
Global Identification and Characterization of C2 Domain-Containing Proteins Associated with Abiotic Stress Response in Rice (Oryza sativa L.)

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Results
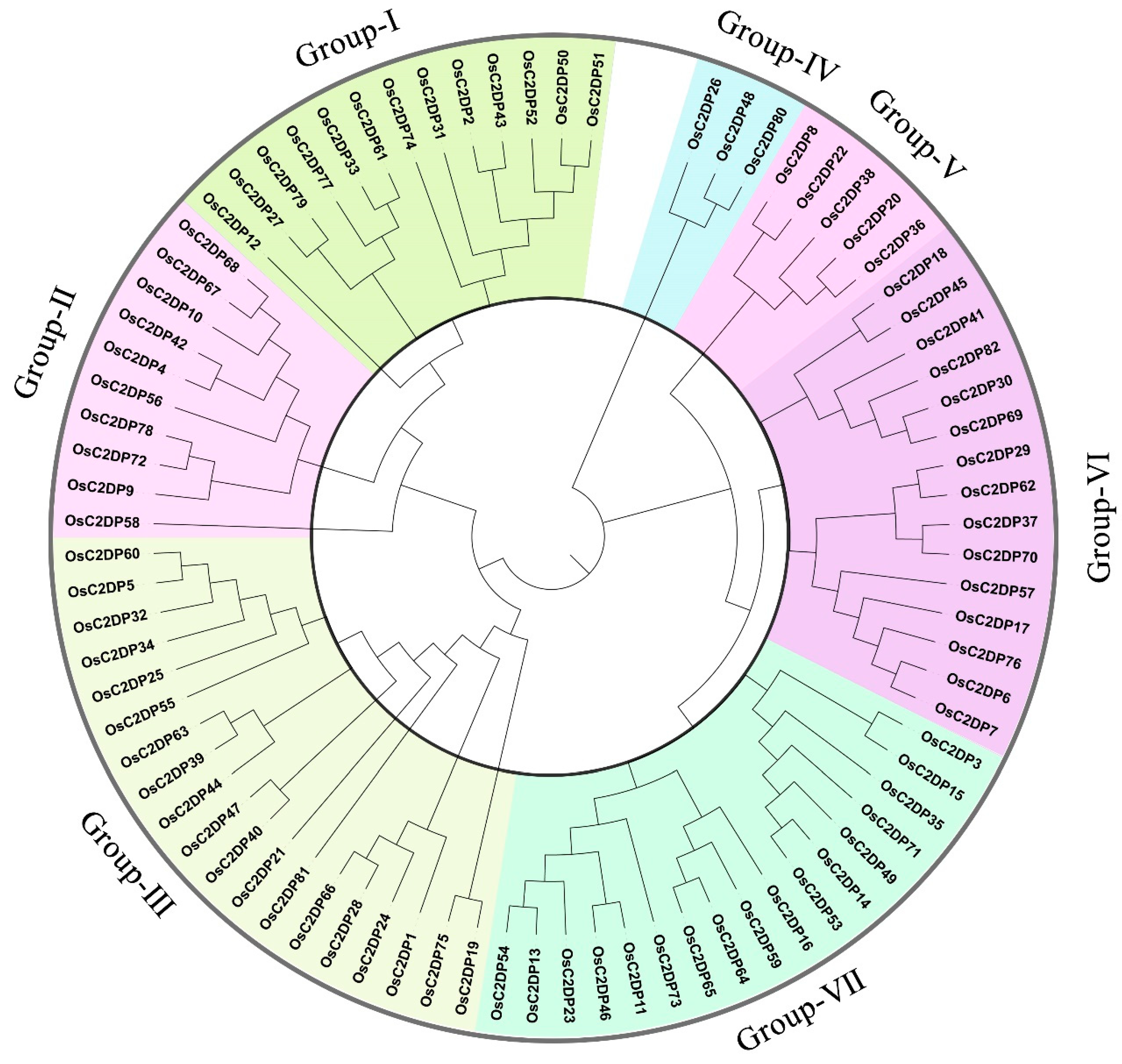
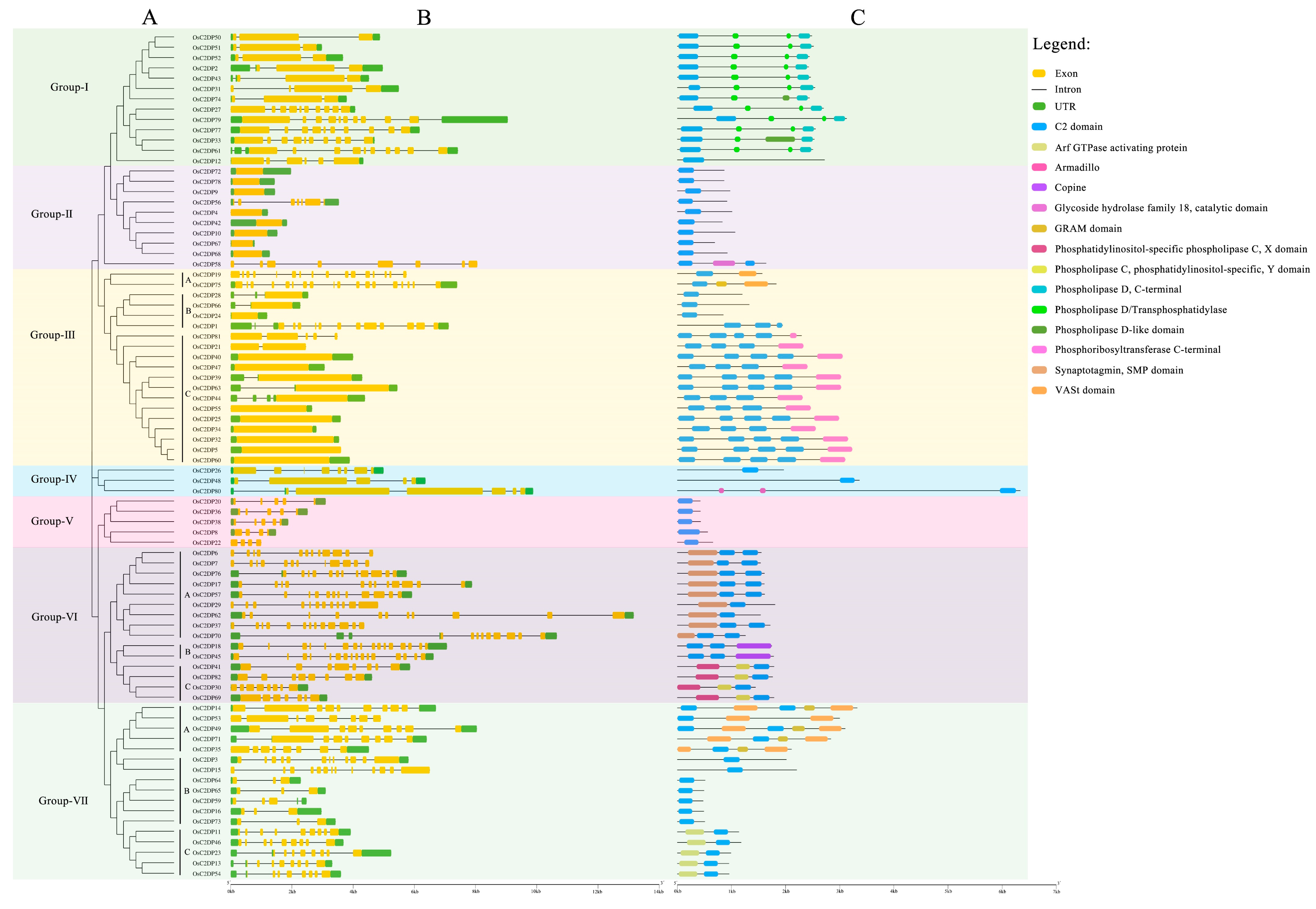
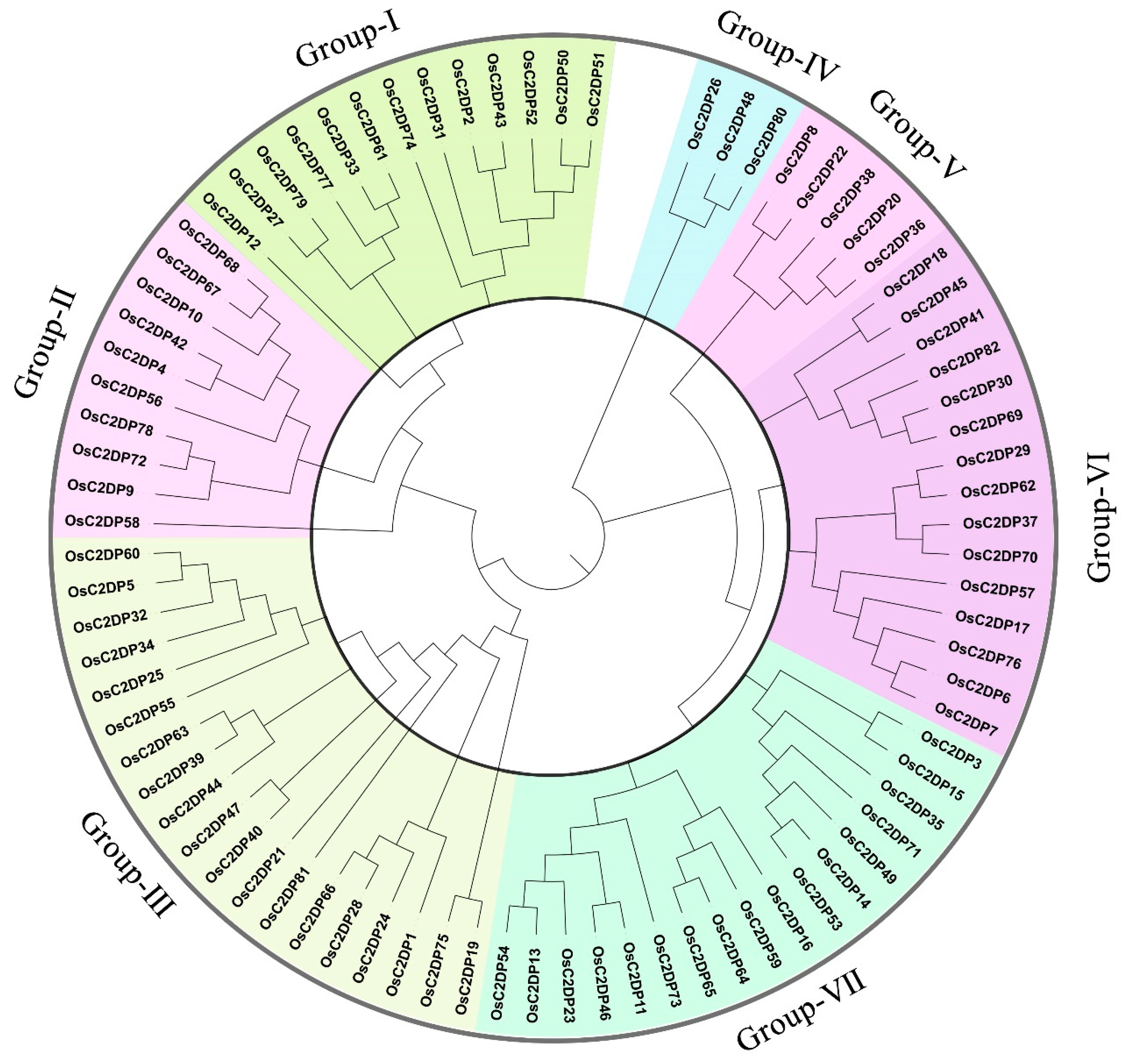
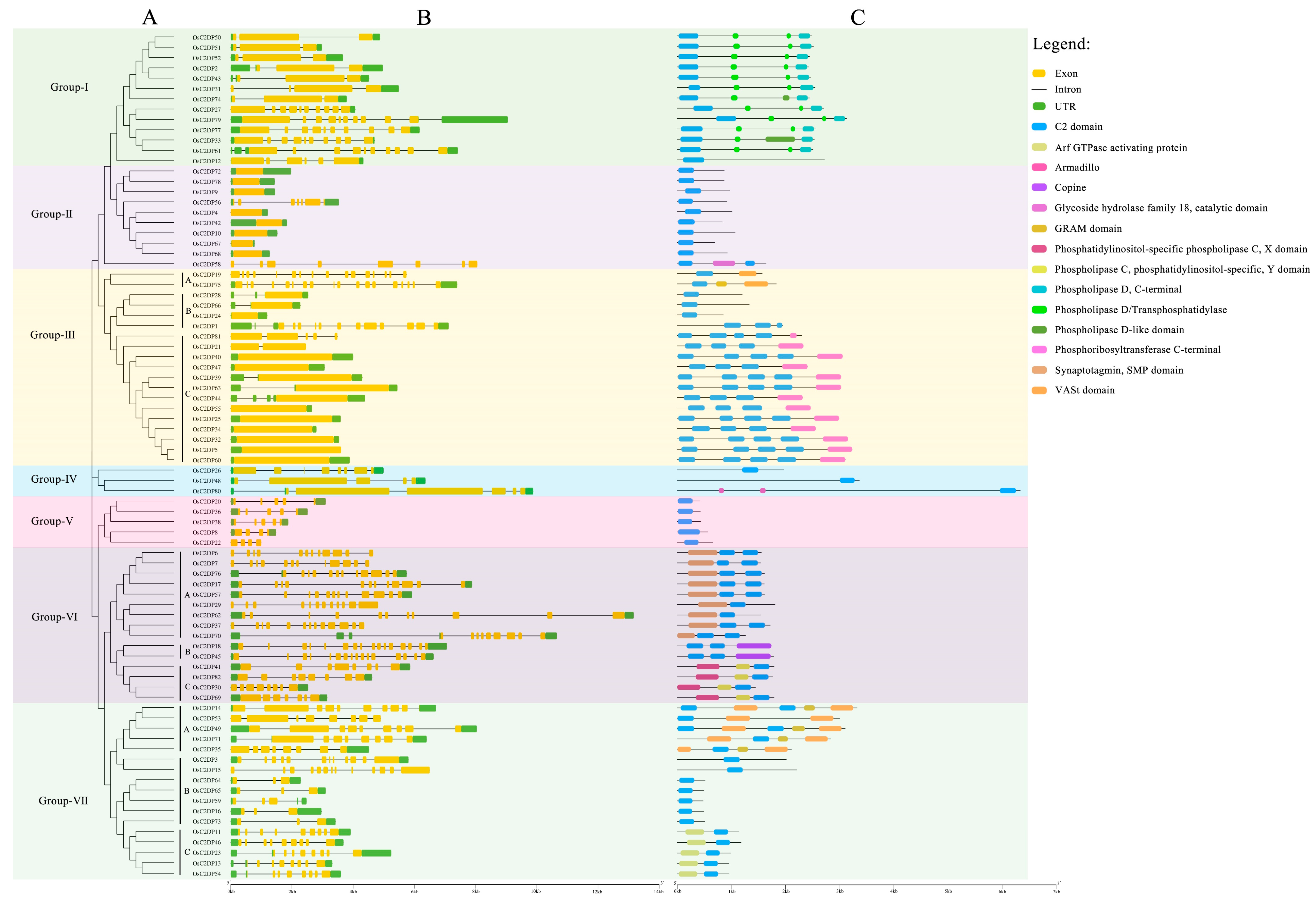
2.1. Identification, Phylogenetic, and Sequence Analyses of OsC2DPs
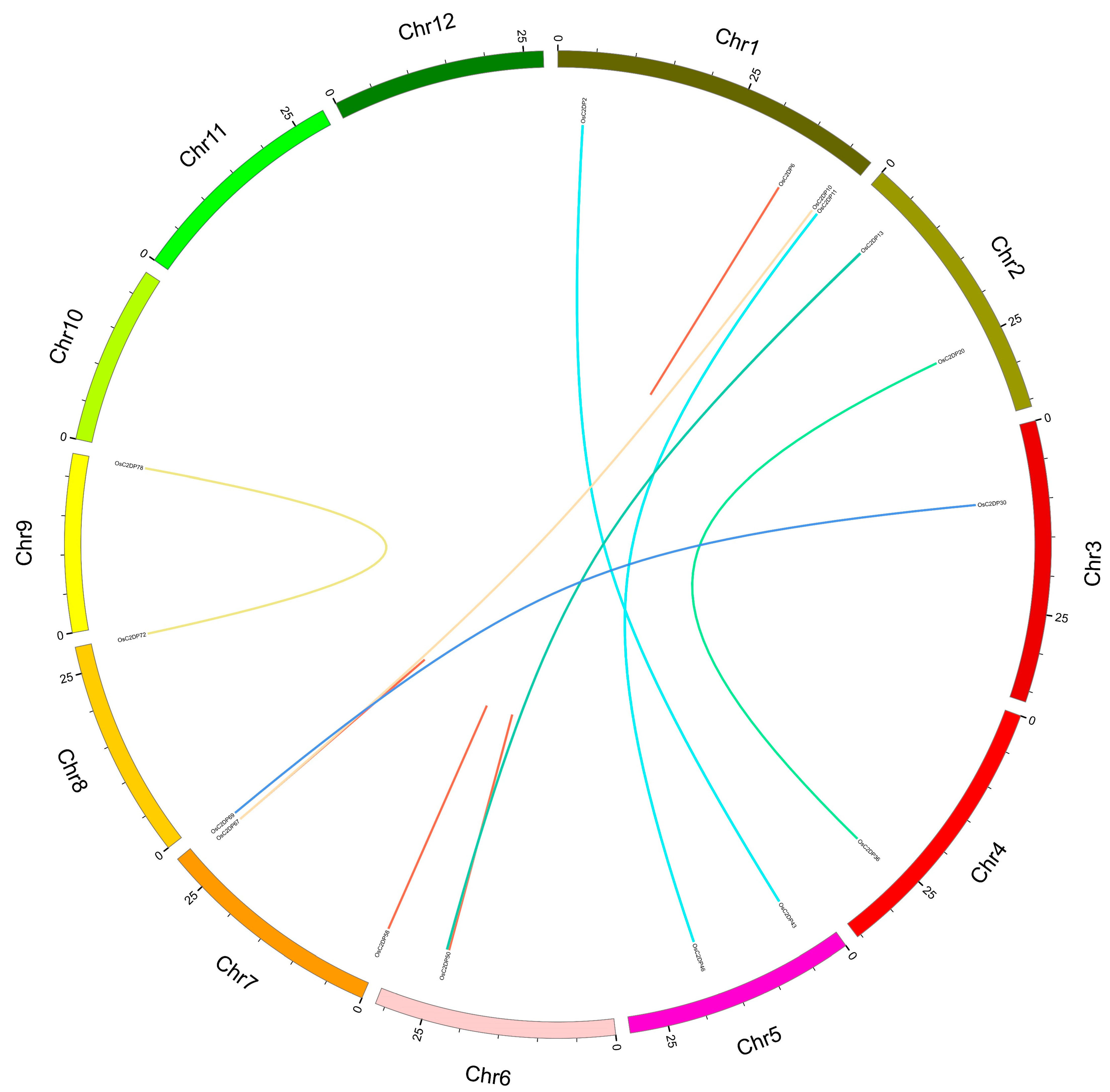
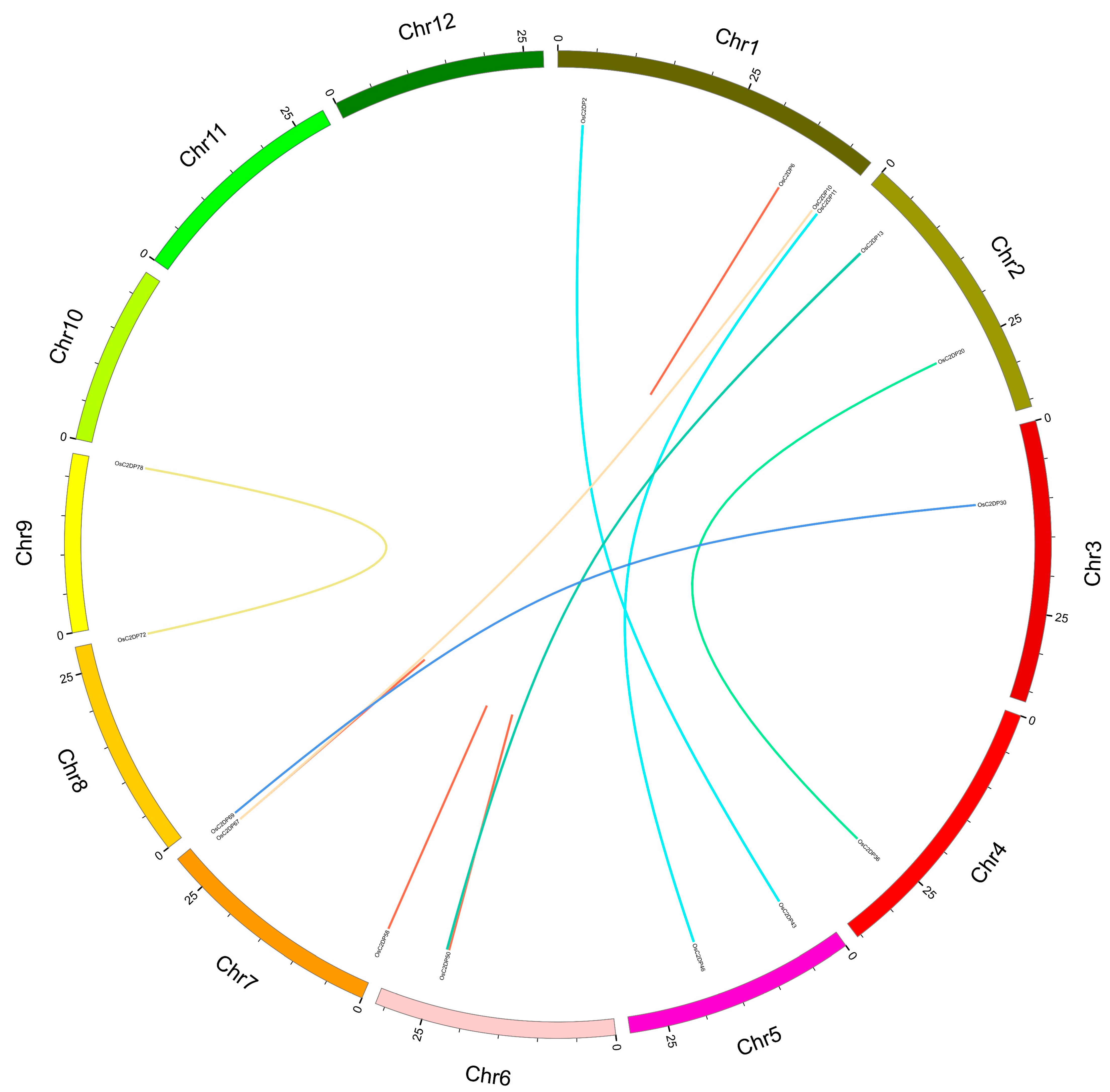
2.2. Synteny Analysis and Ka/Ks Ratio of OsC2DPs in Rice and Others Genome
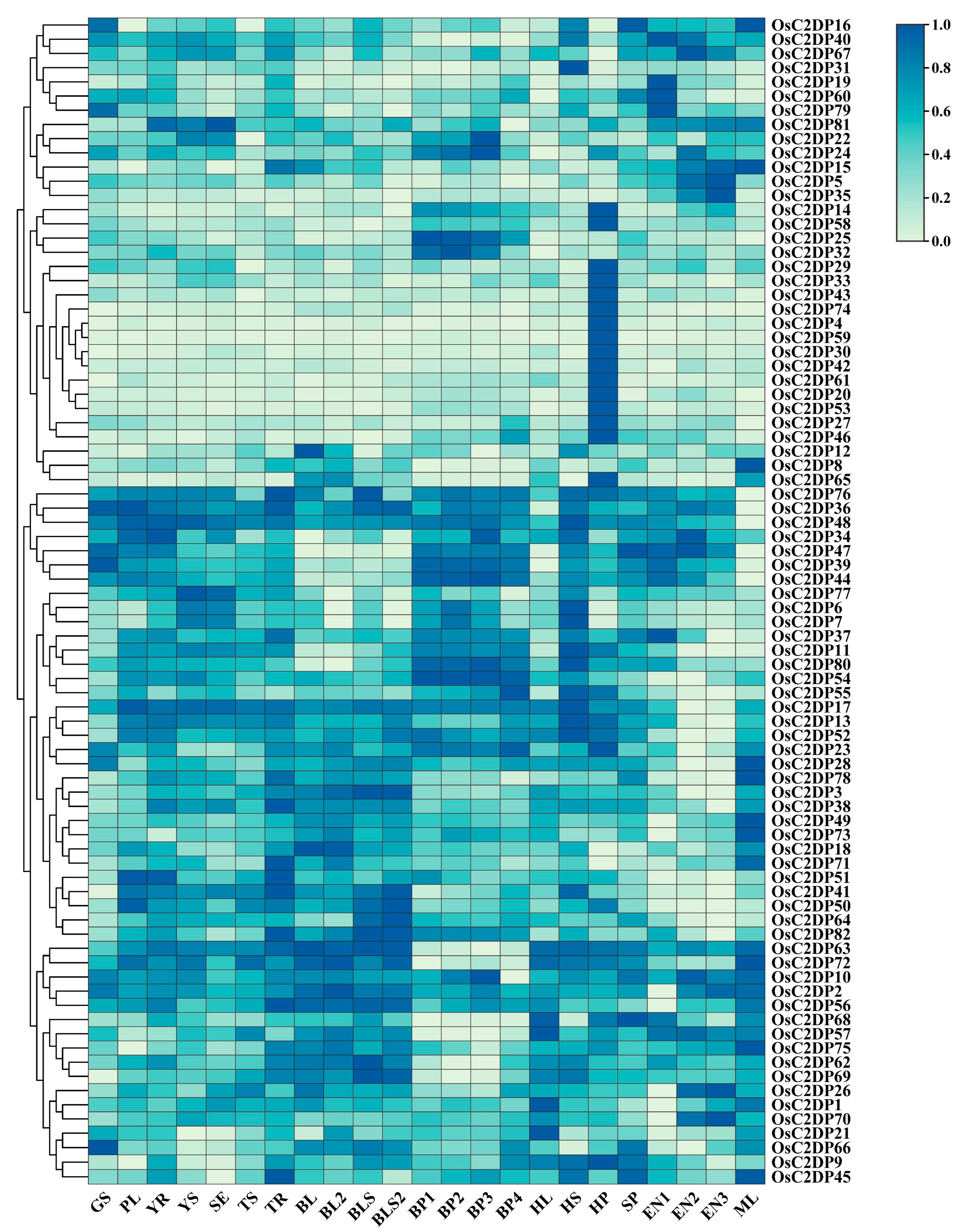
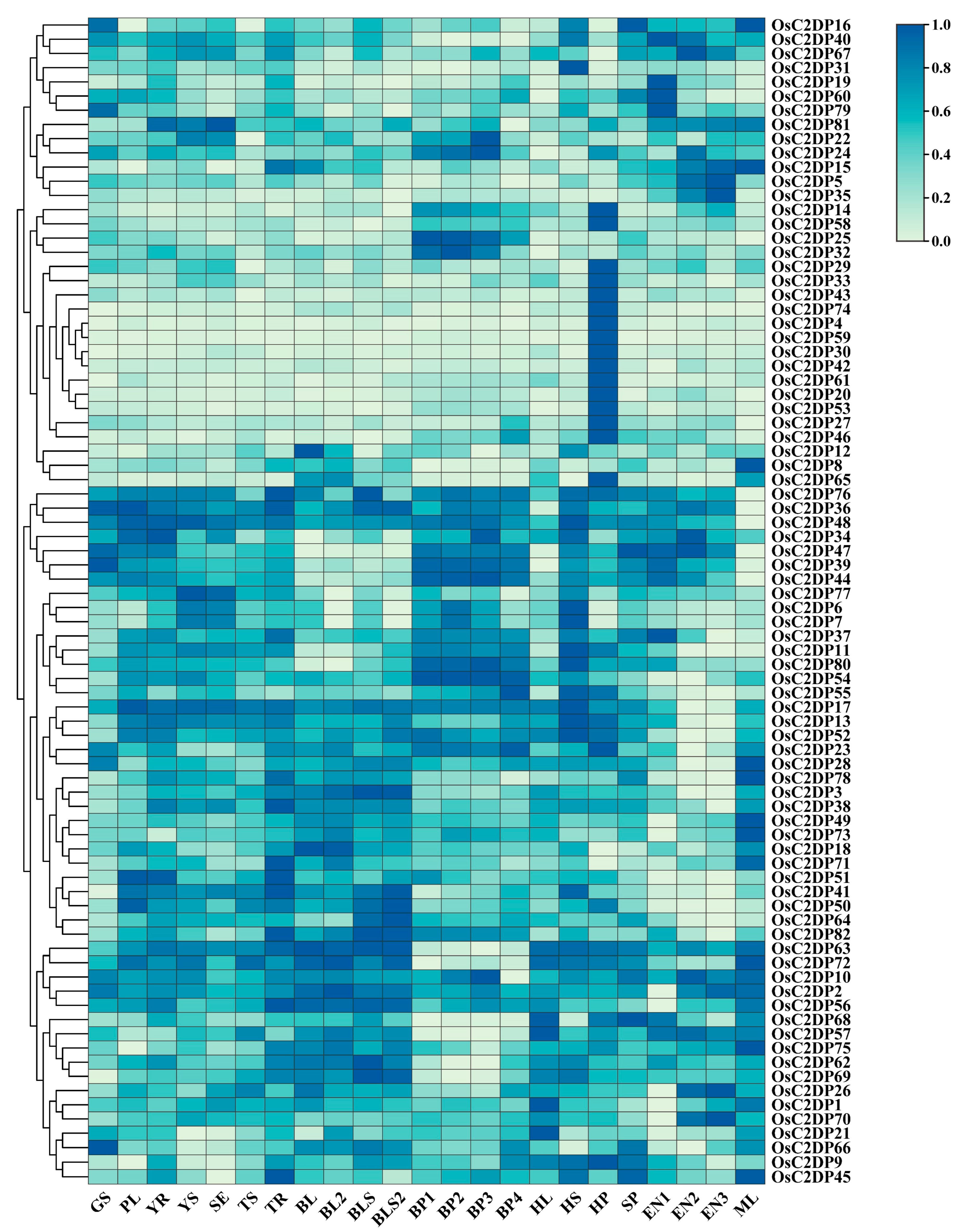
2.3. Expression Profile Analysis of OsC2DPs in Rice Growth Stage
2.4. Prediction and Analysis of the Cis-Acting Elements in Promoter Regions of OsC2DPs
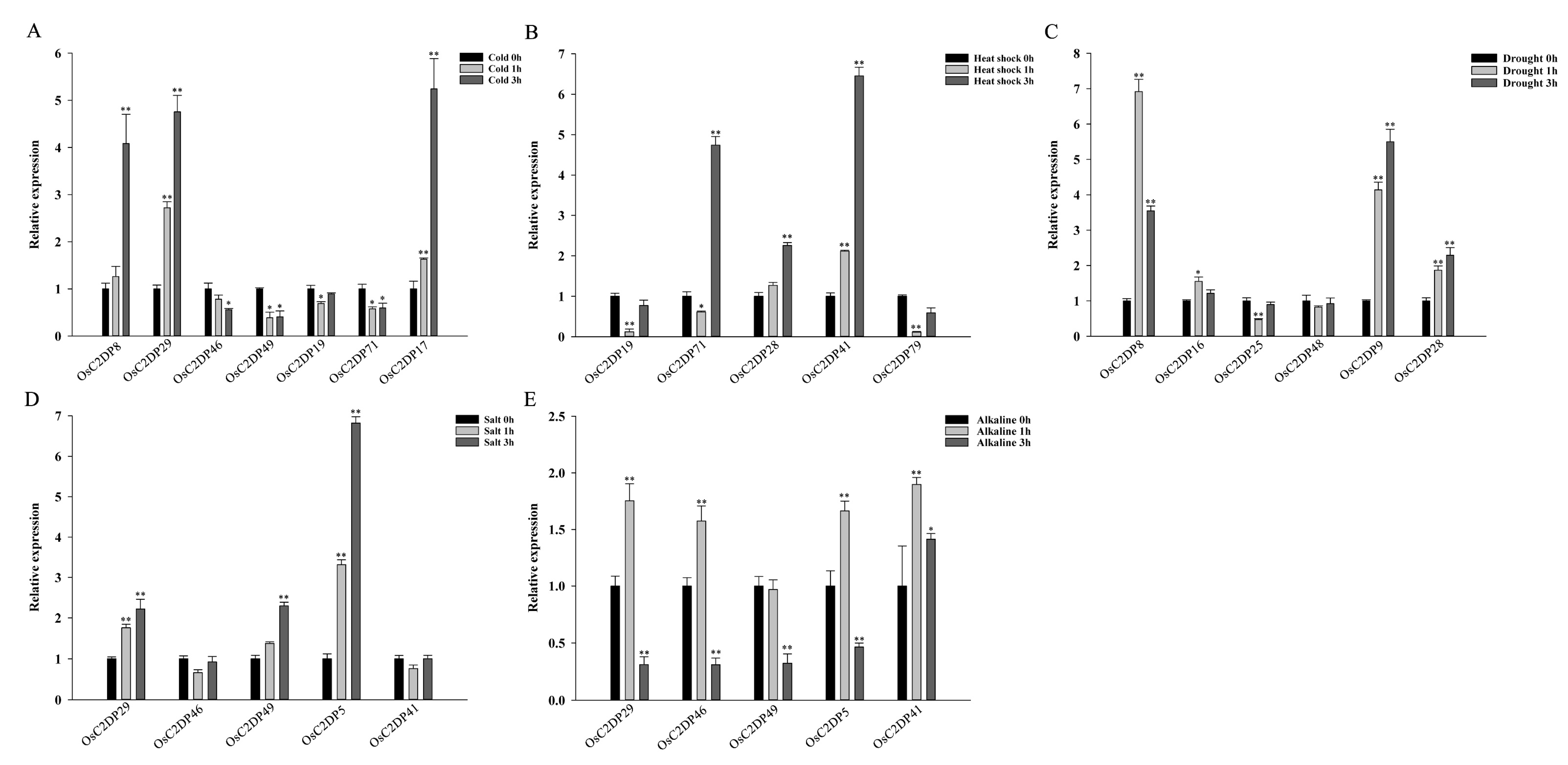
2.5. Expression Analysis of OsC2DPs Involved Abiotic and Exogenous Phytohormone Treatments
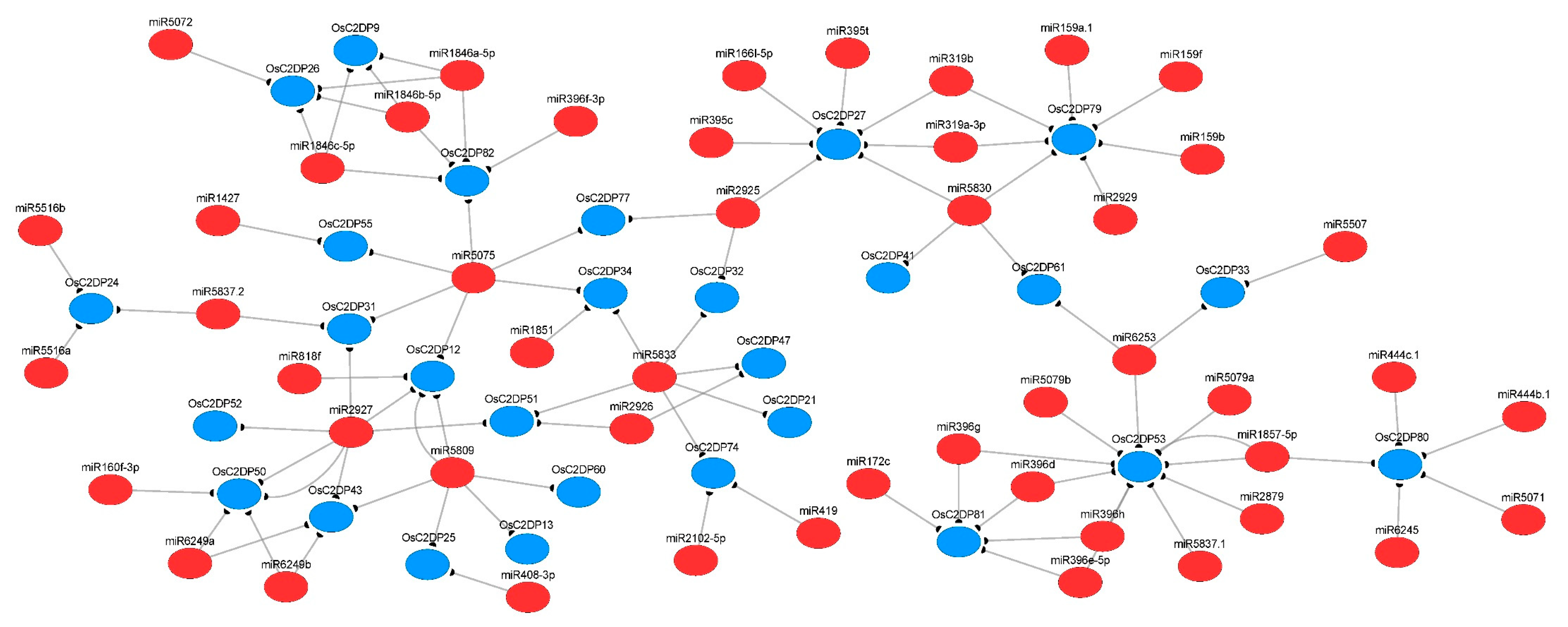
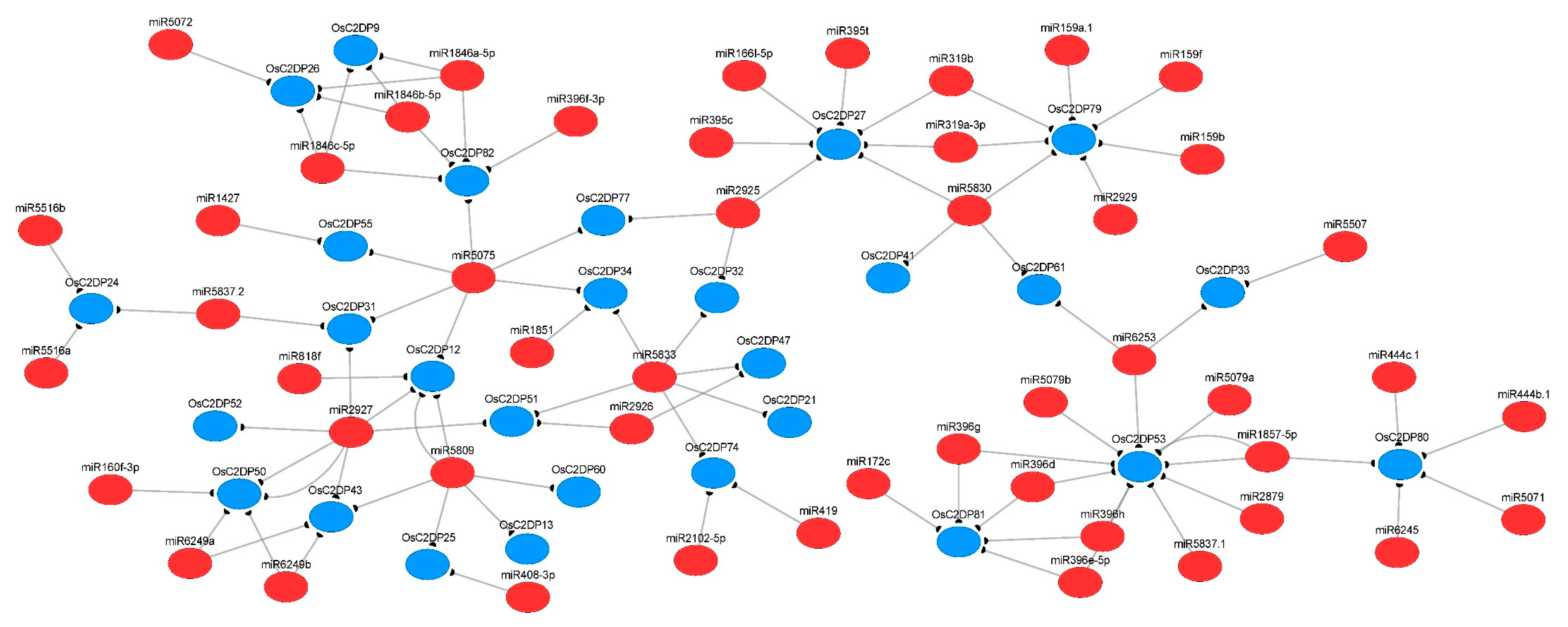
2.6. Prediction of miRNA-Targets and Enrichment Analysis of OsC2DPs
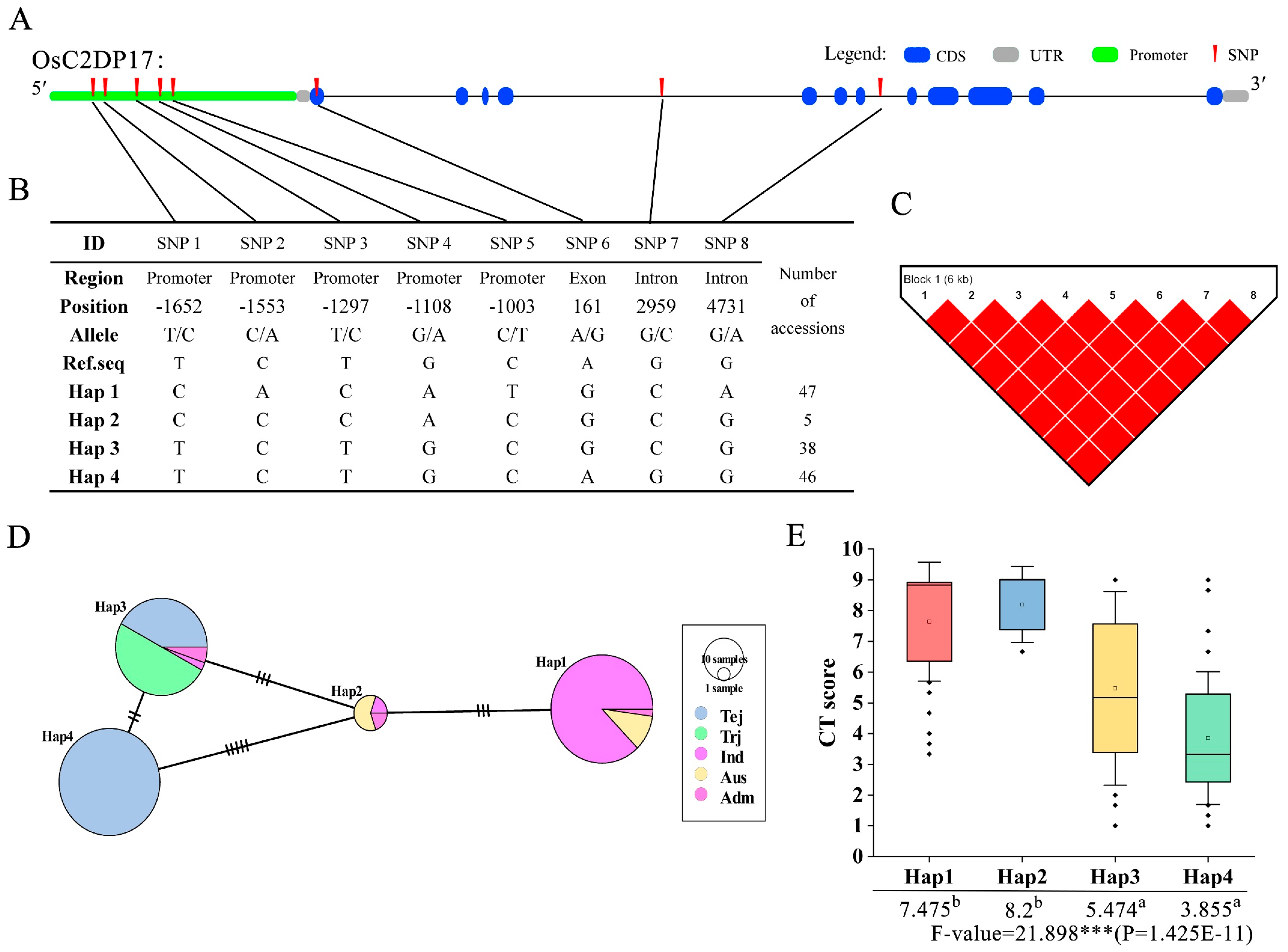
2.7. SNP Genotyping and Haplotype Analysis of OsC2DPs
3. Discussion
3.1. Identification and Phylogenetic Relationship of OsC2DPs in Rice Genome
3.2. Duplications Events Performed in OsC2DPs
3.3. Elements Prediction and Expression Analysis of OsC2DPs
3.4. Identification of Potential Variation Alleles for Future Breeding
4. Materials and Methods
4.1. Identification, Chromosome Distribution, and Localization of OsC2DPs
4.2. Phylogenetic, Structure and Multiple Sequence Alignment Analyses
4.3. Synteny and Ka/Ks Analysis
4.4. Expression Profile Analysis and Prediction of cis-Acting Elements
4.5. Plant Growth, Treatment, and Quantitative Real-Time PCR Analysis
4.6. miRNA Target Prediction and Enrichment Analysis of OsC2DPs
4.7. SNP Genotyping and Haplotype Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kirkby, E.; Pilbeam, D. Calcium as a plant nutrient. Plant Cell Environ. 1984, 7, 397–405. [Google Scholar] [CrossRef]
- Du, L.; Ali, G.S.; Simons, K.A.; Hou, J.; Yang, T.; Reddy, A.; Poovaiah, B. Ca2+/calmodulin regulates salicylic-acid-mediated plant immunity. Nature 2009, 457, 1154–1158. [Google Scholar] [CrossRef] [PubMed]
- Kudla, J.; Batistič, O.; Hashimoto, K. Calcium signals: The lead currency of plant information processing. Plant Cell 2010, 22, 541–563. [Google Scholar] [CrossRef] [PubMed]
