
Alpha 1 Antitrypsin Regulates Trophoblast Syncytialization and Inflammatory Factor Expression


Abstract
:1. Introduction
2. Results
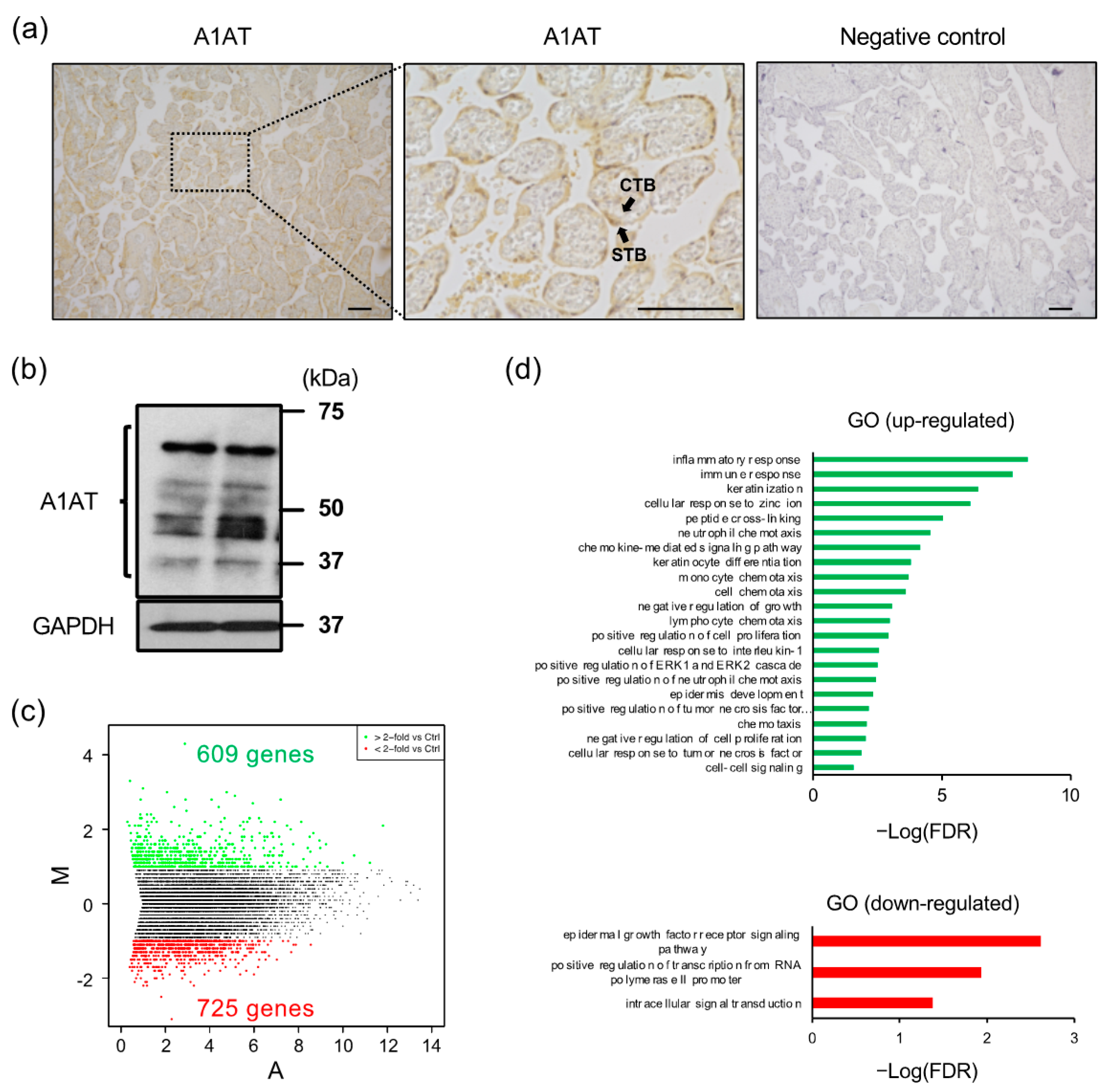
2.1. A1AT Knockdown Upregulates Inflammatory Response-Related Gene Expression in Trophoblasts
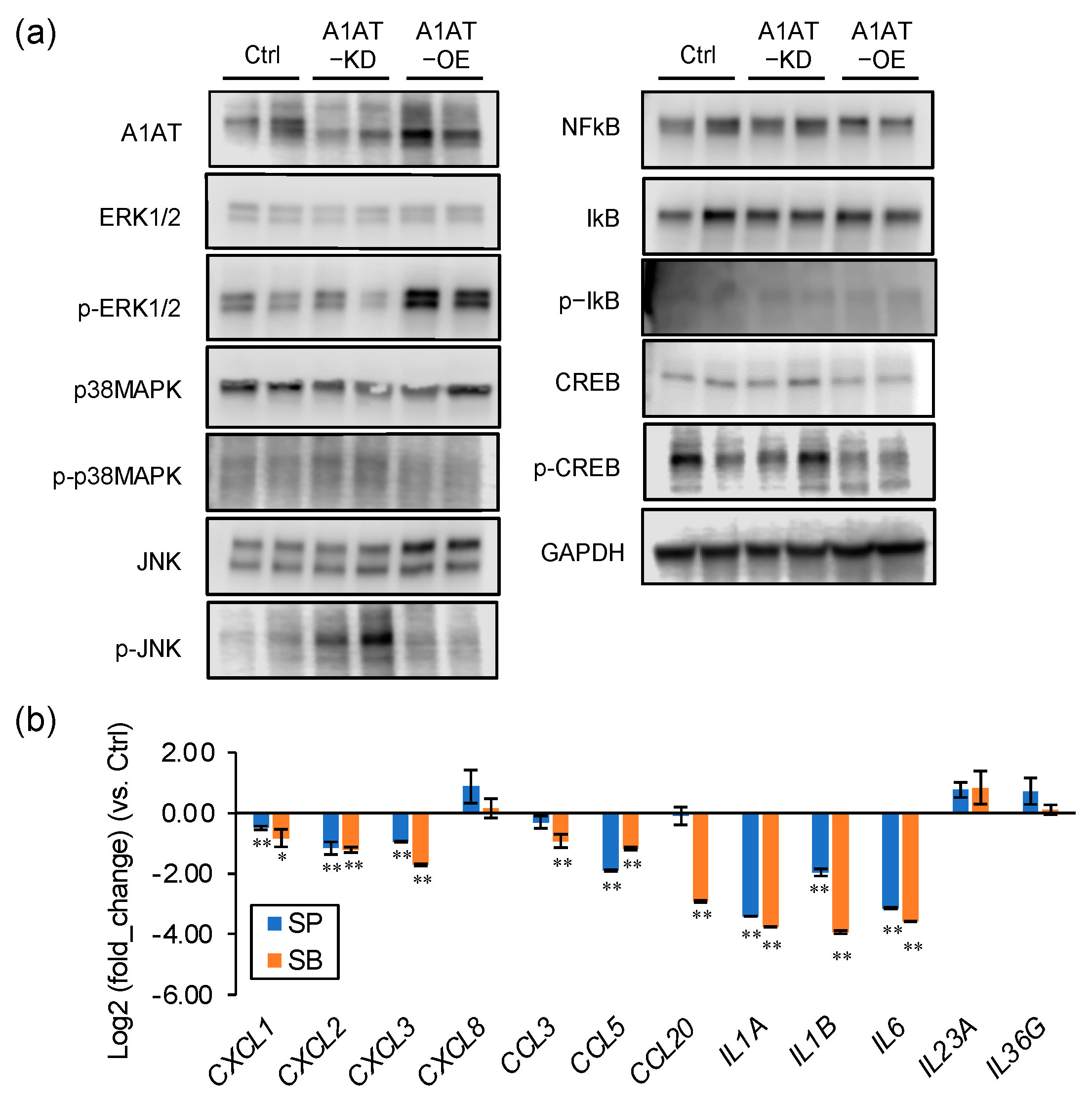
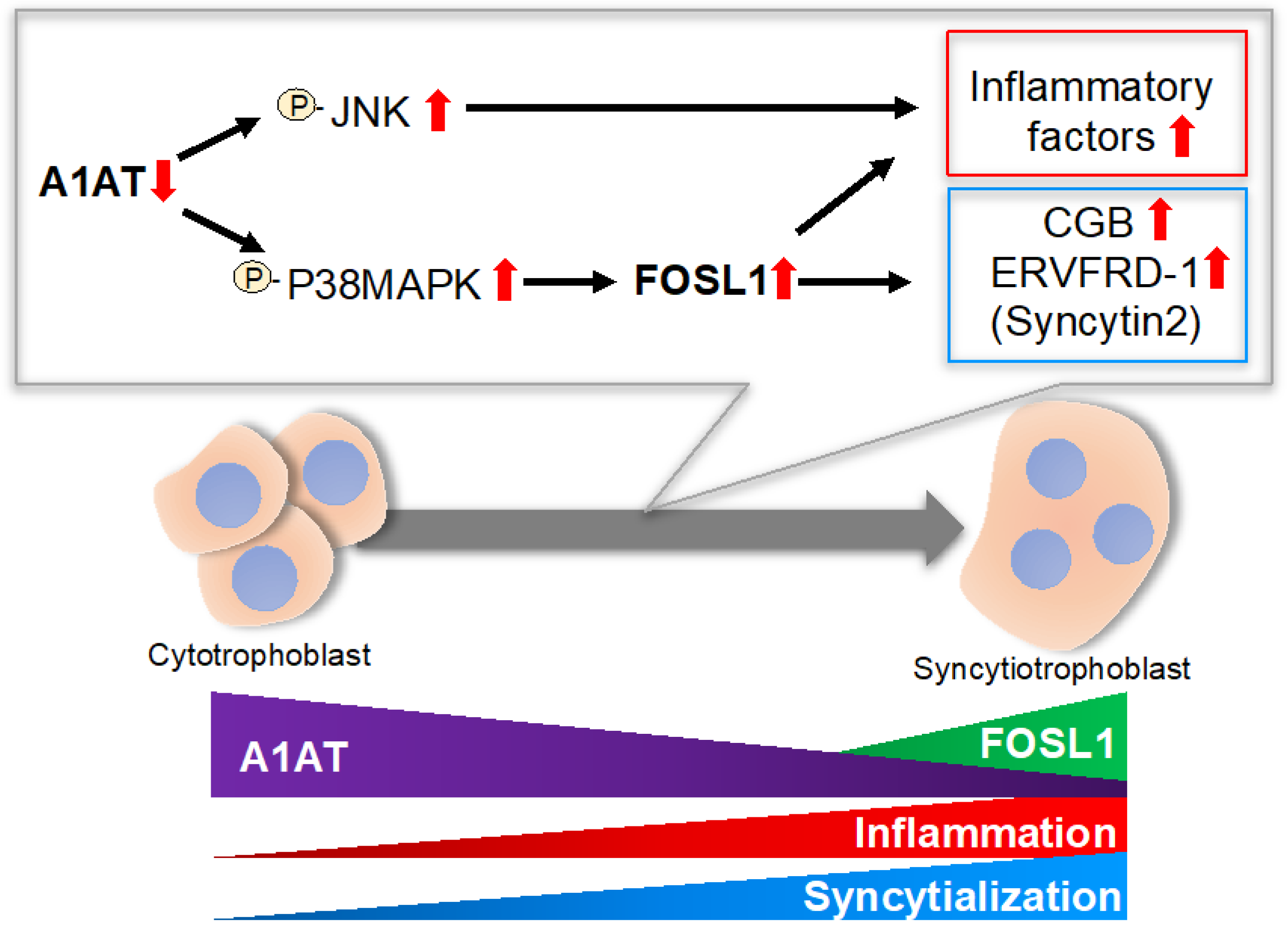
2.2. A1AT Regulates Activation of p38MAPK and JNK and Expression of Inflammatory Cytokines in Trophoblasts
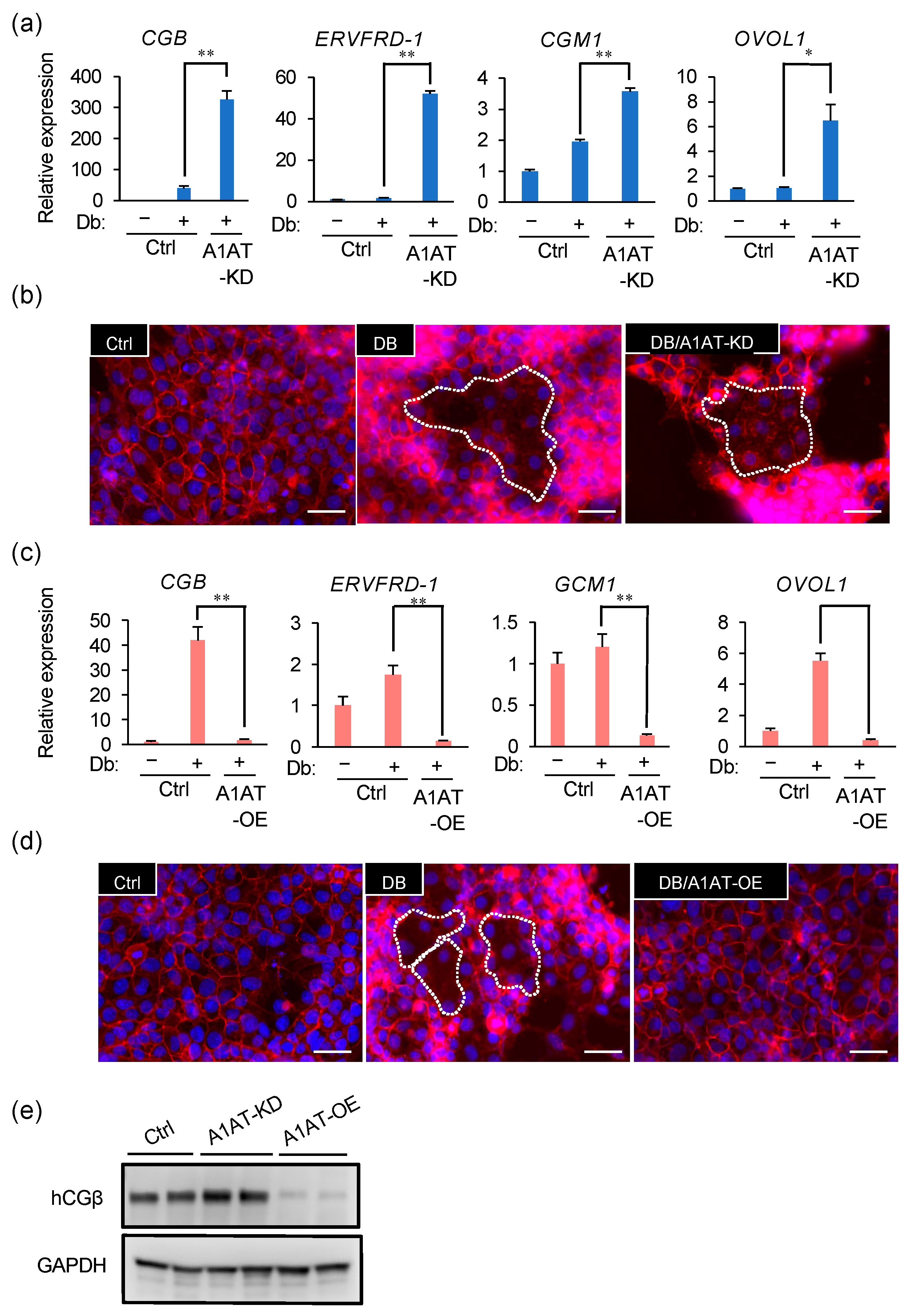
2.3. A1AT Expression Alters the Syncytialization of Trophoblasts
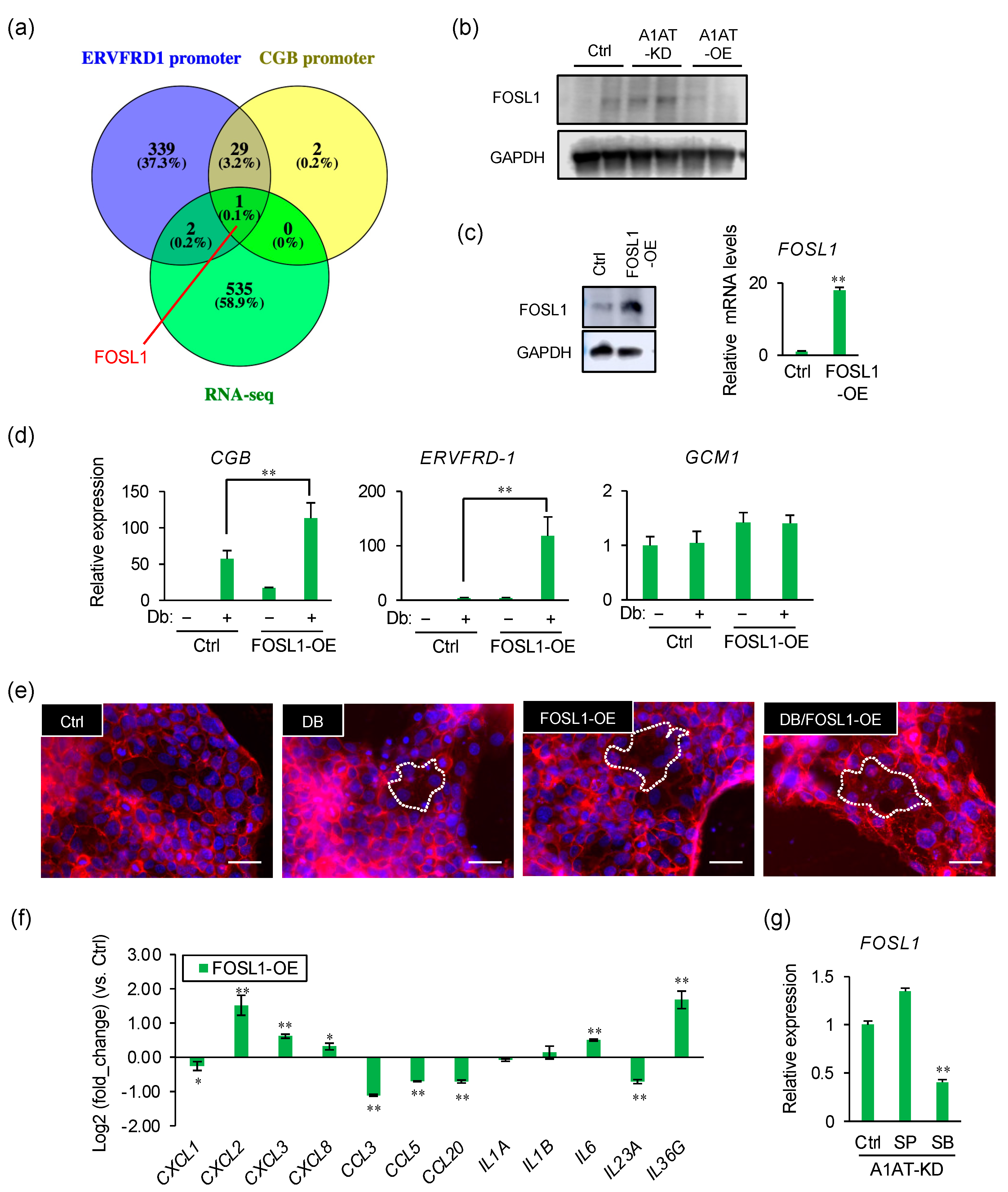
2.4. FOSL1 Mediates A1AT Knockdown-Induced Syncytialization of Trophoblasts
3. Discussion
4. Materials and Methods
4.1. Immunohistochemistry
4.2. Cell Culture
4.3. Western Blotting
4.4. Transfection of Small Interfering (si)RNA
4.5. RNA-Sequencing (RNA-Seq), GO, and Pathway Analyses
4.6. RNA Extraction and Quantitative RT-PCR
4.7. Transfection of the Expression Plasmid Construct
4.8. Cell Fusion Assay
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hunt, J.M.; Tuder, R. Alpha 1 anti-trypsin: One protein, many functions. Curr. Mol. Med. 2012, 12, 827–835. [Google Scholar] [CrossRef]
- Tamura, K.; Takashima, H.; Fumoto, K.; Kajihara, T.; Uchino, S.; Ishihara, O.; Yoshie, M.; Kusama, K.; Tachikawa, E. Possible Role of alpha1-Antitrypsin in Endometriosis-Like Grafts From a Mouse Model of Endometriosis. Reprod. Sci. 2015, 22, 1088–1097. [Google Scholar] [CrossRef] [PubMed]
