
The Role of Exosomal Non-Coding RNAs in Colorectal Cancer Drug Resistance

 ,
,  , and
, and 
Abstract
:1. Introduction
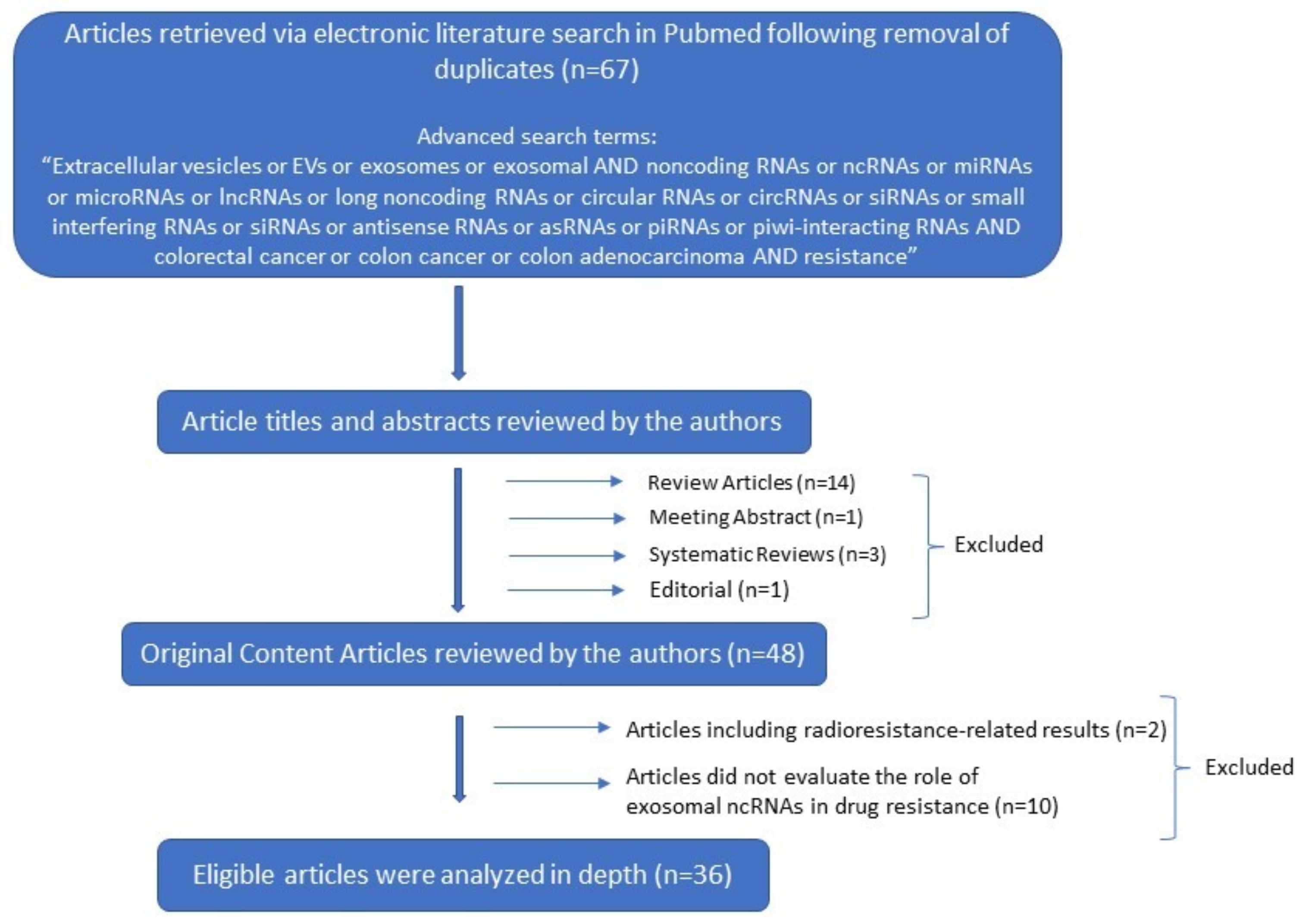
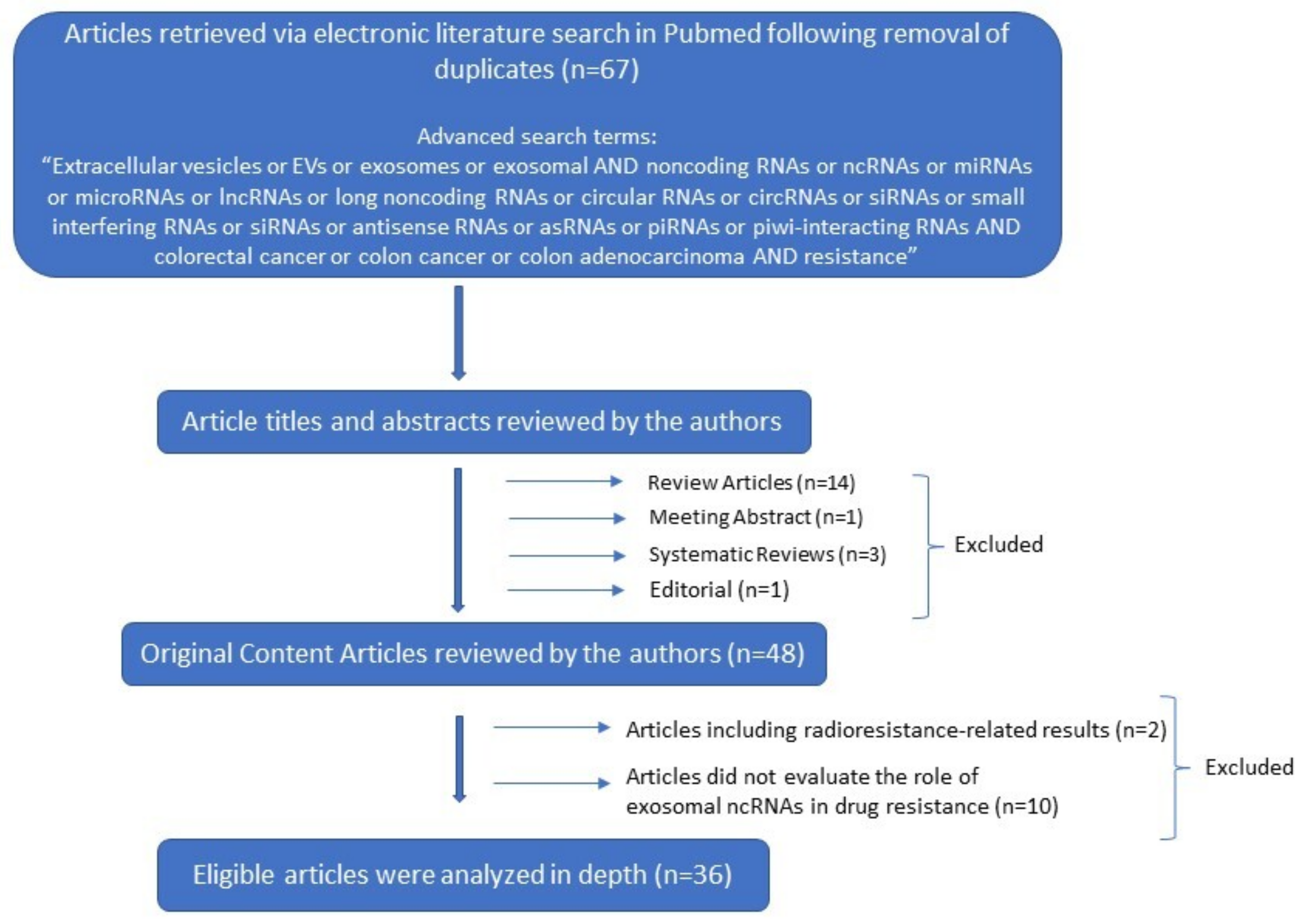
2. Methods and Results
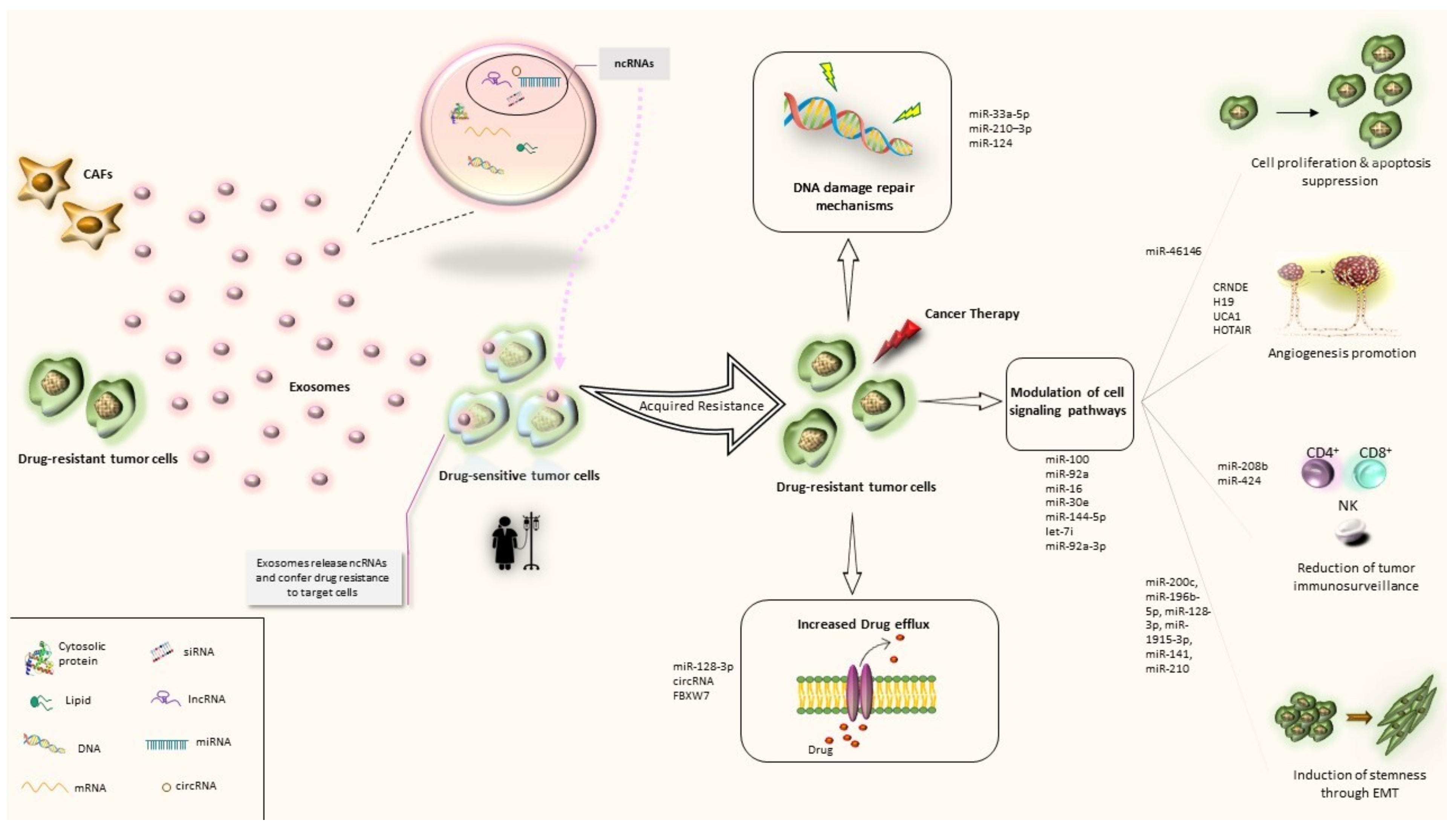
3. Biological Mechanisms of Exosomal ncRNAs in CRC Drug Resistance
4. Exosomal ncRNAs in CRC Drug Resistance: Current Evidence
4.1. Exosomal miRNAs and CRC Drug Resistance
4.2. Exosomal Long ncRNAs and CRC Drug Resistance
4.3. Exosomal circRNAs and CRC Drug Resistance
4.4. Exosomal siRNAs and CRC Drug Resistance
5. Final Remarks and Future Perspectives
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Filip, S.; Vymetalkova, V.; Petera, J.; Vodickova, L.; Kubecek, O.; John, S.; Cecka, F.; Krupova, M.; Manethova, M.; Cervena, K.; et al. Distant Metastasis in Colorectal Cancer Patients-Do We Have New Predicting Clinicopathological and Molecular Biomarkers? A Comprehensive Review. Int. J. Mol. Sci. 2020, 21, 5255. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.L.; Wang, W.; Lan, X.L.; Zeng, Z.C.; Liang, Y.S.; Yan, Y.R.; Song, F.Y.; Wang, F.F.; Zhu, X.H.; Liao, W.J.; et al. CAFs secreted exosomes promote metastasis and chemotherapy resistance by enhancing cell stemness and epithelial-mesenchymal transition in colorectal cancer. Mol. Cancer 2019, 18, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhang, H.; Chen, X. Drug resistance and combating drug resistance in cancer. Cancer Drug Resist 2019, 2, 141–160. [Google Scholar] [CrossRef] [Green Version]
- Bhome, R.; Del Vecchio, F.; Lee, G.H.; Bullock, M.D.; Primrose, J.N.; Sayan, A.E.; Mirnezami, A.H. Exosomal microRNAs (exomiRs): Small molecules with a big role in cancer. Cancer Lett. 2018, 420, 228–235. [Google Scholar] [CrossRef]
