
Genome-Wide Analysis Indicates a Complete Prostaglandin Pathway from Synthesis to Inactivation in Pacific White Shrimp, Litopenaeus vannamei


 and
and 
Abstract
1. Introduction
2. Results
2.1. Screening of Genes in the Prostaglandin Pathway
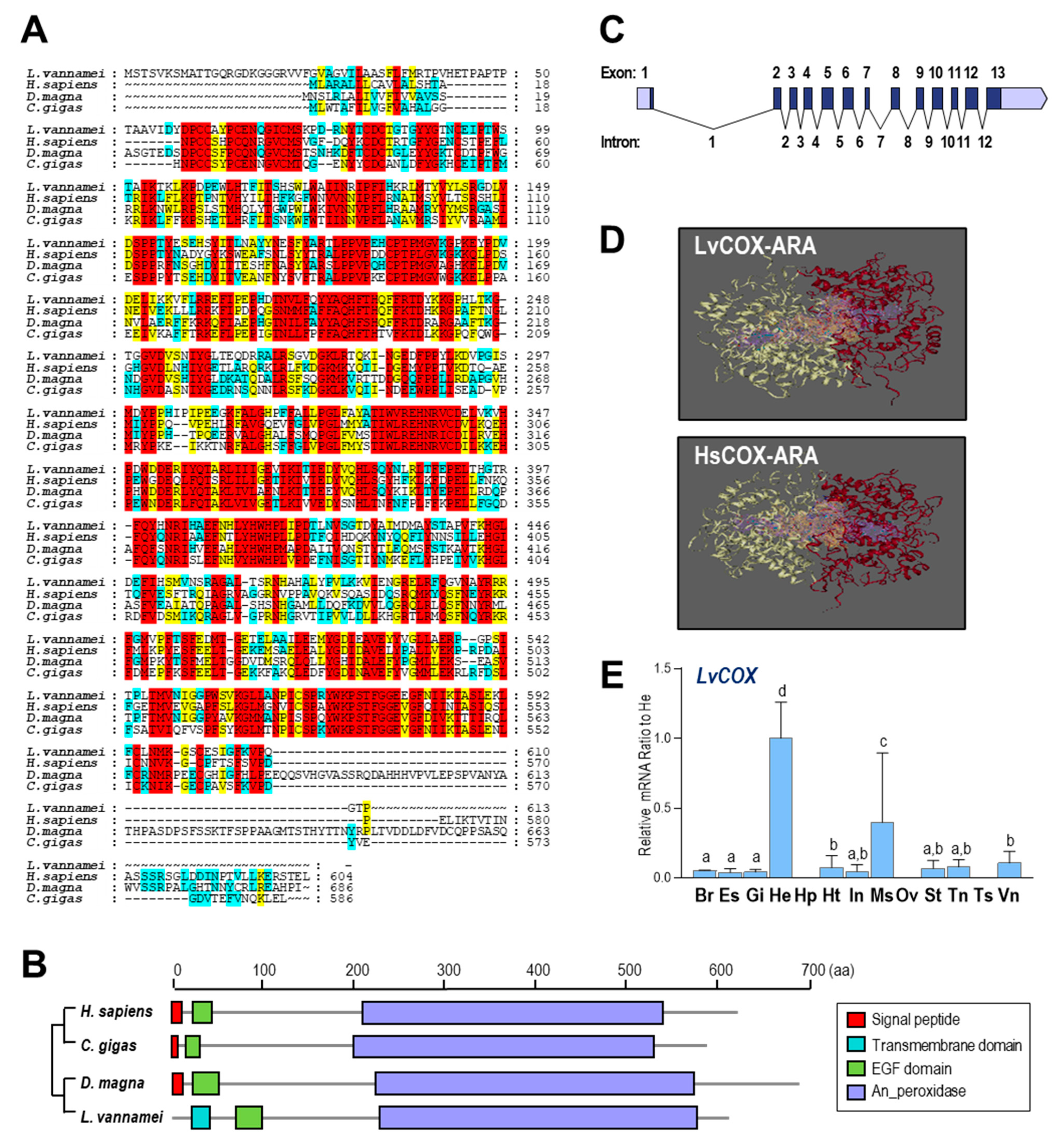
2.2. Comparative Analysis of COX in L. vannamei and Other Species
2.3. Comparative Analysis of PGSs in L. vannamei and Other Species
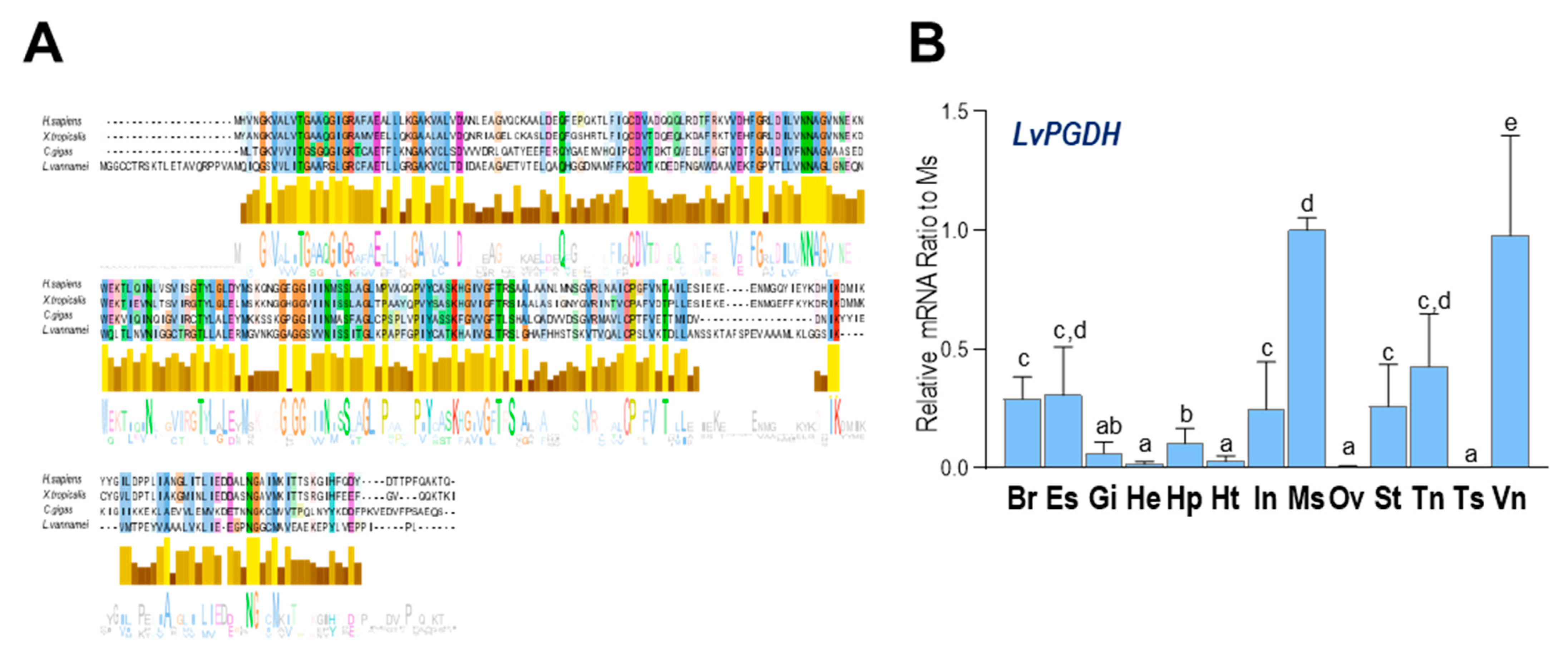
2.4. Comparative Analysis of PGDH in L. vannamei and Other Species
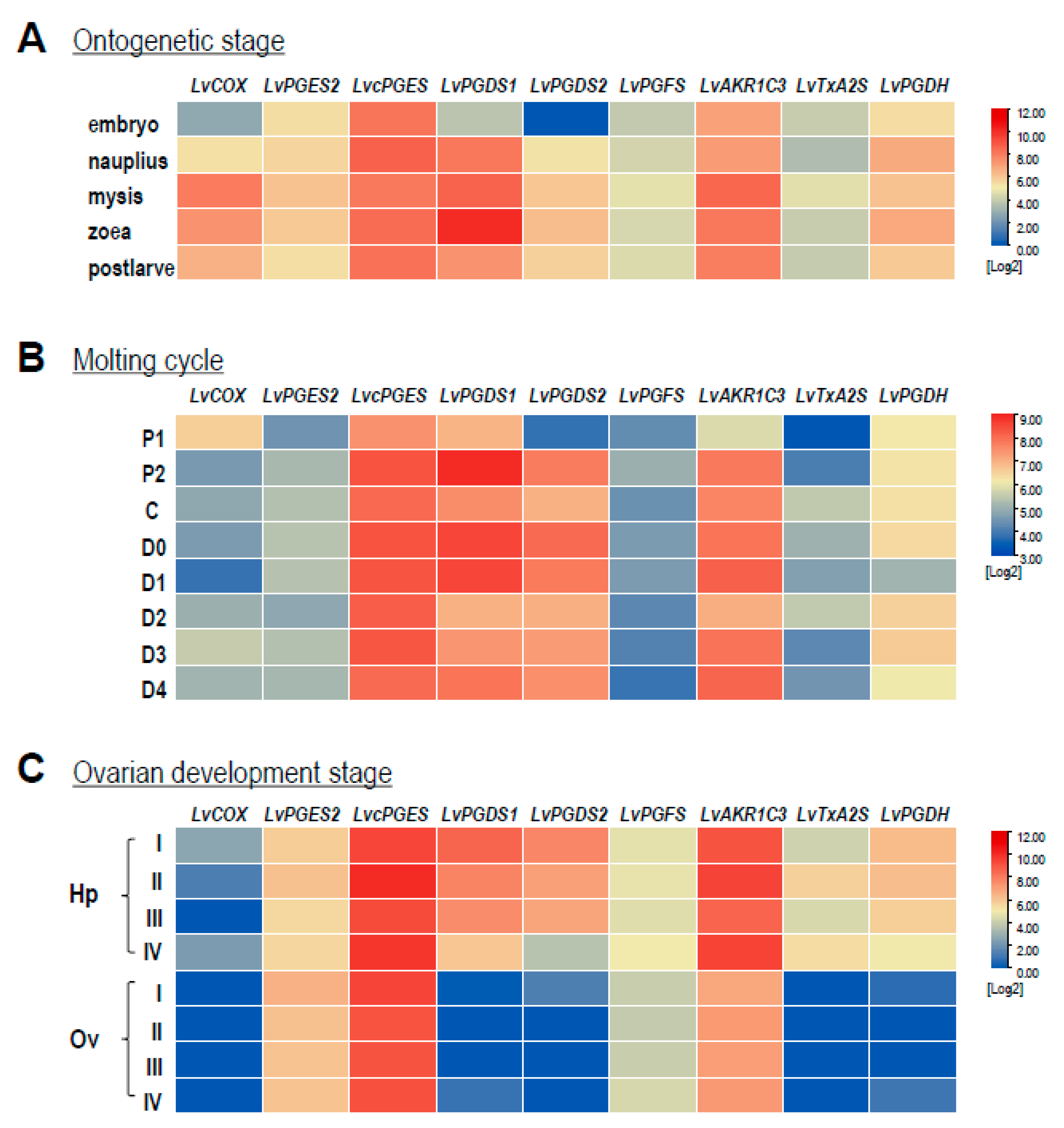
2.5. Expression Patterns during Ontogenetic Development, Ovarian Development, and Molting
3. Discussion
4. Materials and Methods
4.1. Cross-Genomic Analysis of Genes in the Prostaglandin Pathway
4.2. Phylogenetic Tree and Sequence Analysis
4.3. Structural Analysis of Proteins
4.4. Animals and Tissue Samples Collection
4.5. Realtime PCR for Tissue Distribution
4.6. Transcriptomic Analysis for Gene Expression
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| COX | cyclooxygenase |
| PGES | prostaglandin E synthase |
| PGDS | prostaglandin D synthase |
| PGFS | prostaglandin F synthase/PGH 9,11-endoperoxide reductase |
| PGIS | prostaglandin I synthase |
| AKR1C3 | PGD 11-keto reductase |
| TxA2S | thromboxane A2 synthase |
| PGDH | 15-hydroxyprostaglandin dehydrogenases |
References
- Cha, Y.I.; Solnica-Krezel, L.; DuBois, R.N. Fishing for prostanoids: Deciphering the developmental functions of cyclooxygenase-derived prostaglandins. Dev. Biol. 2006, 289, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, Y.; Narumiya, S.; Ichikawa, A. Distribution and function of prostanoid receptors: Studies from knockout mice. Prog. Lipid Res. 2000, 39, 289–314. [Google Scholar] [CrossRef]
- Funk, C.D. Prostaglandins and leukotrienes: Advances in eicosanoid biology. Science 2001, 294, 1871–1875. [Google Scholar] [CrossRef] [PubMed]
- Hata, A.N.; Breyer, R.M. Pharmacology and signaling of prostaglandin receptors: Multiple roles in inflammation and immune modulation. Pharmacol. Ther. 2004, 103, 147–166. [Google Scholar] [CrossRef] [PubMed]
- Christ, E.J.; van Dorp, D.A. Comparative aspects of prostaglandin biosynthesis in animal tissues. Biochim. Biophys. Acta 1972, 270, 534–545. [Google Scholar] [CrossRef]