- Ando, Y.; Kuroda, A.; Kusama, K.; Matsutani, T.; Matsuda, A.; Tamura, K. Impact of serine protease inhibitor alpha1-antitrypsin on expression of endoplasmic reticulum stress-induced proinflammatory factors in adipocytes. Biochem. Biophys. Rep. 2021, 26, 100967. [Google Scholar] [CrossRef] [PubMed]
- Kaner, Z.; Ochayon, D.E.; Shahaf, G.; Baranovski, B.M.; Bahar, N.; Mizrahi, M.; Lewis, E.C. Acute phase protein α1-antitrypsin reduces the bacterial burden in mice by selective modulation of innate cell responses. J. Infect. Dis. 2015, 211, 1489–1498. [Google Scholar] [CrossRef] [Green Version]
- Jonigk, D.; Al-Omari, M.; Maegel, L.; Müller, M.; Izykowski, N.; Hong, J.; Hong, K.; Kim, S.H.; Dorsch, M.; Mahadeva, R.; et al. Anti-inflammatory and immunomodulatory properties of α1-antitrypsin without inhibition of elastase. Proc. Natl. Acad. Sci. USA 2013, 110, 15007–15012. [Google Scholar] [CrossRef] [Green Version]
- Pott, G.B.; Chan, E.D.; Dinarello, C.A.; Shapiro, L. Alpha-1-antitrypsin is an endogenous inhibitor of proinflammatory cytokine production in whole blood. J. Leukoc. Biol. 2009, 85, 886–895. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, K.; Kusama, K.; Fukushima, Y.; Ohmaru-Nakanishi, T.; Kato, K.; Tamura, K. Alpha-1 Antitrypsin-Induced Endoplasmic Reticulum Stress Promotes Invasion by Extravillous Trophoblasts. Int. J. Mol. Sci. 2021, 22, 3683. [Google Scholar] [CrossRef]
- Gou, W.; Wang, J.; Song, L.; Kim, D.S.; Cui, W.; Strange, C.; Wang, H. Alpha-1 antitrypsin suppresses macrophage activation and promotes islet graft survival after intrahepatic islet transplantation. Am. J. Transplant. 2021, 21, 1713–1724. [Google Scholar] [CrossRef]
