the Eye? Exp. Dermatol. 2013, 22, 507–510. [Google Scholar] [CrossRef]
- Maurer, M.; Taube, C.; Schröder, N.W.; Ebmeyer, J.; Siebenhaar, F.; Geldmacher, A.; Schubert, N.; Roers, A. Mast cells drive IgE-mediated disease but might be bystanders in many other inflammatory and neoplastic conditions. J. Allergy Clin. Immunol. 2019, 144, S19–S30. [Google Scholar] [CrossRef] [PubMed]
- Hass, R. Role of MSC in the Tumor Microenvironment. Cancers 2020, 12, 2107. [Google Scholar] [CrossRef]
- Ch’Ng, S.; Wallis, R.A.; Yuan, L.; Davis, P.F.; Tan, S.T. Mast cells and cutaneous malignancies. Mod. Pathol. 2006, 19, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Melo, F.R.; Martin, S.S.; Sommerhoff, C.P.; Pejler, G. Exosome-mediated uptake of mast cell tryptase into the nucleus of melanoma cells: A novel axis for regulating tumor cell proliferation and gene expression. Cell Death Dis. 2019, 10, 1–16. [Google Scholar] [CrossRef]
- Glimelius, I.; Edström, A.; Fischer, M.; Nilsson, G.; Sundström, C.; Molin, D.; Amini, R.M.; Enblad, G. Angiogenesis and Mast Cells in Hodgkin Lymphoma. Leukemia 2005, 19, 2360–2362. [Google Scholar] [CrossRef][Green Version]
- Uguccioni, M.; D’Apuzzo, M.; Loetscher, M.; Dewald, B.; Baggiolini, M. Actions of the chemotactic cytokines MCP-1, MCP-2, MCP-3, RANTES, MIP-1α and MIP-1β on human monocytes. Eur. J. Immunol. 1995, 25, 64–68. [Google Scholar] [CrossRef]
- Conti, P.; Lauritano, D.; Caraffa, A.; Gallenga, C.E.; Kritas, S.K.; Ronconi, G.; Martinotti, S. Microglia and mast cells generate proinflammatory cytokines in the brain and worsen inflammatory state: Suppressor effect of IL-37. Eur. J. Pharmacol. 2020, 875, 173035. [Google Scholar] [CrossRef] [PubMed]
- Maciel, T.; Moura, I.; Hermine, O. The role of mast cells in cancers. F1000Prime Rep. 2015, 7, 09. [Google Scholar] [CrossRef]
- Dundar, E.; Oner, U.; Peker, B.C.; Metintas, M.; Isiksoy, S.; Ak, G. The Significance and Relationship between Mast Cells and Tumour Angiogenesis in Non-small Cell Lung Carcinoma. J. Int. Med Res. 2008, 36, 88–95. [Google Scholar] [CrossRef]
- Lichterman, J.; Reddy, S. Mast Cells: A New Frontier for Cancer Immunotherapy. Cells 2021, 10, 1270. [Google Scholar] [CrossRef]
- Desai, A.; Jung, M.-Y.; Olivera, A.; Gilfillan, A.M.; Prussin, C.; Kirshenbaum, A.S.; Beaven, M.A.; Metcalfe, D.D. IL-6 promotes an increase in human mast cell numbers and reactivity through suppression of suppressor of cytokine signaling 3. J. Allergy Clin. Immunol. 2016, 137, 1863–1871.e6. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, U.; Kjellén, L. Pathophysiology of heparan sulphate: Many diseases, few drugs. J. Intern. Med. 2013, 273, 555–571. [Google Scholar] [CrossRef]
- Davis, D.A.S.; Parish, C.R. Heparan Sulfate: A Ubiquitous Glycosaminoglycan with Multiple Roles in Immunity. Front. Immunol. 2013, 4, 470. [Google Scholar] [CrossRef]
- Morla, S. Glycosaminoglycans and Glycosaminoglycan Mimetics in Cancer and Inflammation. Int. J. Mol. Sci. 2019, 20, 1963. [Google Scholar] [CrossRef] [PubMed]
- Galli, S.J.; Grimbaldeston, A.M.; Tsai, M. Immunomodulatory mast cells: Negative, as well as positive, regulators of immunity. Nat. Rev. Immunol. 2008, 8, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Kissel, H.; Timokhina, I.; Hardy, M.P.; Rothschild, G.; Tajima, Y.; Soares, V.; Angeles, M.; Whitlow, S.R.; Manova, K.; Besmer, P. Point mutation in Kit receptor tyrosine kinase reveals essential roles for Kit signaling in spermatogenesis and oogenesis without affecting other Kit responses. EMBO J. 2000, 19, 1312–1326. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.-C.; Ingram, D.A.; Chen, S.; Zhu, Y.; Yuan, J.; Li, X.; Yang, X.; Knowles, S.; Horn, W.; Li, Y.; et al. Nf1-Dependent Tumors Require a Microenvironment Containing Nf1+/−- and c-kit-Dependent Bone Marrow. Cell 2008, 135, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Hölzel, M.; Landsberg, J.; Glodde, N.; Bald, T.; Rogava, M.; Riesenberg, S.; Becker, A.; Jönsson, G.; Tüting, T. A Preclinical Model of Malignant Peripheral Nerve Sheath Tumor-like Melanoma Is Characterized by Infiltrating Mast Cells. Cancer Res. 2015, 76, 251–263. [Google Scholar] [CrossRef]
- Siebenhaar, F.; Metz, M.; Maurer, M. Mast cells protect from skin tumor development and limit tumor growth during cutaneous de novo carcinogenesis in a Kit-dependent mouse model. Exp. Dermatol. 2014, 23, 159–164. [Google Scholar] [CrossRef]
- Ammendola, M.; Sacco, R.; Sammarco, G.; Donato, G.; Zuccalà, V.; Romano, R.; Luposella, M.; Patruno, R.; Vallicelli, C.; Verdecchia, G.M.; et al. Mast Cells Positive to Tryptase and c-Kit Receptor Expressing Cells Correlates with Angiogenesis in Gastric Cancer Patients Surgically Treated. Gastroenterol. Res. Pract. 2013, 2013, 1–5. [Google Scholar] [CrossRef]
