
Lignin Synthesis, Affected by Sucrose in Lotus (Nelumbo nucifera) Seedlings, Was Involved in Regulation of Root Formation in the Arabidopsis thanliana

Abstract
:1. Introduction
2. Results
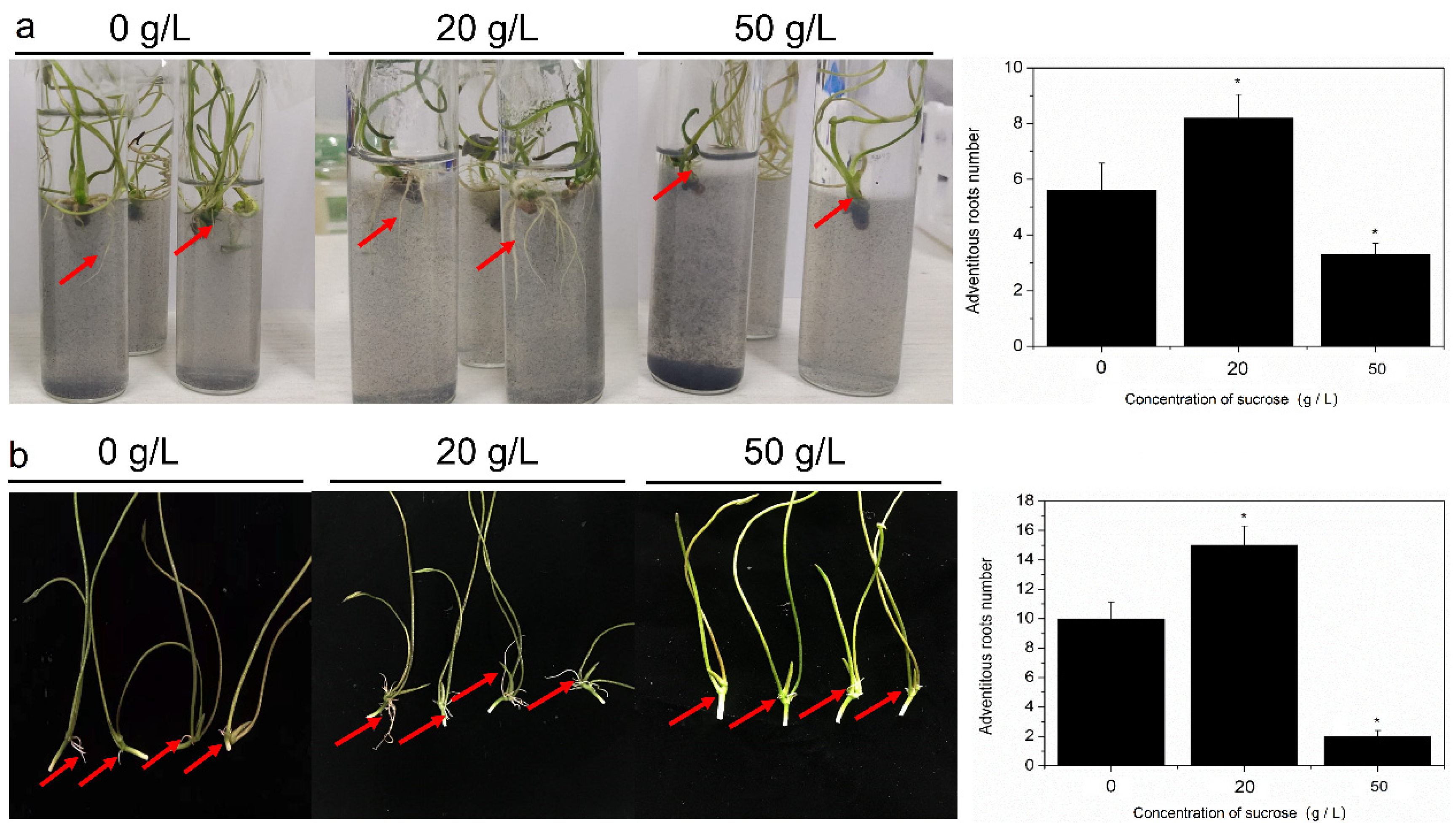
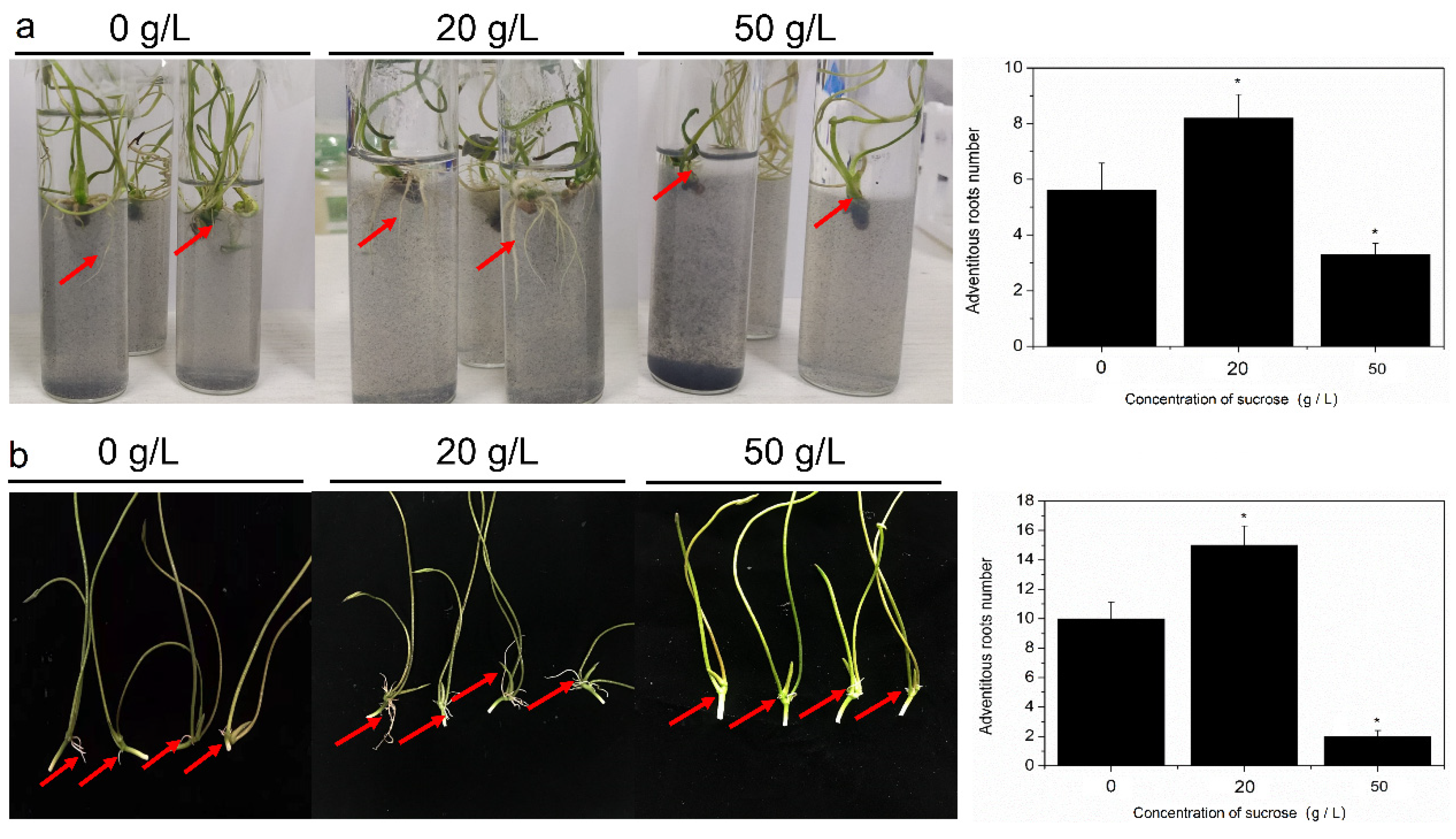
2.1. Effect of Sucrose on AR Formation
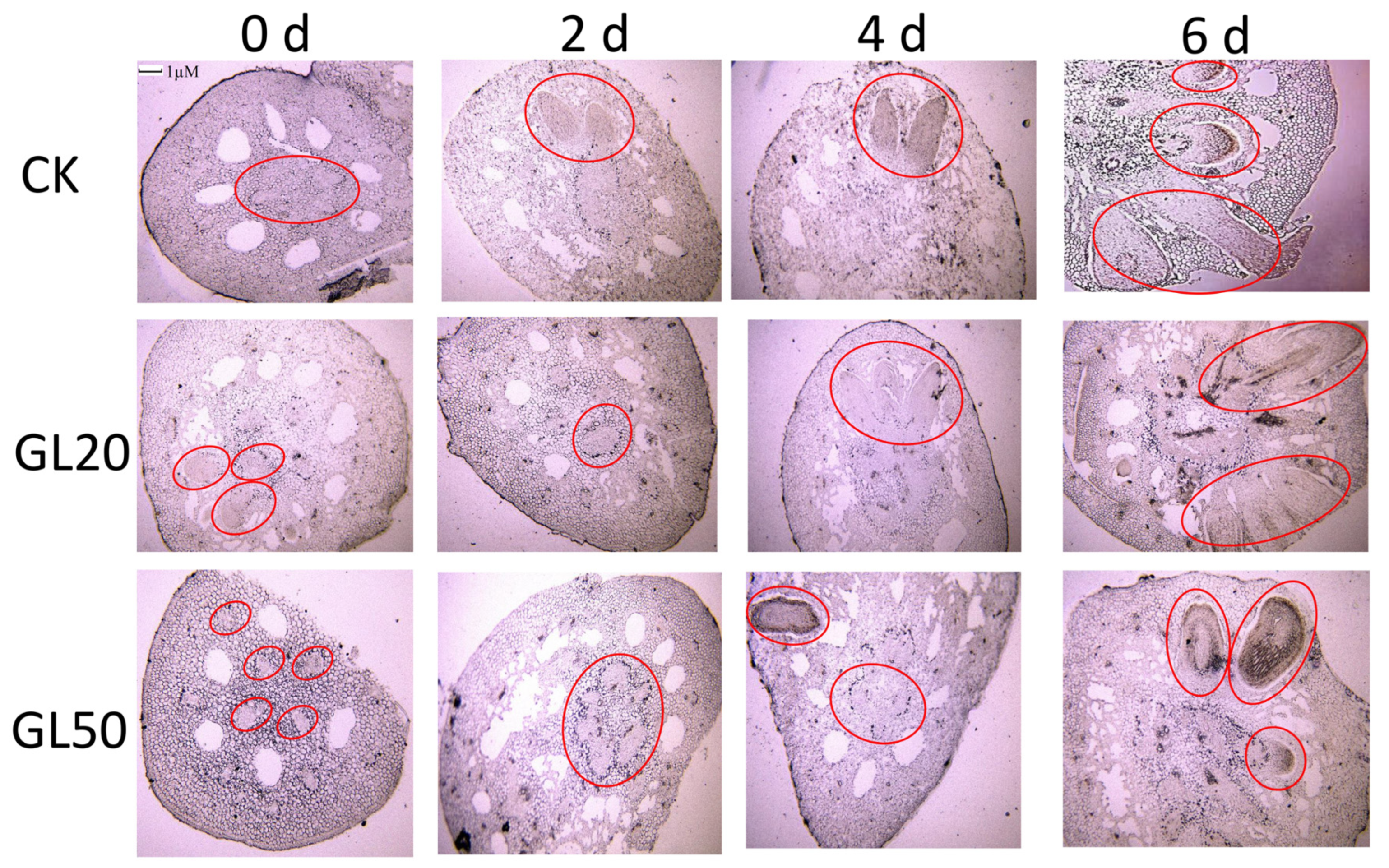
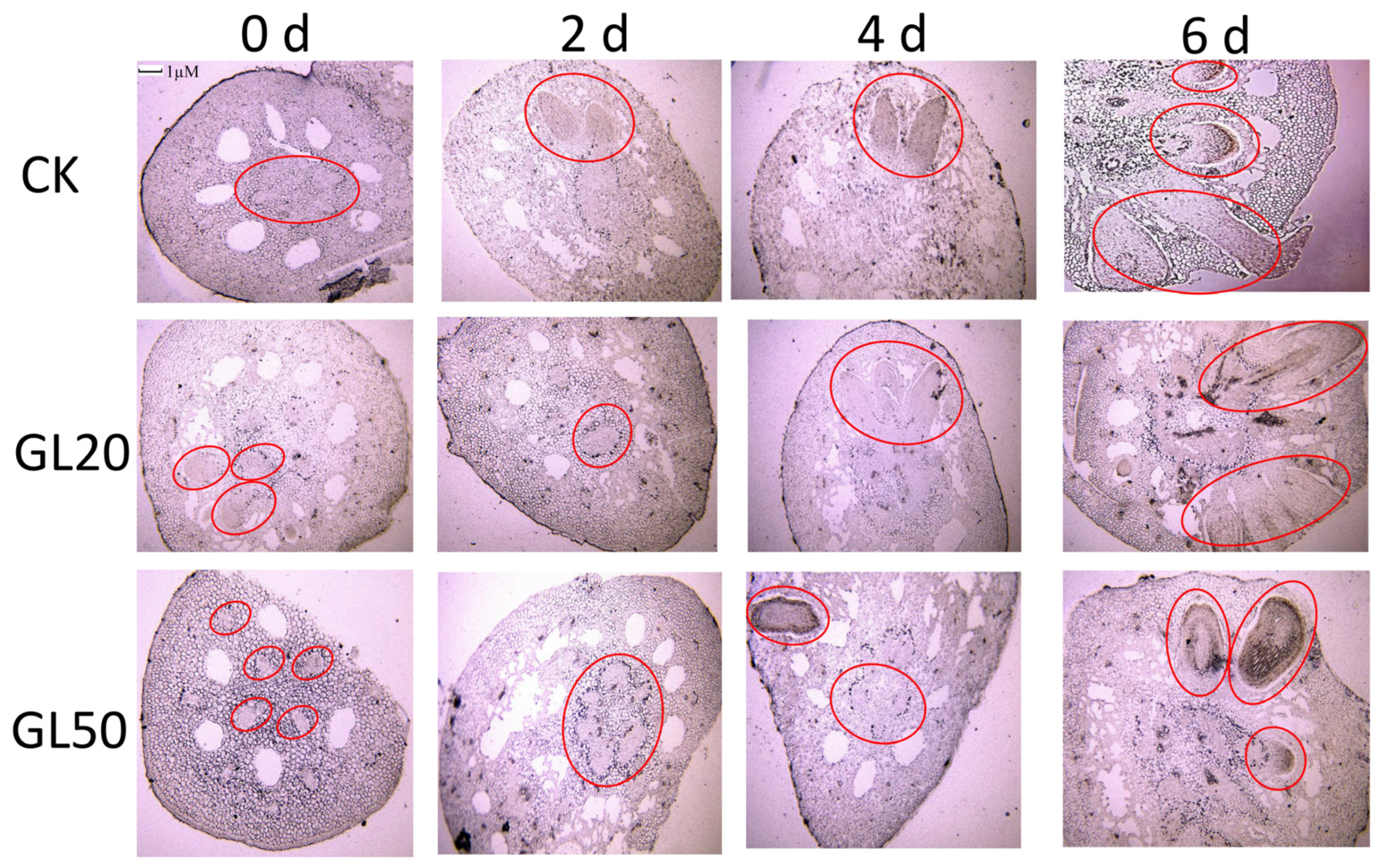
2.2. Observation of the Paraffin Sections
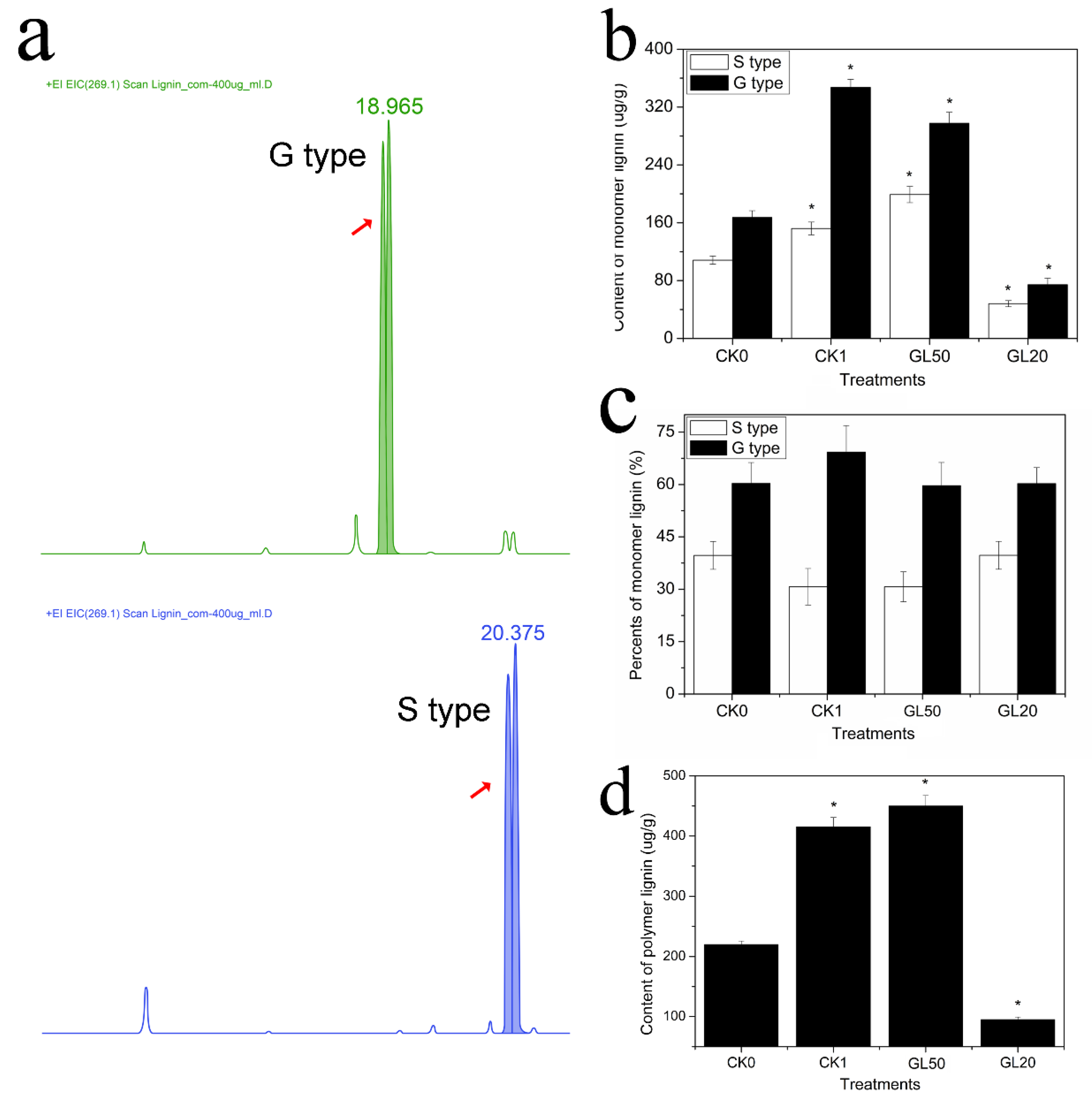
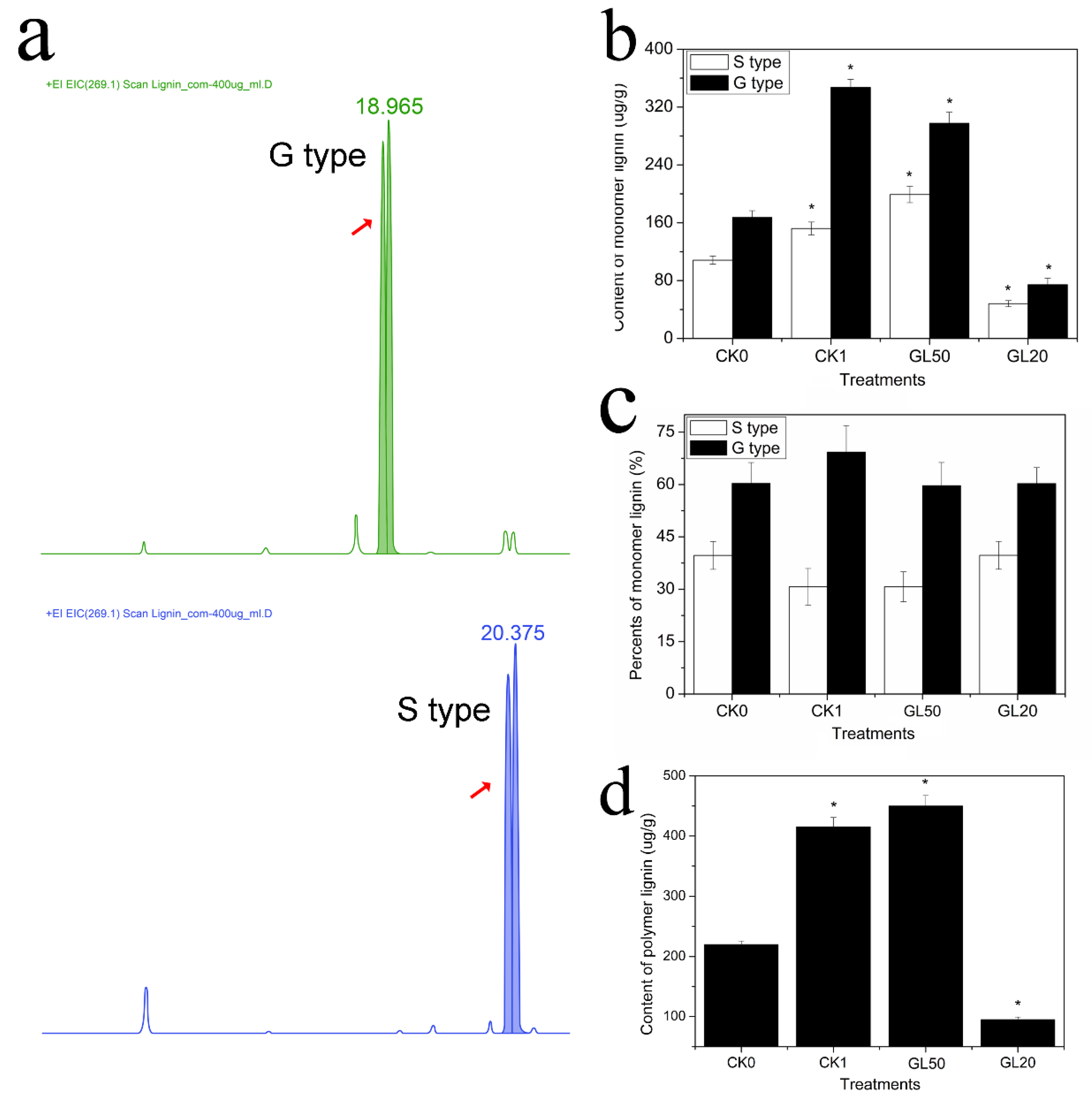
2.3. Determination of Monomer Lignin and Polymer Lignin Contents
2.4. The Change in Metabolites Related to Lignin Biosynthesis Owing to Sucrose Treatment
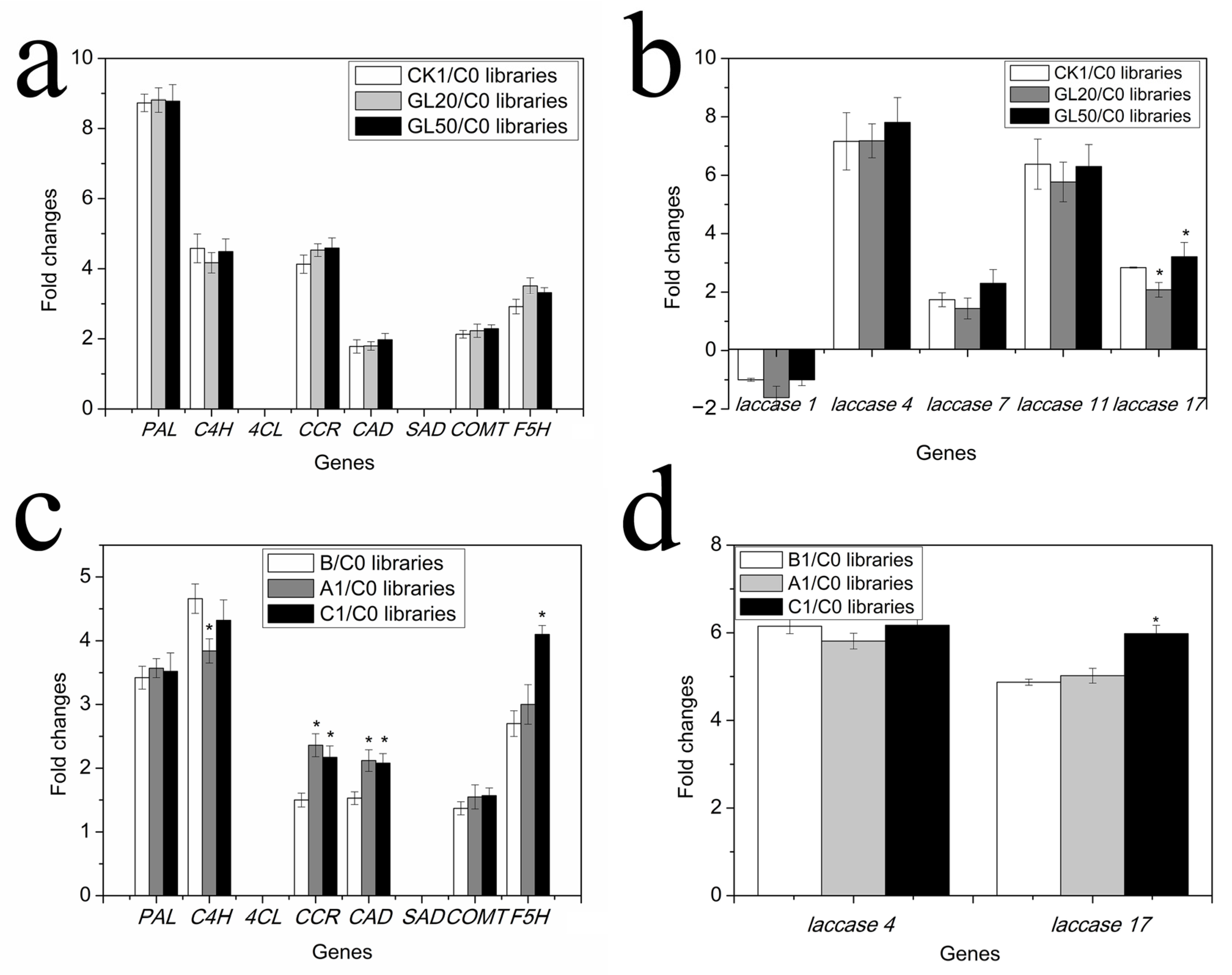
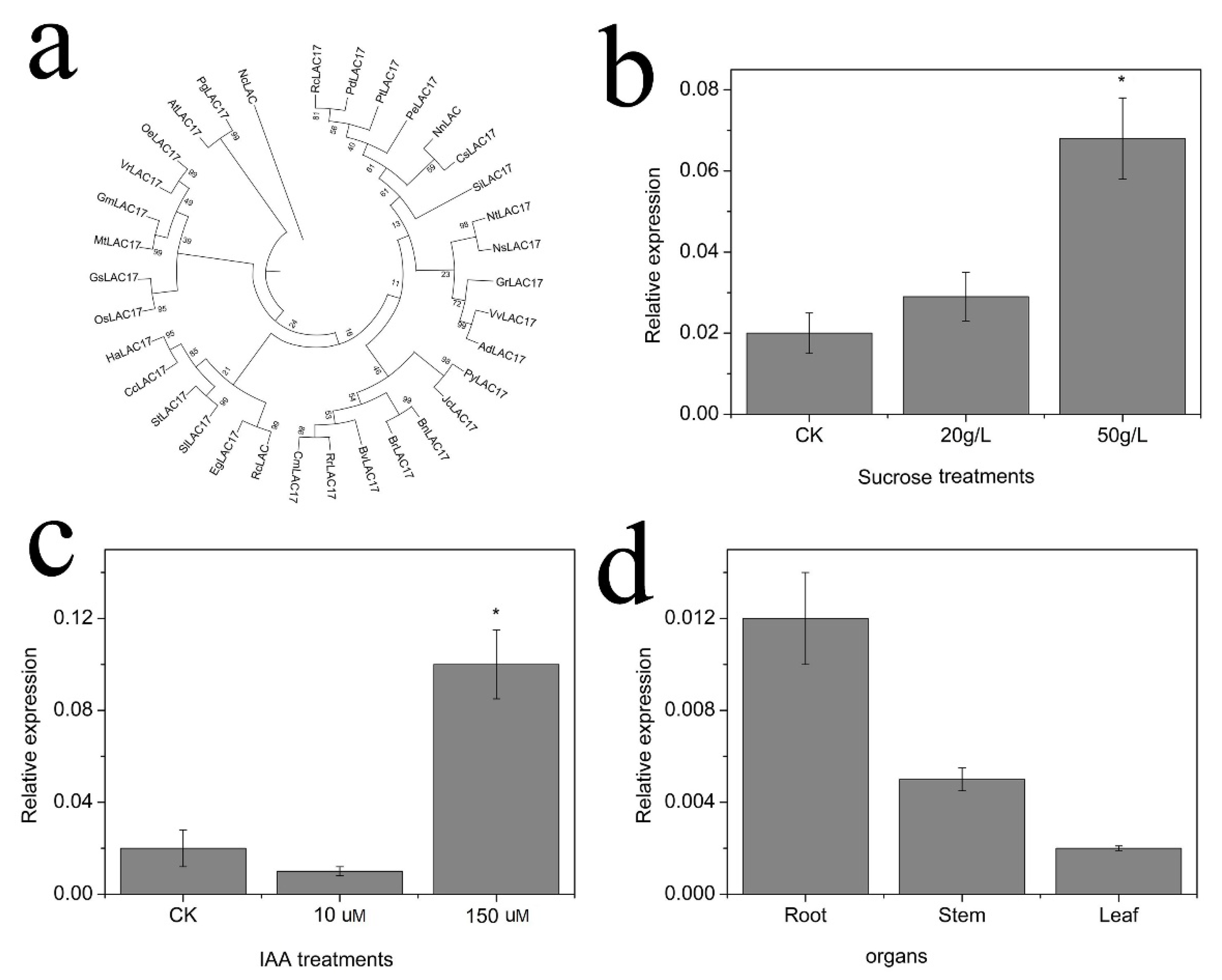
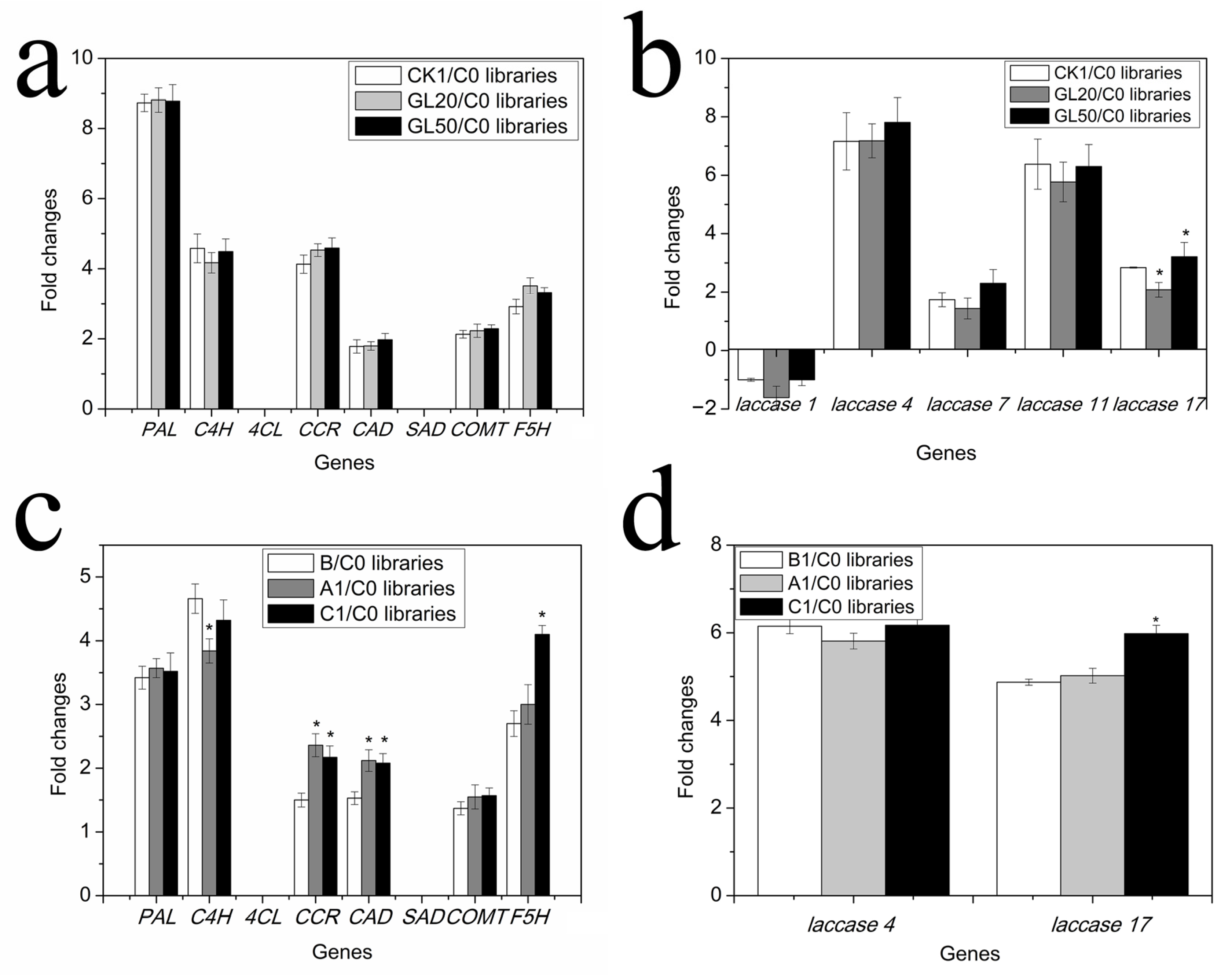
2.5. Expression of Genes Related to Monomer and Polymer Lignin Contents
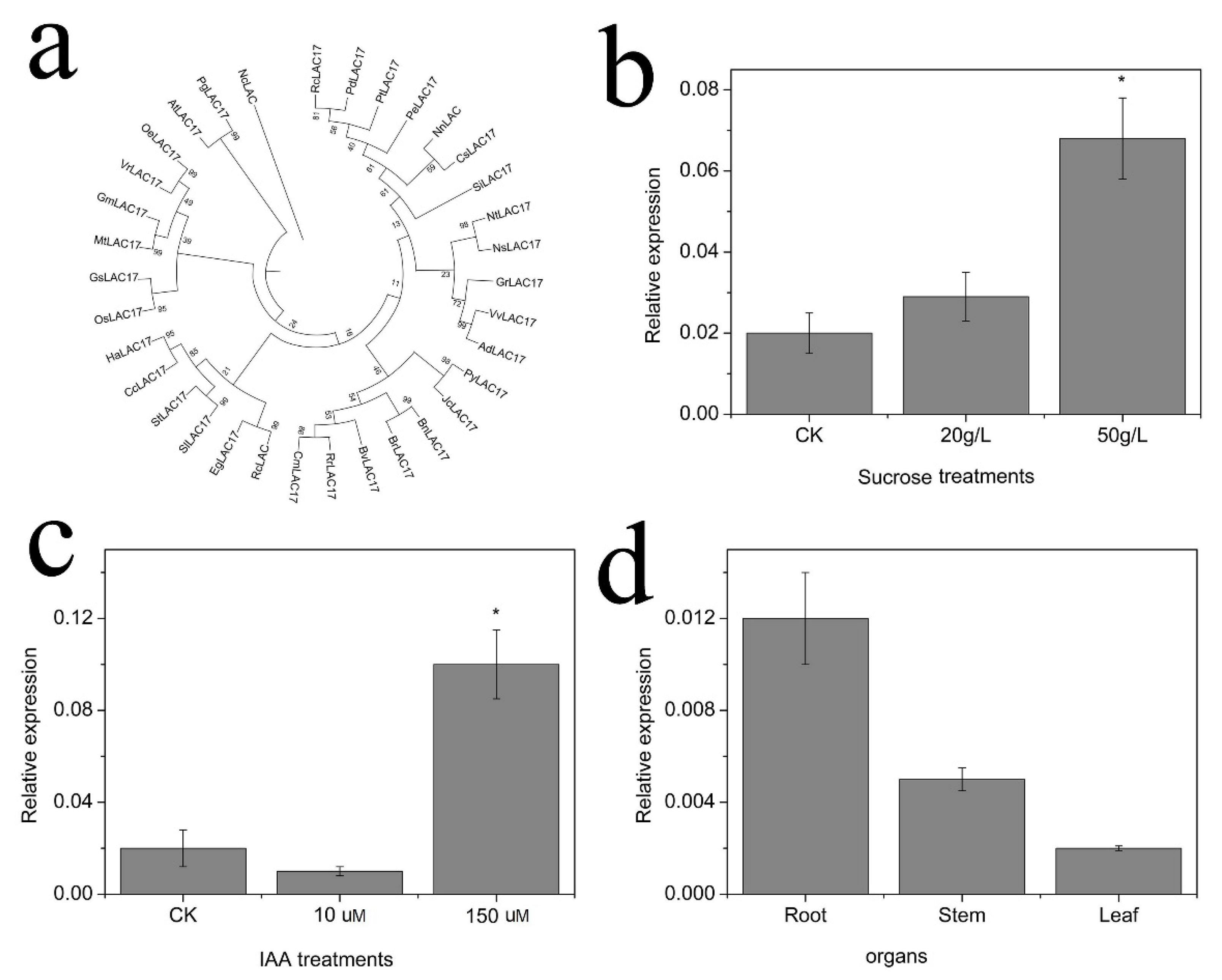
2.6. Cloning and Expression of NnLAC17
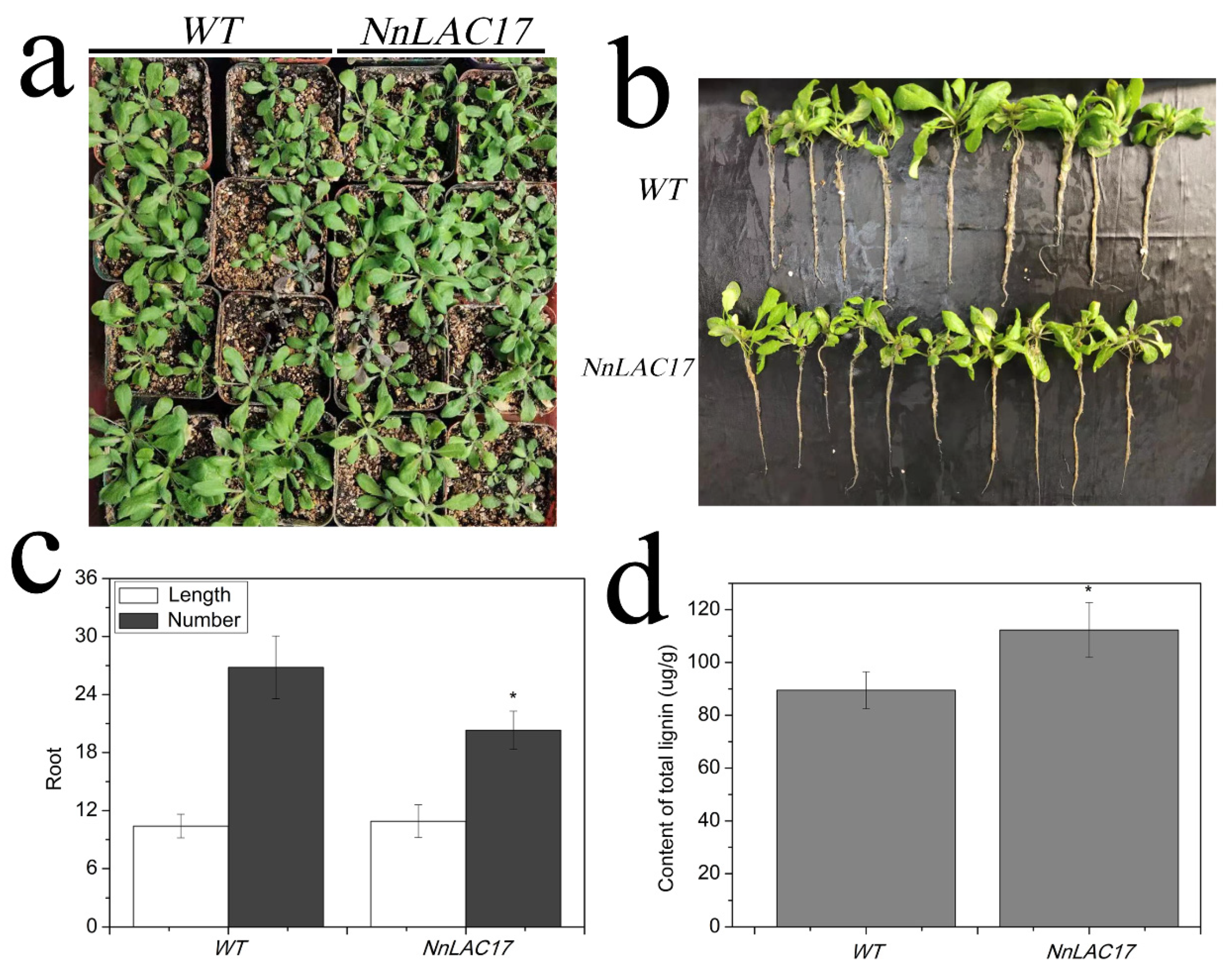
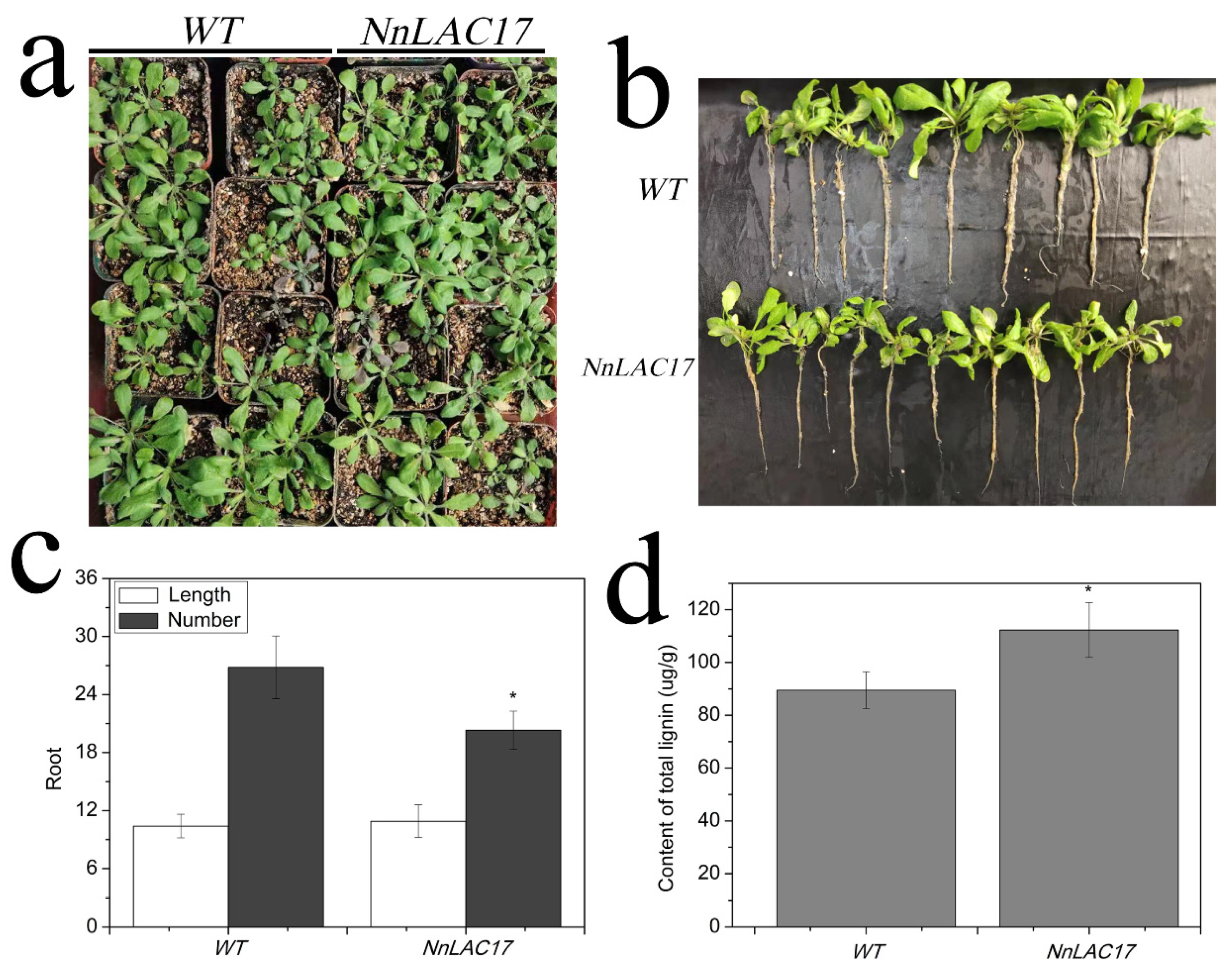
2.7. Functional Analysis of NnLAC17 in Transgenic Arabidopsis Thaliana
3. Discussion
4. Materials and Methods
4.1. Plant Growth
4.2. The Role of Sucrose in ARs Formation
4.3. Observation of the Paraffin Sections
4.4. Determination of Monomer Lignin and Polymer Lignin Contents
4.5. Metabolite Analysis of Lignin during ARs Formation
4.6. UPLC (Ultra Performance Liquid Chromatography) Conditions
4.7. ESI-QTRAP-MS/MS
4.8. RNA-seq Analysis of the Differentially Expressed Genes
4.9. Cloning and Sequence Analysis of NnLAC17
4.10. Expression Analysis of NnLAC17 in Lotus
4.11. Vector Construction
4.12. Identification of NnLAC17 Function in Arabidopsis Plants
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shen-Miller, J. Sacred lotus, the long-living fruits of China Antique. Seed Sci. Res. 2002, 12, 131–143. [Google Scholar]
- Ming, R.; Van Buren, R.; Liu, Y.; Yang, M.; Han, Y.; Li, L. Genome of the Long living sacred lotus (Nelumbo nucifera Gaertn.). Genome Biol. 2013, 14, R41. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.B.; Liu, H.Y.; Jiang, R.Z.; Li, S.Y. A Proteomics analysis of adventitious root formation after leaf removal in lotus (Nelumbo nucifera Gaertn.). ZNC 2018, 73, 375–389. [Google Scholar] [CrossRef]
- Kevers, C.; Hausman, J.F.; Faivre-Rampant, O.; Evers, D.; Gaspar, T. Hormonal control of adventitious rooting: Progress and questions. J. Appl. Bot. Angew. Bot. 1997, 71, 71–79. [Google Scholar]
- Li, S.; Xue, L.; Xu, S.; Feng, H.; An, L. Mediators, genes and signaling in adventitious rooting. Bot. Rev. 2009, 75, 230–247. [Google Scholar] [CrossRef]