- Bischof, P.; Irminger-Finger, I. The human cytotrophoblastic cell, a mononuclear chameleon. Int. J. Biochem. Cell Biol. 2005, 37, 1–16. [Google Scholar] [CrossRef]
- Moffett, A.; Loke, C. Immunology of placentation in eutherian mammals. Nat. Rev. Immunol. 2006, 6, 584–594. [Google Scholar] [CrossRef] [PubMed]
- Loregger, T.; Pollheimer, J.; Knöfler, M. Regulatory Transcription Factors Controlling Function and Differentiation of Human Trophoblast—A Review. Placenta 2003, 24, S104–S110. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.H. Endocrine cell lines from the placenta. Mol. Cell Endocrinol. 2004, 228, 103–119. [Google Scholar] [CrossRef]
- Tuckey, R.C. Progesterone synthesis by the human placenta. Placenta 2005, 26, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.Y.; Wang, L.J.; Chen, C.P.; Chen, L.F.; Chen, Y.H.; Chen, H. GCM1 regulation of the expression of syncytin 2 and its cognate receptor MFSD2A in human placenta. Biol. Reprod. 2010, 83, 387–395. [Google Scholar] [CrossRef] [Green Version]
- Renaud, S.J.; Chakraborty, D.; Mason, C.W.; Rumi, M.A.; Vivian, J.L.; Soares, M.J. OVO-like 1 regulates progenitor cell fate in human trophoblast development. Proc. Natl. Acad. Sci. USA 2015, 112, E6175–E6184. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.; Shen, K.; Lin, M.; Chen, P.; Lin, C.; Chang, G.D.; Chen, H. GCMa regulates the syncytin-mediated trophoblastic fusion. J. Biol. Chem. 2002, 277, 50062–50068. [Google Scholar] [CrossRef] [Green Version]
