
MicroRNA-199b Deregulation Shows Oncogenic Properties and Promising Clinical Value as Circulating Marker in Locally Advanced Rectal Cancer Patients

 , , ,
, , , 
Abstract
:1. Introduction
2. Results
2.1. MiR-199b Regulates Cell Migration and Agressiveness
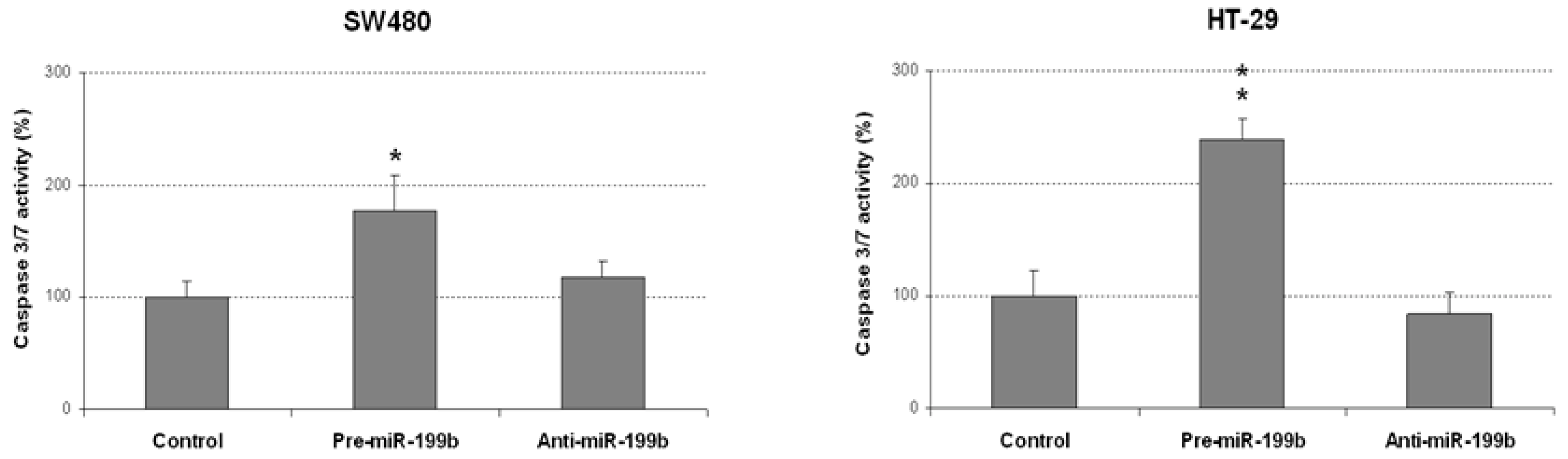
2.2. MiR-199b Induces Apoptosis and Inhibts Colonosphere Formation
2.3. Analysis of miR-199b Expression Levels as Circulating Markers in Larc Patients and Its Association with Molecular and Clinical Parameters
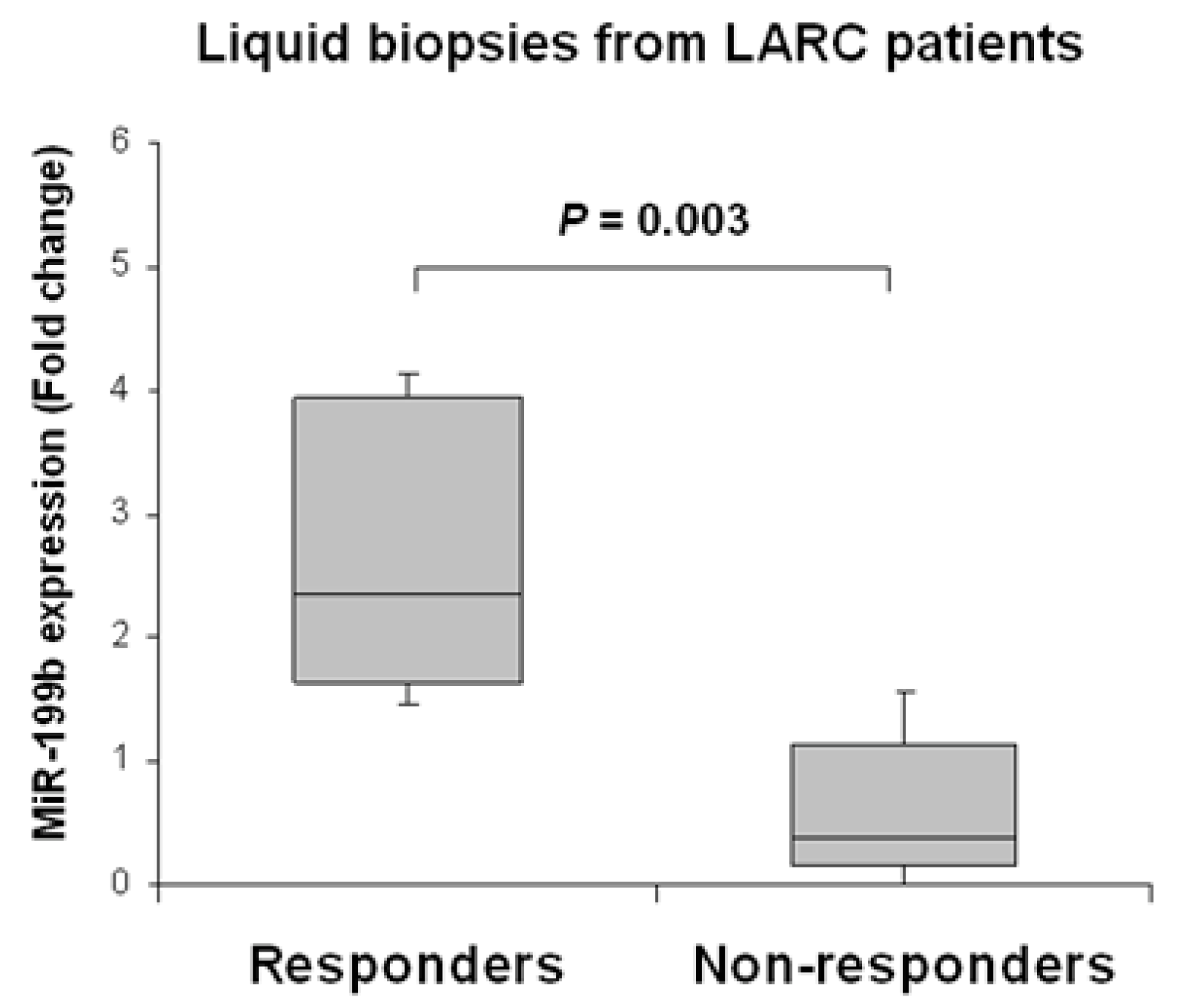
2.4. Circulating miR-199b Expression Levels Predict Pathological Response to nCRT and Recurrence in LARC Patients
3. Discussion
4. Materials and Methods
4.1. Cell Cultures and Transfection
4.2. Patient Samples
4.3. Evaluation of Pathological Response
4.4. Wound-Healing Assay
4.5. Transwell Migration Assay
4.6. Colony-Forming Assay
4.7. Cell Viability Assay
4.8. Analysis of Caspase Activation
4.9. Colonosphere Formation Assay
4.10. Western Blot Analysis
4.11. RNA Isolation
4.12. Quantification of miR Expression Levels
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rawla, P.; Sunkara, T.; Barsouk, A. Epidemiology of colorectal cancer: Incidence, mortality, survival, and risk factors. Prz. Gastroenterol. 2019, 14, 89–103. [Google Scholar] [CrossRef]
- Global Cancer Observatory. Colorectal Cancer. Available online: https://gco.iarc.fr/today/data/factsheets/cancers/10_8_9-Colorectum-fact-sheet.pdf (accessed on 2 November 2021).