- Rasmussen, A.; Hosseini, S.A.; Hajirezaei, M.R.; Druege, U.; Geelen, D. Adventitious rooting declines with the vegetative to reproductive switch and involves a changed auxin homeostasis. J. Exp. Bot. 2015, 66, 1437–1452. [Google Scholar] [CrossRef] [Green Version]
- Teale, W.D.; Paponov, I.A.; Palme, K. Auxin in action: Signaling, transport and the control of plant growth and development. Nat. Rev. Mol. Cell. Biol. 2006, 7, 847–859. [Google Scholar]
- Calvo-Polanco, M.; Senorans, J.; Zwiazek, J.J. Role of adventitious roots in water relations of tamarack (Larix laricina) seedlings exposed to flood. BMC Plant Biol. 2012, 12, 99. [Google Scholar] [CrossRef] [Green Version]
- Rovere, D.; Fattorini, L.; Angeli, S.D.; Veloccia, A.; Falasca, G.; Altamura, M.M. Auxin and cytokinin control formation of the quiescent centre in the adventitious root apex of Arabidopsis. Ann. Bot. 2013, 112, 1395–1407. [Google Scholar]
- Fukaki, H.; Tasaka, M. Hormone interactions during lateral root formation. Plant Mol. Biol. 2009, 69, 437–449. [Google Scholar]
- Mensuali-Sodi, A.; Panizza, M.; Tognoni, F. Endogenous ethylene requirement for adventitious root induction and growth in tomato cotyledons and lavandin microcuttings in vitro. Plant Growth Regul. 1995, 17, 205–212. [Google Scholar] [CrossRef]
- Pan, R.; Wang, J.; Tian, X. Influence of ethylene on adventitious root formation in mung bean hypocotyl cuttings. J. Plant Growth Regul. 2002, 36, 135–139. [Google Scholar]
- Steffens, B.; Wang, J.; Sauter, M. Interactions between ethylene, gibberellin and abscisic acid regulate emergence and growth rate of adventitious roots in deepwater rice. Planta 2006, 223, 604–612. [Google Scholar] [CrossRef]
- Riov, J.; Yang, S. Ethylene and auxin-ethylene interaction in adventitious root formation in mung bean (Vigna radiata) cuttings. J. Plant Growth Regul. 1989, 8, 131–141. [Google Scholar] [CrossRef]
- Negi, S.; Sukumar, P.; Liu, X.; Cohen, J.D.; Muday, G.K. Genetic dissection of the role of ethylene in regulating auxin-dependent lateral and adventitious root formation in tomato. Plant J. 2010, 61, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Visser, E.; Cohen, J.D.; Barendse, G.; Blom, C.; Voesenek, L. An ethylene-mediated increase in sensitivity to auxin induces adventitious root formation in flooded rumex palustrissm. Plant Physiol. 1996, 112, 1687–1692. [Google Scholar] [CrossRef] [Green Version]
- Cano-Delgado, A.I.; Metzlaff, K.; Bevan, M.W. The eli1 mutation reveals a link between cell expansion and secondary cell wall formation in Arabidopsis thaliana. Development 2000, 127, 3395–3405. [Google Scholar]