- Babaei, M.A.; Kamalidehghan, B.; Saleem, M.; Huri, H.Z.; Ahmadipour, F. Receptor tyrosine kinase (c-Kit) inhibitors: A potential therapeutic target in cancer cells. Drug Des. Dev. Ther. 2016, 10, 2443–2459. [Google Scholar] [CrossRef]
- Thomsen, S.F. Epidemiology and natural history of atopic diseases. Eur. Clin. Respir. J. 2015, 2, 24642. [Google Scholar] [CrossRef] [PubMed]
- Abbas, M.; Moussa, M.; Akel, H. Type I Hypersensitivity Reaction. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Galli, S.J.; Tsai, M.; Piliponsky, A.M. The development of allergic inflammation. Nature 2008, 454, 445–454. [Google Scholar] [CrossRef]
- Gimenez-Rivera, V.-A.; Siebenhaar, F.; Zimmermann, C.; Siiskonen, H.; Metz, M.; Maurer, M. Mast Cells Limit the Exacerbation of Chronic Allergic Contact Dermatitis in Response to Repeated Allergen Exposure. J. Immunol. 2016, 197, 4240–4246. [Google Scholar] [CrossRef] [PubMed]
- Turner, H.; Kinet, J.-P. Signalling through the high-affinity IgE receptor FcεRI. Nature 1999, 402, 24–30. [Google Scholar] [CrossRef]
- Kashiwakura, J.-I.; Ando, T.; Matsumoto, K.; Kimura, M.; Kitaura, J.; Matho, M.H.; Zajonc, D.M.; Ozeki, T.; Ra, C.; MacDonald, S.M.; et al. Histamine-releasing factor has a proinflammatory role in mouse models of asthma and allergy. J. Clin. Investig. 2012, 122, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Leick, M.; Azcutia, V.; Newton, G.; Luscinskas, F.W. Leukocyte recruitment in inflammation: Basic concepts and new mechanistic insights based on new models and microscopic imaging technologies. Cell Tissue Res. 2014, 355, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Cruz, S.; Hernández-Pando, R.; Abraham, S.N.; Enciso, J.A. Mast Cell Activation byMycobacterium tuberculosis: Mediator Release and Role of CD48. J. Immunol. 2003, 170, 5590–5596. [Google Scholar] [CrossRef]
- Lyons, D.O.; Pullen, N.A. Beyond IgE: Alternative Mast Cell Activation Across Different Disease States. Int. J. Mol. Sci. 2020, 21, 1498. [Google Scholar] [CrossRef]
- Harvima, I.; Nilsson, G. Mast Cells as Regulators of Skin Inflammation and Immunity. Acta Derm. Venereol. 2011, 91, 644–650. [Google Scholar] [CrossRef] [PubMed]
- Amin, K. The role of mast cells in allergic inflammation. Respir. Med. 2012, 106, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Eissmann, M.F.; Buchert, M.; Ernst, M. IL33 and Mast Cells—The Key Regulators of Immune Responses in Gastrointestinal Cancers? Front. Immunol. 2020, 11, 1389. [Google Scholar] [CrossRef]
- Elst, J.; Sabato, V.; Faber, M.; Bridts, C.; Mertens, C.; Van Houdt, M.; Van Gasse, A.; Hagendorens, M.; Van Tendeloo, V.; Maurer, M.; et al. MRGPRX2 and Immediate Drug Hypersensitivity: Insights from Cultured Human Mast Cells. J. Investig. Allergol. Clin. Immunol. 2020, 31. [Google Scholar] [CrossRef]
- Metcalfe, D.D.; Peavy, R.D.; Gilfillan, A.M. Mechanisms of mast cell signaling in anaphylaxis. J. Allergy Clin. Immunol. 2009, 124, 639–646. [Google Scholar] [CrossRef]
- Zimmermann, C.; Troeltzsch, D.; Giménez-Rivera, V.A.; Galli, S.J.; Metz, M.; Maurer, M.; Siebenhaar, F. Mast cells are critical for controlling the bacterial burden and the healing of infected wounds. Proc. Natl. Acad. Sci. USA 2019, 116, 20500–20504. [Google Scholar] [CrossRef] [PubMed]
- Dahdah, A.; Gautier, G.; Attout, T.; Fiore, F.; Lebourdais, E.; Msallam, R.; Daëron, M.; Monteiro, R.; Benhamou, M.; Charles, N.; et al. Mast cells aggravate sepsis by inhibiting peritoneal macrophage phagocytosis. J. Clin. Investig. 2014, 124, 4577–4589. [Google Scholar] [CrossRef]
- Abe, T.; Sugaya, H.; Ishida, K.; Khan, I.W.; Tasdemir, I.; Yoshimura, K. Intestinal protection against Strongyloides ratti and mastocytosis induced by administration of interleukin-3 in mice. Immunology 1993, 80, 116–121. [Google Scholar] [PubMed]
- Dahlin, J.S.; Ding, Z.; Hallgren, J. Distinguishing Mast Cell Progenitors from Mature Mast Cells in Mice. Stem Cells Dev. 2015, 24, 1703–1711. [Google Scholar] [CrossRef] [PubMed]
- Reitz, M.; Brunn, M.-L.; Rodewald, H.-R.; Feyerabend, T.B.; Roers, A.; Dudeck, A.; Voehringer, D.; Jönsson, F.; Kühl, A.A.; Breloer, M. Mucosal mast cells are indispensable for the timely termination of Strongyloides ratti infection. Mucosal Immunol. 2016, 10, 481–492. [Google Scholar] [CrossRef]
- Renga, G.; Moretti, S.; Oikonomou, V.; Borghi, M.; Zelante, T.; Paolicelli, G.; Costantini, C.; De Zuani, M.; Villella, V.R.; Raia, V.; et al. IL-9 and Mast Cells Are Key Players of Candida albicans Commensalism and Pathogenesis in the Gut. Cell Rep. 2018, 23, 1767–1778. [Google Scholar] [CrossRef]