- Spanish Society of Medical Oncology. Cancer Data in Spain. Available online: https://seom.org/images/Cifras_del_cancer_en_Espnaha_2021.pdf (accessed on 31 October 2021).
- Sanghera, P.; Wong, D.W.; McConkey, C.C.; Geh, J.I.; Hartley, A. Chemoradiotherapy for rectal cancer: An updated analysis of factors affecting pathological response. Clin. Oncol. (R. Coll. Radiol.) 2008, 20, 176–183. [Google Scholar] [CrossRef]
- Dayde, D.; Gunther, J.; Hirayama, Y.; Weksberg, D.C.; Boutin, A.; Parhy, G.; Aguilar-Bonavides, C.; Wang, H.; Katayama, H.; Abe, Y.; et al. Identification of blood-based biomarkers for the prediction of the response to neoadjuvant chemoradiation in rectal cancer. Cancers 2021, 13, 3642. [Google Scholar] [CrossRef]
- São Julião, G.P.; Habr-Gama, A.; Vailati, B.B.; Araujo, S.E.A.; Fernandez, L.M.; Perez, R.O. New strategies in rectal cancer. Surg. Clin. N. Am. 2017, 97, 587–604. [Google Scholar] [CrossRef]
- Papaccio, F.; Roselló, S.; Huerta, M.; Gambardella, V.; Tarazona, N.; Fleitas, T.; Roda, D.; Cervantes, A. Neoadjuvant chemotherapy in locally advanced rectal cancer. Cancers 2020, 12, 3611. [Google Scholar] [CrossRef]
- Bahadoer, R.R.; Dijkstra, E.A.; van Etten, B.; Marijnen, C.A.M.; Putter, H.; Kranenbarg, E.M.-K.; Roodvoets, A.G.H.; Nagtegaal, I.D.; Beets-Tan, R.G.H.; Blomqvist, L.K.; et al. Short-course radiotherapy followed by chemotherapy before total mesorectal excision (TME) versus preoperative chemoradiotherapy, TME, and optional adjuvant chemotherapy in locally advanced rectal cancer (RAPIDO): A randomised, open-label, phase 3 trial. Lancet Oncol. 2021, 22, 29–42. [Google Scholar] [CrossRef]
- Conroy, T.; Bosset, J.-F.; Etienne, P.-L.; Rio, E.; François, É.; Mesgouez-Nebout, N.; Vendrely, V.; Artignan, X.; Bouché, O.; Gargot, D.; et al. Neoadjuvant chemotherapy with FOLFIRINOX and preoperative chemoradiotherapy for patients with locally advanced rectal cancer (UNICANCER-PRODIGE 23): A multicentre, randomised, open-label, phase 3 trial. Lancet Oncol. 2021, 22, 702–715. [Google Scholar] [CrossRef]
- Norcic, G. Liquid biopsy in colorectal cancer-current status and potential clinical applications. Micromachines 2018, 9, 300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vacante, M.; Borzì, A.M.; Basile, F.; Biondi, A. Biomarkers in colorectal cancer: Current clinical utility and future perspectives. World J. Clin. Cases 2018, 6, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Palmirotta, R.; Lovero, D.; Cafforio, P.; Felici, C.; Mannavola, F.; Pellè, E.; Quaresmini, D.; Tucci, M.; Silvestris, F. Liquid biopsy of cancer: A multimodal diagnostic tool in clinical oncology. Ther. Adv. Med. Oncol. 2018, 10, 1758835918794630. [Google Scholar] [CrossRef] [PubMed]
- Malentacchi, F.; Sgromo, C.; Antonuzzo, L.; Pillozzi, S. Liquid biopsy in endometrial cancer. J. Cancer Metastasis Treat. 2020, 6, 34. [Google Scholar] [CrossRef]