- Sanders, D.; Brownlee, C.; Harper, J.F. Communicating with calcium. Plant Cell 1999, 11, 691–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, L.; Schumaker, K.S.; Zhu, J.-K. Cell signaling during cold, drought, and salt stress. Plant Cell 2002, 14, S165–S183. [Google Scholar] [CrossRef] [Green Version]
- Luan, S.; Kudla, J.; Rodriguez-Concepcion, M.; Yalovsky, S.; Gruissem, W. Calmodulins and calcineurin B–like proteins: Calcium sensors for specific signal response coupling in plants. Plant Cell 2002, 14, S389–S400. [Google Scholar] [CrossRef] [Green Version]
- Plant, P.J.; Yeger, H.; Staub, O.; Howard, P.; Rotin, D. The C2 Domain of the Ubiquitin Protein Ligase Nedd4 Mediates Ca2+-dependent Plasma Membrane Localization. J. Biol. Chem. 1997, 272, 32329–32336. [Google Scholar] [CrossRef] [Green Version]
- Farah, C.A.; Sossin, W.S. The role of C2 domains in PKC signaling. Calcium Signal. 2012, 740, 663–683. [Google Scholar]
- Kopka, J.; Pical, C.; Hetherington, A.M.; Müller-Röber, B. Ca2+/phospholipid-binding (C2) domain in multiple plant proteins: Novel components of the calcium-sensing apparatus. Plant Mol. Biol. 1998, 36, 627–637. [Google Scholar] [CrossRef]
- Merithew, E.; Lambright, D.G. Calculating the potential of C2 domains for membrane binding. Dev. Cell 2002, 2, 132–133. [Google Scholar] [CrossRef] [Green Version]
- Schapire, A.L.; Voigt, B.; Jasik, J.; Rosado, A.; Lopez-Cobollo, R.; Menzel, D.; Salinas, J.; Mancuso, S.; Valpuesta, V.; Baluska, F. Arabidopsis synaptotagmin 1 is required for the maintenance of plasma membrane integrity and cell viability. Plant Cell 2008, 20, 3374–3388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molz, L.; Chen, Y.-W.; Hirano, M.; Williams, L.T. Cpk is a novel class of Drosophila PtdIns 3-kinase containing a C2 domain. J. Biol. Chem. 1996, 271, 13892–13899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, I.; Kohara, Y.; Yamamoto, M.; Sugimoto, A. Large-scale analysis of gene function in Caenorhabditis elegans by high-throughput RNAi. Curr. Biol. 2001, 11, 171–176. [Google Scholar] [CrossRef] [Green Version]
- Cho, W.; Stahelin, R.V. Membrane binding and subcellular targeting of C2 domains. Biochim. Et Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2006, 1761, 838–849. [Google Scholar] [CrossRef]
