Abstract
Plant xyloglucan:xyloglucosyl transferases, known as xyloglucan endo-transglycosylases (XETs) are the key players that underlie plant cell wall dynamics and mechanics. These fundamental roles are central for the assembly and modifications of cell walls during embryogenesis, vegetative and reproductive growth, and adaptations to living environments under biotic and abiotic (environmental) stresses. XET enzymes (EC 2.4.1.207) have the β-sandwich architecture and the β-jelly-roll topology, and are classified in the glycoside hydrolase family 16 based on their evolutionary history. XET enzymes catalyse transglycosylation reactions with xyloglucan (XG)-derived and other than XG-derived donors and acceptors, and this poly-specificity originates from the structural plasticity and evolutionary diversification that has evolved through expansion and duplication. In phyletic groups, XETs form the gene families that are differentially expressed in organs and tissues in time- and space-dependent manners, and in response to environmental conditions. Here, we examine higher plant XET enzymes and dissect how their exclusively carbohydrate-linked transglycosylation catalytic function inter-connects complex plant cell wall components. Further, we discuss progress in technologies that advance the knowledge of plant cell walls and how this knowledge defines the roles of XETs. We construe that the broad specificity of the plant XETs underscores their roles in continuous cell wall restructuring and re-modelling.
1. Introduction
This review is presented in the three inter-connected sections that inform on: plant cell walls (CW) as multi-composite hydrogels, including their composition, structure and organisation (Section 2) and plant xyloglucan:xyloglucosyl transferases and their nomenclature, classification, catalytic mechanisms, structural properties and substrate specificity with xyloglucan (XG) and other than XG-derived donor and acceptor substrates (Section 3). In the last Section 4, we focus on plant CW modifications and re-modelling, specifically on the function of xyloglucan:xyloglucosyl transferases in CW re-modelling and dynamics, notwithstanding methods of investigation. We also explore recent progress made in technologies that advance the knowledge of CWs and how this knowledge defines the function of these transferases. The additional goal of this review is to examine the roles of the xyloglucan:xyloglucosyl transferase family in plant CWs and environmental contexts, and not as isolated catalytic entities. We expect that the detailed definition of catalytic properties of these enzymes will contribute to the clarification of the biological function of xyloglucan:xyloglucosyl transferases, in the framework of the recent models of primary and secondary plant CW architectures.
2. Plant Cell Walls Are Multi-Composite Hydrogels
(a) Composition of plant CWs—Plant CWs are multi-composite hydrogels that are predominantly comprised of polysaccharides linked by covalent and non-covalent linkages, which collectively define the properties of CWs [1,2,3]. Other important constituents of plant CWs are proteins including no-catalytic expansins, and organic and inorganic compounds, including non-fermentable polyphenolic lignins. The properties of plant CWs crucially depend on the characteristics of their building blocks, chemical linkages of components, and spatial disposition. Plant CWs have certain common characteristics regardless of their phylogenetic origin, and variations exist between species, organs, tissues, developmental stages during embryogenesis, vegetative and reproductive growth, adaptations to living environments, and biotic and abiotic stresses [4].
Plants form two major types of CWs, termed (i) primary and (ii) secondary CWs. These two major types reflect the chemical composition and structure that fulfil fundamental roles in mono- and di-cotyledonous species. Primary CWs typically contain the networks of cellulose, made of (1,4)-β-d-glucan-linked chains forming micro-fibrils tightly bound via hydrogen bonds. These supra-molecular cellulosic structures form the backbone of primary CWs. Cellulose, as the most abundant polymer, is found in CWs of higher plants, red, brown and green algae, and other species such as Oomycetes, Ameobozoa and Cyanobacteria [4]. Cellulose micro-fibrils are tethered by cross-linking hemicelluloses [5] and embedded in most dicotyledonous plants in the matrix of pectins or glycoproteins, which through hydrogen bonds and other interactions provide the frameworks to activate the CW assembly [6,7]. These hemicelluloses include XGs, which are the major form of cross-linking glycans in dicotyledonous plants, while xylans (arabinoxylans, glucuronoarabinoxylans), mannans (galactomannans, glucomannans, galactoglucomannans) and (1,3;1,4)-β-d-glucans (mixed linkage or MLG glucans) substitute XGs in monocotyledonous and lower plants [8]. Pectins composed of the homogalacturonan main chains could be calcium-cross-linked [9], while branched pectin regions are made of rhamnogalacturonan I and rhamnogalacturonan II [10,11].
Conversely, secondary CWs that are characteristic of woody plants and vascular tissues after their growth ceases, and are more rigid in comparison to the primary CWs, as they contain fewer pectins, and are reinforced by hydrophobic phenylpropanoid lignin polymers [12,13,14]. Wood formation occurs via radial proliferation of vascular cambium cells and the deposition of secondary CW layers [15]. This is critical for the mechanical stability of woody plants as it affects the chemical, mechanical and elastic properties of wood and ligno-cellulosic materials produced from wood [16].
As defined, cellulose micro-fibrils are tethered by the cross-linking hemicelluloses, such as XGs that are the most abundant hemicelluloses in the primary CWs of dicotyledonous plants. The biosynthesis of XGs [17] including green algae [18] was described in plants, while XGs were not found in red and brown algae [4], which is also accompanied by the lack of XG-modifying enzymes [19]. XG forms the chains of repeating (1,4)-β-d-linked glucopyranosyl residues, with the C-6 carbon carrying α-d-xylopyranosyl residues [20], which could be substituted by galactopyranosyl residues on the C-2 carbons (β-d-Galp-(1,2)-α-d-Xylp) and the galactosyl residues that on the C-carbons carry fucopyranosyl substitutions (α-l-Fucp-(1,2)-β-d-Galp-(1,2)-α-d-Xylp) (Figure 1), although, in moss and liverwort XGs, arabinopyranosyl [21] and galacturonate [22] substituents are found instead the fucopyranosyl residues. This suggests that the structure of XGs differs amongst plants, organs and tissues even of the same plant [23]. The XG backbone is synthesised, using UDP-glucose, by XG:glucan synthases encoded by the C subfamily of cellulose synthases-like genes [17,24], implying that the activity of an additional enzyme using UDP-xylose, namely an XG:xylosyltransferase is needed to generate XGs [25,26]. d-Xyl residues are additionally modified with d-Gal, and this decoration is mediated by XG:galactosyltransferases, and d-Gal moieties could in some XGs carry l-fucosyl moieties, which is catalysed by XG:fucosyltransferases [26,27]. Enzymes in each XG biosynthetic step occur in multiple isoforms with a discrete substrate specificity, as it was described for the XG:xylosyltransferase from Arabidopsis thaliana [26].
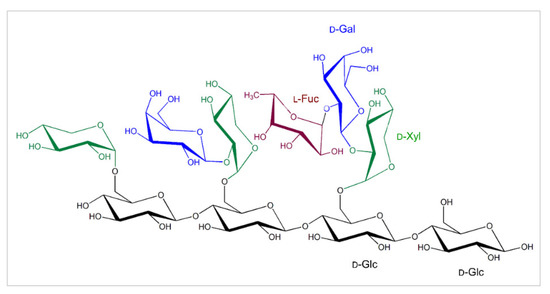
Figure 1.
The common substitutions in the tetragluco-oligosaccharide unit of XGs that forms chains of repeating (1,4)-β-d-linked glucopyranosyl residues (black), with the C-6 carbons carrying α-d-xylopyranosyl moieties (green), which could be substituted by galactopyranosyl residues (blue) on the C-2 carbons (β-d-Galp-(1,2)-α-d-Xylp). The galactopyranosyl residues could carry fucopyranosyl residues (brown) on the C-2 carbons (α-l-Fucp-(1,2)-β-d-Galp-(1,2)-α-d-Xylp). Descriptions of sugar moieties are indicated in matching colours.
(b) Plant cell wall structure and organisation—The chemical and structural parallels between cellulose and XGs underlie the conformational homology, which results in noncovalent interactions [27]. The networks of cellulose and XGs support the structure of primary CWs, which underlines their flexibility and strength [28,29]. The most recognised primary CW model of dicotyledonous plants [30] is based on the linear micro-fibrils of cellulose, each consisting of 32 micro-fibrils associated via hydrogen bonds, where para-crystalline cellulose micro-fibrils are interwoven and bridged by XGs. Consistent with this model is the function of pectin, which is attributed to the function of a gelling milieu that permeates the space between cellulosic and XG polymers.
Microscopic techniques allowed the development of sophisticated CW models that are based on cellulose micro-fibrils, composed of eight or sixteen associating cellulose sub-units [31]. These hot spot micro-fibrils are linked with each other by a thin layer of XG polysaccharides [32], where pectins fulfil the role of the filling materials between cellulosic and XG polymers. Various pectins that make close contacts with both cellulose and XGs, ensure the flexibility but also the strength of CWs [33,34]. The processes of these hot spots formations are unknown, but it is assumed that they could be formed ad hoc or with the participation of enzymes [34].
While sufficient information is available on the role of XGs in CW modifications during plant growth, no information is accessible on the participation of XGs in CW formation. Recent work with the xxt1 xxt2 mutant lacking XGs suggested that the formation of cellulose networks could be XG-independent [35]. As for the necessity of XG for CW regeneration in plant protoplasts, it was established that XG is non-essential for the formation of the cellulose networks and that the cellulose networks formed in the absence of XGs provide sufficient tensile strength to the primary CW in Arabidopsis protoplasts [31].
As alluded to above, CWs are diverse, which underscores the function of cells, and this could be directly linked to the elusive structural changes of polymeric components, their quantity, ratios, and underpins their interactions. The syntheses of CW polysaccharides occur due to the cooperative activities of various glycosyl transferases (GTs) or synthases [17,36,37,38], operating usually in the Golgi apparatus and associated vesicles [39]. These enzymes are transported to CWs by secretory vesicles, but other enzymes, such as cellulose synthases [40,41] and callose synthases [42] locate directly to the plasma membranes [43], meaning that their polysaccharide products are exported to CWs, although the latter structural polysaccharides are often heterogeneous. It is assumed that a wide range of activated sugar donors required for the GTs activities in isomeric forms could give rise to a variety of glycosidic linkages.
As the life processes in plants are guided by CW organisation, involving elongation, expansion and loosening, in addition to GTs and synthases, these processes are also governed by non-catalytic expansins [44,45], and enzymes with hydrolytic, esterase and lyase (non-hydrolytic addition or removal of groups from substrates) activities. There is a large number of these enzymes that could potentially modify the structural polysaccharides in situ by breaking linkages, esterifying or de-esterifying saccharides [46], and incorporating new saccharide components in CWs or re-constructing polymers through cross-linking. The latter processes are mediated by xyloglucan:xyloglucosyl transferases also known as xyloglucan endo-transglycosylases (XETs) [47,48], whereby these enzymes are one of the key glycosidic bond-formation biocatalysts taking part in plant CW expansion, modification, reconstruction and re-modelling.