- Quirico, L.; Orso, F. The power of microRNAs as diagnostic and prognostic biomarkers in liquid biopsies. Cancer Drug Resist. 2020, 3, 117–139. [Google Scholar] [CrossRef] [Green Version]
- Vacante, M.; Ciuni, R.; Basile, F.; Biondi, A. The liquid biopsy in the management of colorectal cancer: An overview. Biomedicines 2020, 8, 308. [Google Scholar] [CrossRef]
- Shigeyasu, K.; Toden, S.; Zumwalt, T.J.; Okugawa, Y.; Goel, A. Emerging role of MicroRNAs as liquid biopsy biomarkers in gastrointestinal cancers. Clin. Cancer Res. 2017, 23, 2391–2399. [Google Scholar] [CrossRef] [Green Version]
- De Palma, F.D.E.; Luglio, G.; Tropeano, F.P.; Pagano, G.; D’Armiento, M.; Kroemer, G.; Maiuri, M.C.; De Palma, G.D. The role of micro-RNAs and circulating tumor markers as predictors of response to neoadjuvant therapy in locally advanced rectal cancer. Int. J. Mol. Sci. 2020, 21, 7040. [Google Scholar] [CrossRef]
- Peng, Y.; Croce, C.M. The role of MicroRNAs in human cancer. Signal Transduct. Target Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.W.; Ferland-McCollough, D.; Jackson, T.J.; Bushell, M. MicroRNAs in cancer management. Lancet Oncol. 2012, 13, e249–e258. [Google Scholar] [CrossRef]
- Imedio, L.; Cristóbal, I.; Rubio, J.; Santos, A.; Rojo, F.; García-Foncillas, J. MicroRNAs in rectal cancer: Functional significance and promising therapeutic value. Cancers 2020, 12, 2040. [Google Scholar] [CrossRef]
- Iorio, M.V.; Ferracin, M.; Liu, C.G.; Veronese, A.; Spizzo, R.; Sabbioni, S.; Magri, E.; Pedriali, M.; Fabbri, M.; Campiglio, M.; et al. MicroRNA gene expression deregulation in human breast cancer. Cancer Res. 2005, 65, 7065–7070. [Google Scholar] [CrossRef] [Green Version]
- Calin, G.A.; Ferracin, M.; Cimmino, A.; Di Leva, G.; Shimizu, M.; Wojcik, S.E.; Iorio, M.V.; Visone, R.; Sever, N.I.; Fabbri, M.; et al. A microRNA signature associated with prognosis and progression in chronic lymphocytic leukemia. N. Engl. J. Med. 2005, 353, 1793–1801. [Google Scholar] [CrossRef]
- Qi, J.; Wang, J.; Katayama, H.; Sen, S.; Liu, S.M. Circulating microRNAs (cmiRNAs) as novel potential biomarkers for hepatocellular carcinoma. Neoplasma 2013, 60, 135–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roldo, C.; Missiaglia, E.; Hagan, J.P.; Falconi, M.; Capelli, P.; Bersani, S.; Calin, G.A.; Volinia, S.; Liu, C.G.; Scarpa, A.; et al. MicroRNA expression abnormalities in pancreatic endocrine and acinar tumors are associated with distinctive pathological features and clinical behavior. J. Clin. Oncol. 2006, 24, 4677–4684. [Google Scholar] [CrossRef] [PubMed]
- Porkka, K.P.; Pfeiffer, M.J.; Waltering, K.K.; Vessella, R.L.; Tammela, T.L.J.; Visakorpi, T. MicroRNA expression profiling in prostate cancer. Cancer Res. 2007, 67, 6130–6135. [Google Scholar] [CrossRef] [Green Version]