- Nalefski, E.A.; Falke, J.J. The C2 domain calcium-binding motif: Structural and functional diversity. Protein Sci. 1996, 5, 2375–2390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, X.; Gao, Y.-G.; Boldyrev, I.A.; Malinina, L.; Patel, D.J.; Molotkovsky, J.G.; Chalfant, C.A.; Brown, R.E. Regulation of Membrane Binding by the C2-Domain of Cytoplasmic Phospholipase A2 by Ceramide-1-Phosphate and Calcium. Biophys. J. 2017, 112, 392a–393a. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Jiang, S.; Mitok, K.A.; Li, L.; Attie, A.D.; Martin, T.F. BAIAP3, a C2 domain–containing Munc13 protein, controls the fate of dense-core vesicles in neuroendocrine cells. J. Cell Biol. 2017, 216, 2151–2166. [Google Scholar] [CrossRef] [PubMed]
- Souroujon, M.C.; Mochly-Rosen, D. Peptide modulators of protein–protein interactions in intracellular signaling. Nat. Biotechnol. 1998, 16, 919–924. [Google Scholar] [CrossRef]
- Brandman, R.; Disatnik, M.-H.; Churchill, E.; Mochly-Rosen, D. Peptides derived from the C2 domain of protein kinase C epsilon (epsilon PKC) modulate epsilon PKC activity and identify potential protein-protein interaction surfaces. J. Biol. Chem. 2006, 282, 4113–4123. [Google Scholar] [CrossRef] [Green Version]
- Rahier, R.; Noiriel, A.; Abousalham, A. Functional characterization of the N-Terminal C2 domain from Arabidopsis thaliana phospholipase Dα and Dβ. BioMed Res. Int. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Premkumar, A.; Lindberg, S.; Lager, I.; Rasmussen, U.; Schulz, A. Arabidopsis PLDs with C2-domain function distinctively in hypoxia. Physiol. Plant 2019, 167, 90–110. [Google Scholar] [CrossRef] [PubMed]
- Cheung, M.-Y.; Ngo, J.C.-K.; Chen, Z.; Jia, Q.; Li, T.; Gou, Y.; Wang, Y.; Lam, H.-M. A structure model explaining the binding between a ubiquitous unconventional G-protein (OsYchF1) and a plant-specific C2-domain protein (OsGAP1) from rice. Biochem. J. 2020, 477, 3935–3949. [Google Scholar] [CrossRef] [PubMed]
- Vaddepalli, P.; Herrmann, A.; Fulton, L.; Oelschner, M.; Hillmer, S.; Stratil, T.F.; Fastner, A.; Hammes, U.Z.; Ott, T.; Robinson, D.G. The C2-domain protein QUIRKY and the receptor-like kinase STRUBBELIG localize to plasmodesmata and mediate tissue morphogenesis in Arabidopsis thaliana. Development 2014, 141, 4139–4148. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.Y.; Koo, Y.D.; Jin, J.B.; Moon, B.C.; Kang, C.H.; Kim, S.T.; Park, B.O.; Lee, S.Y.; Kim, M.L.; Hwang, I. Rice C2-domain proteins are induced and translocated to the plasma membrane in response to a fungal elicitor. Biochemistry 2003, 42, 11625–11633. [Google Scholar] [CrossRef] [PubMed]