3. Plant Xyloglucan:Xyloglucosyl Transferases
Xyloglucan:xyloglucosyl transferases or xyloglucan endo-transglycosylases (XET), classified under EC 2.4.1.207, transfer the glycosyl groups from one glycoside to another. These enzymes were discovered in 1992 independently in bean epicotyls [49], nasturtium seeds [50], and pea, tomato and other plant extracts [51], and since their discovery, significant knowledge has been accumulated on their mode of action.
(a) Nomenclatureand classification—The nomenclature of XET enzymes is defined by the International Union of Biochemistry and Molecular Biology (IUBMB) and International Union of Pure and Applied Chemistry Biochemical Nomenclature Committee [52], the Kyoto Encyclopaedia of Genes and Genomes Enzyme Database, and the BRENDA Comprehensive Enzyme System [53]. These nomenclatures either consider the transfer of the ‘glycosyl’ (xyloglucan endo-transglycosylase) [49,50,51] or ‘glucosyl’ (xyloglucan endo-transglucosylase) [54,55] groups.
The complex view on XET enzyme nomenclature and classification is provided by the Carbohydrate-Active enZYmes Database (CAZy) [56] and CAZypedia [57], which classify entries based on protein tertiary structures, substrate specificities and evolutionary history or phylogenomic relationships. XETs are listed in the glycoside hydrolase (GH) family 16, and not in a glycoside transferase (GT) Leloir group of enzymes. According to the Enzyme Commission and International Union of Biochemistry and Molecular Biology, the latter group involves enzymes that use activated sugars (for example UDP-glucose) as the glycosyl donors, and transfer the glycosyl groups to the nucleophilic glycoside (or other) acceptors. The GH16 family, according to tertiary structural features is sub-divided into 23 subfamilies [58], where the subfamily GH16_20 includes XETs but also xyloglucan endohydrolases (XEHs, EC 3.2.1.151) [59]; the latter group catalyses predominantly hydrolytic reactions of XGs. Thus, the GH16_20 group contains the products of XTH (xyloglucan transglycosylase/hydrolase) genes encoding both types of XG-modifying enzymes which notably display close similarities in tertiary structures [60].
(b) Catalytic mechanism—The catalytic function of XETs is defined by breaking a glycosidic bond between 1,4-β-d-linked glucosyl residues of XGs (or other polysaccharides) and transferring the xyloglucanyl or another glycoside moiety onto the O-4 atom of a non-reducing end of a glycoside acceptor; this acceptor could be XG, XG-oligosaccharide or another glycoside. This constitutes a so-called ping-pong bi-bi reaction mechanism and not a sequential one [61,62]. Recently, the definition of XET substrate specificity has significantly expanded, and thus the previous information on XETs has become obsolete [47,48,63,64,65,66,67,68,69].
The first steps of the transglycosylation and hydrolytic reactions catalysed by XTHs (that is XETs and XEHs) are binding and cleavage of donor glycoside substrates. The difference between XETs and XEHs occurs in the second step, in which the fragment with the original non-reducing end of the donor substrate is transferred to an acceptor, which in the case of a typical transglycosylase is a saccharide, while in the case of a hydrolase, it is an activated water molecule [56,70]. This second step had the key importance for the nomenclature and classification of XETs as transferases.
The transglycosylation mechanism catalysed by XET enzymes proceeds in two stages with two transition states. The first step involves the deprotonation of a carboxylate that operates as a nucleophile and attacks the anomeric carbon of the donor substrate, forming the glycosyl–enzyme intermediate complex, with the participation of an acid/base carboxylate. In the hybrid aspen Populus tremulus x tremuloides PttXET16A, the nucleophile attacking the anomeric carbon is Glu85, while Glu89 acts as an acid/base (has a dual role), that is it protonates the aglycon saccharide and releases it, and subsequently, it de-protonates the glycoside acceptor. The third important residue in PttXET16A is Asp87 positioned between Glu85 and Glu89 (signature motif ExDxE), which forms a tight bond with the Glu85 nucleophile, and interacts with the substrates through hydrogen bonds during substrate binding. The nucleophile must be de-protonated for the donor substrate attack, while Asp87 and Glu89 remain protonated. The preference for the transfer reaction instead of hydrolytic one by XETs leads to the re-ligation of the nascent donor end of one saccharide to the non-reducing end of the acceptor substrate [60,71]. Curiously, in the crystal structure of PttXET16A (‘true’ XET without hydrolytic activity) [71], only a few water molecules were resolved near the donor and acceptor substrate binding sites, while significantly more water molecules were located around the corresponding sites in XEHs.
Typically, hydrolases also transglycosylate under high(er) substrate concentrations [70,72], when in the later stages of reactions, the glycosyl–enzyme intermediate complex interacts with another sugar molecule (instead of an activated water molecule), thus shifting the reaction equilibrium towards the transfer products. Contrary to these high-substrate-induced transglycosylation reactions, ‘true’ transglycosylases such as XETs encounter a second saccharide regardless of a substrate concentration. While transglycosylation reactions catalysed by hydrolases occur under high(er) concentrations of substrates and generated products, this is not the case of XETs.
In addition to XETs, other plant enzymes with transglycosylase activities were reported, often without providing protein sequence information, with trivial descriptions such as mannan endotransglycosylase/hydrolases recognising (1,4)-β-d-mannan-derived polysaccharides [73], xylan endo-transglycosylases functionalising heteroxylan polysaccharides [74,75,76], mixed-linkage glucan:xyloglucan endotransglucosylases acting on (1,3;1,4)-β-d-glucans [77,78], and hetero-trans-β-glucanases (HTGs) functionalising cellulose [48,79]. However, terming these enzymes as such disguises a fact that some of them could in fact be non-specific XETs.
(c) Structural properties, evolutionary relationships, enzyme activity methods—The crystal structures of the XET enzyme PttXET16A (Protein Data Bank—PDB accessions and 1UMZ and 1UN1 at respective 1.80 Å and 2.10 Å resolution; 1UMZ in complex with the XLLG acceptor) [60,71,80] offers insights into its atomic architecture and catalytic mechanism. The crystal structure revealed that PttXET16A has the β-jelly-roll topology and folds into a β-sandwich with convex and concave regions of the two antiparallel β-sheets (Figure 2). The catalytic trio, formed by three Glu85, Glu89 and Asp87 residues, is located around the middle of the convex region of the structure (Figure 2A; magenta cpk sticks). The elongated C-terminus is folded in an α-helix and a β-strand on the concave side of the molecule and stabilised by a disulfide bridge. The PttXET16A structure is also glycosylated at Asn93 with two N-acetylglucosaminyl and one mannosyl residues that are further stabilised by the hydrogen bonds. No other atomic XET structures have been reported; however, structural features of the 3D models of nasturtium, barley (Figure 2B, Figure 3 and Figure 4), and other XETs were defined [64,66,68,69,79,81]. Although, caution should be exercised when extracting and interpreting structural information based on 3D models, where side-chain placements are known to be unreliable even at high similarities between templates and target sequences [82].
Additional information on the PttXET16A enzyme was obtained through molecular dynamics (MD) simulations [60], where the interactions between residues of the PttXET16A and two XLLG nonasaccharides revealed that one XLLG occupied the donor site and created a stable intermediate with the enzyme, while the second XLLG remained bound at the acceptor site. Notably, the reducing-end glucose moiety of XLLG at the donor -1 subsite, altered its low energy 4C1 conformation into 1S3 boat.
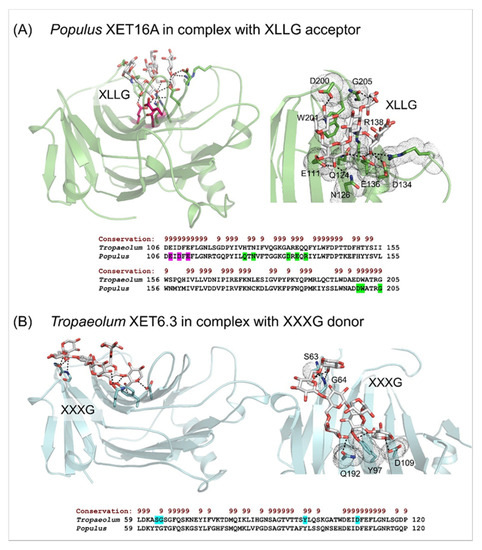
Figure 2.
β-Sandwich architectures and β-jelly-roll topologies of Populus XET16A (PDB accession 1UMZ) in complex with the XLLG acceptor substrate (A), and the Tropaeolum XET6.3 3D molecular model in complex with the XXXG donor substrate (B). (A) Left panel: The XLLG acceptor (cpk sticks) bound in the Populus XET16A structure is indicated in dashed lines at 2.6 Å to 3.5 Å separations. The catalytic trio (Glu85, Glu89 and Asp87) is shown in cpk magenta sticks. Right panel: Details of the XLLG binding in Populus XET16A; interacting residues are marked in green cpk sticks and dots. Bottom panel: Some of the residues of Populues XET16A binding XLLG (highlighted in green) are shown in the alignment by PROMALS3D [83] of the Populus and Tropaeolum XET sequences (numbering includes signal peptides). Conservation of residues on the scale 9–6 is shown on the top of the alignment in brown. (B) Left panel: The XXXG donor (cpk sticks) bound in Tropaeolum XET6.3 (cyan; coordinates from [66]) is indicated in dashed lines at separations between 2.3 Å and 3.0 Å. Right panel: Details of XXXG binding in Tropaeolum XET6.3; interacting residues are marked in cyan cpk sticks in dots. Bottom panel: Some of the residues of Tropaeolum XET6.3 binding XLLG (highlighted in cyan) are shown in the alignment by PROMALS3D [83] of Populus XET16A and Tropaeolum XET6.3 sequences (numbering includes signal peptides). Conservation of residues on the scale 9–6 is shown on the top of the alignment in brown. Images were generated in the PyMOL Molecular Graphics System v2.5.2 (Schrődinger LLC, Portland, OR, USA).
Structural differences, which determine whether the XET/XEH enzymes transglycosylate (PDB accessions 1UN1, 1UMZ) or act as the hydrolases (TmNXG1; PDB accession 2UWA) [71,80,84], were reported including the TmNXG1-DELTAYNIIG mutant (PDB accession 2VH9). Slight structural differences, which led to the stronger binding of a donor substrate combined with larger loop flexibility at the acceptor site and the flexibility of residues, underlined the hydrolytic preference [80,84].
As detailed above, structural differences modulate the binding of donor substrates combined with the loop and residues flexibilities and underline the transfer or hydrolytic preferences of XTH enzymes. This is supported by phylogenomic analyses, where XTHs segregate in the three groups, with XEHs belonging to XTH III clade, while XETs clustered within XTH I and XTH II clades [64,66,80,85]. Based on these analyses, it was concluded that GH16 hydrolases including XEHs have evolved from XETs [60,84,86].