- Ciafrè, S.A.; Galardi, S.; Mangiola, A.; Ferracin, M.; Liu, C.G.; Sabatino, G.; Negrini, M.; Maira, G.; Croce, C.M.; Farace, M.G. Extensive modulation of a set of microRNAs in primary glioblastoma. Biochem. Biophys. Res. Commun. 2005, 334, 1351–1358. [Google Scholar] [CrossRef]
- Caramés, C.; Cristóbal, I.; Moreno, V.; Del Puerto, L.; Moreno, I.; Rodriguez, M.; Marín, J.P.; Correa, A.V.; Hernández, R.; Zenzola, V.; et al. MicroRNA-21 predicts response to preoperative chemoradiotherapy in locally advanced rectal cancer. Int. J. Colorectal Dis. 2015, 30, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Caramés, C.; Cristobal, I.; Moreno, V.; Marín, J.P.; González-Alonso, P.; Torrejón, B.; Minguez, P.; Leon, A.; Martín, J.I.; Hernández, R.; et al. MicroRNA-31 emerges as a predictive biomarker of pathological response and outcome in locally advanced rectal cancer. Int. J. Mol. Sci. 2016, 17, 878. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Peng, Q.; Lin, Y.; Zou, L.; Shen, P.; Chen, F.; Min, M.; Shen, L.; Chen, J.; Shen, B. Identification of biomarker microRNAs for predicting the response of colorectal cancer to neoadjuvant chemoradiotherapy based on microRNA regulatory network. Oncotarget 2016, 8, 2233–2248. [Google Scholar] [CrossRef] [Green Version]
- Roman-Canal, B.; Tarragona, J.; Moiola, C.P.; Gatius, S.; Bonnin, S.; Ruiz-Miró, M.; Sierra, J.E.; Rufas, M.; González, E.; Porcel, J.M.; et al. EV-associated miRNAs from peritoneal lavage as potential diagnostic biomarkers in colorectal cancer. J. Transl. Med. 2019, 17, 208. [Google Scholar] [CrossRef] [Green Version]
- Li, G.L.; Yuan, J.H.; Zhuang, G.D.; Wu, D.Q. MiR-199b exerts tumor suppressive functions in hepatocellular carcinoma by directly targeting JAG1. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7679–7687. [Google Scholar] [CrossRef]
- Qu, X.; Chen, Z.; Fan, D.; Sun, C.; Zeng, Y.; Guo, Z.; Qi, Q.; Li, W. MiR-199b-5p inhibits osteogenic differentiation in ligamentum flavum cells by targeting JAG1 and modulating the Notch signalling pathway. J. Cell Mol. Med. 2017, 21, 1159–1170. [Google Scholar] [CrossRef]
- Fan, X.; Matsui, W.; Khaki, L.; Stearns, D.; Chun, J.; Li, Y.M.; Eberhart, C.G. Notch pathway inhibition depletes stem-like cells and blocks engraftment in embryonal brain tumors. Cancer Res. 2006, 66, 7445–7452. [Google Scholar] [CrossRef] [Green Version]
- Garzia, L.; Andolfo, I.; Cusanelli, E.; Marino, N.; Petrosino, G.; De Martino, D.; Esposito, V.; Galeone, A.; Navas, L.; Esposito, S.; et al. MicroRNA-199b-5p impairs cancer stem cells through negative regulation of HES1 in medulloblastoma. PLoS ONE 2009, 4, e4998. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhao, S.; Luo, L.; Xiang, Q.; Zhu, Z.; Wang, J.; Liu, Y.; Luo, J. MiR-199b-5p-DDR1-ERK signalling axis suppresses prostate cancer metastasis via inhibiting epithelial-mesenchymal transition. Br. J. Cancer 2021, 124, 982–994. [Google Scholar] [CrossRef]