- Leslie, J.B.; Watkins, W.D. Eicosanoids in the central nervous system. J. Neurosurg. 1985, 63, 659–668. [Google Scholar] [CrossRef]
- Yagami, T.; Koma, H.; Yamamoto, Y. Pathophysiological Roles of Cyclooxygenases and Prostaglandins in the Central Nervous System. Mol. Neurobiol. 2016, 53, 4754–4771. [Google Scholar] [CrossRef]
- Heeney, A.; Rogers, A.C.; Mohan, H.; Mc Dermott, F.; Baird, A.W.; Winter, D.C. Prostaglandin E2 receptors and their role in gastrointestinal motility—Potential therapeutic targets. Prostaglandins Other Lipid Mediat. 2021, 152, 106499. [Google Scholar] [CrossRef]
- Wang, D.; DuBois, R.N. Role of prostanoids in gastrointestinal cancer. J. Clin. Investig. 2018, 128, 2732–2742. [Google Scholar] [CrossRef]
- Karim, S.M.; Somers, K.; Hillier, K. Cardiovascular actions of prostaglandin Fa-alpha infusion in man. Eur. J. Pharmacol. 1969, 5, 117–120. [Google Scholar] [CrossRef]
- Sasaguri, T.; Miwa, Y. Prostaglandin J2 family and the cardiovascular system. Curr. Vasc. Pharmacol. 2004, 2, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Fan, X.; Xu, F.; Wang, J.; Chen, Y. Prostaglandin E1 attenuates post-cardiac arrest myocardial dysfunction through inhibition of mitochondria-mediated cardiomyocyte apoptosis. Mol. Med. Rep. 2021, 23, 110. [Google Scholar] [CrossRef]
- Brown, J.A.; Bucknall, R.M. Antidiuretic and cardiovascular actions of prostaglandin E2 in the rainbow trout Salmo gairdneri. Gen. Comp. Endocrinol. 1986, 61, 330–337. [Google Scholar] [CrossRef]
- Goetz, F.W. Compartmentalization of prostaglandin synthesis within the fish ovary. Am. J. Physiol. 1991, 260 Pt 2, R862–R865. [Google Scholar] [CrossRef] [PubMed]
- Stanley, D.; Kim, Y. Prostaglandins and Other Eicosanoids in Insects: Biosynthesis and Biological Actions. Front. Physiol. 2018, 9, 1927. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Kim, Y. Prostaglandin catabolism in Spodoptera exigua, a lepidopteran insect. J. Exp. Biol. 2020, 223, jeb233221. [Google Scholar] [CrossRef]
- Van der Ouderaa, F.J.; Buytenhek, M.; Nugteren, D.H.; Van Dorp, D.A. Purification and characterisation of prostaglandin endoperoxide synthetase from sheep vesicular glands. Biochim. Biophys. Acta 1977, 487, 315–331. [Google Scholar] [CrossRef]
- Urade, Y.; Watanabe, K.; Hayaishi, O. Prostaglandin d, e., and f synthases. J. Lipid Mediat. Cell Signal. 1995, 12, 257–273. [Google Scholar] [CrossRef]