- Gauster, M.; Moser, G.; Orendi, K.; Huppertz, B. Factors involved in regulating trophoblast fusion: Potential role in the development of preeclampsia. Placenta 2009, 30 (Suppl. A), S49–S54. [Google Scholar] [CrossRef] [PubMed]
- Than, N.G.; Romero, R.; Tarca, A.L.; Kekesi, K.A.; Xu, Y.; Xu, Z.; Juhasz, K.; Bhatti, G.; Leavitt, R.J.; Gelencser, Z.; et al. Integrated Systems Biology Approach Identifies Novel Maternal and Placental Pathways of Preeclampsia. Front. Immunol. 2018, 9, 1661. [Google Scholar] [CrossRef]
- Starodubtseva, N.; Nizyaeva, N.; Baev, O.; Bugrova, A.; Gapaeva, M.; Muminova, K.; Kononikhin, A.; Frankevich, V.; Nikolaev, E.; Sukhikh, G. SERPINA1 Peptides in Urine as A Potential Marker of Preeclampsia Severity. Int. J. Mol. Sci. 2020, 21, 914. [Google Scholar] [CrossRef] [Green Version]
- Szilagyi, A.; Gelencser, Z.; Romero, R.; Xu, Y.; Kiraly, P.; Demeter, A.; Palhalmi, J.; Gyorffy, B.A.; Juhasz, K.; Hupuczi, P.; et al. Placenta-Specific Genes, Their Regulation During Villous Trophoblast Differentiation and Dysregulation in Preterm Preeclampsia. Int. J. Mol. Sci. 2020, 21, 628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frochaux, V.; Hildebrand, D.; Talke, A.; Linscheid, M.W.; Schlüter, H. Alpha-1-antitrypsin: A novel human high temperature requirement protease A1 (HTRA1) substrate in human placental tissue. PLoS ONE 2014, 9, e109483. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Xu, J.; Zhou, Q.; Wang, R.; Liu, N.; Wu, Y.; Yuan, H.; Che, H. Alpha-1 Antitrypsin Prevents the Development of Preeclampsia Through Suppression of Oxidative Stress. Front. Physiol. 2016, 7, 176. [Google Scholar] [CrossRef]