- Takahashi, F.; Sato-Nara, K.; Kobayashi, K.; Suzuki, M.; Suzuki, H. Sugar-induced adventitious roots in Arabidopsis seedlings. J. Plant Res. 2003, 116, 83–91. [Google Scholar]
- Cheng, L.B.; Zhao, M.R.; Hu, Z.B.; Liu, H.Y.; Li, S.Y. Comparative transcriptome analysis revealed the cooperative regulation of sucrose and IAA on adventitious root formation in lotus (Nelumbo nucifera Gaertn). BMC Genomics 2020, 21, 653. [Google Scholar]
- Barros, J.; Serk, H.; Granlund, I.; Pesquet, E. The cell biology of lignification in higher plants. Ann. Bot. 2015, 115, 1053–1074. [Google Scholar] [CrossRef] [Green Version]
- Shioya, T.; Akiyama, T.; Yokoyama, T.; Matsumot, Y. Formation rate of benzyl cation intermediate from p-Hydroxyphenyl, guaiacyl, or syringyl nucleus in acidolysis of lignin. J. Wood Chem. Tech. 2017, 37, 75–86. [Google Scholar] [CrossRef]
- Cho, H.; Chang, C.; Huang, L.; Tsai, J.; Liu, Z. Indole-3-butyric acid suppresses the activity of peroxidase while inducing adventitious roots in Cinnamomum kanehirae. Bot. Study 2011, 52, 153–160. [Google Scholar]
- Quiroga, M.; Guerrero, C.; Botella, M.A.; Barcelo, A.; Amaya, I.; Medina, M.I.; Alonso, F.J.; de Forchetti, S.M.; Tigier, H.; Valpuesta, V. A tomato peroxidase involved in the synthesis of lignin and suberin. Plant Physiol. 2000, 122, 1119–1127. [Google Scholar] [CrossRef] [Green Version]
- Chou, C.H.; Huang, Y.C.; Liu, Z. Peroxidase genes differentially respond to auxin during the formation of adventitious roots in soybean hypocotyl. Plant Growth Regul. 2010, 60, 151–161. [Google Scholar] [CrossRef]
- Syros, T.; Yupsanis, T.; Zafiriadis, H.; Economou1, A. Activity and isoforms of peroxidases, lignin and anatomy, during adventitious rooting in cuttings of Ebenus cretica L. J. Plant Physiol. 2004, 161, 69–77. [Google Scholar] [CrossRef]
- Singh, V.; Sergeeva, L.; Ligterink, W.; Aloni, R.; Zemach, H.; Doron-Faigenboim, A.; Yang, J.; Zhang, P.; Shabtai, S.; Firon, N. Gibberellin promotes sweetpotato root vascular lignification and reduces storage-root formation. Front. Plant Sci. 2019, 10, 1320. [Google Scholar] [CrossRef] [PubMed]
- Ponniah, S.K.; Thimmapuram, J.; Bhide, K.; Kalavacharla, V.; Manoharan, M. Comparative analysis of the root transcription of cultivated sweetpotato (Ipomoea batatas [L.] Lam) and its wild ancestor (Ipomoea trifida [Kunth] G. Don). BMC Plant Biol. 2017, 17, 9. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Shi, R.; Leng, Y.; Zhou, Y. Transcriptomic analysis reveals the gene expression profile that specifically responds to IBA during adventitious rooting in mung bean seedlings. BMC Genom. 2016, 17, 43. [Google Scholar] [CrossRef] [Green Version]