- Smith, W.L.; DeWitt, D.L.; Garavito, R.M. Cyclooxygenases: Structural, cellular, and molecular biology. Annu. Rev. Biochem. 2000, 69, 145–182. [Google Scholar] [CrossRef]
- Mendes, R.T.; Stanczyk, C.P.; Sordi, R.; Otuki, M.F.; dos Santos, F.A.; Fernandes, D. Selective inhibition of cyclooxygenase-2: Risks and benefits. Rev. Bras. Reumatol. 2012, 52, 767–782. [Google Scholar]
- Pelley, J.W. 11—Metabolism of Steroids and Other Lipids. In Elsevier’s Integrated Review Biochemistry, 2nd ed.; Pelley, J.W., Ed.; W.B. Saunders: Philadelphia, PA, USA, 2012; pp. 89–98. [Google Scholar]
- Tootle, T.L.; Williams, D.; Hubb, A.; Frederick, R.; Spradling, A. Drosophila eggshell production: Identification of new genes and coordination by Pxt. PLoS ONE 2011, 6, e19943. [Google Scholar] [CrossRef] [PubMed]
- Hara, S.; Kamei, D.; Sasaki, Y.; Tanemoto, A.; Nakatani, Y.; Murakami, M. Prostaglandin E synthases: Understanding their pathophysiological roles through mouse genetic models. Biochimie 2010, 92, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Tanikawa, N.; Ohmiya, Y.; Ohkubo, H.; Hashimoto, K.; Kangawa, K.; Kojima, M.; Ito, S.; Watanabe, K. Identification and characterization of a novel type of membrane-associated prostaglandin E synthase. Biochem. Biophys. Res. Commun. 2002, 291, 884–889. [Google Scholar] [CrossRef] [PubMed]
- Urade, Y.; Hayaishi, O. Prostaglandin D synthase: Structure and function. Vitam. Horm. 2000, 58, 89–120. [Google Scholar] [PubMed]
- Watanabe, K. Prostaglandin F synthase. Prostaglandins Other Lipid Mediat. 2002, 68, 401–407. [Google Scholar] [CrossRef]
- Dai, D.; Chen, B.; Feng, Y.; Wang, W.; Jiang, Y.; Huang, H.; Liu, J. Prognostic value of prostaglandin I2 synthase and its correlation with tumor-infiltrating immune cells in lung cancer, ovarian cancer, and gastric cancer. Aging 2020, 12, 9658–9685. [Google Scholar] [CrossRef]
- Jones, D.A.; Fitzpatrick, F.A. Thromboxane A2 synthase. Modification during “suicide” inactivation. J. Biol. Chem. 1991, 266, 23510–23514. [Google Scholar] [CrossRef]
- Catella, F.; Healy, D.; Lawson, J.A.; FitzGerald, G.A. 11-Dehydrothromboxane B2: A quantitative index of thromboxane A2 formation in the human circulation. Proc. Natl. Acad. Sci. USA 1986, 83, 5861–5865. [Google Scholar] [CrossRef] [PubMed]