As previously summarised [69], XET activity assays use unlabelled donor substrates and radio-chemically or fluorescently labelled acceptors, or radio-labelled donor and unlabelled acceptors. The reaction products in both cases encompass labelled saccharides, while unincorporated donors or acceptors are removed via chromatography [47,49,66], or through a gel [63,87], washed off from a filter paper [88] or ethanol precipitated [89]. When selecting an appropriate approach, it is critical to select the right type of fluorescent tag [90,91] and how to remove efficiently the surplus of substrates [48,69]. Labelled oligosaccharide probes could be also utilised in high-throughput polysaccharide microarrays [92] and through in vivo visualisation [93,94,95,96]. Supplementary XET activity assays include colorimetry [97] that relies on the formation of the blue-green-colored iodine–XG complex, and viscometry that records viscosity changes in substrates.
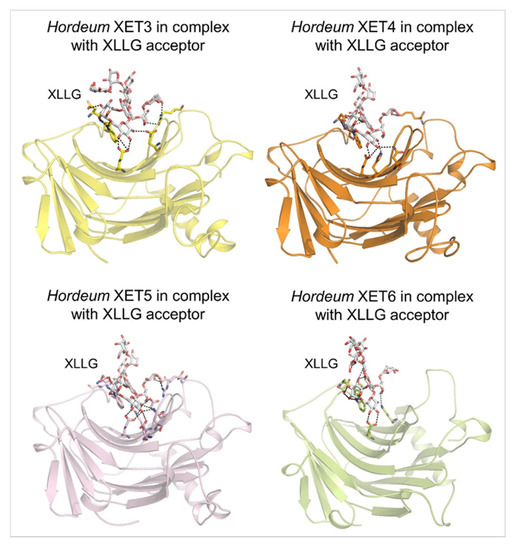
Figure 3.
Molecular models of the β-sandwich architecture with the β-jelly-roll topology of Hordeum XET3 (top left; yellow), Hordeum XET4 (top right; orange), Hordeum XET5 (bottom left; pink), and Hordeum XET6 (bottom right; lemon) in complex with the XLLG acceptors (cpk sticks). Coordinates of XET3, XET4 and XET6 with XLLG were taken from [81] and those of XET5 from [64]. XLLG was docked in XET5 by HDOCK that performs docking based on a hybrid algorithm of template-based modelling and ab initio free docking [98]; the top docking pose is shown for XET5 from 100 poses ranked by energy docking scores. Details of binding of XLLG by Hordeum XETs are shown in dashed lines. Separations are for XET3: 2.7 Å to 3.6 Å; XET4: 2.6 Å to 3.5 Å; XET5: 2.5 Å to 3.6 Å; XET6: 2.5 Å to 3.2 Å. Images were generated in PyMOL as referenced in Figure 2.
(d) Substrate specificity of transfer reactions with xyloglucan and other than xyloglucan-derived donors and acceptors—Substrate specificity of XETs has recently been evaluated in detail ([69]; cf. Table 1, vide infra); thus, here we only succinctly recapitulate key aspects of this topic.
The systematic description of xyloglucan:xyloglucosyl transferases (EC 2.4.1.207) has from the onset of their discoveries stated that these enzymes recognise XG donor (xyloglucan) and acceptor (xyloglucosyl) substrates—this catalytic activity represents (i) homo-transglycosylation reactions. Yet, since 2007 [47], the novel types of transfer reactions were described in barley XETs and later in other plant sources, termed (ii) hetero-transglycosylation reactions [63,66]. These reactions utilise the neutral donor and neutral or ionic acceptor substrates other than XG-derived [69].
(i) Homo-transglycosylation reactions with XG-derived donors and acceptors—The poplar PttXET16A [71,80] and Pinus radiata PrXTH1 [99] XETs, belonging to the I/II cluster of XTH gene products, were characterised as the homo-transglycosylation-type enzymes with a narrow substrate specificity that utilised XG-derived substrates. Hetero-transglycosylation activities in PttXET16A were undetected with cello-oligosaccharides [60], although the data with PrXTH1 suggested the weak binding of cellooctaose [99]. Phylogenomic analyses of the GH16 family, spanning monocots, eudicots and a basal Angiosperm [68] revealed that PttXET16A clustered with barley (Hordeum vulgare L.) XET5 that catalysed the reactions with XG or hydroxyethyl cellulose and XG- or cellulose-derived oligosaccharides [47].
(ii) Hetero-transglycosylation reactions with neutral donor and neutral or ionic acceptors,other than XG-derived—The key question to answer regarding hetero-transglycosylation reactions is, what is the limit for the activity ratio of the XG/XG-oligosaccharide substrate pair and chemically different substrates? Another key point is that the substrate specificity of only a few XETs in near-homogenous forms has been defined, and thus this property cannot be unequivocally assigned to many XETs.
Although the broad XET substrate specificity in Poaceae was predicted based on molecular modelling [100], the first experimental indication that XETs could catalyse hetero-transglycosylation reactions, i.e., to mediate transfers between saccharides other than XG-derived, was presented by Ait Mohand and Farkaš [63]. This study described glycosyl transfers from XG to cello- and laminari-oligosaccharides, and from carboxymethyl and hydroxyethyl cellulose derivatives to XG oligosaccharides, using the crude protein extracts prepared from germinating nasturtium seeds [63]. This study was followed with the barley XET5 isoform [47], the first XET in a near-homogenous form, with a defined primary structure, that catalysed hetero-transglycosylation reactions in vitro. Except for XGs, this enzyme linked covalently carboxymethyl and hydroxyethyl celluloses, and (1,3;1,4)-β-d-glucans as donors and XG- and cellulose-derived oligosaccharide acceptors. The 44% efficiency of barley XET5 with the hydroxyethyl cellulose donor and the XG-oligosaccharides was comparable to that of XG, but it was significantly lower with (1,3;1,4)-β-d-glucan. Subsequently, the hetero-transglycosylation reactions were defined for a near-homogenous barley XET6 isoform [64].
Other XET enzymes recognising cellulose as the donor substrate, were partially purified XET from parsley roots (Petroselinum crispum Mill. Fus) [89] and near-homogenous XTH3 from Arabidopsis thaliana L. Heynh, which catalysed the hetero-transglycosylation reactions between cellulose and cello-oligosaccharides or cellulose and XG-oligosaccharides [65]. Despite differences in the specificity of hetero-transglycosylation reactions, barley XET5 [47] and Arabidopsis XTH3 [65] clustered in the same phylogenetic group of XTH I, as presumably XG-specific PttXET16A and PrXTH1 [80,99], although they segregated to different sub-clades [69]. The next XETs with known primary structures and utilising besides XG, cellulose or (1,3;1,4)-β-d-glucans as donors, were three acidic EfXTH-A, EfXTH-H and EfXTH-I isoforms from Equisetum fluviatile L. [101], although the homogeneity of these enzymes was not shown. While EfXTH-A displayed the comparable transfer of (1,3;1,4)-β-d-glucan fragments to XG-oligosaccharides, as was the case of barley XET5 (0.2–0.3%) [47], the activity with cellulose was higher with barley XET5. EfXTH-H and EfXTH-I showed the equivalent hetero-transglycosylation activities with (1,3;1,4)-β-d-glucans and cellulose donors [101]. Conversely, HTG and MLG (1,3;1,4-β-d-glucan): xyloglucan endotransglycosylase enzymes from Equisetum preferred cellulose and (1,3;1,4)-β-d-glucans with XG-oligosaccharides as respective donors and acceptors [101]. Such enzymes were so far found only in Equisetum and the charophytic algae [74,102], where they are predicted to re-model hemicelluloses in horsetails shoots [78]. Molecular modelling of HTG suggested the amino acid residues responsible for the evolution of their substrate specificity [79].

Figure 4.
Details of the XXXG donor (cpk sticks) and the [α(1-4)GalAp]5 acceptor (magenta cpk sticks) binding substrates in Hordeum XET3 (A) and Hordeum XET4 (B). In both panels, blue and black dashed lines indicate residue separations between donors or acceptors that are 2.6 Å–3.6 Å (XET3) and 2.7 Å–3.4 Å (XET4). Interacting residues are shown in yellow (XET3) and orange (XET4) cpk sticks and emphasised in dots. Subsites at -4 to -1 for XXXG and +1 to +5 for [α(1-4)GalAp]5 are indicated. Images were generated in PyMOL as referenced in Figure 2. Bottom panel: Some of the residues of Hordeum XETs that bind [α(1-4)GalAp]5 (highlighted in yellow) are shown in the alignment by PROMALS3D [83] of the Hordeum XET sequences (numbering includes signal peptides). Conservation of residues on the scale 9–6 is shown on the top of the alignment in brown.
Recently, Tropaeolum majus L. XET6.3 classified in the XTH II clade [69] in its near-homogenous form displayed hetero-transglycosylation activity with the donor XG or hydroxyethyl cellulose and neutral oligosaccharide acceptors, such as those derived from cellulose, (1,3;1,4)-β-d-glucan, laminarin, pustulan, arabinoxylan, xylan, arabinan, arabinogalactan, mannan, glucomannan and galactomannan [66].
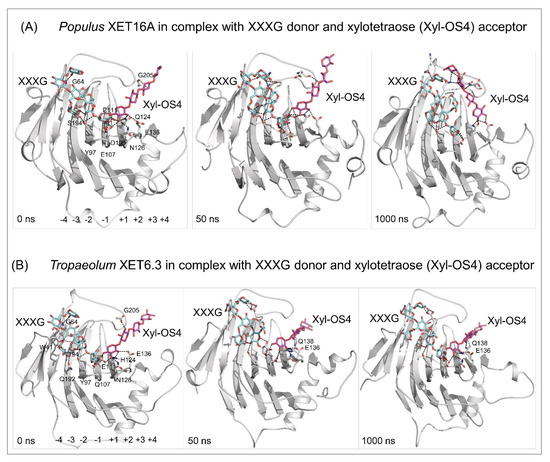
Figure 5.
Docking of the XXXG donor (cpk cyan sticks) and the xylotetraose (Xyl-OS4) acceptor (cpk magenta sticks) substrates in the active sites of Populus XET16A (PDB accession 1UN1) (A) and Tropaeolum XET6.3 [66] (B), and MD simulations of enzyme/substrate complexes after 0, 50, and 1000 ns (simulation times indicated in bottom-left corners). MD simulations were carried out similarly as described in [103]. Separations between the donors and acceptor substrates are between 2.6 Å and 3.6 Å for Populus XET16A and 2.7 Å–3.4 Å for Tropaeolum XET6.3. Residues (numbering includes signal peptides) mediating contacts with substrates are shown in cpk sticks. In Tropaeolum XET6.3, E136 and Q138 stabilise the binding of the Xyl-OS4 acceptor. For clarity, the residues and the subsite binding sites (−4 to −1 for XXXG and +1 to +4 for Xyl-OS) are shown in left panels only. Images were generated in PyMOL as referenced in Figure 2.
