
Disaggregation of Islet Amyloid Polypeptide Fibrils as a Potential Anti-Fibrillation Mechanism of Tetrapeptide TNGQ




Abstract
:1. Introduction
2. Results
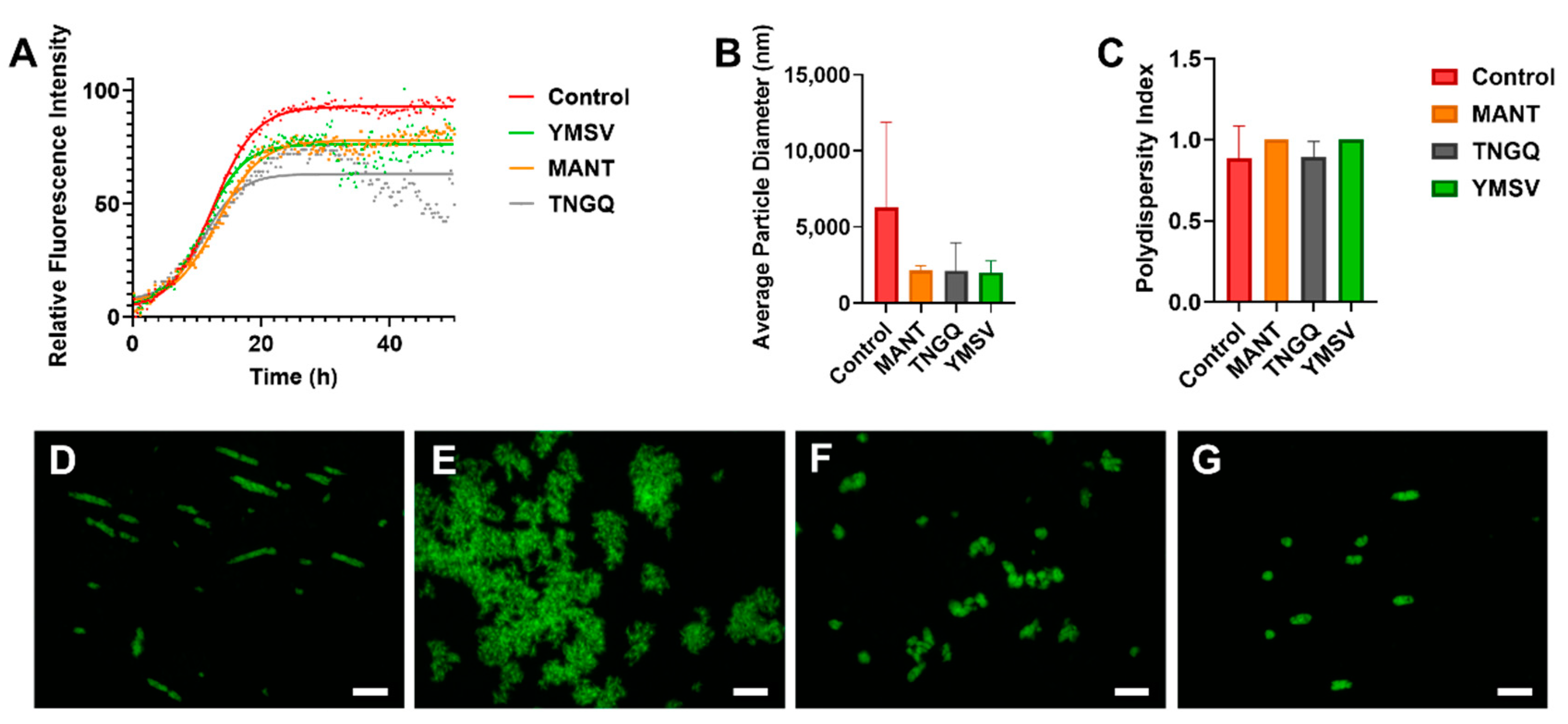
2.1. Thioflavin T Fluorescence Kinetics
2.2. Fluorescence Morphology of IAPP Fibrils
2.3. IAPP Fibrillation Progression
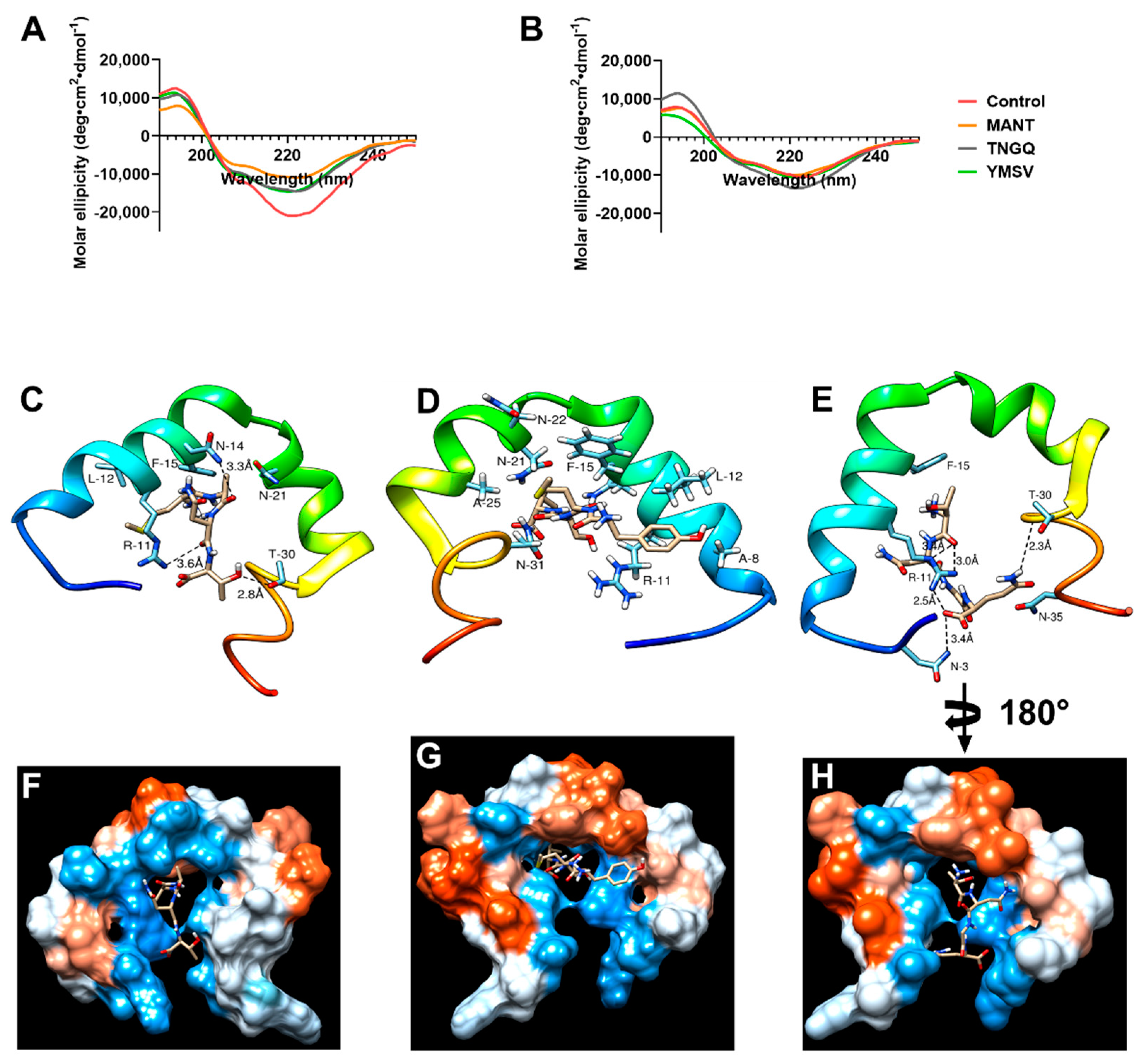
2.4. IAPP Secondary Structure during Fibrillation
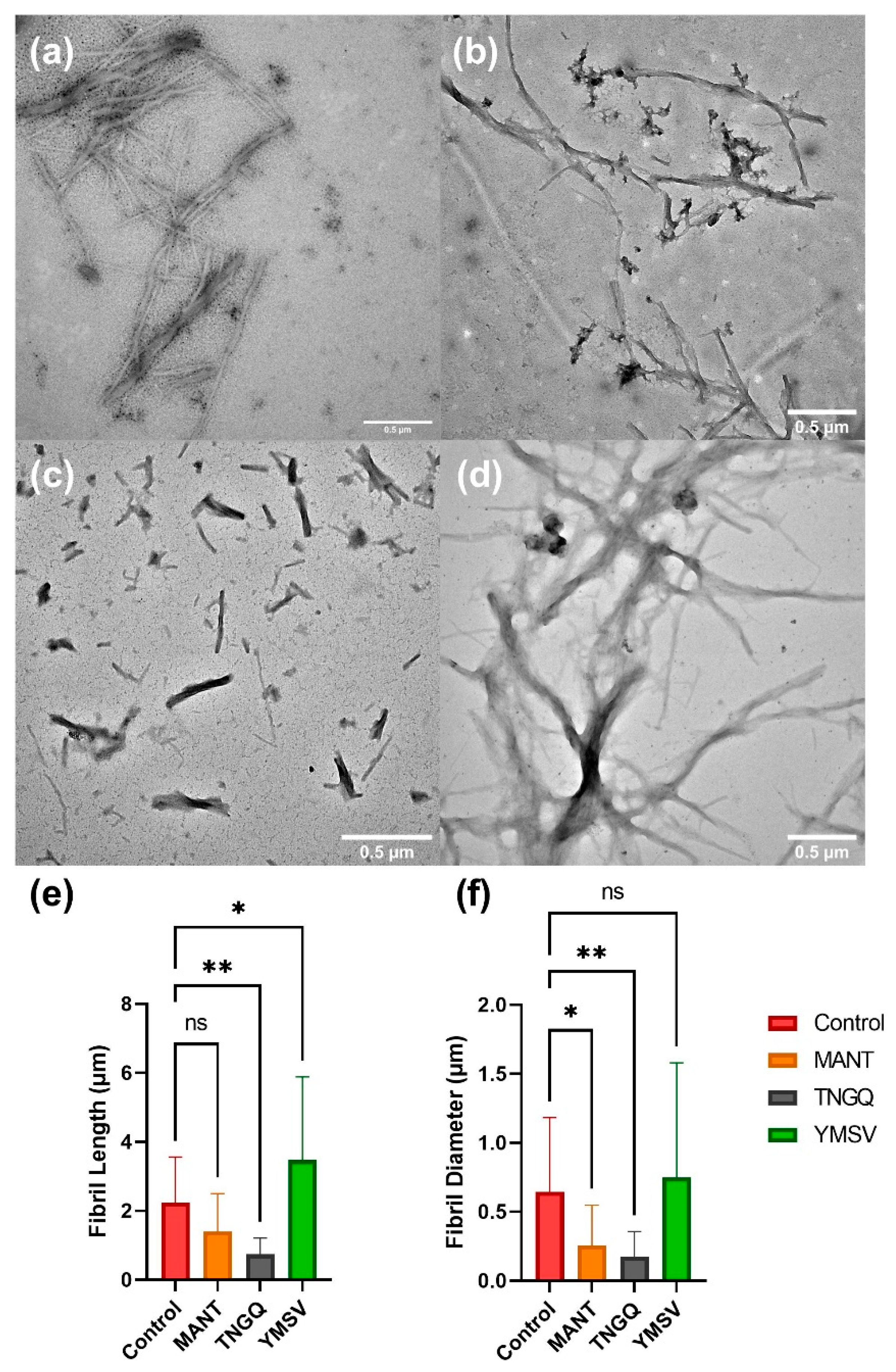
2.5. IAPP Fibril Morphology
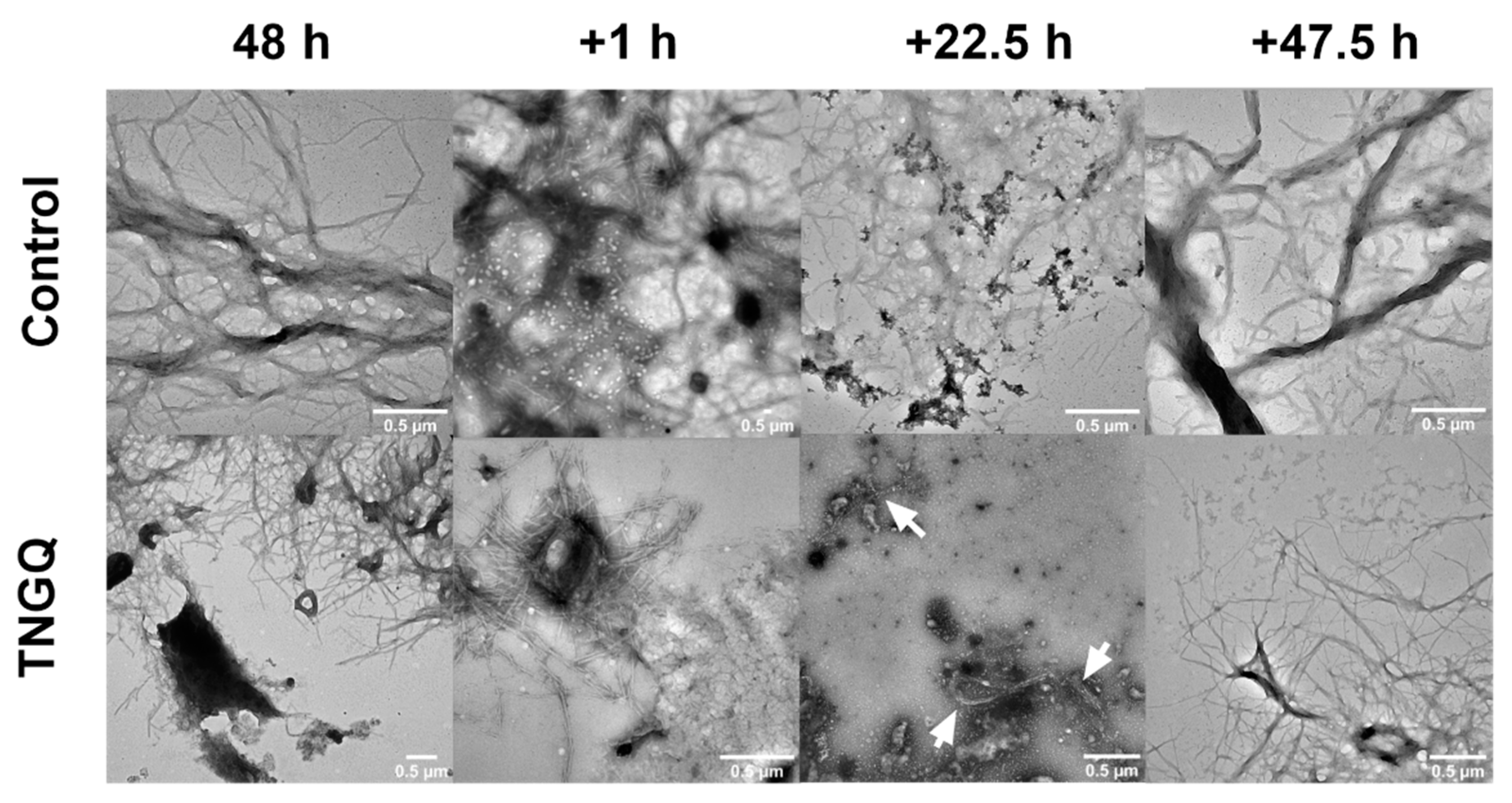
2.6. Disaggregation of Pre-Formed IAPP Fibrils
2.7. In Silico Drug-Likeness of the Peptides
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. IAPP Preparation
4.3. Thioflavin T (ThT) Fluorescence Assay
4.4. Circular Dichroism (CD) Spectroscopy
4.5. Molecular Docking of IAPP-Tetrapeptide Interaction for Determination of Substantive Binding Site and Relative Binding Affinity
4.6. Fluorescence Microscopy
4.7. Dynamic Light Scattering (DLS)
4.8. Transmission Electron Microscopy (TEM)
4.9. In Silico ADME/Tox and Physicochemical Properties of the Peptides
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abioye, R.O.; Udenigwe, C.C. Potential of peptides and phytochemicals in attenuating different phases of islet amyloid polypeptide fibrillation for type 2 diabetes. Food Sci. Hum. Wellness 2021, 10, 258–268. [Google Scholar] [CrossRef]
- Kahn, S.E.; Andrikopoulos, S.; Verchere, C.B. Islet amyloid. Diabetes 1999, 48, 241–242. [Google Scholar] [CrossRef]
- Zraika, S.; Hull, R.L.; Udayasankar, J.; Aston-Mourney, K.; Subramanian, S.L.; Kisilevsky, R.; Szarek, W.A.; Kahn, S.E. Oxidative Stress Is Induced by Islet amyloid formation and time-dependently mediates amyloid-induced beta cell apoptosis. Diabetologia 2009, 52, 