- Hsin-Hsiung, T.; Charles, M.E.; Min, T. Prostaglandin catabolizing enzymes. Prostaglandins Other Lipid Mediat. 2002, 68, 483–493. [Google Scholar]
- Ensor, C.M.; Tai, H.H. 15-Hydroxyprostaglandin dehydrogenase. J. Lipid Mediat Cell Signal. 1995, 12, 313–319. [Google Scholar] [CrossRef]
- Luo, X.; Chen, T.; Zhong, M.; Jiang, X.; Zhang, L.; Ren, C.; Hu, C. Differential regulation of hepatopancreatic vitellogenin (VTG) gene expression by two putative molt-inhibiting hormones (MIH1/2) in Pacific white shrimp (Litopenaeus vannamei). Peptides 2015, 68, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Ren, C.; Wang, Y.; Gao, Y.; Wong, N.-K.; Zhang, L.; Hu, C. Crustacean cardioactive peptide (CCAP) of the Pacific white shrimp (Litopenaeus vannamei): Molecular characterization and its potential roles in osmoregulation and freshwater tolerance. Aquaculture 2016, 451, 405–412. [Google Scholar] [CrossRef]
- Chen, T.; Ren, C.; Jiang, X.; Zhang, L.; Li, H.; Huang, W.; Hu, C. Mechanisms for type-II vitellogenesis-inhibiting hormone suppression of vitellogenin transcription in shrimp hepatopancreas: Crosstalk of GC/cGMP pathway with different MAPK-dependent cascades. PLoS ONE 2018, 13, e0194459. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Shi, W.; Lin, D.; Ye, H. A Possible Role of Allatostatin C in Inhibiting Ecdysone Biosynthesis Revealed in the Mud Crab Scylla Paramamosain. Front. Mar. Sci. 2021, 8, 1293. [Google Scholar] [CrossRef]
- Liu, J.; Liu, A.; Liu, F.; Huang, H.; Ye, H. Role of neuroparsin 1 in vitellogenesis in the mud crab, Scylla paramamosain. Gen. Comp. Endocrinol. 2020, 285, 113248. [Google Scholar] [CrossRef]
- Pan, J.; Liu, M.; Chen, T.; Cheng, Y.; Wu, X. Immunolocalization and changes of 17beta-estradiol during ovarian development of Chinese mitten crab Eriocheir sinensis. Cell Tissue Res. 2018, 373, 509–520. [Google Scholar] [CrossRef]
- Tahara, D.; Yano, I. Maturation-related variations in prostaglandin and fatty acid content of ovary in the kuruma prawn (Marsupenaeus japonicus). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2004, 137, 631–637. [Google Scholar] [CrossRef]