Finally, it is important to describe hetero-transglycosylation reactions that accept charged acceptors that were shown in several near-homogenous barley XETs [68] (Figure 4), where all tested neutral oligosaccharides served as the acceptors with barley XET3-XET6 isoforms, although with different efficiency. Barley XET3 and XET4 also transferred efficiently the fragments of XG or hydroxyethyl cellulose to the penta-galacturonic acid ([α(1-4)GalAp]5), a fragment of the linear part of pectin, [68]. These hetero-transglycosylation activities by barley XETs were demonstrated in vitro in CWs of barley roots using fluorescently labelled [α(1-4)GalAp]5) [68]. These findings were supported by the structural modelling of barley XET3 and XET4 with the docked XG heptasaccharide (XXXG) donor and the [α(1-4)GalAp]5 acceptor substrates in the −4 to +5 active site subsites (Figure 4), and suggested that the XET substrate poly-specificity resulted from protein sequences alterations that evoked structural re-arrangements [66,67,68,69,104].
In summary, only specific combinations of residues could underlie certain substrate specificity in XETs, which underpins the functionalisation of substrates other than XG-derived. Despite categorising these residues [66,68], the future understanding of the mechanisms of these reactions depends on the resolution of atomic structures of XETs. Future research will also be focused on in vitro and in silico studies to dissect the molecular mechanisms that drive the transglycosylation reactions catalysed by XETs (Figure 5).
4. Plant Cell Wall Modifications and Re-Modelling, and Methods of Investigation
(a) Plant cell wall function, dynamics and methods of investigation—Polysaccharide-rich nature is a key characteristic of plant CWs that through evolution reflected on a multitude of roles. These include mechanical support to counter osmotically driven turgor pressure or in the maintenance of cellular structural integrity by resisting internal hydrostatic pressures, regulation of plant growth, providing flexibility to support cell division and expansion during tissue differentiation, acting as an environmental barrier that protects cells through defence via metabolites to counteract biotic and abiotic (environmental) stresses, and for providing a milieu that through diffusion ensures cell-to-cell communication [1,2,3,4,69,105]. The evolution of changes in the plant CWs during these functions is characteristic for individual species and relies on the process of natural selection. It is exactly the pressure from surrounding environments, microorganisms, and other life forms that confront plants, which in return form CW structures that reflect and embody plant responses. This pressure, specifically from microorganisms to use plant CWs as an energy source to survive, simultaneously allows the evolution of novel biocatalysts cleaving most of the covalent linkages contained in CWs.
These multifunctional roles that underscored the evolution of plant CWs, require the formation of highly complex bio-composites, which underlines the economic feasibility of CW bioconversion to biofuels, biochemicals and other high-value products. The success of utilising the plant CWs, including the plant biomass, relies on the knowledge of the composition, structure, biosynthesis and biodegradation of CWs. Although this knowledge is not completely accessible, the structure–function relationships of CWs have become more tractable in recent years. Here, an astonishing increase in the accumulation of knowledge has occurred in the last twenty years to understand features that underlie the complexity and dynamics of plant CWs. This is fundamentally due to the progress in the physico-chemical and analytical methods, and the biological and biochemical technologies at multi-disciplinary levels, that allowed advancements to examine CWs.
These applications include the use of atomic force and scanning probe microscopy [106,107] to characterise the plant CW structure. This allows unique insights into the morphological features and molecular characteristics, such as heights, width, contour length, end-to-end, distance, and polydispersity of CW components. Other promising avenues include looking into the molecular motions and assembly formations and visualising the conformation behaviour under various conditions [107]. This technique was applied to image cellulose micro-fibrils up to a single molecular level under the near-native conditions, although other polysaccharides, namely, pectin, xanthan, carrageenan, curdlan, scleroglucan, XG, arabinoxylan and their detailed nanostructures were also characterised [107].
On the other hand, Raman spectroscopy and surface-enhanced Raman spectroscopic chemical imaging allowed investigating the mutual distribution and co-localisation of CW polysaccharides, such as pectin and XGs in, for example, onion cells [108]. Further, Felhofer and co-workers [109] described how CW thickening indentations were formed in primary CWs in algae at exactly defined areas, specifically where pectins and XGs co-localised, while in the secondary CWs, crystalline cellulose dominated in thickening areas. These observations allowed concluding that a denser network of cellulose micro-fibrils stiffened CWs at the indent and modified the CW extensibility.
Additional experimental approaches included applying one or two-dimensional Nuclear Magnetic Resonance spectroscopy to directly observe the random coil and flexible conformations of XGs in plant seeds [110], solid-state NMR to assess and quantify chemical effects (such as pH) on plant CWs [111], and confocal microscopy demonstrating that XG and cellulose formed the molecular cross-bridges connecting root border cells in pea [112]. The combination of these approaches and the equilibrium adsorptions of hemicelluloses offered new ideas, for example, that XGs could be involved in the lignification processes in primary wood CWs [113].
In conjunction with the experimental avenues, it is vital to mention theoretical MD calculations [114], based on the existing chemical and biological knowledge of plant CWs, where mutual interactions could be studied and, thus, provide the working hypotheses on how individual polysaccharides interact in CWs. For example, Zhang and co-workers [115] reported that in the water environments, van der Waals interactions prevail over electrostatic ones during the adsorption of XGs onto the Iβ 1–10 cellulose surface, and that variation in one side chain of XG had no influence on the interaction energy and the binding affinity between XGs and cellulose. Similar data provided the nano-scale views of polysaccharide interactions and showed that from five polymers (galacto-glucomannan, acetyl-galacto-glucomannan, fuco-galacto-XG, methylglucurono-xylan, methyl-glucurono-arabinoxylan), glucurono-arabinoxylan had the highest free energy of binding to cellulose nanocrystals, whereas XGs had the lowest energies amongst five models [116]. It was established that the lack of forming these interactions occurred at the inter-molecular level of hemicelluloses, rather than at the interface of cellulose, and that water molecules had a weakening effect on the hemicelluloses–cellulose interactions [115,116]—these findings might have implications for the roles of XETs in CW dynamics. Finally, a steered MD model using pull-out simulations (where force is applied to molecules along a chosen direction to understand interactions) provided the quantitative information on forces between cellulose micro-fibril surfaces [117]. These strategies are central to the deeper understanding of plant CWs, and collectively emphasise that with the mathematical modelling [118], it is vital at a microscale to tackle the challenges of understanding CW mechanics, plant tip growth, morphogenesis and stress feedback. Looking towards the future, one would expect burgeoning advances in this field because CWs are essential and indispensable to the life of plants.
(b) Roles of xyloglucan:xyloglucosyl transferases in cell wall re-modelling—The fundamental roles of XET enzymes in plants are in the assembly of the CW structures and in modifications during plant growth and development, in adaptations to living environments, and during biotic and abiotic stresses. This also underpins the major role of XET enzymes in the CW assembly and thickening, and in the regulation of its mechanical properties, through affecting the load-bearing framework of the XG-cellulose composites. This function of XET (together with XEH) enzymes is believed to regulate the CW plasticity along with non-catalytic expansins and other matrix polysaccharide-modifying enzymes. However, the precise molecular mechanisms of the XET activity in CWs are far from being clear [33,69]. To this end, the functional role of the specific Arabidopsis thaliana XET19 isoform (with undetectable hydrolase activity) was shown to increase the freezing tolerance after a low-temperature acclimation, providing a hypothesis that these enzymes modify (re-model) CWs and change their physico-chemical properties under the stress conditions [119]. Although XET enzymes have appeared concurrently with XGs in CWs of green algae (but not red or brown algae) [4,19], it remains to find out if other enzymes recognising cellulose as a donor substrate, had occurred in red or brown algae before the emergence of XET enzymes, as it is the case of HTGs [78].
The expression of XTH genes is regulated by hormones in response to a variety of environmental fluctuations. For this reason, XETs have evolved in multiple isoforms, which fulfil the specific roles during various stages of plant development and in response to biotic and abiotic stresses, such as drought, heat and metal toxicity [55,120,121,122,123,124,125,126,127,128]. XTH genes also play a unique role in the saccharide metabolism and energy storage that are connected to plant growth, as their elevated expression during elongation of cells directly affects the CW characteristics that underlie the growth and function of plant tissues through structuralisation and re-modelling [129,130].
It has now been accepted that XETs control polymerisation of plant structural polysaccharides such as XGs, and recognise cellulose [47,48,65,67,71,128,131,132,133]. XET enzymes primarily work on the XG molecules associated with cellulose micro-fibrils such that this process does not influence the integrity of CWs [1,47]. The discovery of XET isoforms that link not only XG molecules but also those of cellulose and celluose-derived oligosaccharides by barley XET5 [47] and Arabidopsis XTH3 [65], or XG, (1,3;1,4)-β-d-glucans and cellulose catalysed by barley XET5 [47] and Equisetum HTGs [67,77,78,133], or XG and a wide spectrum of structurally diverse neutral saccharides, catalysed by Tropaeolum XET6.3 [66], and XG and pectin fragments by barley XET isoforms [64], suggested that plants established the sophisticated and variable mechanisms to influence firmness, porosity, and flexibility (stiffness and extensibility) of CWs [47]. This allows plants to continuously modify the properties of CWs that otherwise would lack this adaptability when needed. Although it is not always known how precisely are these specific or broad specific XET isoforms expressed in plant tissues during a defined plant developmental stage, this information could be obtained from the localisation studies and gene expression analyses [66,68,77,100,134,135,136]. The significance of the transglycosylation reactions catalysed by broad specific XETs may be related to the structural roles of polysaccharides in plant CWs. These specific expression patterns thus provide a piece of evidence that it is the flexibility and properties of CWs that are required to fulfill their dynamic roles during embryogenesis, vegetative and reproductive growth, and environmental responses of plants, as these CW polysaccharides are synthesised and interact [44,45].
In conclusion, the fundamental complexities of structural polysaccharides in plant CWs have been defined [1,34,137,138,139,140], although, these complexities are not completely transparent, including the precise significance of homo- and hetero-transglycosylation reactions catalysed by XETs. Although the definition of the catalytic function of xyloglucan:xyloglucosyl transferases and more broadly of the XTH family have significantly progressed during the past fifteen years, there are limitations (and still a lack of information) as to how this knowledge can be organically implemented in the function of plant CWs. One example of how these complexities could be better tackled includes the generation of XET/XEH mutant knockout plants (in explicit plant tissues of different taxa) that would be deficient in certain XET/XEH isoforms, to understand the fine structure of plant CWs. This approach could also answer the prodigious question of the potential interchangeability in hemicellulose polymers [8], and if additional mutations to certain hemicellulose biosynthetic enzymes would activate compensatory mechanisms, while other mutations would activate feedback processes between biosynthetic pathways and endomembrane compartments [141]. The latter notions combined with the definitions of roles of broad specific XETs that generate novel hetero-polymeric networks could modify our views on plant CW architectures. Examining these novel relationships will spur a better understanding of the roles of XTH enzymes in plant CWs from macro to atomic levels. Thus, it is imperative to use the knowledge of XTHs in a CW context, amongst other applications, for the rational design of economically important crops using genetic engineering and more broadly bio-technology to increase stress tolerance, tackle drought and other abiotic stresses [105], and produce modified (thermo-responsive) XG hydrogels for bio-medicine [142] and wound healing [143].