- Rönnberg, E.; Guss, B.; Pejler, G. Infection of Mast Cells with Live Streptococci Causes a Toll-Like Receptor 2- and Cell-Cell Contact-Dependent Cytokine and Chemokine Response. Infect. Immun. 2010, 78, 854–864. [Google Scholar] [CrossRef]
- Ebmeyer, J.; Furukawa, M.; Pak, K.; Ebmeyer, U.; Sudhoff, H.; Broide, D.; Ryan, A.F.; Wasserman, S. Role of mast cells in otitis media. J. Allergy Clin. Immunol. 2005, 116, 1129–1135. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhang, D.; Lyubynska, N.; Wolters, P.J.; Killeen, N.P.; Baluk, P.; McDonald, D.M.; Hawgood, S.; Caughey, G.H. Mast Cells Protect Mice from Mycoplasma Pneumonia. Am. J. Respir. Crit. Care Med. 2006, 173, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Lopes, J.P.; Stylianou, M.; Nilsson, G.; Urban, C.F. Opportunistic pathogen Candida albicans elicits a temporal response in primary human mast cells. Sci. Rep. 2015, 5, 12287. [Google Scholar] [CrossRef] [PubMed]
- Genovese, A.; Borgia, G.; Björck, L.; Petraroli, A.; De Paulis, A.; Piazza, M.; Marone, G. Immunoglobulin Superantigen Protein L Induces IL-4 and IL-13 Secretion from Human FcεRI+Cells Through Interaction with theκLight Chains of IgE. J. Immunol. 2003, 170, 1854–1861. [Google Scholar] [CrossRef] [PubMed]
- Paul, C.; Wolff, S.; Zapf, T.; Raifer, H.; Feyerabend, T.B.; Bollig, N.; Camara, B.; Trier, C.; Schleicher, U.; Rodewald, H.-R.; et al. Mast cells have no impact on cutaneous leishmaniasis severity and related Th2 differentiation in resistant and susceptible mice. Eur. J. Immunol. 2015, 46, 114–121. [Google Scholar] [CrossRef]
- Maurer, M.; Kostka, S.L.; Siebenhaar, F.; Moelle, K.; Metz, M.; Knop, J.; von Stebut, E. Skin mast cells control T cell-dependent host defense in Leishmania major infections. FASEB J. 2006, 20, 2460–2467. [Google Scholar] [CrossRef]
- Huang, B.; Huang, S.; Chen, X.; Liu, X.B.; Wu, Q.; Wang, Y.; Li, K.; Gao, H.; Cen, S.; Lin, R.; et al. Activation of Mast Cells Promote Plasmodium berghei ANKA Infection in Murine Model. Front. Cell. Infect. Microbiol. 2019, 9, 322. [Google Scholar] [CrossRef]
- Brown, M.G.; Hermann, L.L.; Issekutz, A.C.; Marshall, J.S.; Rowter, D.; Al-Afif, A.; Anderson, R. Dengue Virus Infection of Mast Cells Triggers Endothelial Cell Activation. J. Virol. 2010, 85, 1145–1150. [Google Scholar] [CrossRef]
- Sager, G.; Gabaglio, S.; Sztul, E.; Belov, G.A. Role of Host Cell Secretory Machinery in Zika Virus Life Cycle. Viruses 2018, 10, 559. [Google Scholar] [CrossRef]
- Marcet, C.W.; Laurent, C.D.S.; Moon, T.C.; Singh, N.; Befus, A.D. Limited replication of influenza A virus in human mast cells. Immunol. Res. 2012, 56, 32–43. [Google Scholar] [CrossRef]
- Sundstrom, J.B.; Little, D.M.; Villinger, F.; Ellis, J.E.; Ansari, A.A. Signaling through Toll-Like Receptors Triggers HIV-1 Replication in Latently Infected Mast Cells. J. Immunol. 2004, 172, 4391–4401. [Google Scholar] [CrossRef] [PubMed]
- Kritas, S.K.; Ronconi, G.; Caraffa, A.; Gallenga, C.E.; Ross, R.; Conti, P. Mast Cells Contribute to Coronavirus-Induced Inflammation: New Anti-Inflammatory Strategy. J. Biol. Regul. Homeost. Agents 2020, 34, 9–14. [Google Scholar] [CrossRef]
- Ribbing, C.; Engblom, C.; Lappalainen, J.; Lindstedt, K.; Kovanen, P.T.; Karlsson, M.A.; Lundeberg, L.; Johansson, C.; Nilsson, G.; Lunderius-Andersson, C.; et al. Mast cells generated from patients with atopic eczema have enhanced levels of granule mediators and an impaired Dectin-1 expression. Allergy 2010, 66, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Horii, Y.; Tiuria, R.; Sato, Y.; Nawa, Y. Mucosal mast cells and the expulsive mechanisms of mice against Strongyloides venezuelensis. Int. J. Parasitol. 1993, 23, 551–555. [Google Scholar] [CrossRef]
- Friend, D.S.; Ghildyal, N.; Austen, K.F.; Gurish, M.F.; Matsumoto, R.; Stevens, R.L. Mast cells that reside at different locations in the jejunum of mice infected with Trichinella spiralis exhibit sequential changes in their granule ultrastructure and chymase phenotype. J. Cell Biol. 1996, 135, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Hepworth, M.R.; Danilowicz-Luebert, E.; Rausch, S.; Metz, M.; Klotz, C.; Maurer, M.; Hartmann, S. Mast cells orchestrate type 2 immunity to helminths through regulation of tissue-derived cytokines. Proc. Natl. Acad. Sci. USA 2012, 109, 6644–6649. [Google Scholar] [CrossRef]
- Jiao, Q.; Luo, Y.; Scheffel, J.; Geng, P.; Wang, Y.; Frischbutter, S.; Li, R.; Maurer, M.; Zhao, Z. Skin Mast Cells Contribute to Sporothrix schenckii Infection. Front. Immunol. 2020, 11, 469. [Google Scholar] [CrossRef]
- Pinke, K.H.; de Lima, H.G.; Cunha, F.Q.; Lara, V.S. Mast cells phagocyte Candida albicans and produce nitric oxide by mechanisms involving TLR2 and Dectin-1. Immunobiology 2016, 221, 220–227. [Google Scholar] [CrossRef]