- Laschos, K.; Lampropoulou, D.I.; Aravantinos, G.; Piperis, M.; Filippou, D.; Theodoropoulos, G.; Gazouli, M. Exosomal noncoding RNAs in cholangiocarcinoma: Laboratory noise or hope? World J. Gastrointest Surg. 2020, 12, 407–424. [Google Scholar] [CrossRef]
- Weng, W.; Liu, N.; Toiyama, Y.; Kusunoki, M.; Nagasaka, T.; Fujiwara, T.; Wei, Q.; Qin, H.; Lin, H.; Ma, Y.; et al. Novel evidence for a PIWI-interacting RNA (piRNA) as an oncogenic mediator of disease progression, and a potential prognostic biomarker in colorectal cancer. Mol. Cancer 2018, 17, 16. [Google Scholar] [CrossRef]
- Guo, Q.R.; Wang, H.; Yan, Y.D.; Liu, Y.; Su, C.Y.; Chen, H.B.; Yan, Y.Y.; Adhikari, R.; Wu, Q.; Zhang, J.Y. The Role of Exosomal microRNA in Cancer Drug Resistance. Front. Oncol. 2020, 10, 472. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, S. Roles of circular RNAs in colorectal cancer. Oncol Lett. 2021, 22, 602. [Google Scholar] [CrossRef]
- Dashti, F.; Mirazimi, S.M.A.; Rabiei, N.; Fathazam, R.; Rabiei, N.; Piroozmand, H.; Vosough, M.; Rahimian, N.; Hamblin, M.R.; Mirzaei, H. The role of non-coding RNAs in chemotherapy for gastrointestinal cancers. Mol. Ther. Nucleic Acids 2021, 26, 892–926. [Google Scholar] [CrossRef]
- Maleki, M.; Golchin, A.; Javadi, S.; Khelghati, N.; Morovat, P.; Asemi, Z.; Alemi, F.; Vaghari-Tabari, M.; Yousefi, B.; Majidinia, M. Role of exosomal miRNA in chemotherapy resistance of Colorectal cancer: A systematic review. Chem. Biol. Drug Des. 2021. Online ahead of print. [Google Scholar] [CrossRef]
- Xitong, D.; Xiaorong, Z. Targeted therapeutic delivery using engineered exosomes and its applications in cardiovascular diseases. Gene 2016, 575, 377–384. [Google Scholar] [CrossRef]