Author Contributions
Conceptualisation and original draft preparation—M.H.; generation of figures—M.H. and B.S.; editing and review—B.S., E.S. and M.H. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by DP120100900 from the Australian Research Council (Australia) to M.H., and VEGA 2/0137/20 (Slovakia).
Acknowledgments
The authors thank Stanislav Kozmon (Institute of Chemistry, Centre for Glycomics, Slovak Academy of Sciences, Slovakia) for advice in molecular docking. M.H. acknowledges the Huaiyin Normal University (China) and the University of Adelaide (Australia) for additional funding.
Conflicts of Interest
The authors declare no conflict of interest.
List of Abbreviations in Alphabetical Order
| [α(1-4)GalAp]5 | penta-galacturonic acid |
| CW(s) | cell wall(s) |
| CAZy | Carbohydrate-Active enZYmes |
| GH(s) | glycoside hydrolase(s) |
| GT(s) | glycosyl transferase(s) |
| HTG(s) | hetero-trans-β-glucanase(s) |
| MD | molecular dynamics |
| MLG(s) | (1,3;1,4)-β-d-glucan(s) or mix-linkage glucan(s) |
| PDB | Protein Data Bank |
| XEH(s) | xyloglucan endohydrolase(s) |
| XET(s) | xyloglucan endo-transglycosylase(s) |
| XG(s) | xyloglucan(s) |
| XTH(s) | xyloglucan transglycosylase/hydrolase(s) |
References
- Cosgrove, D. Growth of the plant cell wall. Nat. Rev. Mol. Cell. Biol. 2005, 6, 850–861. [Google Scholar] [CrossRef]
- Farrokhi, N.; Burton, R.A.; Brownfield, L.; Hrmova, M.; Wilson, S.M.; Bacic, A.; Fincher, G.B. Plant cell wall biosynthesis: Genetic, biochemical and functional genomics approaches to the identification of key genes. Plant Biotech. J. 2009, 4, 145–167. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, D.J. Plant cell wall extensibility: Connecting plant cell growth with cell wall structure, mechanics, and the action of wall modifying enzymes. J. Exp. Bot. 2015, 67, 463–476. [Google Scholar] [CrossRef]
- Popper, Z.A.; Michel, G.; Hervé, C.; Domozych, D.S.; Willats, W.G.; Tuohy, M.G.; Kloareg, B.; Stengel, D.B. Evolution and diversity of plant cell walls: From algae to flowering plants. Annu. Rev. Plant Biol. 2011, 62, 567–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef] [PubMed]
- Popper, Z.A.; Fry, S.C. Widespread occurrence of a covalent linkage between xyloglucan and acidic polysaccharides in suspension-cultured angiosperm cells. Ann. Bot. 2005, 96, 91–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcus, S.E.; Verhertbruggen, Y.; Hervé, C.; Ordaz-Ortiz, J.J.; Farkaš, V.; Pedersen, H.L.; Willats, W.G.T.; Knox, J.P. Pectic homogalacturonan masks abundant sets of xyloglucan epitopes in plant cell walls. BMC Plant Biol. 2008, 8, 60. [Google Scholar] [CrossRef] [Green Version]
- McCann, M.C.; Knox, J.P. Plant Cell Wall Biology: Polysaccharides in architectural and developmental contexts. Ann. Plant Rev. 2018, 343–366. [Google Scholar] [CrossRef]
- Harholt, J.; Suttangkakul, A.; Scheller, V.H. Biosynthesis of pectin. Plant Physiol. 2010, 153, 384–395. [Google Scholar] [CrossRef] [Green Version]
- Mohnen, D. Pectin structure and biosynthesis. Curr. Opin. Plant Biol. 2008, 11, 266–277. [Google Scholar] [CrossRef]
- Caffall, K.H.; Mohnen, D. The structure, function, and biosynthesis of plant cell wall pectic polysaccharides. Carbohydr. Res. 2009, 344, 1879–1900. [Google Scholar] [CrossRef] [PubMed]
- Boerjan, W.; Ralph, J.; Baucher, M. Lignin biosynthesis. Annu. Rev. Plant Biol. 2003, 54, 519–546. [Google Scholar] [CrossRef]
- Weng, J.K.; Chapple, C. The origin and evolution of lignin biosynthesis. New Phytol. 2010, 187, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Mnich, E.; Bjarnholt, N.; Eudes, A.; Harholt, J.; Holland, C.; Jørgensen, B.; Larsen, F.H.; Liu, M.; Manat, R.; Meyer, A.S.; et al. Phenolic cross-links: Building and de-constructing the plant cell wall. Nat. Prod. Rep. 2020, 37, 919–961. [Google Scholar] [CrossRef]
- Déjardin, A.; Laurans, F.; Arnaud, D.; Breton, C.; Pilate, G.; Leplé, J.-C. Wood formation in angiosperms. Comptes Rendus Biol. 2010, 333, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Kushwah, S.; Banasiak, A.; Nishikubo, N.; Derba-Maceluch, M.; Majda, M.; Endo, S.; Kumar, V.; Gomez, L.; Gorzsas, A.; McQueen-Mason, S. Arabidopsis XTH4 and XTH9 contribute to wood cell expansion and secondary wall formation. Plant Physiol. 2020, 182, 1946–1965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.J.; Chandrasekar, B.; Rea, A.C.; Danhof, L.; Zemelis-Durfee, S.; Thrower, N.; Shepard, Z.S.; Pauly, M.; Brandizzi, F.; Keegstra, K. The synthesis of xyloglucan, an abundant plant cell wall polysaccharide, requires CSLC function. Proc. Natl. Acad. Sci. USA 2020, 117, 20316–20324. [Google Scholar] [CrossRef] [PubMed]
- Ikegaya, H.; Hayashi, T.; Kaku, T.; Iwata, K.; Sonobe, S.; Shimmen, T. Presence of xyloglucan-like polysaccharide in Spirogyra and possible involvement in cell–cell attachment. Phycol. Res. 2008, 56, 216–222. [Google Scholar] [CrossRef]
- Van Sandt, V.S.; Stieperaere, H.; Guisez, Y.; Verbelen, J.P.; Vissenberg, K. XET activity is found near sites of growth and cell elongation in bryophytes and some green algae: New insights into the evolution of primary cell wall elongation. Ann. Bot. 2007, 99, 39–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- York, W.S.; Van Halbeek, H.; Darvill, A.G.; Albersheim, P. Structural analysis of xyloglucan oligosaccharides by 1H-n.m.r. spectroscopy and fast-atom-bombardment mass spectrometry. Carbohydr. Res. 1990, 200, 9–31. [Google Scholar] [CrossRef]
- Peña, M.J.; Darvill, A.G.; Eberhard, S.; York, W.S.; O’Neill, M.A. Moss and liverwort xyloglucans contain galacturonic acid and are structurally distinct from the xyloglucans synthesized by hornworts and vascular plants. Glycobiology 2008, 18, 891–904. [Google Scholar] [CrossRef] [PubMed]
- Peña, M.J.; Kong, Y.; York, W.S.; O’neill, M.A. A galacturonic acid-containing xyloglucan is involved in Arabidopsis root hair tip growth. Plant Cell 2012, 24, 4511–4524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, Y.S.Y.; Harris, P.J. Xyloglucans of monocotyledons have diverse structures. Mol. Plant 2009, 2, 943–965. [Google Scholar] [CrossRef]
- Cocuron, J.C.; Lerouxel, O.; Drakakaki, G.; Alonso, A.P.; Liepman, A.H.; Keegstra, K.; Raikhel, N.; Wilkerson, C.G. A gene from the cellulose synthase-like C family encodes a β-1,4 glucan synthase. Proc. Natl. Acad. Sci. USA 2007, 104, 8550–8555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauly, M.; Keegstra, K. Biosynthesis of the plant cell wall matrix polysaccharide xyloglucan. Annu. Rev. Plant Biol. 2016, 67, 235–259. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Phillips, D.R.; Ye, Z.-H. A single xyloglucan xylosyltransferase is sufficient for generation of the XXXG xylosylation pattern of xyloglucan. Plant Cell Physiol. 2021, 62, 1589–1602. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T. Xyloglucans in the primary cell wall. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 139–168. [Google Scholar] [CrossRef]
- Carpita, N.C.; Gibeaut, D.M. Structural models of primary cell walls in flowering plants: Consistency of molecular structure with the physical properties of the walls during growth. Plant J. 1993, 3, 1–30. [Google Scholar] [CrossRef]
- Pauly, M.; Albersheim, P.; Darvill, A.; York, W.S. Molecular domains of the cellulose/xyloglucan network in the cell walls of higher plants. Plant J. 1999, 20, 629–639. [Google Scholar] [CrossRef]
- Albersheim, P.; Darvill, A.; Roberts, K.; Sederoff, R.; Staehelin, A. Plant Cell Walls, from Chemistry to Biology; Garland Science: New York, NY, USA, 2011; pp. 227–272. ISBN 978-0815319962. [Google Scholar]
- Cosgrove, D.J. Re-constructing our models of cellulose and primary cell wall assembly. Curr. Opin. Plant Biol. 2014, 22, 122–131. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.B.; Cosgrove, D.J. A revised architecture of primary cell walls based on biomechanical changes induced by substrate-specific endoglucanases. Plant Physiol. 2012, 158, 1933–1943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, Y.B.; Cosgrove, D.J. Xyloglucan and its interactions with other components of the growing cell wall. Plant Cell Physiol. 2015, 56, 180–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosgrove, D.J. Diffuse growth of plant cell walls. Plant Physiol. 2018, 176, 16–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuki, H.; Yokoyama, R.; Kuroha, T.; Nishitani, K. Xyloglucan is not essential for the formation and integrity of the cellulose network in the primary cell wall regenerated from Arabidopsis protoplasts. Plants 2020, 9, 629. [Google Scholar] [CrossRef] [PubMed]
- Scheible, W.R.; Pauly, M. Glycosyltransferases and cell wall biosynthesis: Novel players and insights. Curr. Opin. Plant Biol. 2004, 7, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Burton, R.A.; Wilson, S.M.; Hrmova, M.; Harvey, A.J.; Shirley, N.J.; Medhurst, A.; Stone, B.A.; Newbigin, E.J.; Bacic, A.; Fincher, G.B. Cellulose synthase- like CslF genes mediate the synthesis of cell wall (1,3;1,4)-β-d-glucans. Science 2006, 311, 1940–1942. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.-M.; Hörnblad, E.; Voxeur, A.; Gerber, L.; Rihouey, C.; Lerouge, P.; Marchant, A. Analysis of the Arabidopsis IRX9/IRX9-L and IRX14/IRX14-L pairs of glycosyltransferase genes reveals critical contributions to biosynthesis of the hemicellulose glucuronoxylan. Plant Physiol. 2010, 153, 542–554. [Google Scholar] [CrossRef] [Green Version]