- Kraal, G.; van der Laan, L.; Elomaa, O.; Tryggvason, K. The macrophage receptor MARCO. Microbes Infect. 2000, 2, 313–316. [Google Scholar] [CrossRef]
- Urb, M.; Sheppard, D.C. The Role of Mast Cells in the Defence against Pathogens. PLoS Pathog. 2012, 8, e1002619. [Google Scholar] [CrossRef] [PubMed]
- Ermert, D.; Urban, C.F.; Laube, B.; Goosmann, C.; Zychlinsky, A.; Brinkmann, V. Mouse Neutrophil Extracellular Traps in Microbial Infections. J. Innate Immun. 2009, 1, 181–193. [Google Scholar] [CrossRef]
- Airola, K.; Petman, L.; Mäkinen-Kiljunen, S. Clustered sensitivity to fungi: Anaphylactic reactions caused by ingestive allergy to yeasts. Ann. Allergy Asthma Immunol. 2006, 97, 294–297. [Google Scholar] [CrossRef]
- Moretti, S.; Renga, G.; Oikonomou, V.; Galosi, C.; Pariano, M.; Iannitti, R.G.; Borghi, M.; Puccetti, M.; De Zuani, M.; Pucillo, C.E.; et al. A mast cell-ILC2-Th9 pathway promotes lung inflammation in cystic fibrosis. Nat. Commun. 2017, 8, 14017. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.S.; Portales-Cervantes, L.; Leong, E. Mast Cell Responses to Viruses and Pathogen Products. Int. J. Mol. Sci. 2019, 20, 4241. [Google Scholar] [CrossRef]
- Gotlib, J.; Kluin-Nelemans, J.C.; George, T.I.; Akin, C.; Sotlar, K.; Hermine, O.; Awan, F.T.; Hexner, E.; Mauro, M.J.; Sternberg, D.W.; et al. Efficacy and Safety of Midostaurin in Advanced Systemic Mastocytosis. N. Engl. J. Med. 2016, 374, 2530–2541. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Feunang, Y.D.; Guo, A.C.; Lo, E.J.; Marcu, A.; Grant, J.R.; Sajed, T.; Johnson, D.; Li, C.; Sayeeda, Z.; et al. DrugBank 5.0: A Major Update to the DrugBank Database for 2018. Nucleic Acids Res. 2018, 46, D1074–D1082. [Google Scholar] [CrossRef]
- Amer, S.M.; Kadi, A.A.; Darwish, H.W.; Attwa, M.W. Identification and characterization of in vitro phase I and reactive metabolites of masitinib using a LC-MS/MS method: Bioactivation pathway elucidation. RSC Adv. 2017, 7, 4479–4491. [Google Scholar] [CrossRef]
- Dhillon, S. Avapritinib: First Approval. Drugs 2020, 80, 433–439. [Google Scholar] [CrossRef]
- Alzofon, N.; Jimeno, A. Avapritinib for metastatic or unresectable gastrointestinal stromal tumors. Drugs Today 2020, 56, 561–571. [Google Scholar] [CrossRef]
- Huang, S.; Okumura, K.; Sinicrope, F.A. BH3 Mimetic Obatoclax Enhances TRAIL-Mediated Apoptosis in Human Pancreatic Cancer Cells. Clin. Cancer Res. 2008, 15, 150–159. [Google Scholar] [CrossRef]
- Peter, B.; Cerny-Reiterer, S.; Hadzijusufovic, E.; Schuch, K.; Stefanzl, G.; Eisenwort, G.; Gleixner, K.V.; Hoermann, G.; Mayerhofer, M.; Kundi, M.; et al. The pan-Bcl-2 blocker obatoclax promotes the expression of Puma, Noxa, and Bim mRNA and induces apoptosis in neoplastic mast cells. J. Leukoc. Biol. 2013, 95, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Mannargudi, M.B.; Deb, S. Clinical Pharmacology and Clinical Trials of Ribonucleotide Reductase Inhibitors: Is It a Viable Cancer Therapy? J. Cancer Res. Clin. Oncol. 2017, 143, 1499–1529. [Google Scholar] [CrossRef]
- Schmidt, J.; Welsch, T.; De Jager, D.J.; Muhlradt, P.F.; Büchler, M.W.; Marten, A. Intratumoural injection of the toll-like receptor-2/6 agonist ‘macrophage-activating lipopeptide-2′ in patients with pancreatic carcinoma: A phase I/II trial. Br. J. Cancer 2007, 97, 598–604. [Google Scholar] [CrossRef] [PubMed]
- Smolich, B.D.; Yuen, H.A.; West, K.A.; Giles, F.J.; Albitar, M.; Cherrington, J.M. The antiangiogenic protein kinase inhibitors SU5416 and SU6668 inhibit the SCF receptor (c-kit) in a human myeloid leukemia cell line and in acute myeloid leukemia blasts. Blood 2001, 97, 1413–1421. [Google Scholar] [CrossRef]
- Qian, B.; Yao, Y.; Liu, C.; Zhang, J.; Chen, H.; Li, H. SU6668 modulates prostate cancer progression by downregulating MTDH/AKT signaling pathway. Int. J. Oncol. 2017, 50, 1601–1611. [Google Scholar] [CrossRef]
- Murphy, S.; Kelly, H.W. Cromolyn Sodium: A Review of Mechanisms and Clinical Use in Asthma. Drug Intell. Clin. Pharm. 1987, 21, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, E.; Manley, P.; Mestan, J.; Cowan-Jacob, S.; Ray, A.; Griffin, J.D. AMN107 (nilotinib): A novel and selective inhibitor of BCR-ABL. Br. J. Cancer 2006, 94, 1765–1769. [Google Scholar] [CrossRef]
- Yuksel, M.; Okajima, K.; Uchiba, M.; Okabe, H. Gabexate Mesilate, a Synthetic Protease Inhibitor, Inhibits Lipopolysaccharide-Induced Tumor Necrosis Factor-α Production by Inhibiting Activation of Both Nuclear Factor-κB and Activator Protein-1 in Human Monocytes. J. Pharmacol. Exp. Ther. 2003, 305, 298–305. [Google Scholar] [CrossRef]
- Nakatsuka, M.; Asagiri, K.; Noguchi, S.; Habara, T.; Kudo, T. Nafamostat mesilate, a serine protease inhibitor, suppresses lipopolysaccharide-induced nitric oxide synthesis and apoptosis in cultured human trophoblasts. Life Sci. 2000, 67, 1243–1250. [Google Scholar] [CrossRef]