- Piacentinia, D.; Ronzana, M.; Fattorinia, L.; DellaRoverea, F.; Massimib, L.; Altamuraa, M.M. Nitric oxide alleviates cadmium-but not arsenic-induced damages in rice roots. Plant Physiol. Biochem. 2020, 151, 729–742. [Google Scholar] [CrossRef]
- Cheng, L.B.; Han, Y.Y.; Liu, H.Y.; Jiang, R.Z.; Li, S.Y. Transcriptomic analysis reveals ethylene’s regulation involved in adventitious roots formation in lotus (Nelumbo nucifera Gaertn.). Acta Physiol. Plant. 2019, 41, 97. [Google Scholar]
- Cheng, L.B.; Han, Y.Y.; Zhao, M.R.; Li, S.Y. Gene expression profiling reveals the effects of light on adventitious root formation in lotus seedlings (Nelumbo nucifera Gaertn.). BMC Genomics 2020, 21, 707. [Google Scholar]
- Cheng, L.B.; Jiang, R.Z.; Yang, J.J.; Xu, X.Y.; Zeng, H.T.; Li, S.Y. Transcriptome profiling reveals an IAA-regulated response to adventitious root formation in lotus seedling. Z. Naturforsch. 2018, 73, 229–240. [Google Scholar]
- Hobbie, L.; Estelle, E. The axr4 auxin-resistant mutants of Arabidopsis thaliana define a gene important for root gravitropism and lateral root initiation. Plant J. 1995, 7, 211–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casimiro, I.; Marchant, A.; Bhalerao, R.P.; Beeckman, T.; Dhooge, S.; Swarup, R.; Graham, N.; Inzé, D.; Sandberg, G.; Casero, P.J.; et al. Auxin transport promotes Arabidopsis lateral root nitiation. Plant Cell. 2001, 13, 843–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchant, A.; Bhalerao, R.; Casimiro, I.; Eklöf, J.; Casero, P.J.; Bennet, M.; Sandberg, G. AUX1 promotes lateral root formation by facilitating indole-3-acetic acid distribution between sink and source tissues in the Arabidopsis seedling. Plant Cell. 2002, 14, 589–597. [Google Scholar] [CrossRef] [Green Version]
- Khadr, A.; Wang, G.; Wang, Y.; Zhang, R.; Wang, X.; Xu, Z.; Tian, Y.-S.; Xiong, A.-S. Effects of auxin (indole-3-butyric acid) on growth characteristics, lignification, and expression profiles of genes involved in lignin biosynthesis in carrot taproot. PeerJ 2020, 8, e10492. [Google Scholar] [CrossRef]
- Brunetti, C.; Fini, A.; Sebastiani, F.; Gori, A. Modulation of phytohormone signaling: A primary function of flavonoids in plant environment interactions. Front. Plant Sci. 2018, 9, 1042. [Google Scholar] [CrossRef] [Green Version]
- Vanholme, R.; De Meester, B.; Ralph, J.; Boerjan, W. Lignin biosynthesis and its integration into metabolism. Curr. Opin. Biotech. 2019, 56, 230–239. [Google Scholar] [CrossRef]
- Dolan, W.L.; Dilkes, B.P.; Stout, J.M.; Bonawitz, N.D.; Chapple, C. Mediato omplex subunits MED2, MED5, MED16, and MED23 genetically interact in the regulation of phenylpropanoid biosynthesis. Plant Cell. 2017, 29, 3269–3285. [Google Scholar] [CrossRef]