- Srivastava, A.; Amreddy, N.; Babu, A.; Panneerselvam, J.; Mehta, M.; Muralidharan, R.; Chen, A.; Zhao, Y.D.; Rzaq, M.; Riedinger, N.; et al. Nanosomes carrying doxorubicin exhibit potent anticancer activity against human lung cancer cells. Sci. Rep. 2016, 6, 38541. [Google Scholar] [CrossRef]
- Ota, Y.; Takahashi, K.; Otake, S.; Tamaki, Y.; Okada, M.; Aso, K.; Makino, Y.; Fujii, S.; Ota, T.; Haneda, M. Extracellular vesicle-encapsulated miR-30e suppresses cholangiocarcinoma cell invasion and migration via inhibiting epithelial-mesenchymal transition. Oncotarget 2018, 9, 16400–16417. [Google Scholar] [CrossRef] [Green Version]
- Ha, D.; Yang, N.; Nadithe, V. Exosomes as therapeutic drug carriers and delivery vehicles across biological membranes: Current perspectives and future challenges. Acta Pharm. Sin. B 2016, 6, 287–296. [Google Scholar] [CrossRef] [Green Version]
- Mansoori, B.; Mohammadi, A.; Davudian, S.; Shirjang, S.; Baradaran, B. The Different Mechanisms of Cancer Drug Resistance: A Brief Review. Adv. Pharm. Bull. 2017, 7, 339–348. [Google Scholar] [CrossRef]
- Shedden, K.; Xie, X.T.; Chandaroy, P.; Chang, Y.T.; Rosania, G.R. Expulsion of small molecules in vesicles shed by cancer cells: Association with gene expression and chemosensitivity profiles. Cancer Res. 2003, 63, 4331–4337. [Google Scholar]
- Lv, M.M.; Zhu, X.Y.; Chen, W.X.; Zhong, S.L.; Hu, Q.; Ma, T.F.; Zhang, J.; Chen, L.; Tang, J.H.; Zhao, J.H. Exosomes mediate drug resistance transfer in MCF-7 breast cancer cells and a probable mechanism is delivery of P-glycoprotein. Tumour Biol. 2014, 35, 10773–10779. [Google Scholar] [CrossRef]
- Bebawy, M.; Combes, V.; Lee, E.; Jaiswal, R.; Gong, J.; Bonhoure, A.; Grau, G.E. Membrane microparticles mediate transfer of P-glycoprotein to drug sensitive cancer cells. Leukemia 2009, 23, 1643–1649. [Google Scholar] [CrossRef] [Green Version]