626–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahn, S.E.; D’Alessio, D.A.; Schwartz, M.W.; Fujimoto, W.Y.; Ensinck, J.W.; Taborsky, G.J., Jr.; Porte, D., Jr. Evidence of cosecretion of islet amyloid polypeptide and insulin by beta-cells. Diabetes 1990, 39, 634–639. [Google Scholar] [CrossRef] [Green Version]
- Westermark, P.; Andersson, A.; Westermark, G.T. Islet amyloid polypeptide, islet amyloid, and diabetes mellitus. Physiol. Rev. 2011, 91, 795–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asthana, S.; Mallick, B.; Alexandrescu, A.T.; Jha, S. IAPP in type II diabetes: Basic research on structure, molecular interactions, and disease mechanisms suggests potential intervention strategies. Biochim. Et Biophys. Acta (BBA)-Biomembr. 2018, 1860, 1765–1782. [Google Scholar] [CrossRef]
- Höppener, J.W.M.; Ahrén, B.; Lips, C.J.M. Islet amyloid and type 2 diabetes mellitus. N. Engl. J. Med. 2000, 343, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Kapurniotu, A. Amyloidogenicity and cytotoxicity of islet amyloid polypeptide. Pept. Sci. Orig. Res. Biomol. 2001, 60, 438–459. [Google Scholar] [CrossRef]
- Lorenzo, A.; Yankner, B.A. Beta-amyloid neurotoxicity requires fibril formation and is inhibited by congo red. Proc. Natl. Acad. Sci. USA 1994, 91, 12243–12247. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo, A.; Razzaboni, B.; Weir, G.C.; Yankner, B.A. Pancreatic islet cell toxicity of amylin associated with type-2 diabetes mellitus. Nature 1994, 368, 756–760. [Google Scholar] [CrossRef]
- Brender, J.R.; Salamekh, S.; Ramamoorthy, A. Membrane disruption and early events in the aggregation of the diabetes related peptide IAPP from a molecular prospective. Acc. Chem. Res. 2012, 45, 