- Papich, M.G. Masitinib Mesylate. In Saunders Handbook of Veterinary Drugs; Elsevier: Amsterdam, The Netherlands, 2016; pp. 476–477. [Google Scholar]
- Garland, W.; Benezra, R.; Chaudhary, J. Targeting Protein–Protein Interactions to Treat Cancer—Recent Progress and Future Directions. Annu. Rep. Med. Chem. 2013, 48, 227–245. [Google Scholar] [CrossRef]
- Ciccolini, J.; Serdjebi, C.; Peters, G.J.; Giovannetti, E. Pharmacokinetics and pharmacogenetics of Gemcitabine as a mainstay in adult and pediatric oncology: An EORTC-PAMM perspective. Cancer Chemother. Pharmacol. 2016, 78, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Koop, E.A.; Voest, E.E. Tumor Vasculature As A Target. In Anticancer Drug Development; Elsevier: Amsterdam, The Netherlands, 2002; pp. 123–136. [Google Scholar]
- Conti, P.; Pregliasco, F.; Bellomo, R.; Gallenga, C.; Caraffa, A.; Kritas, S.; Lauritano, D.; Ronconi, G. Mast Cell Cytokines IL-1, IL-33, and IL-36 Mediate Skin Inflammation in Psoriasis: A Novel Therapeutic Approach with the Anti-Inflammatory Cytokines IL-37, IL-38, and IL-1Ra. Int. J. Mol. Sci. 2021, 22, 8076. [Google Scholar] [CrossRef]
- Conti, P.; Stellin, L.; Caraffa, A.; Gallenga, C.E.; Ross, R.; Kritas, S.K.; Frydas, I.; Younes, A.; Di Emidio, P.; Ronconi, G. Advances in Mast Cell Activation by IL-1 and IL-33 in Sjögren’s Syndrome: Promising Inhibitory Effect of IL-37. Int. J. Mol. Sci. 2020, 21, 4297. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Ding, F.; Zhai, Y.; Tao, W.; Bi, J.; Fan, H.; Yin, N.; Wang, Z. IL-37 is protective in allergic contact dermatitis through mast cell inhibition. Int. Immunopharmacol. 2020, 83, 106476. [Google Scholar] [CrossRef]
- Zhang, M.; Liang, W.; Gong, W.; Yoshimura, T.; Chen, K.; Wang, J.M. The Critical Role of the Antimicrobial Peptide LL-37/ CRAMP in Protection of Colon Microbiota Balance, Mucosal Homeostasis, Anti-Inflammatory Responses, and Resistance to Carcinogenesis. Crit. Rev. Immunol. 2019, 39, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, L.; Shi, R.; Liu, X.; Zhang, J.; Zou, Z.; Hao, Z.; Tao, A. Mast Cell Targeted Chimeric Toxin Can Be Developed as an Adjunctive Therapy in Colon Cancer Treatment. Toxins 2016, 8, 71. [Google Scholar] [CrossRef]



{kind=link}
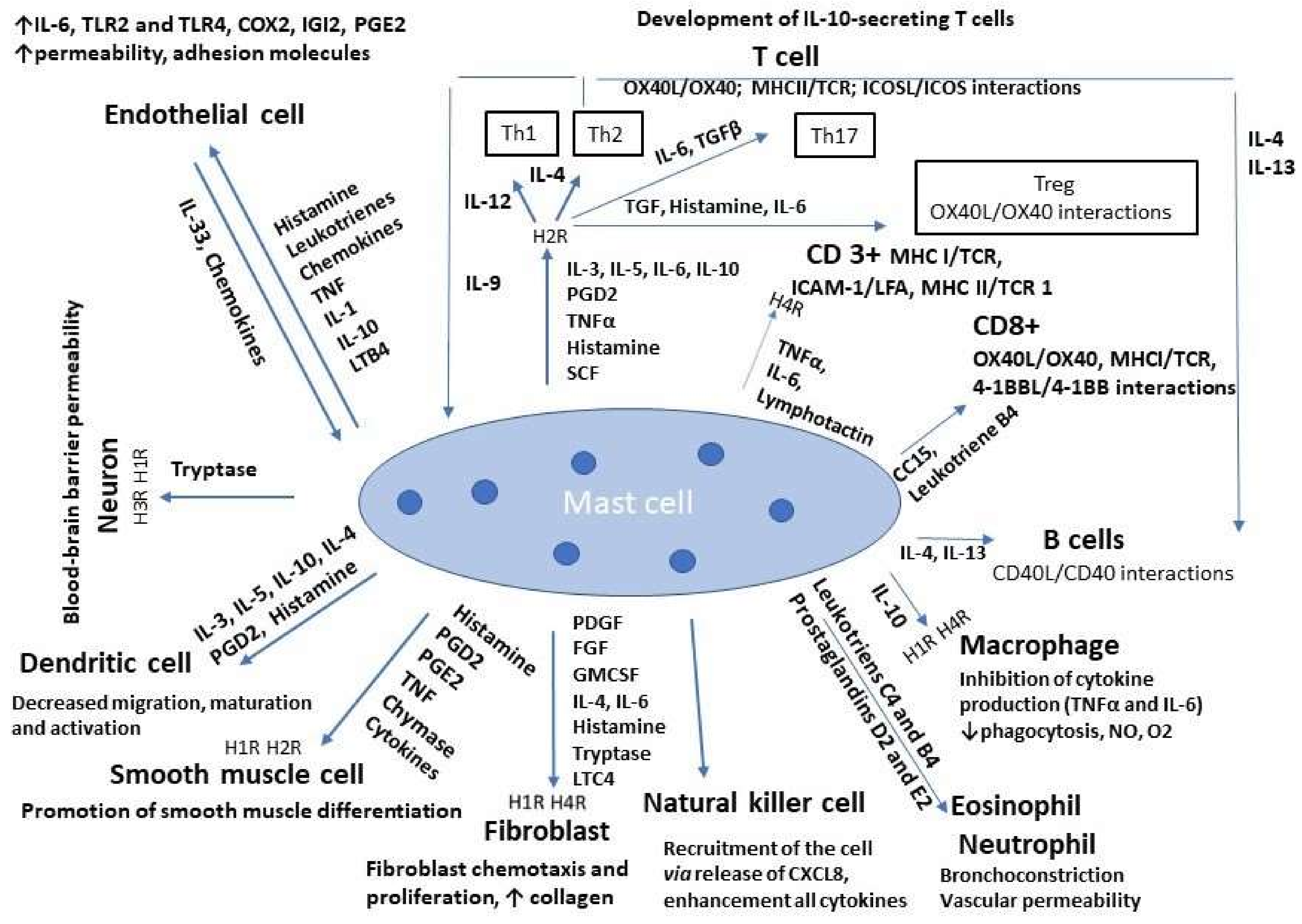{kind=link}
| Cell Origin | MC-Released Mediators | References | |
|---|---|---|---|
| Immune cells | Eosinophils | Histamine, IL-5, IL-9, SCF, LTD4, PAF, PGD2, VEGF-A | [9,53,55] |
| Basophils | Histamine, PAF, PGD2 | [9,35,56] | |
| Neutrophils | Histamine, LTB4, PAF, heparin | [7,8,9,35,49,53,54,57,58] | |
| Platelets | PAF | [58] | |
| Monocytes | Histamine, LTD4, VEGF-A, PAF | [9,53,55,59] | |
| Macrophages | Histamine, IL-13, IL-6, PAF, PGD2 | [5,60,61,62,63] | |
| Dendritic cells | Histamine, PGE2, PGD2, VEGF-C, IL-13 | [9,11,35,54,62,64,65,66] | |
| Innate lymphoid cells | IL-1β, IL-9, PGD2, LTD4 | [59,67,68] | |
| NK cells | Histamine, heparin | [35,49,64] | |
| CD4+/CD8+ T cells | Histamine, LTC4, LTD4, TNF-α | [40,55,60] | |
| Th1 cells | Histamine | [11,60] | |
| Th2 cells | Histamine, LTC4, LTD4, PGD2 | [9,11,35,54] | |
| Tfh cells | IL-6 | [54] | |
| B cells | Histamine, PAF, IL-5 | [35,58,63] | |
| Treg cells | Histamine | [40] | |
| Non-immune cells | Blood endothelial cells | Histamine, LTC4, LTD4, PGD2, PAF, VEGF-A, IL-13, IL-1β | [55,69,70,71,72] |
| Lymphatic endothelial cells | VEGF-C, VEGF-D | [73,74] | |
| Bronchial epithelia | IL-13, TNF-α, IL-9, TGF-β, PGD2 | [31,55,56,75] | |
| Smooth-muscle cells | Histamine, LTC4, LTD4, PGD2, tryptase | [9,76,77,78,79] | |
| Goblet cells | IL-13, LTE4 | [80] | |
| Cardiomyocytes | Tryptase | [31,54,72,81,82] | |
| Osteo-blasts/clasts | Histamine, osteopontin | [35,54,68,83,84,85,86] | |
| Neurons | Histamine, NGF, SP, PGD2 | [9,53,85,86] | |
| Adipocytes | IL-13 | [31,54,68,87] | |
| Fibroblasts | Tryptase, PGD2, TNF-α, TGF-β, IL-13, IL-1β | [9,53,55,60,88] | |
| Keratinocytes | Tryptase, PAF | [11,53] |
| Pro-Tumorigenic Role of Mast Cells in Human Cancers | Pro-Tumorigenic Mechanisms | References |
|---|---|---|
| Bladder | Angiogenesis (VEGF-A, VEGF-B, FGF-2, IL-8), Genetic instability, DNA & RNA damage (ROS), Tumor cell growth (PAF), Immunosuppression (adenosine), Mesenchymal transition (beta TGF, IL-8), Tissue remodelling (Tryptase), Activation of STAT-3 (IL-6) | [29,50,54,55,56,62,63,69,93,94,95,96] |
| Colorectal | Angiogenesis (VEGF-A, VEGF-B, FGF-2, IL-8), Genetic instability, DNA & RNA damage (ROS), Tumor cell growth (PAF) | [64,95,97,98] |
| Esophagus | Angiogenesis (VEGF-A, VEGF-B, FGF-2, IL-8), Genetic instability, DNA & RNA damage (ROS), Tumor cell growth (PAF) | [55,56,62,63,78,99,100] |