- Dhugga, K.S. Biosynthesis of non-cellulosic polysaccharides of plant cell walls. Phytochemistry 2012, 74, 8–19. [Google Scholar] [CrossRef]
- Roberts, A.W.; Roberts, E.M.; Delmer, D.P. Cellulose synthase (CesA) genes in the green alga Mesotaenium caldariorum. Eukaryot. Cell 2002, 1, 847–855. [Google Scholar] [CrossRef] [Green Version]
- Roberts, A.W.; Roberts, E. Evolution of the cellulose synthase (CesA) gene family: Insights from green algae and seedless plants. In Cellulose: Molecular and structural Biology; Brown, R.M., Saxena, I.M., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 17–34. [Google Scholar] [CrossRef]
- Abercrombie, J.M.; O’Meara, B.C.; Moffatt, A.R.; Williams, J.H. Developmental evolution of flowering plant pollen tube cell walls: Callose synthase (CalS) gene expression patterns. EvoDevo 2011, 2, 14. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Szumlanski, A.L.; Gu, F.; Guo, F.; Nielsen, E. A role for CSLD3 during cell-wall synthesis in a pical plasma membranes of tip-growing root-hair cells. Nat. Cell Biol. 2011, 13, 973–980. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Loosening of plant cell walls by expansins. Nature 2000, 407, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.M.; Zuo, D.Y.; Wang, X.F.; Cheng, H.L.; Zhang, Y.P.; Wang, Q.L.; Song, G.L.; Ma, Z.Y. Genome-wide identification of the expansin gene family reveals that expansin genes are involved in fibre cell growth in cotton. BMC Plant Biol. 2020, 20, 223. [Google Scholar] [CrossRef] [PubMed]
- Willats, W.G.T.; Orfila, C.; Limberg, G.; Buchholt, H.C.; van Alebeek, G.-J.W.M.; Voragen, A.G.J.; Marcus, S.E.; Christensen, T.M.I.E.; Mikkelsen, J.D.; Murray, B.S.; et al. Modulation of the degree and pattern of methyl-esterification of pectic homogalacturonan in plant cell walls: Implications for pectin methylesterase action, matrix properties and cell adhesion. J. Biol. Chem. 2001, 276, 19404–19413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hrmova, M.; Farkaš, V.; Lahnstein, J.; Fincher, G.B. A barley xyloglucan xyloglucosyl transferase covalently links xyloglucan, cellulosic substrates, and (1,3;1,4)-β-d-glucans. J. Biol. Chem. 2007, 283, 27344. [Google Scholar] [CrossRef]
- Franková, L.; Fry, S.C. Biochemistry and physiological roles of enzymes that ‘cut and paste’ plant cell-wall polysaccharides. J. Exp. Bot. 2013, 64, 3519–3550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishitani, K.; Tominaga, R. Endo-xyloglucan transferase, a novel class of glycosyltransferase that catalyzes transfer of a segment of xyloglucan molecule to another xyloglucan molecule. J. Biol. Chem. 1992, 267, 21058–21064. [Google Scholar] [CrossRef]
- Farkaš, V.; Sulová, Z.; Stratilová, E.; Hanna, R.; Maclachlan, G. Cleavage of xyloglucan by nasturtium seed xyloglucanase and transglycosylation to xyloglucan subunit oligosaccharides. Arch. Biochem. Biophys. 1992, 298, 365–370. [Google Scholar] [CrossRef]
- Fry, S.; Smith, R.; Renwick, K.; Martin, D.; Hodge, S.; Matthews, K. Xyloglucan endotransglycosylase, a new wall-loosening enzyme activity from plants. Biochem. J. 1992, 282, 821–828. [Google Scholar] [CrossRef] [Green Version]
- Webb, E.C. Enzyme Nomenclature 1992: Recommendations of the Nomenclature Committee of the International Union of Biochemistry and Molecular Biology on the Nomenclature and Classification of Enzymes; Academic Press Inc.: Cambridge, MA, USA; Harcourt Brace Jovanovich Publishers: San Diego, CA, USA, 1992; ISBN 0-12-227164-5. [Google Scholar]
- Placzek, S.; Schomburg, I.; Chang, A.; Jeske, L.; Ulbrich, M.; Tillack, J.; Schomburg, D. BRENDA in 2017: New perspectives and new tools in BRENDA. Nucleic Acids Res. 2017, 45, D380–D388. [Google Scholar] [CrossRef]
- Rose, J.K.C.; Braam, J.; Fry, S.C.; Nishitani, K. The XTH family of enzymes involved in xyloglucan endotransglucosylation and endohydrolysis: Current perspectives and a new unifying nomenclature. Plant Cell Physiol. 2002, 43, 1421–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atkinson, R.G.; Johnston, S.L.; Yauk, Y.K.; Sharma, N.N.; Schröder, R. Analysis of xyloglucan endotransglucosylase/hydrolase (XTH) gene families in kiwifruit and apple. Postharvest Biol. Technol. 2009, 51, 149–157. [Google Scholar] [CrossRef]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CAZypedia Consortium. Ten years of CAZypedia: A living encyclopedia of carbohydrate-active enzymes. Glycobiology 2018, 28, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Viborg, A.H.; Terrapon, N.; Lombard, V.; Michel, G.; Gurvan, M.; Czjzek, M.; Henrissat, B.; Brumer, H. A subfamily roadmap for functional glycogenomics of the evolutionarily diverse Glycoside Hydrolase Family 16 (GH16). J. Biol. Chem. 2019, 294, 15973–15986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eklöf, J.M.; Brumer, H. The XTH gene family: An update on enzyme structure, function, and phylogeny in xyloglucan remodeling. Plant Physiol. 2010, 153, 456–466. [Google Scholar] [CrossRef] [Green Version]
- Mark, P.; Baumann, M.J.; Eklöf, J.M.; Gullfot, F.; Michel, G.; Kallas, A.M.; Teeri, T.T.; Brumer, H.; Czjek, M. Analysis of nasturtium TmNXG1 complexes by crystallography and molecular dynamics provides detailed insight into substrate recognition by family GH16 xyloglucan endo-transglycosylases and endo-hydrolases. Proteins 2009, 75, 820–836. [Google Scholar] [CrossRef]
- Baran, R.; Sulová, Z.; Stratilová, E.; Farkaš, V. Ping-Pong character of nasturtium-seed xyloglucan endotransglycosylase (XET) Reaction. Gen. Physiol. Biophys. 2000, 19, 427–440. [Google Scholar]
- Saura-Valls, M.; Faure, R.; Ragas, S.; Piens, K.; Brumer, H.; Teeri, T.T.; Cottaz, S.; Driguez, H.; Planas, A. Kinetic analysis using low-molecular mass xyloglucan oligosaccharides defines the catalytic mechanism of a Populus xyloglucan endotransglycosylase. Biochem. J. 2006, 395, 99–106. [Google Scholar] [CrossRef]
- Ait Mohand, F.; Farkaš, V. Screening for hetero-transglycosylating activities in extracts from nasturtium (Tropaeolum majus). Carbohydr. Res. 2006, 34, 577–581. [Google Scholar] [CrossRef]
- Hrmova, M.; Farkaš, V.; Harvey, A.J.; Lahnstein, J.; Wischmann, B.; Kaewthai, N.; Ezcurra, I.; Teeri, T.T.; Fincher, G.B. Substrate specificity and catalytic mechanism of a xyloglucan xyloglucosyl transferase HvXET6 from barley (Hordeum vulgare L.). FEBS J. 2009, 276, 437–456. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, N.; Sunagawa, N.; Tamura, S.; Yokoyama, R.; Ueda, M.; Igarashi, K.; Nishitani, K. The plant cell-wall enzyme AtXTH3 catalyses covalent cross-linking between cellulose and cello-oligosaccharide. Sci. Rep. 2017, 7, 46099. [Google Scholar] [CrossRef] [PubMed]
- Stratilová, B.; Firáková, Z.; Klaudiny, J.; Šesták, S.; Kozmon, S.; Strouhalová, D.; Garajová, S.; Ait-Mohand, F.; Horváthová, Á.; Farkaš, V.; et al. Engineering the acceptor substrate specificity in the xyloglucan endotransglycosylase TmXET6.3 from nasturtium seeds (Tropaeolum majus L.). Plant Mol. Biol. 2019, 100, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Herburger, K.; Franková, L.; Sanhueza, D.; Roig-Sanchez, S.; Meulewaeter, F.; Hudson, A.; Thomson, A.; Laromaine, A.; Budtova, T.; Fry, S.C. Enzymically attaching oligosaccharide-linked ‘cargoes’ to cellulose and other commercial polysaccharides via stable covalent bonds. Int. J. Biol. Macromol. 2020, 164, 4359–4369. [Google Scholar] [CrossRef]
- Stratilová, B.; Šesták, S.; Mravec, J.; Garajová, S.; Pakanová, Z.; Vadinová, K.; Kučerová, D.; Kozmon, S.; Schwerdt, J.G.; Shirley, N.; et al. Another building block in the plant cell wall: Barley xyloglucan xyloglucosyl transferases link covalently xyloglucan and anionic oligosaccharides derived from pectin. Plant J. 2020, 104, 752–767. [Google Scholar] [CrossRef]
- Stratilová, B.; Kozmon, S.; Stratilová, E.; Hrmova, M. Xyloglucan xyloglucosyl transferases and the cell wall structure: Subtle but significant. Molecules 2020, 25, 5619. [Google Scholar] [CrossRef]
- Hrmova, M.; MacGregor, E.A.; Biely, P.; Stewart, R.J.; Fincher, G.B. Substrate binding and catalytic mechanism of a barley β-d-glucosidase/(1,4)-β-d-glucan exohydrolase. J. Biol. Chem. 1998, 273, 11134–11143. [Google Scholar] [CrossRef] [Green Version]
- Johansson, P.; Brumer, H.; Baumann, M.J.; Kallas, A.M.; Henriksson, H.; Denman, S.E.; Teeri, T.T.T.; Jones, T.A. Crystal structures of a poplar xyloglucan endotransglycosylase reveal details of transglycosylation acceptor binding. Plant Cell 2004, 16, 874–886. [Google Scholar] [CrossRef] [Green Version]
- Hrmova, M.; Fincher, G.B. Barley β-d-glucan exohydrolases. Substrate specificity and kinetic properties. Carbohydr. Res. 1998, 305, 209–221. [Google Scholar] [CrossRef]
- Schröder, R.; Atkinson, R.G.; Redgwell, R.J. Re-interpreting the role of endo-β-mannanases as mannan endotransglycosylase/hydrolases in the plant cell wall. Ann. Bot. 2009, 104, 197–204. [Google Scholar] [CrossRef] [Green Version]