- Wu, A.; Chen, Y.; Liu, Y.; Lai, Y.; Liu, D. MiR-199b-5p inhibits triple negative breast cancer cell proliferation, migration and invasion by targeting DDR1. Oncol. Lett. 2018, 16, 4889–4896. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Qiu, W.; Xiao, Y.; Ma, J.; Xu, F.; Zhang, K.; Gao, Y.; Chen, Q.; Li, Y.; Li, H.; et al. MiR-199b-5p Suppresses tumor angiogenesis mediated by vascular endothelial cells in breast cancer by targeting ALK1. Front. Genet. 2020, 10, 1397. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.Y.; Zhi, Z.; Wang, L.; Zhao, Y.Y.; Deng, M.; Liu, Y.H.; Qin, Y.; Tian, M.M.; Liu, Y.; Shen, T.; et al. NSD2 circular RNA promotes metastasis of colorectal cancer by targeting miR-199b-5p-mediated DDR1 and JAG1 signalling. J. Pathol. 2019, 248, 103–115. [Google Scholar] [CrossRef]
- Cristóbal, I.; Caramés, C.; Rincón, R.; Manso, R.; Madoz-Gúrpide, J.; Torrejón, B.; González-Alonso, P.; Rojo, F.; García-Foncillas, J. Downregulation of microRNA-199b predicts unfavorable prognosis and emerges as a novel therapeutic target which contributes to PP2A inhibition in metastatic colorectal cancer. Oncotarget 2017, 8, 40169–40180. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Wang, J.H.; Xu, C.; Sun, B.; Kang, S.O. Activin pathway enhances colorectal cancer stem cell self-renew and tumor progression. Biochem. Biophys. Res. Commun. 2016, 479, 715–720. [Google Scholar] [CrossRef]
- Jung, B.; Staudacher, J.J.; Beauchamp, D. Transforming growth factor β superfamily signaling in development of colorectal cancer. Gastroenterology 2017, 152, 36–52. [Google Scholar] [CrossRef] [Green Version]
- Shen, Z.L.; Wang, B.; Jiang, K.W.; Ye, C.X.; Cheng, C.; Yan, Y.C.; Zhang, J.Z.; Yang, Y.; Gao, Z.D.; Ye, Y.J.; et al. Downregulation of miR-199b is associated with distant metastasis in colorectal cancer via activation of SIRT1 and inhibition of CREB/KISS1 signaling. Oncotarget 2016, 7, 35092–35105. [Google Scholar] [CrossRef] [Green Version]
- Cristóbal, I.; Rubio, J.; Torrejón, B.; Santos, A.; Caramés, C.; Luque, M.; Sanz-Álvarez, M.; Alonso, R.; Zazo, S.; Madoz-Gúrpide, J.; et al. MicroRNA-199b deregulation shows a strong SET-independent prognostic value in early-stage colorectal cancer. J. Clin. Med. 2020, 9, 2419. [Google Scholar] [CrossRef] [PubMed]
- Cristóbal, I.; Rubio, J.; Santos, A.; Torrejón, B.; Caramés, C.; Imedio, L.; Mariblanca, S.; Luque, M.; Sanz-Alvarez, M.; Zazo, S.; et al. MicroRNA-199b Downregulation confers resistance to 5-fluorouracil treatment and predicts poor outcome and response to neoadjuvant chemoradiotherapy in locally advanced rectal cancer patients. Cancers 2020, 12, 1655. [Google Scholar] [CrossRef] [PubMed]
- Cristóbal, I.; Santos, A.; Rubio, J.; Caramés, C.; Zazo, S.; Sanz-Álvarez, M.; Luque, M.; Madoz-Gúrpide, J.; Rojo, F.; García-Foncillas, J. Validation of microRNA-199b as A promising predictor of outcome and response to neoadjuvant treatment in locally advanced rectal cancer patients. Cancers 2021, 13, 5003. [Google Scholar] [CrossRef] [PubMed]