| Gastric | Angiogenesis (VEGF-A, VEGF-B, FGF-2, IL-8), Genetic instability, DNA & RNA damage (ROS) | [9,49,100] |
| Hepatocellular | Angiogenesis (VEGF-A, VEGF-B, FGF-2, IL-8), Genetic instability, DNA & RNA damage (ROS), Tumor cell growth (PAF), Activation of STAT-3 (IL-6) | [55,101] |
| Hodgkin’s lymphoma | Angiogenesis (VEGF-A, VEGF-B, FGF-2, IL-8), Immunosuppression (adenosine), Mesenchymal transition (beta TGF, IL-8), IL-13 (Macrophage 2 polarization) | [99,102] |
| Melanoma | Angiogenesis (VEGF-A, VEGF-B, FGF-2, IL-8), Activation of STAT-3 (IL-6) | [9,55,60,62,63,76,103] |
| Pancreas | Angiogenesis (VEGF-A, VEGF-B, FGF-2, IL-8), Genetic instability, DNA & RNA damage (ROS), Tumor cell growth (PAF) | [55,62,70,74,98,104,105] |
| Prostate | Angiogenesis (VEGF-A, VEGF-B, FGF-2, IL-8), Genetic instability, DNA & RNA damage (ROS) | [94,100,106] |
| Thyroid | Tissue remodelling (Tryptase), Mesenchymal transition (beta TGF, IL-8), Genetic instability, DNA & RNA damage (ROS) | [55,96,107] |
| Anti-tumorigenic Role of Mast Cells in Human Tumors | Antitumorigenic Role | |
| Breast | Cytotoxicity (TNF alpha), Cytotoxicity (ROS), Tumor growth inhibition (heparin), Tumor cell inhibition (cytolytic activity), Tumor cell apoptosis (IL-4, TNF) | [58,59,62,100,102] |
| Colorectal | Cytotoxicity (TNF alpha), Cytotoxicity (ROS), Tumor growth inhibition (heparin), Tumor cell growth inhibition (IL-9) | [9,54,62] |
| Lung | Cytotoxicity (TNF alpha), Cytotoxicity (ROS), Tumor growth inhibition (heparin), Tumor cell apoptosis (IL-4, TNF) | [9,53,55,59,108] |
| Mesothelioma | Cytotoxicity (TNF alpha), Cytotoxicity (ROS), Tumor growth inhibition (heparin) | [55,109] |
| Melanoma | Cytotoxicity (TNF alpha), Cytotoxicity (ROS), Tumor growth inhibition (heparin), Tumor cell growth inhibition (IL-9), APC maturation (histamine), Tumor cell inhibition (cytolytic activity) | [9,55,60,62,103] |
| Ovarian | Cytotoxicity (TNF alpha), Cytotoxicity (ROS), Tumor growth inhibition (heparin), | [76,94,110,111] |
| Pancreas | Cytotoxicity (TNF alpha), Cytotoxicity (ROS), Tumor growth inhibition (heparin), | [55,62,70,98,105] |
| Prostate | Tumor cell apoptosis (IL-4, TNF), Immune cell recruitment and activation, Tumor cell growth inhibition (IL-9) | [94,100,106] |
| Tumors in Which Mast Cells Play Both a Pro- and Anti-Tumorigenic Role | Pro-and Anti-Tumorigenic Mechanisms | |
| Colorectal | Cytotoxicity (ROS), DNA & RNA damage (ROS) | [9,53,56,112] |
| Lung | Cytotoxicity (ROS), DNA & RNA damage (ROS), Genetic instability | |
| Prostate | DNA & RNA damage (ROS), Genetic instability | |
| Melanoma | DNA & RNA damage (ROS), Genetic instability | |
| Pancreatic | Cytotoxicity (ROS), DNA & RNA damage (ROS), Genetic instability | |
| Non-Contributing Role of Mast Cells in Tumors | ||
| Colorectal | Mutations in beta-catenin or axin Inactivation of TGF-beta signaling pathway | [9,55,62,78] |
| Non-small cell lung cancer | Mutation in EGFR Mutation in anaplastic lymphoma kinase | [55,62,100,102] |
| Renal | Biallelic inactivation of VHL, Deletion of SETD2, BAP1, PBRM1, Constitutive activation of the HIF pathway (angiogenesis, anaerobic glycolysis, pentose phosphate pathway, epithelial–mesenchymal transition, invasion, and metastasis) | [113,114,115,116] |
| Mediators | MC-Interacting Cell Types | Biological Effects | References |
|---|---|---|---|
| Proteases Tryptase, MMPs Proteoglycans heparin Chemokines (MCP-1, RANTES, TARC Cytokines IL-1, IL-3, IL-4, IL-6, IL-18, IL-33, TNF-α, SCF, TGF-β Growth factors VEGF, bFGF, NGF, GM-CSF, M-CSF Lipid-derived LTC4, LTD4, LTE4, PGD2, PAF Neuropeptides CRH, VIP | Endothelial cells, Bronchial, Smooth muscle, Neurons, Vascular, Gastric parietal cells, Cells of the central nervous system, Basophils and eosinophils | Type-1 hypersensitivity, Pro-inflammatory effects, Wheal and flare reaction in skin, Bronchoconstriction, Drop in blood pressure, Gastrointestinal hyperactivity | [11,35,79,84,85,92,151] |
| Growth factors bFGF, VEGF, NGF Cytokines IL-1, IL-3, IL-4, IL-6, IL-18, IL-33, TNF-α, SCF, TGF-β | Vascular, Gastric parietal cells | Regulation of the immune response, Th1 lymphocyte cytokine production | [8,9,11,35,92,152] |
| Proteases tryptase, MMPs Neuropeptides CRH, VIP Lipid-derived LTC4, LTD4, LTE4, PGD2, PAF | Cells of the central nervous system, Neurons | Blood–brain barrier permeability | [11,40,41,82,125,129,153,154] |
| Chemokines MCP-1, RANTES, TARC Cytokines IL-1, IL-3, IL-4, IL-6, IL-18, IL-33, TNF-α, SCF, TGF-β | Basophils and eosinophils | Pro-inflammatory effects, Process of chemotaxis, Stimulation of histamine and cytokine generation |