- Franková, L.; Fry, S.C. Phylogenetic variation in glycosidases and glycanases acting on plant cell wall polysaccharides, and the detection of transglycosidase and trans-β-xylanase activities. Plant J. 2011, 67, 662–681. [Google Scholar] [CrossRef] [PubMed]
- Johnston, S.; Prakash, R.; Chen, N.J.; Kumagai, M.H.; Turano, H.M.; Cooney, J.M.; Atkinson, R.G.; Paull, R.E.; Cheetamun, R.; Bacic, A.; et al. An enzyme activity capable of endotransglycosylation of heteroxylan polysaccharides is present in plant primary cell walls. Planta 2013, 237, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Derba-Maceluch, M.; Awano, T.; Takahashi, J.; Lucenius, J.; Ratke, C.; Kontro, I.; Busse-Wicher, M.; Kosik, O.; Tanaka, R.; Winzéll, A.; et al. Suppression of xylan endotransglycosylase PtxtXyn10A affects cellulose microfibril angle in secondary wall in aspen wood. New Phytol. 2015, 205, 666–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fry, S.C.; Mohler, K.E.; Nesselrode, B.H.W.A.; Franková, L. Mixed-linkage β-glucan:xyloglucan endotransglucosylase, a novel wall-remodelling enzyme from Equisetum (horsetails) and charophytic algae. Plant J. 2008, 55, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Mohler, K.E.; Simmons, T.J.; Fry, S.C. Mixed-linkage glucan:xyloglucan endotransglucosylase (MXE) re-models hemicelluloses in Equisetum shoots but not in barley shoots or Equisetum callus. New Phytol. 2013, 197, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Simmons, T.J.; Mohler, K.E.; Holland, C.; Goubet, F.; Franková, L.; Houston, D.R.; Hudson, A.D.; Meulewaeter, F.; Fry, S.C. Hetero-trans-β-glucanase, an enzyme unique to Equisetum plants, functionalizes cellulose. Plant J. 2015, 83, 753–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumann, M.J.; Eklöf, J.M.; Michel, G.; Kallas, A.M.; Teeri, T.T.; Czjzek, M.; Brumer, H. Structural evidence for the evolution of xyloglucanase activity from xyloglucan endo-transglycosylases: Biological implications for cell wall metabolism. Plant Cell 2007, 19, 1947–1963. [Google Scholar] [CrossRef] [Green Version]
- Vaaje-Kolstad, G.; Farkaš, V.; Fincher, G.B.; Hrmova, M. Barley xyloglucan xyloglucosyl transferases bind xyloglucan-derived oligosaccharides in their acceptor-binding regions in multiple conformational states. Arch. Biochem. Biophys. 2010, 496, 61–68. [Google Scholar] [CrossRef]
- Eramian, D.; Eswar, N.; Shen, M.; Sali, A. How well can the accuracy of comparative protein structure models be predicted? Prot. Sci. 2008, 17, 1881–1893. [Google Scholar] [CrossRef] [Green Version]
- Pei, J.; Kim, B.-H.; Grishin, N.V. PROMALS3D: A tool for multiple protein sequence and structure alignments. Nucleic Acids Res. 2008, 36, 2295–2300. [Google Scholar] [CrossRef]
- McGregor, N.; Yin, V.; Tung, C.C.; Van Petegem, F.; Brumer, H. Crystallographic insight into the evolutionary origins of xyloglucan endotransglycosylases and endohydrolases. Plant J. 2017, 89, 651–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaewthai, N.; Harvey, A.J.; Hrmova, M.; Brumer, H.; Ezcurra, I.; Teeri, T.T.; Fincher, G.B. Heterologous expression of diverse barley XTH genes in the yeast Pichia pastoris. Plant Biotechnol. 2010, 27, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Behar, H.; Graham, S.W.; Brumer, H. Comprehensive cross-genome survey and phylogeny of glycoside hydrolase family 16 members reveals the evolutionary origin of EG 16 and XTH proteins in plant lineages. Plant J. 2018, 95, 1114–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farkaš, V.; Ait-Mohand, F.; Stratilová, E. Sensitive detection of transglycosylating activity of xyloglucan endotransglycosylase/hydrolase (XTH) after isoelectric focusing in polyacrylamide gels. Plant Physiol. Biochem. 2005, 43, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Fry, S.C. Novel ‘dot-blot’ assays for glycosyltransferases and glycosylhydrolases: Optimization for xyloglucan endotransglycosylase (XET) activity. Plant J. 1997, 11, 1141–1150. [Google Scholar] [CrossRef]
- Garajová, S.; Flodrová, D.; Ait-Mohand, F.; Farkaš, V.; Stratilová, E. Characterization of two partially purified xyloglucan endotransglycosylases from parsley (Petroselinum crispum) roots. Biologia 2008, 63, 313–319. [Google Scholar] [CrossRef]
- Vaaje-Kolstad, G.; Farkaš, V.; Hrmova, M.; Fincher, G.B. Xyloglucan xyloglucosyl transferases from barley (Hordeum vulgare L.) bind oligomeric and polymeric xyloglucan molecules in their acceptor binding sites. Biochim. Biophys. Acta 2010, 1800, 674–684. [Google Scholar] [CrossRef]
- Kosík, O.; Garajová, S.; Matulová, M.; Řehulka, P.; Stratilová, E.; Farkaš, V. Effect of the label of oligosaccharide acceptors on the kinetic parameters of nasturtium seed xyloglucan endotransglycosylase (XET). Carbohydr. Res. 2011, 346, 357–361. [Google Scholar] [CrossRef]
- Kosík, O.; Auburn, R.P.; Russell, S.; Stratilová, E.; Garajová, S.; Hrmova, M.; Farkaš, V. Polysaccharide microarrays for high-throughput screening of transglycosylase activities in plant extracts. Glycoconj. J. 2010, 27, 79–87. [Google Scholar] [CrossRef]
- Vissenberg, K.; Martinez-Vilchez, I.M.; Verbelen, J.P.; Miller, J.G.; Fry, S.C. In vivo colocalization of xyloglucan endotransglycosylase activity and its donor substrate in the elongation zone of Arabidopsis roots. Plant Cell 2000, 12, 1229–1237. [Google Scholar] [CrossRef] [Green Version]
- Nishikubo, N.; Awano, T.; Banasiak, A.; Bourquin, V.; Ibatullin, F.; Funada, R.; Brumer, H.; Teeri, T.T.; Hayashi, T.; Sundberg, B.; et al. Xyloglucan endo-transglycosylase (XET) functions in gelatinous layers of tension wood fibers in poplar—a glimpse into the mechanism of the balancing act of trees. Plant Cell Phys. 2007, 48, 843–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibatullin, F.M.; Banasiak, A.; Baumann, M.J.; Greffe, L.; Takahashi, J.; Mellerowicz, E.J.; Brumer, H.A. Real-time fluorogenic assay for the visualization of glycoside hydrolase activity in planta. Plant Phys. 2009, 151, 1741–1750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mravec, J.; Kračun, S.K.; Rydahl, M.G.; Westereng, B.; Miart, F.; Clausen, M.H.; Fangel, J.U.; Daugaard, M.; Van Cutsem, P.; De Fine Licht, H.H.; et al. Tracking developmentally regulated post-synthetic processing of homogalacturonan and chitin using reciprocal oligosaccharide probes. Development 2014, 141, 4841–4850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulová, Z.; Lednická, M.; Farkaš, V. A colorimetric assay for xyloglucan endotransglycosylase from germinating seeds. Anal. Biochem. 1995, 229, 80–85. [Google Scholar] [CrossRef]
- Yan, Y.; Zhang, D.; Zhou, P.; Li, B.; Huang, S.-Y. HDOCK: A web server for protein-protein and protein-DNA/RNA docking based on a hybrid strategy. Nucleic Acids Res. 2017, 45, W365–W373. [Google Scholar] [CrossRef]
- Morales-Quintana, L.; Carrasco-Orellana, C.; Beltrán, D.; Moya-León, M.A.; Herrera, R. Molecular insights of a xyloglucan endo-transglycosylase/hydrolase of radiata pine (PrXTH1) expressed in response to inclination: Kinetics and computational study. Plant Physiol. Biochem. 2019, 136, 155–161. [Google Scholar] [CrossRef]
- Strohmeier, M.; Hrmova, M.; Fischer, M.; Harvey, A.J.; Fincher, J.B.; Pleiss, J. Molecular modeling of family GH16 glycoside hydrolases: Potential roles for xyloglucan transglucosylases/hydrolases in cell wall modification in the Poaceae. Prot. Sci. 2004, 13, 3200–3213. [Google Scholar] [CrossRef] [Green Version]
- Holland, C.; Simmons, T.J.; Meulewaeter, F.; Hudson, A.; Fry, S.C. Three highly acidic Equisetum XTHs differ from hetero-trans-β-glucanase in donor substrate specificity and are predominantly xyloglucan homo-transglucosylases. J. Plant Physiol. 2020, 251, 153210. [Google Scholar] [CrossRef]
- Herburger, K.; Ryan, L.M.; Popper, Z.A.; Holzinger, A. Localisation and substrate specificities of transglycanases in charophyte algae relate to development and morphology. J. Cell Sci. 2018, 131, jcs203208. [Google Scholar] [CrossRef] [Green Version]
- Stratilová, B.; Řehulka, P.; Garajová, S.; Řehulková, H.; Stratilová, E.; Hrmova, M.; Kozmon, S. Structural characterization of the Pet c 1.0201 PR-10 protein isolated from roots of Petroselinum crispum (Mill.) Fuss. Phytochemistry 2020, 175, 112368. [Google Scholar] [CrossRef]
- Seven, M.; Derman, Ü.C.; Harvey, A.J. Enzymatic characterization of ancestral/group-IV clade xyloglucan endotransglycosylase/hydrolase enzymes reveals broad substrate specificities. Plant J. 2021, 106, 1060–1073. [Google Scholar] [CrossRef] [PubMed]
- Ezquer, I.; Salameh, I.; Colombo, L.; Kalaitzis, P. Plant cell walls tackling climate change: Insights into plant cell wall remodeling, its regulation, and biotechnological strategies to improve crop adaptations and photosynthesis in response to global warming. Plants 2020, 9, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yarbrough, J.M.; Himmel, M.E.; Ding, S.Y. Plant cell wall characterization using scanning probe microscopy techniques. Biotechnol. Biofuels 2009, 2, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Nie, S. Application of atomic force microscopy in microscopic analysis of polysaccharide. Trends Food Sci. Technol. 2019, 87, 35–46. [Google Scholar] [CrossRef]