- Feng, Y.L.; Yin, Y.X.; Ding, J.; Yuan, H.; Yang, L.; Xu, J.J.; Hu, L.Q. Alpha-1-antitrypsin suppresses oxidative stress in preeclampsia by inhibiting the p38MAPK signaling pathway: An in vivo and in vitro study. PLoS ONE 2017, 12, e0173711. [Google Scholar] [CrossRef]
- Kaner, Z.; Engelman, R.; Schuster, R.; Rider, P.; Greenberg, D.; Av-Gay, Y.; Benhar, M.; Lewis, E.C. S-Nitrosylation of α1-Antitrypsin Triggers Macrophages Toward Inflammatory Phenotype and Enhances Intra-Cellular Bacteria Elimination. Front. Immunol. 2019, 10, 590. [Google Scholar] [CrossRef] [PubMed]
- Cindrova-Davies, T.; Spasic-Boskovic, O.; Jauniaux, E.; Charnock-Jones, D.S.; Burton, G.J. Nuclear factor-kappa B, p38, and stress-activated protein kinase mitogen-activated protein kinase signaling pathways regulate proinflammatory cytokines and apoptosis in human placental explants in response to oxidative stress: Effects of antioxidant vitamins. Am. J. Pathol. 2007, 170, 1511–1520. [Google Scholar] [CrossRef] [PubMed]
- Szabo, S.; Mody, M.; Romero, R.; Xu, Y.; Karaszi, K.; Mihalik, N.; Xu, Z.; Bhatti, G.; Fule, T.; Hupuczi, P.; et al. Activation of villous trophoblastic p38 and ERK1/2 signaling pathways in preterm preeclampsia and HELLP syndrome. Pathol. Oncol. Res. 2015, 21, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Vaillancourt, C.; Lanoix, D.; Le Bellego, F.; Daoud, G.; Lafond, J. Involvement of MAPK signalling in human villous trophoblast differentiation. Mini. Rev. Med. Chem. 2009, 9, 962–973. [Google Scholar] [CrossRef] [PubMed]