- Sumpownon, C.; Engsusophon, A.; Siangcham, T.; Sugiyama, E.; Soonklang, N.; Meeratana, P.; Wanichanon, C.; Hanna, P.J.; Setou, M.; Sobhon, P. Variation of prostaglandin E2 concentrations in ovaries and its effects on ovarian maturation and oocyte proliferation in the giant fresh water prawn, Macrobrachium rosenbergii. Gen. Comp. Endocrinol. 2015, 223, 129–138. [Google Scholar] [CrossRef]
- Tahara, D.; Yano, I. Development of hemolymph prostaglandins assay systems and their concentration variations during ovarian maturation in the kuruma prawn, Penaeus japonicus. Aquaculture 2003, 220, 791–800. [Google Scholar] [CrossRef]
- Duangprom, S.; Ampansri, W.; Suwansa-Ard, S.; Chotwiwatthanakun, C.; Sobhon, P.; Kornthong, N. Identification and expression of prostaglandin E synthase (PGES) gene in the central nervous system and ovary during ovarian maturation of the female mud crab, Scylla olivacea. Anim. Reprod. Sci. 2018, 198, 220–232. [Google Scholar] [CrossRef]
- Swetha, C.H.; Girish, B.P.; Hemalatha, M.; Reddy, P.S. Induction of vitellogenesis, methyl farnesoate synthesis and ecdysteroidogenesis in two edible crabs by arachidonic acid and prostaglandins. J. Exp. Biol. 2020, 223 Pt 3, jeb212381. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Chen, T.; Luo, P.; Huang, W.; Huo, D.; Yun, L.; Hu, C.; Cheng, C. A fibrinogen-related protein, LvFREP2, from Litopenaeus vannamei facilitates the clearance of Vibrio harveyi. Fish Shellfish Immunol. 2018, 78, 364–371. [Google Scholar] [CrossRef]
- Chen, T.; Lin, T.; Li, H.; Lu, T.; Li, J.; Huang, W.; Sun, H.; Jiang, X.; Zhang, J.; Yan, A.; et al. Heat Shock Protein 40 (HSP40) in Pacific White Shrimp (Litopenaeus vannamei): Molecular Cloning, Tissue Distribution and Ontogeny, Response to Temperature, Acidity/Alkalinity and Salinity Stresses, and Potential Role in Ovarian Development. Front. Physiol. 2018, 9, 1784. [Google Scholar] [CrossRef] [PubMed]
- Meunpol, O.; Duangjai, E.; Yoonpun, R.; Piyatiratitivorakul, S. Detection of prostaglandin E2 in polychaete Perinereis sp. and its effect on Penaeus monodon oocyte development in vitro. Fish. Sci. 2010, 76, 281–286. [Google Scholar] [CrossRef]
- Vane, J.R.; Bakhle, Y.S.; Botting, R.M. Cyclooxygenases 1 and 2. Annu. Rev. Pharmacol. Toxicol. 1998, 38, 97–120. [Google Scholar] [CrossRef] [PubMed]
- Otto, J.C.; Smith, W.L. Prostaglandin endoperoxide synthases-1 and -2. J. Lipid Mediat. Cell Signal. 1995, 12, 139–156. [Google Scholar] [CrossRef]