- Yokotani, N.; Ichikawa, T.; Kondou, Y.; Maeda, S.; Iwabuchi, M.; Mori, M.; Hirochika, H.; Matsui, M.; Oda, K. Overexpression of a rice gene encoding a small C2 domain protein OsSMCP1 increases tolerance to abiotic and biotic stresses in transgenic Arabidopsis. Plant Mol. Biol. 2009, 71, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Fu, L.; Zhang, X.; Huang, J.; Yang, G.; Wang, Z.; Liu, Y.-G.; Zhang, G.; Wu, D.; Xia, J. OsC2DP, a novel C2 domain-containing protein is required for salt tolerance in rice. Plant Cell Physiol. 2019, 60, 2220–2230. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, C.; Liang, Z.; Yu, H. Characterization of multiple C2 domain and transmembrane region proteins in Arabidopsis. Plant Physiol. 2018, 176, 2119–2132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, P.; Wang, H.; Ma, L.; Wu, A.; Chen, P.; Cheng, S.; Wei, H.; Yu, S. Genome-wide identification and characterization of multiple C2 domains and transmembrane region proteins in Gossypium hirsutum. BMC Genom. 2020, 21, 445. [Google Scholar] [CrossRef]
- Huang, R.; Zhao, J.; Liu, J.; Wang, Y.; Han, S.; Zhao, H. Genome-wide analysis and expression profiles of NTMC2 family genes in Oryza sativa. Gene 2017, 637, 130–137. [Google Scholar] [CrossRef]
- Hammoudi, V.; Vlachakis, G.; Schranz, M.E.; van den Burg, H.A. Whole-genome duplications followed by tandem duplications drive diversification of the protein modifier SUMO in Angiosperms. New Phytol. 2016, 211, 172–185. [Google Scholar] [CrossRef] [Green Version]
- Chow, C.-N.; Chiang-Hsieh, Y.-F.; Chien, C.-H.; Zheng, H.-Q.; Lee, T.-Y.; Wu, N.-Y.; Tseng, K.-C.; Hou, P.-F.; Chang, W.-C. Delineation of condition specific Cis-and Trans-acting elements in plant promoters under various Endo-and exogenous stimuli. BMC Genom. 2018, 19, 109–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Song, S.; Xiao, Y.; Fan, F.; Zhou, Y.; Jia, G.; Tang, W.; Peng, J. Circadian clock-coordinated response to chilling stress in rice. Environ. Exp. Bot. 2021, 185, 104398. [Google Scholar] [CrossRef]
- Zheng, L.; Zhang, C.; Shi, C.; Wang, Y.; Zhou, T.; Sun, F.; Wang, H.; Zhao, S.; Qin, Q.; Qiao, R. Rice stripe virus NS3 protein regulates primary miRNA processing through association with the miRNA biogenesis factor OsDRB1 and facilitates virus infection in rice. PLoS Pathog. 2017, 13, e1006662. [Google Scholar] [CrossRef] [Green Version]
- Archak, S.; Nagaraju, J. Computational prediction of rice (Oryza sativa) miRNA targets. Genom. Proteom. Bioinform. 2007, 5, 196–206. [Google Scholar] [CrossRef] [Green Version]
- Nalefski, E.A.; Wisner, M.A.; Chen, J.Z.; Sprang, S.R.; Fukuda, M.; Mikoshiba, K.; Falke, J.J. C2 domains from different Ca2+ signaling pathways display functional and mechanistic diversity. Biochemistry 2001, 40, 3089–3100. [Google Scholar] [CrossRef] [PubMed]