- Shaulian, E.; Karin, M. AP-1 as a regulator of cell life and death. Nat. Cell Biol. 2002, 4, E131–E136. [Google Scholar] [CrossRef]
- Kent, L.N.; Rumi, M.A.; Kubota, K.; Lee, D.S.; Soares, M.J. FOSL1 is integral to establishing the maternal-fetal interface. Mol. Cell Biol. 2011, 31, 4801–4813. [Google Scholar] [CrossRef] [Green Version]
- Kent, L.N.; Konno, T.; Soares, M.J. Phosphatidylinositol 3 kinase modulation of trophoblast cell differentiation. BMC Dev. Biol. 2010, 10, 97. [Google Scholar] [CrossRef] [Green Version]
- Kubota, K.; Kent, L.N.; Rumi, M.A.; Roby, K.F.; Soares, M.J. Dynamic Regulation of AP-1 Transcriptional Complexes Directs Trophoblast Differentiation. Mol. Cell Biol. 2015, 35, 3163–3177. [Google Scholar] [CrossRef] [Green Version]
- Renaud, S.J.; Kubota, K.; Rumi, M.A.; Soares, M.J. The FOS transcription factor family differentially controls trophoblast migration and invasion. J. Biol. Chem. 2014, 289, 5025–5039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wongwananuruk, T.; Sato, T.; Kajihara, T.; Matsumoto, S.; Akita, M.; Tamura, K.; Brosens, J.J.; Ishihara, O. Endometrial androgen signaling and decidualization regulate trophoblast expansion and invasion in co-culture: A time-lapse study. Placenta 2016, 47, 56–62. [Google Scholar] [CrossRef]