454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milardi, D.; Gazit, E.; Radford, S.E.; Xu, Y.; Gallardo, R.U.; Caflisch, A.; Westermark, G.T.; Westermark, P.; La Rosa, C.; Ramamoorthy, A. Proteostasis of islet amyloid polypeptide: A molecular perspective of risk factors and protective strategies for type II diabetes. Chem. Rev. 2021, 121, 1845–1893. [Google Scholar] [CrossRef] [PubMed]
- Porat, Y.; Abramowitz, A.; Gazit, E. Inhibition of amyloid fibril formation by polyphenols: Structural similarity and aromatic interactions as a common inhibition mechanism. Chem. Biol. Drug Des. 2006, 67, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Pithadia, A.; Brender, J.R.; Fierke, C.A.; Ramamoorthy, A. Inhibition of IAPP aggregation and toxicity by natural products. J. Diabetes Res. 2016, 2016, 2046327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tak, Y.; Kaur, M.; Amarowicz, R.; Bhatia, S.; Gautam, C. Pulse derived bioactive peptides as novel nutraceuticals: A review. Int. J. Pept. Res. Ther. 2021, 27, 2057–2068. [Google Scholar] [CrossRef]
- Daroit, D.J.; Brandelli, A. In vivo bioactivities of food protein-derived peptides—A current review. Curr. Opin. Food Sci. 2021, 39, 120–129. [Google Scholar] [CrossRef]
- Zhou, P.; Jin, B.; Li, H.; Huang, S.Y. HPEPDOCK: A web server for blind peptide–protein docking based on a hierarchical algorithm. Nucleic Acids Res. 2018, 46, W443–W450. [Google Scholar] [CrossRef]
- Huang, S.Y.; Zou, X. An iterative knowledge-based scoring function for protein-protein recognition. Proteins. 2008, 72, 557–579. [Google Scholar] [CrossRef]
- Van de Waterbeemd, H. In silico models to predict oral absorption. Compr. Med. Chem. II 2007, 5, 669–697. [Google Scholar] [CrossRef]