| Biological Agents | Localization | MC Subpopulation | Signaling/Recruitment | References |
|---|---|---|---|---|
| Pathogenesis | ||||
| Bacteroidetes Proteobacteria | Gut | MMCs; MCPT1 | IL-9, ILC-2; Th9 | [163] |
| Streptococcus equis | Peritoneum | BMMC-derived MCs | IL-4, IL-6, IL-12, IL-13, TNF-α, chemokines (CCL2/MCP-1, CCL7/MCP-3, CXCL2/MIP-2, CCL5/RANTES) | [164] |
| Haemophilus influenzae | Middle ear | BMMC-derived MCs | - | [165] |
| Mycoplasma pulmonis | Lung | BMMC-derived MCs | TNF-α, MCP-1, MIP-2, IL-6, Histamine | [166] |
| Pseudomonas aeruginosa | Skin | BMMC-derived MCs | NF-α, MCP-1, MIP-2, and IL-6 | [59] |
| C. albicans (yeast and hyphae) | Mucous membranes | BMMC-derived MCs, MMCs; MCPT1 | IL-9, ILC-2; Th9, IL-4, IL-6, IL-8; IL-13, TNF-α; Neutrophils, Macrophage crawling and migration | [16,163,167] |
| Malassezia sympodialis | Skin | BMMC-derived MCs | IL-6; IL-8 β-hexosaminidase, IgE | [13] |
| Staphylococcus aureus Peptostreptococcus magnus | Skin | BMMC-derived MCs | IL-4; IL-13; IgE | [168] |
| Strongyloides ratti and S. venezuelensisis | Mucosal intestine | MMCs | IL-4, IgE | [162] |
| Trichinella spiralis | Mucosal intestine; vessels | BM-resident HPC | mMCP-1, IgE, IL-2, | [36] |
| Leishmania major L. infantum | Cutaneous tissues | BMMCs | Β-hexosaminidase, TNF-α, Chymases (mMCP-1, mMCP-9), NO, IgE; IL-12; IFN-γ | [169,170] |
| Plasmodium berghei | CLN; skin | MMCs | TNF-α; CCL2; CXCL1; MMP-9; IFN-γ | [171] |
| DENV | Mucous membranes | MMCs | Tryptase, IL-1β | [172] |
| Zika virus | Mucous membranes | MMCs | Histame, IL-9, Th2 | [173,174] |
| Influenza and parainfluenza | Mucous membranes | MMCs | Th2, type I INF | [174] |
| HIV-1gp120 | Vascularized tissues | MC progenitors in the blood | IgE | [175] |
| Hepatitis C | Vascularized tissues | MC progenitors in the blood | IL-10, type I TNF | [101] |
| Coronavirus | Mucous membranes of lungs | MMCs | Histamine, Protease, PGD2, LTC4, IL-1, IL-6, IL-33 | [176] |
| Homeostatic Conditions | ||||
| C. albicans | Mucosal gut | CTMC | TGF-β; ILC2; IL-9; Th9; Treg Foxp3; IL-10 | [163] |
| C. albicans | Mucous membrane | BMMC-derived MCs | Restrain myeloid cells | [16] |
| Malassezia sympodialis | Skin | BMMC-derived MCs | IL-8, IgE | [177] |
| Drug | Target | Mode of Action | References |
|---|---|---|---|
| Midostaurin Chemical name: N-[(2S,3R,4R,6R)-3-methoxy-2-methyl-16-oxo-29-oxa-1,7,17-triazaoctacyclo [1 2.12.2.12,6.07,28.08,13.015,19.020,27.021,26]nonacosa-8,10,12,14,19,21,23,25,27-nonaen-4-yl]-N-methylbenzamide Brand name: Rydapt | PKC alpha, VEGFR2, KIT | Antagonist and inhibitor | PubChem CID: 9829523 [103] |
| WT and/or mutant FLT3 tyrosine kinases | Apoptosis of target leukemia cells expressing target receptors and mast cells, Antiproliferative activity, Interacts with OATP | ||
| Imatinib Chemical name: 4-[(4-methylpiperazin-1-yl) methyl]-N-[4-methyl-3-[(4-pyridin-3-ylpyrimidin-2-yl)amino]phenyl]benzamide Brand name: Gleevec (USA) or Glivec (Europe/Australia) | BCRP | Inhibitor | PubChem CID: 5291 [64] |
| MSCGFR KIT | Antagonist, multitarget | ||
| RET proto-oncogene | Inhibitor | ||
| HANGFR | Antagonist | ||
| PDGFRα | Antagonist | ||
| EDDCR 1 | Antagonist | ||
| KIT ABL1 | Inhibitor | ||
| Platelet-derived growth factor receptor beta | Antagonist | ||
| Dasatinib Chemical name: N-(2-chloro-6-methylphenyl)-2-[[6-[4-(2-hydroxyethyl)piperazin-1-yl]-2-methylpyrimidin-4-yl]amino]-1,3-thiazole-5-carboxamide | KIT ABL1 | Multitarget-bind to both the active and inactive conformation of the ABL kinase domain | PubChem CID: 3062316 [59,83] |
| Proto-oncogene KIT Src | Multitarget | ||
| Ephrin type-A receptor 2 | Antagonist | ||
| KIT Lck | Multitarget | ||
| KIT | Inhibitor | ||
| MSCGFR Kit | Antagonist | ||
| PDGFRP | Antagonist | ||
| STAT5B | Inhibitor | ||
| Abelson KIT2 | Multitarget | ||
| KIT Fyn | Multitarget | ||
| Nilotinib Chemical name: 4-methyl-N-[3-(4-methylimidazol-1-yl)-5-(trifluoromethyl)phenyl]-3-[(4-pyridin-3-ylpyrimidin-2-yl)amino]benzamide | KIT ABL1 | Inhibitor of BCR–ABL-binds to the inactive conformation of ABL | PubChem CID: 644241 [201] |
| MSCGFR Kit | Antagonist | ||
| Tranilast Chemical name: 2-[[(E)-3-(3,4-dimethoxyphenyl)prop-2-enoyl]amino]benzoic acid Brand name: Rizaben | Hematopoietic prostaglandin D synthase | Inhibitor | PubChem CID: 5282230 [103,189] |
| MAP kinases (extracellularly regulated kinase 1 and 2 and JNK) | Calcium channel blocker, Antineoplastic agent, Aryl hydrocarbon receptor agonist and a hepatoprotective agent, Inhibit TGF-β production, interferon-gamma, IL-6, IL-10, and IL-17 by lymphoid cells | ||