- DeWitt, D.L.; Day, J.S.; Sonnenburg, W.K.; Smith, W.L. Concentrations of prostaglandin endoperoxide synthase and prostaglandin I2 synthase in the endothelium and smooth muscle of bovine aorta. J. Clin. Investig. 1983, 72, 1882–1888. [Google Scholar] [CrossRef]
- Hatae, T.; Hara, S.; Yokoyama, C.; Yabuki, T.; Tanabe, T.I.J.F.L. Site-directed mutagenesis of human prostacyclin synthase: Alteration of Cys441 of the Cys-pocket, and Glu347 and Arg350 of the EXXR motif. FEBS Lett. 1996, 389, 268–272. [Google Scholar] [CrossRef]
- Pritchard, K.J.J.J.B.C. Induction of cyclooxygenase-2 in rat vascular smooth muscle cells in vitro and in vivo. J. Biol. Chem. 1994, 269, 8504–8509. [Google Scholar] [CrossRef]
- Ruan, Y.; Wong, N.K.; Zhang, X.; Zhu, C.; Wu, X.; Ren, C.; Luo, P.; Jiang, X.; Ji, J.; Wu, X.; et al. Vitellogenin Receptor (VgR) Mediates Oocyte Maturation and Ovarian Development in the Pacific White Shrimp (Litopenaeus vannamei). Front. Physiol. 2020, 11, 485. [Google Scholar] [CrossRef]
- Murakami, M.; Naraba, H.; Tanioka, T.; Semmyo, N.; Nakatani, Y.; Kojima, F.; Ikeda, T.; Fueki, M.; Ueno, A.; Oh, S.; et al. Regulation of prostaglandin E2 biosynthesis by inducible membrane-associated prostaglandin E2 synthase that acts in concert with cyclooxygenase-2. J. Biol. Chem. 2000, 275, 32783–32792. [Google Scholar] [CrossRef] [PubMed]
- Tanioka, T.; Nakatani, Y.; Semmyo, N.; Murakami, M.; Kudo, I. Molecular identification of cytosolic prostaglandin E2 synthase that is functionally coupled with cyclooxygenase-1 in immediate prostaglandin E2 biosynthesis. J. Biol. Chem. 2000, 275, 32775–32782. [Google Scholar] [CrossRef] [PubMed]
- Urade, Y.; Eguchi, N. Lipocalin-type and hematopoietic prostaglandin D synthases as a novel example of functional convergence. Prostaglandins Other Lipid Mediat. 2002, 68–69, 375–382. [Google Scholar] [CrossRef]
- Christ-Hazelhof, E.J.B.B.A. Purification and characterisation of prostaglandin endoperoxide D-isomerase, a cytoplasmic, glutathione-requiring enzyme. Biochim. Biophys. Acta (BBA) Lipids Lipid Metab. 1979, 572, 43–51. [Google Scholar] [CrossRef]
- Urade, Y.; Fujimoto, N.; Ujihara, M.; Hayaishi, O. Biochemical and immunological characterization of rat spleen prostaglandin D synthetase. J. Biol. Chem. 1987, 262, 3820–3825. [Google Scholar] [CrossRef]