- Ertl, P.; Rohde, B.; Selzer, P. Fast calculation of molecular polar surface area as a sum of fragment-based contributions and its application to the prediction of drug transport properties. J. Med. Chem. 2000, 43, 3714–3717. [Google Scholar] [CrossRef]
- Veber, D.F.; Johnson, S.R.; Cheng, H.; Smit, B.R.; Ward, K.W.; Kopple, K.D. Molecular properties that influence the oral bioavailability of drug candidates. J. Med. Chem. 2002, 45, 2615–2623. [Google Scholar] [CrossRef]
- Delaney, J.S. ESOL: Estimating aqueous solubility directly from molecular structure. J. Chem. Inf. Comput. Sci. 2004, 44, 1000–1005. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Kapoor, P.; Chaudhary, K.; Gautam, A.; Kumar, R.; Consortium, O.S.D.D.; Raghava, G.P.S. In silico approach for predicting toxicity of peptides and proteins. PLoS ONE 2013, 8, e73957. [Google Scholar] [CrossRef] [Green Version]
- Martin, Y.C. A bioavailability score. J. Med. Chem. 2005, 48, 3164–3170. [Google Scholar] [CrossRef]
- Ratha, B.N.; Ghosh, A.; Brender, J.R.; Gayen, N.; Ilyas, H.; Neeraja, C.; Das, K.P.; Mandal, A.K.; Bhunia, A. Inhibition of insulin amyloid fibrillation by a novel amphipathic heptapeptide. J. Biol. Chem. 2016, 291, 23545–23556. [Google Scholar] [CrossRef] [Green Version]
- Paslawski, W.; Andreasen, M.; Nielsen, S.; Lorenzen, N.; Thomsen, K.; Kaspersen, J.; Pedersen, J.; Otzen, D. High stability and cooperative unfolding of α-synuclein oligomers. Biochemistry 2014, 53, 6252–6263. [Google Scholar] [CrossRef]
- Christensen, M.; Skeby, K.K.; Schiøtt, B. Identification of key interactions in the initial self-assembly of amylin in a membrane environment. Biochemistry 2017, 56, 4884–4894. [Google Scholar] [CrossRef] [PubMed]
- Nagel-Steger, L.; Owen, M.C.; Strodel, B. An account of amyloid oligomers: Facts and figures obtained from experiments and simulations. ChemBioChem 2016, 17, 657–676. [Google Scholar] [CrossRef]