- Leister, D. Tandem and segmental gene duplication and recombination in the evolution of plant disease resistance genes. Trends Genet. 2004, 20, 116–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Wu, Y.; Liu, Y.; Yang, L.; Dong, R.; Jiang, L.; Liu, P.; Liu, G.; Wang, Z.; Luo, L. Genome-wide analysis of tandem duplicated genes and their contribution to stress resistance in pigeonpea (Cajanus cajan). Genomics 2021, 113, 728–735. [Google Scholar] [CrossRef]
- Yu, J.; Ke, T.; Tehrim, S.; Sun, F.; Liao, B.; Hua, W. PTGBase: An integrated database to study tandem duplicated genes in plants. Database 2015, 2015. [Google Scholar] [CrossRef] [Green Version]
- Clark, R.M.; Schweikert, G.; Toomajian, C.; Ossowski, S.; Zeller, G.; Shinn, P.; Warthmann, N.; Hu, T.T.; Fu, G.; Hinds, D.A. Common sequence polymorphisms shaping genetic diversity in Arabidopsis thaliana. Science 2007, 317, 338–342. [Google Scholar] [CrossRef] [Green Version]
- Rizzon, C.; Ponger, L.; Gaut, B.S. Striking similarities in the genomic distribution of tandemly arrayed genes in Arabidopsis and rice. PLoS Comput. Biol. 2006, 2, e115. [Google Scholar] [CrossRef]
- Bellieny-Rabelo, D.; Oliveira, A.E.A.; Venancio, T.M. Impact of whole-genome and tandem duplications in the expansion and functional diversification of the F-box family in legumes (Fabaceae). PLoS ONE 2013, 8, e55127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.-Q.; Lai, Y.; Li, M.-N.; Xu, W.-Y.; Xue, Y.-B. A novel C2-domain phospholipid-binding protein, OsPBP1, is required for pollen fertility in rice. Mol. Plant 2008, 1, 770–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, S.Y.; Cai, M.; Ramachandran, S. The Oryza sativa no pollen (Osnop) gene plays a role in male gametophyte development and most likely encodes a C2-GRAM domain-containing protein. Plant Mol. Biol. 2005, 57, 835–853. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Ou, S.; Mao, B.; Tang, J.; Wang, W.; Wang, H.; Cao, S.; Schläppi, M.R.; Zhao, B.; Xiao, G. Early selection of bZIP73 facilitated adaptation of japonica rice to cold climates. Nat. Commun. 2018, 9, 3302. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Schläppi, M.R.; Mao, B.; Wang, W.; Wang, A.; Chu, C. The bZIP 73 transcription factor controls rice cold tolerance at the reproductive stage. Plant Biotechnol. J. 2019, 17, 1834–1849. [Google Scholar] [CrossRef] [Green Version]