- Urade, Y.; Ujihara, M.; Horiguchi, Y.; Igarashi, M.; Nagata, A.; Ikai, K.; Hayaishi, O. Mast cells contain spleen-type prostaglandin D synthetase. J. Biol. Chem. 1990, 265, 371–375. [Google Scholar] [CrossRef]
- Hansen, H.S.J.P. 15-Hydroxyprostaglandin dehydrogenase. A review. Prostaglandins 1976, 12, 647–679. [Google Scholar] [CrossRef]
- Persson, B.; Krook, M.; Jornvall, H. Characteristics of short-chain alcohol dehydrogenases and related enzymes. Eur. J. Biochem. 1991, 200, 537–543. [Google Scholar] [CrossRef]
- Lister, A.L.; Van Der Kraak, G. An investigation into the role of prostaglandins in zebrafish oocyte maturation and ovulation. Gen. Comp. Endocrinol. 2008, 159, 46–57. [Google Scholar] [CrossRef]
- Ruggeri, B.; Thoroughgood, C.A. Prostaglandins in aquatic fauna: A comprehensive review. Mar. Ecol. Prog. Ser. 1985, 23, 301–306. [Google Scholar] [CrossRef]
- Boeckmann, B.; Bairoch, A.; Apweiler, R.; Blatter, M.C.; Estreicher, A.; Gasteiger, E.; Martin, M.J.; Michoud, K.; O’Donovan, C.; Phan, I.; et al. The SWISS-PROT protein knowledgebase and its supplement TrEMBL in 2003. Nucleic Acids Res. 2003, 31, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Quevillon, E.; Silventoinen, V.; Pillai, S.; Harte, N.; Mulder, N.; Apweiler, R.; Lopez, R. InterProScan: Protein domains identifier. Nucleic Acids Res. 2005, 33, W116–W120. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Wei, J.; Zhang, X.; Yu, Y.; Huang, H.; Li, F.; Xiang, J. Comparative transcriptomic characterization of the early development in Pacific white shrimp Litopenaeus vannamei. PLoS ONE 2014, 9, e106201. [Google Scholar]
- Gao, Y.; Zhang, X.; Wei, J.; Sun, X.; Yuan, J.; Li, F.; Xiang, J. Whole Transcriptome Analysis Provides Insights into Molecular Mechanisms for Molting in Litopenaeus vannamei. PLoS ONE 2015, 10, e0144350. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein ID | Annotation |
|---|---|
| XP027210685.1 | Phospholipase A2 |
| XP027215735.1 | Phospholipase A2 |
| XP027216925.1 | Phospholipase A2 |
| XP027224729.1 | Phospholipase A2 |
| XP027229029.1 | Phospholipase A2 |
| XP027230031.1 | Phospholipase A2 |
| XP027239642.1 | Phospholipase A2 |
| XP027218437.1 | Cyclooxygenase/Prostaglandin G/H synthase |
| XP027219753.1 | Prostaglandin E synthase |
| XP027224753.1 | Prostaglandin E synthase |
| XP027210104.1 | Prostaglandin E synthase |