- Schilmiller, A.L.; Stout, J.; Weng, J.K.; Humphreys, J.; Ruegger, M.O.; Chapple, C. Mutations in the cinnamate 4-hydroxylase gene impact metabolism, growth and development in Arabidopsis. Plant J. 2009, 60, 771–782. [Google Scholar] [CrossRef]
- Singh, V.; Zemach, H.; Shabtai, S.; Aloni, R.; Yang, J.; Zhang, P.; Sergeeva, L.; Ligterink, W.; Firon, N. Proximal and distal parts of sweet potato adventitious roots display differences in root architecture, lignin, and starch metabolism and their developmental fates. Front. Plant Sci. 2021, 11, 609923. [Google Scholar] [CrossRef] [PubMed]
- Houari, I.E.; Beirs, C.V.; Arents, H.E.; Han, H.B.; Chanoca, A.; Opdenacker, D.; Pollier, J.; Storme, V.; Steenackers, W.; Quareshy, M.; et al. Seedling developmental defects upon blocking CINNAMATE-4-HYDROXYLASE are caused by perturbations in auxin transport. New Phytol. 2021, 230, 2275–2291. [Google Scholar] [CrossRef] [PubMed]
- Durkovic, J.; Kanuchova, A.; Kacık, F.; Solar, R.; Lengyelova, A. Genotype- and age-dependent patterns of lignin and cellulose in regenerants derived from 80-year-old trees of black mulberry (Morus nigra L.). Plant Cell Tissue Organ. Cult. 2011, 108, 359–370. [Google Scholar] [CrossRef]
- Eliyahu, A.; Duman, Z.; Sherf, S.; Genin, O.; Cinnamon, Y.; Abu-Abied, M.; Weinstain, R.; Dag, A.; Sadot, E. Vegetative propagation of elite Eucalyptus clones as food source for honeybees (Apis mellifera); adventitious roots versus callus formation. Israel J. Plant Sci. 2020, 67, 83–97. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Teng, R.; Wang, W.; Wang, Y.; Shen, W.; Zhuang, J. Identification of genes revealed differential expression profiles and lignin accumulation during leaf and stem development in tea plant [Camellia sinensis (L.) O. Kuntze]. Protoplasma 2019, 256, 359–370. [Google Scholar] [CrossRef]
- Liu, Q.; Luo, L.; Zheng, L. Lignins: Biosynthesis and biological functions in plants. Int. J. Mol. Sci. 2018, 19, 335. [Google Scholar] [CrossRef] [Green Version]
- Han, M.; Yang, N.; Wan, Q.; Teng, R.; Duan, A.; Wang, Y.; Zhuang, J. Exogenous melatonin positively regulates lignin biosynthesis in Camellia sinensis. Int. J. Biol. Macromol. 2021, 179, 485–499. [Google Scholar] [CrossRef]
- Zhang, X.; Lu, M.; Ludlow, R.A.; Ma, W.; An, H. Transcriptome analysis reveals candidate genes for dietary fiber metabolism in Rosa roxburghii fruit grown under different light intensities. Hort. Environ. Biol. 2021, 62, 751–764. [Google Scholar] [CrossRef]
- Lu, C.; Yang, T.; Zhang, Y.; Miao, X.; Jin, C.; Xu, X. Genome-wide analyses and expression patterns under abiotic stress of LAC gene family in pear (Pyrus bretschneideri). Plant Biotech. Rep. 2021, 15, 403–416. [Google Scholar] [CrossRef]