- Baek, D.W.; Kim, G.; Kang, B.W.; Kim, H.J.; Park, S.Y.; Park, J.S.; Choi, G.S.; Kang, M.K.; Hur, K.; Kim, J.G. High expression of microRNA-199a-5p is associated with superior clinical outcomes in patients with locally advanced rectal cancer. J. Cancer Res. Clin. Oncol. 2020, 146, 105–115. [Google Scholar] [CrossRef]
- Calin, G.A.; Croce, C.M. MicroRNA signatures in human cancers. Nat. Rev. Cancer 2006, 6, 857–866. [Google Scholar] [CrossRef]
- Suer, I.; Guzel, E.; Karatas, O.F.; Creighton, C.J.; Ittmann, M.; Ozen, M. MicroRNAs as prognostic markers in prostate cancer. Prostate 2019, 79, 265–271. [Google Scholar] [CrossRef]
- Lai, Y.; Quan, J.; Lin, C.; Li, H.; Hu, J.; Chen, P.; Xu, J.; Guan, X.; Xu, W.; Lai, Y.; et al. miR-199b-5p serves as a tumor suppressor in renal cell carcinoma. Exp. Ther. Med. 2018, 16, 436–444. [Google Scholar] [CrossRef]
- Favreau, A.J.; McGlauflin, R.E.; Duarte, C.W.; Sathyanarayana, P. MiR-199b, a novel tumor suppressor miRNA in acute myeloid leukemia with prognostic implications. Exp. Hematol. Oncol. 2016, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Ten Klooster, J.P.; Leeuwen, I.V.; Scheres, N.; Anthony, E.C.; Hordijk, P.L. Rac-1induced cell migration requires membranes recruitment of the nuclear oncogene SET. EMBO J. 2007, 26, 336–345. [Google Scholar] [CrossRef] [Green Version]
- Cristóbal, I.; Torrejón, B.; Rubio, J.; Santos, A.; Pedregal, M.; Caramés, C.; Zazo, S.; Luque, M.; Sanz-Alvarez, M.; Madoz-Gúrpide, J.; et al. Deregulation of SET is associated with tumor progression and predicts adverse outcome in patients with early-stage colorectal cancer. J. Clin. Med. 2019, 8, 346. [Google Scholar] [CrossRef] [Green Version]
- Greenson, J.K.; Huang, S.-C.; Herron, C.; Moreno, V.; Bonner, J.D.; Tomsho, L.P.; Ben-Izhak, O.; Cohen, H.I.; Trougouboff, P.; Bejhar, J.; et al. Pathologic predictors of microsatellite instability in colorectal cancer. Am. J. Surg. Pathol. 2009, 33, 126–133. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-Time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- McShane, L.M.; Altman, D.G.; Sauerbrei, W.; Taube, S.E.; Gion, M.; Clark, G.M. Statistics Subcommittee of the NCI-EORTC Working Group on Cancer Diagnostics. Reporting recommendations for tumor marker prognostic studies. J. Clin. Oncol. 2005, 23, 9067–9072. [Google Scholar] [CrossRef] [Green Version]
- Sauerbrei, W.; Taube, S.E.; McShane, L.M.; Cavenagh, M.M.; Altman, D.G. Reporting Recommendations for Tumor Marker Prognostic Studies (REMARK): An abridged explanation and elaboration. J. Natl. Cancer Inst. 2018, 110, 803–811. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | No. Cases | No. miR-199b High (%) | No. miR-199 Low (%) | p |
|---|---|---|---|---|
| MiR-199b | 22 | 17 (77.3) | 5 (22.7) | |
| Gender | 22 | 17 | 5 | 0.078 |
| Male | 12 | 11 (91.7) | 1 (8.3) | |