| XP027217033.1 | prostaglandin D synthase |
| XP027217035.1 | prostaglandin D synthase |
| XP027216546.1 | Prostamide/prostaglandin F synthase |
| XP027237263.1 | 9,11-endoperoxide prostaglandin H2 reductase |
| Gene Name. | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Product (bp) |
|---|---|---|---|
| LvCOX | ATGTCAACGTCTGTGAAAAGCATGG | CCATGCTTTTCACAGACGTTGACAT | 174 |
| LvPGES2 | CTCTGAGGGCAGCGAGTA | GCAACAGAATGGGCAAGT | 133 |
| LvPGES3 | CAGCACTGGCTCAAAGTA | GTCGGAATCTTCATCCTC | 168 |
| LvPGDS1 | TGGCAGACAAGGACGAGG | GCCGAGGCTGATGATGAA | 162 |
| LvPGDS2 | GATGTCTTGGGTCGATCTGA | AAGGTGTCTGGGCGTCTTT | 159 |
| LvPGFS | CGGAATGAAACTGAAATAGG | GAGGTAGGTCAGCTGCAGGTTACTT | 178 |
| LvAKR1C3 | GAGGCTCATTGGCATCGG | AATCATCGCAAGAACACC | 145 |
| LvTxA2S1 | CGGAATGAAACTGAAATAGG | GGTAAAGGTCGAGGTAGGT | 178 |
| LvPGDH | CTGACCTCCTCGCCAACTCC | AGGGCTCTTTCTCCGCCTC | 180 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Chen, X.; Li, Z.; Wu, X.; Zhou, M.; Zhang, X.; Liu, Y.; Sun, Y.; Zhu, C.; Guo, Q.; et al. Genome-Wide Analysis Indicates a Complete Prostaglandin Pathway from Synthesis to Inactivation in Pacific White Shrimp, Litopenaeus vannamei. Int. J. Mol. Sci. 2022, 23, 1654. https://doi.org/10.3390/ijms23031654
Yang H, Chen X, Li Z, Wu X, Zhou M, Zhang X, Liu Y, Sun Y, Zhu C, Guo Q, et al. Genome-Wide Analysis Indicates a Complete Prostaglandin Pathway from Synthesis to Inactivation in Pacific White Shrimp, Litopenaeus vannamei. International Journal of Molecular Sciences. 2022; 23(3):1654. https://doi.org/10.3390/ijms23031654
Chicago/Turabian StyleYang, Hao, Xiaoli Chen, Zhi Li, Xugan Wu, Mingyu Zhou, Xin Zhang, Yujie Liu, Yuying Sun, Chunhua Zhu, Qiuhui Guo, and et al. 2022. "Genome-Wide Analysis Indicates a Complete Prostaglandin Pathway from Synthesis to Inactivation in Pacific White Shrimp, Litopenaeus vannamei" International Journal of Molecular Sciences 23, no. 3: 1654. https://doi.org/10.3390/ijms23031654
APA StyleYang, H., Chen, X., Li, Z., Wu, X., Zhou, M., Zhang, X., Liu, Y., Sun, Y., Zhu, C., Guo, Q., Chen, T., & Zhang, J. (2022). Genome-Wide Analysis Indicates a Complete Prostaglandin Pathway from Synthesis to Inactivation in Pacific White Shrimp, Litopenaeus vannamei. International Journal of Molecular Sciences, 23(3), 1654. https://doi.org/10.3390/ijms23031654