- Shao, C.; Qiu, C.; Qian, Y.; Liu, G. Nitrate deficiency decreased photosynthesis and oxidation-reduction processes, but increased cellular, transport, lignin biosynthesis and flavonoid metabolism revealed by RNA-Seq in Oryza sativa leaves. PLoS ONE 2020, 15, e0235975. [Google Scholar] [CrossRef]
- Wang, G.; Huang, Y.; Zhang, X.; Xu, Z.; Wang, F.; Xiong, A. Transcriptome-based identification of genes revealed differential expression profiles and lignin accumulation during root development in cultivated and wild carrots. Plant Cell Rep. 2016, 35, 1743–1755. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Zhong, R.; Morrison, W.H.; Himmelsbach, D.S. Caffeoyl coenzyme A O-methyltransferase and lignin biosynthesis. Phytochemistry 2001, 57, 1177–1185. [Google Scholar] [CrossRef]
- Duan, A.; Feng, K.; Wang, G.; Liu, J.; Xu, Z.; Xiong, A. Elevated gibberellin enhances lignin accumulation in celery (Apium graveolens L.) leaves. Protoplasma 2019, 256, 777–788. [Google Scholar] [CrossRef]
- Hoffmann, L.; Maury, S.; Martz, F.; Geoffroy, P.; Legrand, M. Purification, cloning, and properties of an acyltransferase controlling shikimate and quinate ester intermediate in phenylpropanoid metabolism. J. Biol. Chem. 2003, 278, 95–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Gu, M.; Lai, Z.; Fan, B.; Shi, K.; Zhou, Y.; Yu, J.-Q.; Chen, Z. Functional analysis of the Arabidopsis PAL gene family in plant growth, development, and response to environmental stress. Plant Physiol. 2018, 153, 1526–1538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gui, J.; Shen, J.; Li, L. Functional characterization of evolutionarily divergent 4-coumarate: Coenzyme a ligases in rice. Plant Physiol. 2011, 157, 574–586. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.H.; Kim, C.Y.; Song, W.K.; Park, D.S.; Kwon, S.Y.; Lee, H.S.; Bang, J.-W.; Kwak, S.-S. Overexpression of sweetpotato swpa4 peroxidase results in increased hydrogen peroxide production and enhances stress tolerance in tobacco. Planta 2008, 227, 867–881. [Google Scholar] [CrossRef]
- Hiraide, H.; Tobimatsu, Y.; Yoshinaga, A.; Lam, P.Y.; Kobayashi, M.; Matsushita, Y.; Fukushima, K.; Takabe, K. Localised laccase activity modulates distribution of lignin polymers in gymnosperm compression wood. New Phytol. 2021, 230, 2186–2199. [Google Scholar] [CrossRef]
- Niu, Z.; Li, G.; Hu, H.; Lv, J.; Zheng, Q.; Liu, J.; Wan, D. A gene that underwent adaptive evolution, LAC2 (LACCASE), in Populus euphratica improves drought tolerance by improving water transport capacity. Hort. Res. 2021, 8, 88. [Google Scholar] [CrossRef]