- Cohen, S.I.A.; Cukalevski, R.; Michaels, T.C.T.; Šarić, A.; Törnquist, M.; Vendruscolo, M.; Dobson, C.M.; Buell, A.K.; Knowles, T.P.J.; Linse, S. Distinct thermodynamic signatures of oligomer generation in the aggregation of the amyloid-β peptide. Nat. Chem. 2018, 10, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Kapurniotu, A.; Schmauder, A.; Tenidis, K. Structure-based design and study of non-amyloidogenic, double n-methylated iapp amyloid core sequences as inhibitors of IAPP amyloid formation and cytotoxicity. J. Mol. Biol. 2002, 315, 339–350. [Google Scholar] [CrossRef]
- Scrocchi, L.A.; Chen, Y.; Waschuk, S.; Wang, F.; Cheung, S.; Darabie, A.A.; McLaurin, J.A.; Fraser, P.E. Design of peptide-based inhibitors of human islet amyloid polypeptide fibrillogenesis. J. Mol. Biol. 2002, 318, 697–706. [Google Scholar] [CrossRef]
- Sivanesam, K.; Andersen, N.H. Inhibition of human amylin amyloidogenesis by human amylin-fragment peptides: Exploring the effects of serine residues and oligomerization upon inhibitory potency. Biochemistry 2017, 56, 5373–5379. [Google Scholar] [CrossRef]
- Wang, L.; Lei, L.; Li, Y.; Wang, L.; Li, F. A hIAPP-Derived all-D-amino-acid inhibits hIAPP fibrillation efficiently at membrane surface by targeting α-helical oligomeric intermediates. FEBS Lett. 2014, 588, 884–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Q.; Boyer, D.R.; Sawaya, M.R.; Ge, P.; Eisenberg, D.S. Cryo-EM structure and inhibitor design of human IAPP (amylin) fibrils. Nat. Struct. Mol. Biol. 2020, 27, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Lv, W.; Jiao, A.; Zhang, C.; Zhang, J. A novel pentapeptide inhibitor reduces amyloid deposit formation by direct interaction with hIAPP. Int. J. Endocrinol. 2019, 2019, 9062032. [Google Scholar] [CrossRef] [PubMed]