- He, Q.; Yang, J.; Zabotina, O.A.; Yu, C. Surface-enhanced Raman spectroscopic chemical imaging reveals distribution of pectin and its co-localization with xyloglucan inside onion epidermal cell wall. PLoS ONE 2021, 16, e0250650. [Google Scholar] [CrossRef] [PubMed]
- Felhofer, M.; Mayr, K.; Lütz-Meindl, U.; Gierlinger, N. Raman imaging of Micrasterias: New insights into shape formation. Protoplasma 2021, 258, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Lucyszyn, N.; Lubambo, A.; Ono, L.; Tatiane, A.; Jó, C.F.; Souza, M. Sierakowski. Chemical, physico-chemical and cytotoxicity characterisation of xyloglucan from Guibourtia hymenifolia (Moric.) J. Leonard seeds. Food Hydrocoll. 2011, 25, 1242–1250. [Google Scholar] [CrossRef] [Green Version]
- Phyo, P.; Gu, Y.; Hong, M. Impact of acidic pH on plant cell wall polysaccharide structure and dynamics: Insights into the mechanism of acid growth in plants from solid-state NMR. Cellulose 2019, 26, 291–304. [Google Scholar] [CrossRef]
- Ropitaux, M.; Bernard, S.; Follet-Gueye, M.L.; Vicré, M.; Boulogne, I.; Driouich, A. Xyloglucan and cellulose form molecular cross-bridges connecting root border cells in pea (Pisum sativum). Plant Physiol. Biochem. 2019, 139, 191–196. [Google Scholar] [CrossRef]
- Lyu, Y.; Matsumoto, T.; Taira, S.; Ijiri, K.; Yoshinaga, A.; Shigetomi, K.; Uraki, Y. Influences of polysaccharides in wood cell walls on lignification in vitro. Cellulose 2021, 28, 9907–9917. [Google Scholar] [CrossRef]
- Kishani, S.; Benselfelt, T.; Wågberg, L.; Wohlert, J. Entropy drives the adsorption of xyloglucan to cellulose surfaces—A molecular dynamics study. J. Colloid Interface Sci. 2021, 588, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Brumer, H.; Ågren, H.; Tu, Y. The adsorption of xyloglucan on cellulose: Effects of explicit water and side chain variation. Carbohydr. Res. 2011, 346, 2595–2602. [Google Scholar] [CrossRef] [PubMed]
- Khodayari, A.; Thielemans, W.; Hirn, U.; Van Vuure, A.W.; Seveno, D. Cellulose-hemicellulose interactions—A nanoscale view. Carbohydr. Polym. 2021, 270, 118364. [Google Scholar] [CrossRef] [PubMed]
- Ezquerro, C.S.; Minana, C.C.; Izquierdo, S.; Laspalas, M. A molecular dynamics model to measure forces between cellulose fibril surfaces: On the effect of noncovalent polyelectrolyte adsorption. Cellulose 2019, 26, 1449–1466. [Google Scholar] [CrossRef] [Green Version]
- Smithers, E.T.; Luo, J.; Dyson, R.J. Mathematical principles and models of plant growth mechanics: From cell wall dynamics to tissue morphogenesis. J. Exp. Bot. 2019, 70, 3587–3600. [Google Scholar] [CrossRef]
- Takahashi, D.; Johnson, K.L.; Hao, P.; Tuong, T.; Erban, A.; Sampathkumar, A.; Bacic, A.; Livingston, D.P., 3rd; Kopka, J.; Kuroha, T.; et al. Cell wall modification by the xyloglucan endotransglucosylase/hydrolase XTH19 influences freezing tolerance after cold and sub-zero acclimation. Plant Cell Environ. 2021, 44, 915–930. [Google Scholar] [CrossRef]
- De Caroli, M.; Manno, E.; Piro, G.; Lenucci, M.S. Ride to cell wall: Arabidopsis XTH11, XTH29 and XTH33 exhibit different secretion pathways and responses to heat and drought stress. Plant J. 2021, 107, 448–466. [Google Scholar] [CrossRef]
- Muñoz-Bertomeu, J.; Miedes, E.; Lorences, E.P. Expression of xyloglucan endotransglucosylase/hydrolase (XTH) genes and XET activity in ethylene treated apple and tomato fruits. J. Plant Physiol. 2013, 170, 1194–1201. [Google Scholar] [CrossRef] [Green Version]
- Zemková, Z.; Garajová, S.; Flodrová, D.; Řehulka, P.; Zelko, I.; Vadkertiová, R.; Farkaš, V.; Stratilová, E. Incorporation of β-(1,6)-linked glucooligosaccharides (pustulooligosaccharides) into plant cell wall structures. Chem. Pap. 2012, 66, 814–820. [Google Scholar] [CrossRef]
- Catala, C.; Rose, J.K.C.; York, W.S.; Albersheim, P.; Darvill, A.G.; Bennett, A.B. Characterization of a tomato xyloglucan endotransglycosylase gene that is down-regulated by auxin in etiolated hypocotyls. Plant Physiol. 2001, 127, 1180–1192. [Google Scholar] [CrossRef]
- Matsui, A.; Yokoyama, R.; Seki, M.; Ito, T.; Shinozaki, K.; Takahashi, T.; Komeda, Y.; Nishitani, K. AtXTH27 plays an essential role in cell wall modification during the development of tracheary elements. Plant J. 2005, 42, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Nardi, C.F.; Villarreal, N.M.; Opazo, M.C.; Martinez, G.A.; Moya León, M.A.; Civello, P.M. Expression of FaXTH1 and FaXTH2 genes in strawberry fruit. Cloning of promoter regions and effect of plant growth regulators. Sci. Hortic. 2014, 165, 111–122. [Google Scholar] [CrossRef]
- Han, Y.; Zhu, Q.; Zhang, Z.; Meng, K.; Hou, Y.; Ban, Q.; Suo, J.; Rao, J. Analysis of xyloglucan endotransglycosylase/hydrolase (XTH) genes and diverse roles of isoenzymes during persimmon fruit development and postharvest softening. PLoS ONE 2015, 10, e0123668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Ban, Q.; Hou, Y.; Meng, K.; Suo, J.; Rao, J. Isolation and characterization of two persimmon xyloglucan endotransglycosylase/hydrolase (XTH) genes that have divergent functions in cell wall modification and fruit postharvest softening. Front. Plant Sci. 2016, 7, 624. [Google Scholar] [CrossRef] [Green Version]
- Campbell, P.; Braam, J. In vitro activities of four xyloglucan endotransglycosylases from Arabidopsis. Plant J. 1999, 18, 371–382. [Google Scholar] [CrossRef]
- Bourquin, V.; Nishikubo, N.; Abe, H.; Brumer, H.; Denman, S.; Eklund, M.; Christiernin, M.; Teeri, T.T.; Sundberg, B.; Mellerowicz, E.J. Xyloglucan endotransglycosylases have a function during the formation of secondary cell walls of vascular tissues. Plant Cell 2002, 14, 3073–3088. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.E.; Smith, R.C.; Fry, S.C. Xyloglucan undergoes interpolymeric transglycosylation during binding to the plant cell wall in vivo: Evidence from 13C/3H dual labelling and isopycnic centrifugation in caesium trifluoroacetate. Biochem. J. 1997, 327, 699–708. [Google Scholar] [CrossRef] [Green Version]
- Oh, M.H.; Romanow, W.G.; Smith, R.C.; Zamski, E.; Sasse, J.; Clouse, S.D. Soybean BRU1 encodes a functional xyloglucan endotransglycosylase that is highly expressed in inner epicotyl tissues during brassinosteroid-promoted elongation. Plant Cell Physiol. 1998, 39, 124–130. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.E.; Fry, S.C. Restructuring of wall-bound xyloglucan by transglycosylation in living plant cells. Plant J. 2001, 26, 23–34. [Google Scholar] [CrossRef]
- Simmons, T.J.; Fry, S.C. Bonds broken and formed during the mixed-linkage glucan:xyloglucan endotransglucosylase reaction catalysed by Equisetum hetero-trans-β-glucanase. Biochem. J. 2017, 474, 1055–1070. [Google Scholar] [CrossRef] [Green Version]
- Tucker, M.R.; Lou, H.; Aubert, M.K.; Wilkinson, L.G.; Little, A.; Houston, K.; Pinto, S.C.; Shirley, N.J. Exploring the role of cell wall-related genes and polysaccharides during plant development. Plants 2018, 7, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Li, H.; Yin, C.; Wang, X.; Jiang, Q.; Zhang, R.; Ge, F.; Chen, Y.; Yang, L. Genome-wide identification and characterization of xyloglucan endotransglycosylase/ hydrolase in Ananas comosus during Development. Genes 2019, 10, 537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Xie, F.; He, Q.; Li, J.; Liu, J.; Sun, B.; Luo, Y.; Zhang, Y.; Chen, Q.; Zhang, F.; et al. Expression Analysis of XTH in stem swelling of stem mustard and selection of reference genes. Genes 2020, 11, 113. [Google Scholar] [CrossRef] [Green Version]
- Voiniciuc, C.; Engle, K.A.; Günl, M.; Dieluweit, S.; Schmidt, M.H.; Yang, J.Y.; Moremen, K.W.; Mohnen, D.; Usadel, B. Identification of key enzymes for pectin synthesis in seed mucilage. Plant Physiol. 2018, 178, 1045–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpita, N.; McCann, M. The cell wall. In Biochemistry and Molecular Biology of Plants; Buchanan, B.B., Wilhelm, G., Jones, R.L., Eds.; American Society of Plant Physiologists: Rockville, IL, USA, 2000; pp. 52–108. ISBN 978-0-470-71421-8. [Google Scholar]
- Thompson, D.S. How do cell walls regulate plant growth? J. Exp. Bot. 2005, 56, 2275–2285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Xiao, G.; Wang, X.; Zhang, X.; Friml, J. Evolution of fast root gravitropism in seed plants. Nat. Commun. 2019, 10, 3480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, N.; King, S.; Samuels, A.L.; McFarlane, H.E. Subcellular coordination of plant cell wall synthesis. Dev. Cell 2021, 56, 933–948. [Google Scholar] [CrossRef] [PubMed]
- Talantikite, M.; Stimpson, T.C.; Gourlay, A.; Le-Gall, S.; Moreau, C.; Cranston, E.D.; Moran-Mirabal, J.M.; Cathala, B. Bioinspired thermoresponsive xyloglucan-cellulose nanocrystal hydrogels. Biomacromolecules 2021, 22, 743–753. [Google Scholar] [CrossRef]
- Ajovalasit, A.; Redondo-Gómez, C.; Sabatino, M.A.; Okesola, B.O.; Braun, K.; Mata, A.; Dispenza, C. Carboxylated-xyloglucan and peptide amphiphile co-assembly in wound healing. Regen. Biomater. 2021, 8, rbab040. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).