- Ambudkar, S.V.; Sauna, Z.E.; Gottesman, M.M.; Szakacs, G. A novel way to spread drug resistance in tumor cells: Functional intercellular transfer of P-glycoprotein (ABCB1). Trends Pharmacol. Sci. 2005, 26, 385–387. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.F.; Luk, F.; Gong, J.; Jaiswal, R.; Grau, G.E.; Bebawy, M. Microparticles mediate MRP1 intercellular transfer and the re-templating of intrinsic resistance pathways. Pharmacol. Res. 2013, 76, 77–83. [Google Scholar] [CrossRef]
- Goler-Baron, V.; Assaraf, Y.G. Structure and function of ABCG2-rich extracellular vesicles mediating multidrug resistance. PLoS ONE 2011, 6, e16007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapuy, B.; Koch, R.; Radunski, U.; Corsham, S.; Cheong, N.; Inagaki, N.; Ban, N.; Wenzel, D.; Reinhardt, D.; Zapf, A.; et al. Intracellular ABC transporter A3 confers multidrug resistance in leukemia cells by lysosomal drug sequestration. Leukemia 2008, 22, 1576–1586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.B.; Yan, C.; Mu, L.; Mi, Y.L.; Zhao, H.; Hu, H.; Li, X.L.; Tao, D.D.; Wu, Y.Q.; Gong, J.P.; et al. Exosomal Wnt-induced dedifferentiation of colorectal cancer cells contributes to chemotherapy resistance. Oncogene 2019, 38, 1951–1965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Yan, C.; Mu, L.; Huang, K.; Li, X.; Tao, D.; Wu, Y.; Qin, J. Fibroblast-derived exosomes contribute to chemoresistance through priming cancer stem cells in colorectal cancer. PloS ONE 2015, 10, e0125625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahrami, A.; Moradi Binabaj, M.; Ferns, G.A. Exosomes: Emerging modulators of signal transduction in colorectal cancer from molecular understanding to clinical application. Biomed. Pharmacother. 