- Li, S.; Xue, L.; Xu, S.; Fent, H.; An, H. IBA-induced changes in antioxidant enzymes during adventitious rooting in mung bean seedlings: The role of H2O2. Environ. Exp. Bot. 2009, 66, 442–450. [Google Scholar] [CrossRef]
- Abaturov, B.D.; Kolesnikov, M.P.; Likhnova, O.P.; Petrishchev, B.I.; Nikonova, O.A. Use of plant forage lignin and silica as digestibility indicators for quantitative estimation of food consumption by free-ranging saigas. Zoologicheskii Zhurnal 1997, 76, 106–113. [Google Scholar]
- Tao, J.M.; Li, S.; Ye, F.Y.; Zhou, Y.; Lei, L.; Zhao, G.H. Lignin-An underutilized, renewable and valuable material for food industry. Crit. Rev. Food Sci. Nutr. 2020, 60, 2011–2033. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cui, J.H.; Yin, T.T.; Sun, L.Z.; Li, G.X. Activated effect of lignin on α-amylase. Food Chem. 2013, 141, 2229–2237. [Google Scholar] [CrossRef]
- Kaewtatip, K.; Menut, P.; Auvergne, R.; Tanrattanakul, V.; Morel, M.H.; Guilbert, S. Interactions of kraft lignin and wheat gluten during biomaterial processing: Evidence for the role of phenolic groups. J. Agric. Food Chem. 2010, 58, 4185–4192. [Google Scholar] [CrossRef] [PubMed]
- Teng, R.M.; Wang, Y.X.; Li, H.; Lin, S.J.; Liu, H.; Zhang, J. Effects of shading on lignin biosynthesis in the leaf of tea plant (Camellia sinensis (L.) O. Kuntze). Mol. Genet. Genom. 2021, 296, 165–177. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | p Value | Fold Change | Types |
|---|---|---|---|
| CK1/CK0 libraries | |||
| p-Coumaric acid | 0.0112 | 0.2743 | Down-regulated |
| p-Coumaraldehyde | NA | NA | |
| Sinapic acid | NA | NA | |
| Sinapyl alcohol | NA | NA | |
| p-Coumaryl alcohol | NA | NA | |
| Caffeyl alcohol | NA | NA | |
| Caffeyl aldehyde | NA | NA | |
| Sinapinaldehyde | 0.0015 | 3.4747 | Up-regulated |
| L-Phenylalanine | 0.0052 | 1.8344 | Up-regulated |
| Coniferyl alcohol | 0.0225 | 23.0678 | Up-regulated |
| Caffeate | 0.0305 | 1.4309 | Up-regulated |
| Ferulic acid | 0.0564 | 1.6682 | Up-regulated |
| 4-Hydroxy-3-methoxycinnamaldehyde | 0.0019 | 10.5479 | Up-regulated |
| Cinnamic acid | 0.0036 | 2.5194 | Up-regulated |
| GL20/CK0 | |||
| p-Coumaric acid | 0.0407 | 0.1107 | Down-regulated |
| p-Coumaraldehyde | NA | NA | |
| Sinapic acid | NA | NA | |
| Caffeate | 0.0369 | 1.156 | |
| Sinapyl alcohol | NA | NA | |
| p-Coumaryl alcohol | NA | NA | |
| Caffeyl alcohol | NA | NA | |
| Caffeyl aldehyde | NA | NA | |
| Sinapinal dehyde | 0.0147 | 7.2121 | Up-regulated |
| L-Phenylalanine | 0.0497 | 1.6486 | Up-regulated |
| Coniferyl alcohol | 0.01762 | 19.5263 | Up-regulated |