| Female | 10 | 6 (60) | 4 (40) | |
| Age | 22 | 17 | 5 | 0.962 |
| <70 | 13 | 10 (76.9) | 3 (23.1) | |
| ≥70 | 9 | 7 (77.8) | 2 (22.2) | |
| Grade pre-CRT 1 | 22 | 17 | 5 | 0.211 |
| Low | 14 | 12 (85.7) | 2 (14.3) | |
| Moderate-High | 8 | 5 (62.5) | 3 (37.5) | |
| Clinical stage pre-CRT | 22 | 17 | 5 | 0.312 |
| II | 3 | 3 (100) | 0 (0) | |
| III | 19 | 14 (73.7) | 5 (26.3) | |
| ECOG 2 | 22 | 17 | 5 | 0.962 |
| 0 | 13 | 10 (76.9) | 3 (23.1) | |
| 1 | 9 | 7 (77.8) | 2 (22.2) | |
| ypT 3 | 20 | 15 | 5 | 0.002 |
| 0–2 | 12 | 12 (100) | 0 (0) | |
| 3–4 | 8 | 3 (37.5) | 5 (62.5) | |
| ypN 4 | 20 | 15 | 5 | 0.091 |
| 0 | 14 | 12 (85.7) | 2 (14.3) | |
| 1–2 | 6 | 3 (50) | 3 (50) | |
| Pathological stage | 20 | 15 | 5 | 0.020 |
| yp0-I | 9 | 19 (100) | 0 (0) | |
| ypII-III | 11 | 6 (54.5) | 5 (45.5) |
| Response to Neoadjuvant CRT 1 | ||||
|---|---|---|---|---|
| Response | No. Cases | Responders 2 (%) | Non-Responders 3 (%) | p |
| MiR-199b expression | 22 | 11 | 11 | 0.011 |
| Low | 5 | 0 (0) | 5 (100) | |
| High | 17 | 11 (64.7) | 6 (35.3) | |
| Downstaging | No. Cases | Yes (%) | No (%) | p |
|---|---|---|---|---|
| MiR-199b expression | 22 | 15 | 7 | 0.009 |
| Low | 5 | 1 (20) | 4 (80) | |
| High | 17 | 14 (82.4) | 3 (17.6) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, A.; Cristóbal, I.; Rubio, J.; Caramés, C.; Luque, M.; Sanz-Alvarez, M.; Morales-Gallego, M.; Madoz-Gúrpide, J.; Rojo, F.; García-Foncillas, J. MicroRNA-199b Deregulation Shows Oncogenic Properties and Promising Clinical Value as Circulating Marker in Locally Advanced Rectal Cancer Patients. Int. J. Mol. Sci. 2022, 23, 2203. https://doi.org/10.3390/ijms23042203
Santos A, Cristóbal I, Rubio J, Caramés C, Luque M, Sanz-Alvarez M, Morales-Gallego M, Madoz-Gúrpide J, Rojo F, García-Foncillas J. MicroRNA-199b Deregulation Shows Oncogenic Properties and Promising Clinical Value as Circulating Marker in Locally Advanced Rectal Cancer Patients. International Journal of Molecular Sciences. 2022; 23(4):2203. https://doi.org/10.3390/ijms23042203
Chicago/Turabian StyleSantos, Andrea, Ion Cristóbal, Jaime Rubio, Cristina Caramés, Melani Luque, Marta Sanz-Alvarez, Miriam Morales-Gallego, Juan Madoz-Gúrpide, Federico Rojo, and Jesús García-Foncillas. 2022. "MicroRNA-199b Deregulation Shows Oncogenic Properties and Promising Clinical Value as Circulating Marker in Locally Advanced Rectal Cancer Patients" International Journal of Molecular Sciences 23, no. 4: 2203. https://doi.org/10.3390/ijms23042203
APA StyleSantos, A., Cristóbal, I., Rubio, J., Caramés, C., Luque, M., Sanz-Alvarez, M., Morales-Gallego, M., Madoz-Gúrpide, J., Rojo, F., & García-Foncillas, J. (2022). MicroRNA-199b Deregulation Shows Oncogenic Properties and Promising Clinical Value as Circulating Marker in Locally Advanced Rectal Cancer Patients. International Journal of Molecular Sciences, 23(4), 2203. https://doi.org/10.3390/ijms23042203