2021, 141, 111882. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Yuan, T.; Tschannen, M.; Sun, Z.; Jacob, H.; Du, M.; Liang, M.; Dittmar, R.L.; Liu, Y.; Liang, M.; et al. Characterization of human plasma-derived exosomal RNAs by deep sequencing. BMC Genom. 2013, 14, 319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.X.; Cai, Y.Q.; Lv, M.M.; Chen, L.; Zhong, S.L.; Ma, T.F.; Zhao, J.H.; Tang, J.H. Exosomes from docetaxel-resistant breast cancer cells alter chemosensitivity by delivering microRNAs. Tumour Biol. 2014, 35, 9649–9659. [Google Scholar] [CrossRef]
- Chen, W.X.; Liu, X.M.; Lv, M.M.; Chen, L.; Zhao, J.H.; Zhong, S.L.; Ji, M.H.; Hu, Q.; Luo, Z.; Wu, J.Z.; et al. Exosomes from drug-resistant breast cancer cells transmit chemoresistance by a horizontal transfer of microRNAs. PLoS ONE 2014, 16, e95240. [Google Scholar] [CrossRef]
- Wei, Y.; Lai, X.; Yu, S.; Chen, S.; Ma, Y.; Zhang, Y.; Li, H.; Zhu, X.; Yao, L.; Zhang, J. Exosomal miR-221/222 enhances tamoxifen resistance in recipient ER-positive breast cancer cells. Breast Cancer Res. Treat. 2014, 147, 423–431. [Google Scholar] [CrossRef]
- Salvatore, L.; Calegari, M.A.; Loupakis, F.; Fassan, M.; Di Stefano, B.; Bensi, M.; Bria, E.; Tortora, G. PTEN in Colorectal Cancer: Shedding Light on Its Role as Predictor and Target. Cancers 2019, 11, 1765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Shen, H.; Yin, X.; Yang, M.; Wei, H.; Chen, Q.; Feng, F.; Liu, Y.; Xu, W.; Li, Y. Macrophages derived exosomes deliver miR-223 to epithelial ovarian cancer cells to elicit a chemoresistant phenotype. J. Exp. Clin. Cancer Res. 2019, 38, 81. [Google Scholar] [CrossRef]