- Kusama, K.; Bai, R.; Imakawa, K. Regulation of human trophoblast cell syncytialization by transcription factors STAT5B and NR4A3. J. Cell Biochem. 2018, 119, 4918–4927. [Google Scholar] [CrossRef] [PubMed]
- Kusama, K.; Fukushima, Y.; Yoshida, K.; Azumi, M.; Yoshie, M.; Mizuno, Y.; Kajihara, T.; Tamura, K. PGE2 and Thrombin Induce Myofibroblast Transdifferentiation via Activin A and CTGF in Endometrial Stromal Cells. Endocrinology 2021, 162, bqab207. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name (Accession No.) | Sequence (5′---3′) | Product Length (bp) |
|---|---|---|
| GAPDH NM_002046.7 | AGCCACATCGCTCAGACA | 66 |
| GCCCAATACGACCAAATCC | ||
| CXCL1 NM_001511.4 | AGCTTGCCTCAATCCTGCATCC | 119 |
| TCCTTCAGGAACAGCCACCAGT | ||
| CXCL2 NM_002089.4 | GGCAGAAAGCTTGTCTCAACCC | 127 |
| CTCCTTCAGGAACAGCCACCAA | ||
| CXCL3 NM_002090.3 | TTCACCTCAAGAACATCCAAAGTG | 94 |
| TTCTTCCCATTCTTGAGTGTGGC | ||
| CXCL8 | GAGAGTGATTGAGAGTGGACCAC | 112 |
| NM_000584.4 | CACAACCCTCTGCACCCAGTTT | |
| CCL3 | ACTTTGAGACGAGCAGCCAGTG | 101 |
| NM_002983.3 | TTTCTGGACCCACTCCTCACTG | |
| CCL5 | CCTGCTGCTTTGCCTACATTGC | 125 |
| NM_002985.3 | ACACACTTGGCGGTTCTTTCGG | |
| CCL20 | AAGTTGTCTGTGTGCGCAAATCC | 107 |
| NM_004591.3 | CCATTCCAGAAAAGCCACAGTTTT | |
| IL1A | TGTATGTGACTGCCCAAGATGAAG | 96 |
| NM_000575.5 | AGAGGAGGTTGGTCTCACTACC | |
| IL1B | TGATGGCTTATTACAGTGGCAATG | 140 |
| NM_000576.3 | GTAGTGGTGGTGGGAGATTCG | |
| IL6 | CAGGAGCCCAGCTATGAACT | 85 |
| NM_000600.5 | AGCAGGCAACACCAGGAG | |
| IL23A | GAGCCTTCTCTGCTCCCTGATA | 121 |
| NM_016584.3 | GACTGAGGCTTGGAATCTGCTG | |
| IL36G | GAAGGTTGGAGAACAGCCCACA | 130 |