- Profit, A.A.; Felsen, V.; Chinwong, J.; Mojica, E.-R.E.; Desamero, R.Z.B. Evidence of π-stacking interactions in the self-assembly of hIAPP22-29. Proteins 2013, 81, 690–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gazit, E. Mechanisms of amyloid fibril self-assembly and inhibition. FEBS J. 2005, 272, 5971–5978. [Google Scholar] [CrossRef]
- Chaari, A.; Ladjimi, M. Human islet amyloid polypeptide (hIAPP) aggregation in type 2 diabetes: Correlation between intrinsic physicochemical properties of hIAPP aggregates and their cytotoxicity. Int. J. Biol. Macromol. 2019, 136, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Miles, A.J.; Wallace, B.A. CDtoolX, a downloadable software package for processing and analyses of circular dichroism spectroscopic data. Protein Sci. 2018, 27, 1717–1722. [Google Scholar] [CrossRef]
- Micsonai, A.; Wien, F.; Bulyáki, É.; Kun, J.; Moussong, É.; Lee, Y.H.; Goto, Y.; Réfrégiers, M.; Kardos, J. BeStSel: A web server for accurate protein secondary structure prediction and fold recognition from the circular dichroism spectra. Nucleic Acids Res. 2018, 46, W315. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physicochemical Properties | Fibrillation Parameters | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| MW (Da) | Hydrophobicity | Net Charge | Boman Index | Instability Index | Aliphatic Index | Fmax | t50 (h) | τ | K | Lag Time (h) | |
| Control | n/a | n/a | n/a | n/a | n/a | n/a | 92.89 | 12.45 | 3.33 | 0.30 | 5.79 |
| MANT | 435.50 | −0.13 | −0.002 | 1.26 | 17.13 | 25 | 77.98 | 13.13 | 3.41 | 0.29 | 6.31 |
| TNGQ | 418.40 | −2.03 | −0.002 | 3.45 | −67.65 | 0 | 63.11 | 11.00 | 2.89 | 0.35 | 5.22 |
| YMSV | 498.59 | 1.00 | −0.003 | −0.71 | 227.2 | 72.5 | 76.15 | 11.30 | 2.95 | 0.34 | 5.39 |