- Fjell, C.D.; Jenssen, H.; Fries, P.; Aich, P.; Griebel, P.; Hilpert, K.; Hancock, R.E.; Cherkasov, A. Identification of novel host defense peptides and the absence of α-defensins in the bovine genome. Proteins Struct. Funct. Bioinf. 2008, 73, 420–430. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stothard, P. The sequence manipulation suite: JavaScript programs for analyzing and formatting protein and DNA sequences. Biotechniques 2000, 28, 1102–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-h.; Jin, H.; Marler, B.; Guo, H. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Yang, G.; Yan, J.; Pan, Y.; Nie, X. Genome-wide identification, expression profiles and regulatory network of MAPK cascade gene family in barley. BMC Genom. 2019, 20, 750. [Google Scholar] [CrossRef] [Green Version]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Zhang, Y.; Zhang, Z.; Zhu, J.; Yu, J. KaKs_Calculator 2.0: A toolkit incorporating gamma-series methods and sliding window strategies. Genom. Proteom. Bioinform. 2010, 8, 77–80. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Xie, W.; Chen, Y.; Tang, W.; Yang, J.; Ye, R.; Liu, L.; Lin, Y.; Xu, C.; Xiao, J. A dynamic gene expression atlas covering the entire life cycle of rice. Plant J. 2010, 61, 752–766. [Google Scholar] [CrossRef]
- Garg, R.; Tyagi, A.K.; Jain, M. Microarray analysis reveals overlapping and specific transcriptional responses to different plant hormones in rice. Plant Signal. Behav. 2012, 7, 951–956. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Takehisa, H.; Kamatsuki, K.; Minami, H.; Namiki, N.; Ikawa, H.; Ohyanagi, H.; Sugimoto, K.; Antonio, B.A.; Nagamura, Y. RiceXPro version 3.0: Expanding the informatics resource for rice transcriptome. Nucleic Acids Res. 2013, 41, D1206–D1213. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2−ΔΔCT method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinform. Biomath. 2013, 3, 71. [Google Scholar]
- Dai, X.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server. Nucleic Acids Res. 2011, 39, W155–W159. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Yi, X.; Du, Z.; Su, Z. PlantGSEA: A gene set enrichment analysis toolkit for plant community. Nucleic Acids Res. 2013, 41, W98–W103. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.-S.; He, Q.; Kim, K.-W.; Yoon, M.-Y.; Ra, W.-H.; Li, F.P.; Tong, W.; Yu, J.; Oo, W.H.; Choi, B. Genome-wide resequencing of KRICE_CORE reveals their potential for future breeding, as well as functional and evolutionary studies in the post-genomic era. BMC Genom. 2016, 17, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; San, M.L.; Jang, S.-G.; Lee, J.-H.; Kim, N.-E.; Lee, A.-R.; Park, S.-Y.; Cao, F.-Y.; Chin, J.-H.; Kwon, S.