| NM_019618.4 | AGACTCAAGGGTGGAGGTCCTA | |
| CGB | CCTGGCCTTGTCTACCTCTT | 108 |
| NM_000737.3 | GGCTTTATACCTCGGGGTTG | |
| ERVFRD-1 | CCAAATTCCCTCCTCTCCTC | 115 |
| NM_207582.2 | CGGGTGTTAGTTTGCTTGGT | |
| GCM1 | GCAACACCAACAACCACAAC | 100 |
| NM_003643.3 | GTAAATCTTGCGGCCTTCCT | |
| OVOL1 | AGACATGGGCCACTTGACAG | 104 |
| NM_004561.3 | AGGTGAACAGGTCTCCACTG | |
| FOSL1 | GGAGGAAGGAACTGACCGACTT | 113 |
| NM_005438.5 | CTCTAGGCGCTCCTTCTGCTTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoshida, K.; Yano, A.; Kusama, K.; Ishikawa, G.; Tamura, K. Alpha 1 Antitrypsin Regulates Trophoblast Syncytialization and Inflammatory Factor Expression. Int. J. Mol. Sci. 2022, 23, 1955. https://doi.org/10.3390/ijms23041955
Yoshida K, Yano A, Kusama K, Ishikawa G, Tamura K. Alpha 1 Antitrypsin Regulates Trophoblast Syncytialization and Inflammatory Factor Expression. International Journal of Molecular Sciences. 2022; 23(4):1955. https://doi.org/10.3390/ijms23041955
Chicago/Turabian StyleYoshida, Kanoko, Aruto Yano, Kazuya Kusama, Gen Ishikawa, and Kazuhiro Tamura. 2022. "Alpha 1 Antitrypsin Regulates Trophoblast Syncytialization and Inflammatory Factor Expression" International Journal of Molecular Sciences 23, no. 4: 1955. https://doi.org/10.3390/ijms23041955
APA StyleYoshida, K., Yano, A., Kusama, K., Ishikawa, G., & Tamura, K. (2022). Alpha 1 Antitrypsin Regulates Trophoblast Syncytialization and Inflammatory Factor Expression. International Journal of Molecular Sciences, 23(4), 1955. https://doi.org/10.3390/ijms23041955