| Gabexate Chemical name: 4-[[6-[(Aminoiminomethyl)amino]-1-oxohexyl]oxy]-benzoic acid ethyl ester mesylate Brand name: Gabexate mesylate (Japan), Foy (Taiwan), Reminaron (Japan) | Serine protease Thrombin Trypsin Plasminogen Plasma kallikrein | Inhibitor, Antithrombotic in vitro and in vivo, inhibits LPS-induced TNF-α production, Inhibiting NF-κB and AP-1 activation | PubChem CID: 3447 [202] |
| Nafamostat mesylate Chemical name: (6-carbamimidoylnaphthalen-2-yl) 4-(diaminomethylideneamino)benzoate Brand name: Fusan | Serine protease | Inhibits enzyme: thrombin, Xa, and XIIa), The kallikrein–kinin system, The complement system, Pancreatic proteases and activation of protease-activated receptors (PARs), Lipopolysaccharide-induced nitric oxide production, apoptosis, Interleukin (IL)-6 and IL-8. Antioxidant in TNF-α-induced ROS production | PubChem CID: 5311180 [203] |
| Prothrombin Coagulation factor X Coagulation factor XII Trypsin-1 Kallikrein-1 Intracellular adhesion molecule 1 | Inhibitor | ||
| Masitinib mesylate Chemical name: N-(4-methyl-3-{[4-(pyridin-3-yl)-1,3-thiazol-2-yl]amino}phenyl)-4-[(4-methylpiperazin-1-yl)methyl]benzamide Brand name: Masivet, Kinavet | Tyrosine-kinase PDGF and KIT | Inhibiting the stem cell factor that regulates mast cell tumor proliferation, Antiproliferative actions- targets the c-KIT pathway | PubChem CID: 25024769 [103] |
| Avapritinib Chemical name: (1S)-1-(4-fluorophenyl)-1-(2-{4-[6-(1-methyl-1H-pyrazol-4-yl)pyrrolo[2,1-f][1,2,4]triazin-4-yl]piperazin-1-yl}pyrimidin-5-yl)ethan-1-amine Brand name: Ayvakit | MSCGFR KIT and PDGFR alpha |
Inhibitor Negatively modulates the action of cell transporters | PubChem CID: 118023034 [204] |
| Obatoclax Chemical name: 2-[(2Z)-2-[(3,5-dimethyl-1H-pyrrol-2-yl)methylidene]-3-methoxy-2H-pyrrol-5-yl]-1H-indole | Apoptosis regulator Bcl-2 | Leads to release of apoptosis-inducing cytochromec | PubChem CID: 11404337 [205] |
| Bcl-2 family of proteins | Displaces BH3 domains by activation of a pocket of the BcL-2 family member | ||
| Gemcitabine Chemical name: 4-amino-1-[(2R,4R,5R)-3,3-difluoro-4-hydroxy-5-(hydroxymethyl)oxolan-2-yl]-1,2-dihydropyrimidin-2-one Brand name: Gemzar, Infugem | Block DNA Replication DNA during the “S” phase (or DNA synthesis phase of the cell cycle), stopping normal development and division | Cross-linking/Alkylation | PubChem CID: 60750 [206] |
| Ribonucleoside-diphosphate reductase large subunit | Blocks an enzyme which converts the cytosine nucleotide into the deoxy derivative | ||
| Thymidylate synthase | Blocks the incorporation of the thymidine nucleotide | ||
| UMP-CMP kinase | Inhibitor | ||
| Orantinib Chemical name: 3-(2,4-dimethyl-5-{[(3Z)-2-oxo-2,3-dihydro-1H-indol-3-ylidene]methyl}-1H-pyrrol-3-yl)propanoic acid | KIT autophosphorylation VEGFR2 PDGFR FGFR angiogenesis proliferation | Inhibitor | PubChem CID: 5329099 [207] |
| Cromolyn Chemical name: 5-{3-[(2-carboxy-4-oxo-4H-chromen-5-yl)oxy]-2-hydroxypropoxy}-4-oxo-4H-chromene-2-carboxylic acid Brand name: Spinhaler, nebulizer solution, metered-dose inhaler, NasalCrom, Nasal Allergy Control, Nasalcrom Child | Protein S100-P | Antagonist | PubChem CID: 2882 [99,200] |
| Calcium sensor Capsaicin receptor | Ion channel blockade Blockade | ||
| Cellular calcium signaling | Blockade of signaling of heat shock protein or G-protein | ||
| Stabilizing mast cells | Prevents the subsequent release of mediators; Precise mechanism/clinical activity not delineated |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sobiepanek, A.; Kuryk, Ł.; Garofalo, M.; Kumar, S.; Baran, J.; Musolf, P.; Siebenhaar, F.; Fluhr, J.W.; Kobiela, T.; Plasenzotti, R.; et al. The Multifaceted Roles of Mast Cells in Immune Homeostasis, Infections and Cancers. Int. J. Mol. Sci. 2022, 23, 2249. https://doi.org/10.3390/ijms23042249
Sobiepanek A, Kuryk Ł, Garofalo M, Kumar S, Baran J, Musolf P, Siebenhaar F, Fluhr JW, Kobiela T, Plasenzotti R, et al. The Multifaceted Roles of Mast Cells in Immune Homeostasis, Infections and Cancers. International Journal of Molecular Sciences. 2022; 23(4):2249. https://doi.org/10.3390/ijms23042249
Chicago/Turabian StyleSobiepanek, Anna, Łukasz Kuryk, Mariangela Garofalo, Sandeep Kumar, Joanna Baran, Paulina Musolf, Frank Siebenhaar, Joachim Wilhelm Fluhr, Tomasz Kobiela, Roberto Plasenzotti, and et al. 2022. "The Multifaceted Roles of Mast Cells in Immune Homeostasis, Infections and Cancers" International Journal of Molecular Sciences 23, no. 4: 2249. https://doi.org/10.3390/ijms23042249
APA StyleSobiepanek, A., Kuryk, Ł., Garofalo, M., Kumar, S., Baran, J., Musolf, P., Siebenhaar, F., Fluhr, J. W., Kobiela, T., Plasenzotti, R., Kuchler, K., & Staniszewska, M. (2022). The Multifaceted Roles of Mast Cells in Immune Homeostasis, Infections and Cancers. International Journal of Molecular Sciences, 23(4), 2249. https://doi.org/10.3390/ijms23042249