-W. Genome-Wide Association Study of Root System Development at Seedling Stage in Rice. Genes 2020, 11, 1395. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wu, T.; Li, Z.; Huang, K.; Kim, N.-E.; Ma, Z.; Kwon, S.-W.; Jiang, W.; Du, X. OsGATA16, a GATA Transcription Factor, Confers Cold Tolerance by Repressing OsWRKY45–1 at the Seedling Stage in Rice. Rice 2021, 14, 42. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. popart: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tandem Duplication | Ka | Ks | Ka/Ks | S | N | Effective Len | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene Symbol | Gene ID | Chr. | Start | End | Gene Symbol | Gene ID | Chr. | Start | End | ||||||
| OsC2DP6 | LOC_Os01g60340 | Chr1 | 34,902,303 | 34,906,955 | OsC2DP7 | LOC_Os01g60350 | Chr1 | 34,908,079 | 34,908,079 | 0.15904 | 0.34162 | 0.46554 | 326.167 | 1161.83 | 1488 |
| OsC2DP50 | LOC_Os06g40170 | Chr6 | 23,908,918 | 23,913,797 | OsC2DP51 | LOC_Os06g40180 | Chr6 | 23,921,940 | 23,921,940 | 0.09049 | 0.14449 | 0.62625 | 590.333 | 1893.67 | 2484 |
| OsC2DP51 | LOC_Os06g40180 | Chr6 | 23,921,940 | 23,924,921 | OsC2DP52 | LOC_Os06g40190 | Chr6 | 23,928,702 | 23,928,702 | 0.16077 | 0.48728 | 0.32993 | 590.667 | 1854.33 | 2445 |
| OsC2DP50 | LOC_Os06g40170 | Chr6 | 23,908,918 | 23,913,797 | OsC2DP52 | LOC_Os06g40190 | Chr6 | 23,928,702 | 23,928,702 | 0.16616 | 0.45379 | 0.36617 | 588.167 | 1856.83 | 2445 |
| OsC2DP58 | LOC_Os07g01770 | Chr7 | 452,093 | 460,151 | OsC2DP59 | LOC_Os07g01780 | Chr7 | 457,738 | 457,738 | 0.00405 | 0.00448 | 0.90514 | 111.975 | 371.025 | 483 |
| OsC2DP67 | LOC_Os07g47390 | Chr7 | 28,339,687 | 28,340,470 | OsC2DP68 | LOC_Os07g47400 | Chr7 | 28,344,133 | 28,344,133 | 0.40704 | 0.60314 | 0.67487 | 190.833 | 493.167 | 684 |
| Segmental Duplication | Ka | Ks | Ka/Ks | S | N | Effective Len | |||||||||
| Gene Symbol | Gene ID | Chr. | Start | End | Gene Symbol | Gene ID | Chr. | Start | End | ||||||
| OsC2DP11 | LOC_Os01g72230 | Chr1 | 41,882,488 | 41,886,410 | OsC2DP46 | LOC_Os05g31720 | Chr5 | 18,466,963 | 18,470,649 | 0.26074 | 1.11476 | 0.2339 | 246.833 | 875.167 | 1122 |
| OsC2DP2 | LOC_Os01g07760 | Chr1 | 3,724,314 | 3,729,284 | OsC2DP43 | LOC_Os05g07880 | Chr5 | 4,255,763 | 4,260,333 | 0.11203 | 5.56155 | 0.02014 | 396.932 | 2039.07 | 2436 |
| OsC2DP10 | LOC_Os01g70790 | Chr1 | 40,973,255 | 40,974,782 | OsC2DP67 | LOC_Os07g47390 | Chr7 | 28,339,687 | 28,340,470 | 0.40741 | 0.63562 | 0.64096 | 190.917 | 484.083 | 675 |