| Ferulic acid | 0.0113 | 2.3898 | Up-regulated |
| 4-Hydroxy-3-methoxycinnamaldehyde | 0.0415 | 16.7289 | Up-regulated |
| Cinnamic acid | 0.02470 | 2.1252 | Up-regulated |
| GL50/CK0 | |||
| p-Coumaric acid | 0.04766 | 0.079 | Down-regulated |
| p-Coumaraldehyde | NA | NA | |
| Sinapic acid | NA | NA | |
| Caffeate | 0.0325 | 0.899 | |
| Sinapyl alcohol | NA | NA | |
| p-Coumaryl alcohol | NA | NA | |
| Caffeyl alcohol | NA | NA | |
| Caffeyl aldehyde | NA | NA | |
| Sinapinal dehyde | 0.0057 | 5.2929 | Up-regulated |
| L-Phenylalanine | 0.0073 | 1.5573 | Up-regulated |
| Coniferyl alcohol | 0.0072 | 20.4520 | Up-regulated |
| Ferulic acid | 0.03586 | 1.9201 | Up-regulated |
| 4-Hydroxy-3-methoxycinnamaldehyde | 0.0001 | 17.3515 | Up-regulated |
| Cinnamic acid | 0.0004 | 2.4017 | Up-regulated |
| GL50/GL20 | |||
| p-Coumaric acid | 0.0386 | 0.7140 | Down-regulated |
| p-Coumaraldehyde | NA | NA | |
| Sinapic acid | NA | NA | |
| Caffeate | 0.0339 | 0.7823 | Down-regulated |
| Sinapyl alcohol | NA | NA | |
| p-Coumaryl alcohol | NA | NA | |
| Caffeyl alcohol | NA | NA | |
| Caffeyl aldehyde | NA | NA | |
| Sinapinal dehyde | 0.0157 | 0.7338 | Down-regulated |
| L-Phenylalanine | 0.0014 | -------- | |
| Coniferyl alcohol | 0.0082 | -------- | |
| Ferulic acid | 0.0254 | 0.7034 | Down-regulated |
| 4-Hydroxy-3-methoxycinnamaldehyde | 0.0497 | -------- | |
| Cinnamic acid | 0.0145 | -------- | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, L.; Zhao, C.; Zhao, M.; Han, Y.; Li, S. Lignin Synthesis, Affected by Sucrose in Lotus (Nelumbo nucifera) Seedlings, Was Involved in Regulation of Root Formation in the Arabidopsis thanliana. Int. J. Mol. Sci. 2022, 23, 2250. https://doi.org/10.3390/ijms23042250
Cheng L, Zhao C, Zhao M, Han Y, Li S. Lignin Synthesis, Affected by Sucrose in Lotus (Nelumbo nucifera) Seedlings, Was Involved in Regulation of Root Formation in the Arabidopsis thanliana. International Journal of Molecular Sciences. 2022; 23(4):2250. https://doi.org/10.3390/ijms23042250
Chicago/Turabian StyleCheng, Libao, Chen Zhao, Minrong Zhao, Yuyan Han, and Shuyan Li. 2022. "Lignin Synthesis, Affected by Sucrose in Lotus (Nelumbo nucifera) Seedlings, Was Involved in Regulation of Root Formation in the Arabidopsis thanliana" International Journal of Molecular Sciences 23, no. 4: 2250. https://doi.org/10.3390/ijms23042250
APA StyleCheng, L., Zhao, C., Zhao, M., Han, Y., & Li, S. (2022). Lignin Synthesis, Affected by Sucrose in Lotus (Nelumbo nucifera) Seedlings, Was Involved in Regulation of Root Formation in the Arabidopsis thanliana. International Journal of Molecular Sciences, 23(4), 2250. https://doi.org/10.3390/ijms23042250