| Peptide | Physicochemical Properties | Toxicity | Lipophilicity | Drug-Likeness | Pharmacokinetics | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ROTB (n) < 10 | HBA (n) < 10 | HBD (n) < 5 | ESOL Log S | SVM Score (<0.0) | TPSA (Å2) < 140 | ClogP (o/w) < 5 | Bioavailability Score | Lipinski Filter | GIA | P-Gly Substrate | CYP3A4 Inhibitor | |
| MANT | 16 | 8 | 7 | 1.83 (HS) | −0.81 Non-toxin | 239.24 | −2.33 | 0.17 | No | Low | Yes | No |
| TNGQ | 16 | 9 | 8 | 3.32 (HS) | −0.72 Non-toxin | 257.03 | −3.94 | 0.17 | No | Low | Yes | No |
| YMSV | 17 | 8 | 7 | 0.23 (HS) | −0.89 Non-toxin | 216.38 | −0.39 | 0.17 | No | Low | Yes | No |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abioye, R.O.; Okagu, O.D.; Udenigwe, C.C. Disaggregation of Islet Amyloid Polypeptide Fibrils as a Potential Anti-Fibrillation Mechanism of Tetrapeptide TNGQ. Int. J. Mol. Sci. 2022, 23, 1972. https://doi.org/10.3390/ijms23041972
Abioye RO, Okagu OD, Udenigwe CC. Disaggregation of Islet Amyloid Polypeptide Fibrils as a Potential Anti-Fibrillation Mechanism of Tetrapeptide TNGQ. International Journal of Molecular Sciences. 2022; 23(4):1972. https://doi.org/10.3390/ijms23041972
Chicago/Turabian StyleAbioye, Raliat O., Ogadimma D. Okagu, and Chibuike C. Udenigwe. 2022. "Disaggregation of Islet Amyloid Polypeptide Fibrils as a Potential Anti-Fibrillation Mechanism of Tetrapeptide TNGQ" International Journal of Molecular Sciences 23, no. 4: 1972. https://doi.org/10.3390/ijms23041972
APA StyleAbioye, R. O., Okagu, O. D., & Udenigwe, C. C. (2022). Disaggregation of Islet Amyloid Polypeptide Fibrils as a Potential Anti-Fibrillation Mechanism of Tetrapeptide TNGQ. International Journal of Molecular Sciences, 23(4), 1972. https://doi.org/10.3390/ijms23041972