| OsC2DP20 | LOC_Os02g42710 | Chr2 | 25,684,044 | 25,687,149 | OsC2DP36 | LOC_Os04g44870 | Chr4 | 26,558,810 | 26,561,326 | 0.1123 | 0.57604 | 0.19495 | 97 | 326 | 423 |
| OsC2DP13 | LOC_Os02g10480 | Chr2 | 5,505,016 | 5,508,425 | OsC2DP54 | LOC_Os06g40704 | Chr6 | 24,264,404 | 24,268,015 | 0.15437 | 0.84849 | 0.18193 | 217.5 | 727.5 | 945 |
| OsC2DP14 | LOC_Os02g10630 | Chr2 | 5,586,939 | 5,593,645 | OsC2DP53 | LOC_Os06g40570 | Chr6 | 24,187,249 | 24,192,151 | 0.22964 | 0.83077 | 0.27643 | 664.167 | 2266.83 | 2931 |
| OsC2DP30 | LOC_Os03g18010 | Chr3 | 10,025,401 | 10,027,931 | OsC2DP69 | LOC_Os07g49330 | Chr7 | 29,545,825 | 29,548,981 | 0.17977 | 1.14005 | 0.15769 | 325.667 | 1108.33 | 1434 |
| OsC2DP72 | LOC_Os08g44850 | Chr8 | 28,170,358 | 28,172,330 | OsC2DP78 | LOC_Os09g39770 | Chr9 | 22,805,610 | 22,807,049 | 0.16323 | 0.41331 | 0.39494 | 231.833 | 602.167 | 834 |
| Variation Type | Count | Variation in Exon | Count |
|---|---|---|---|
| Promoter variation | 424 | ||
| Upstream variation | 575 | ||
| Downstream variation | 73 | ||
| Upstream and downstream variation | 265 | ||
| 5’UTR variation | 47 | ||
| 3’UTR variation | 172 | ||
| Intergenic variation | 178 | ||
| Exonic variation | 223 | Synonymous variant | 124 |
| Non-synonymous variant | 99 | ||
| Intronic variation | 829 | ||
| ncRNA_exonic variation | 75 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Zeng, Y.; Seo, J.; Kim, Y.-J.; Kim, S.T.; Kwon, S.-W. Global Identification and Characterization of C2 Domain-Containing Proteins Associated with Abiotic Stress Response in Rice (Oryza sativa L.). Int. J. Mol. Sci. 2022, 23, 2221. https://doi.org/10.3390/ijms23042221
Zhang H, Zeng Y, Seo J, Kim Y-J, Kim ST, Kwon S-W. Global Identification and Characterization of C2 Domain-Containing Proteins Associated with Abiotic Stress Response in Rice (Oryza sativa L.). International Journal of Molecular Sciences. 2022; 23(4):2221. https://doi.org/10.3390/ijms23042221
Chicago/Turabian StyleZhang, Hongjia, Yuting Zeng, Jeonghwan Seo, Yu-Jin Kim, Sun Tae Kim, and Soon-Wook Kwon. 2022. "Global Identification and Characterization of C2 Domain-Containing Proteins Associated with Abiotic Stress Response in Rice (Oryza sativa L.)" International Journal of Molecular Sciences 23, no. 4: 2221. https://doi.org/10.3390/ijms23042221
APA StyleZhang, H., Zeng, Y., Seo, J., Kim, Y.-J., Kim, S. T., & Kwon, S.-W. (2022). Global Identification and Characterization of C2 Domain-Containing Proteins Associated with Abiotic Stress Response in Rice (Oryza sativa L.). International Journal of Molecular Sciences, 23(4), 2221. https://doi.org/10.3390/ijms23042221