- Ragusa, M.; Statello, L.; Maugeri, M.; Barbagallo, C.; Passanisi, R.; Alhamdani, M.S.; Li Destri, G.; Cappellani, A.; Barbagallo, D.; Scalia, M.; et al. Highly skewed distribution of miRNAs and proteins between colorectal cancer cells and their exosomes following Cetuximab treatment: Biomolecular, genetic and translational implications. Oncoscience 2014, 1, 132–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, W.L.; Lan, H.Y.; Cheng, W.C.; Huang, S.C.; Yang, M.H. Tumor stem-like cell-derived exosomal RNAs prime neutrophils for facilitating tumorigenesis of colon cancer. J. Hematol Oncol. 2019, 12, 10. [Google Scholar] [CrossRef] [Green Version]
- Eguchi, T.; Taha, E.A.; Calderwood, S.K.; Ono, K. A Novel Model of Cancer Drug Resistance: Oncosomal Release of Cytotoxic and Antibody-Based Drugs. Biology 2020, 9, 47. [Google Scholar] [CrossRef] [Green Version]
- Paulus, P.; Stanley, E.R.; Schäfer, R.; Abraham, D.; Aharinejad, S. Colony-stimulating factor-1 antibody reverses chemoresistance in human MCF-7 breast cancer xenografts. Cancer Res. 2006, 66, 4349–4356. [Google Scholar] [CrossRef] [Green Version]
- Escamilla, J.; Schokrpur, S.; Liu, C.; Priceman, S.J.; Moughon, D.; Jiang, Z.; Pouliot, F.; Magyar, C.; Sung, J.L.; Xu, J.; et al. CSF1 receptor targeting in prostate cancer reverses macrophage-mediated resistance to androgen blockade therapy. Cancer Res. 2015, 75, 950–962. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Chen, Z.; Hua, D.; He, D.; Wang, L.; Zhang, P.; Wang, J.; Cai, Y.; Gao, C.; Zhang, X.; et al. Essential role for TrpC5-containing extracellular vesicles in breast cancer with chemotherapeutic resistance. Proc. Natl. Acad. Sci. USA 2014, 111, 6389–6394. [Google Scholar] [CrossRef] [Green Version]
- Binenbaum, Y.; Fridman, E.; Yaari, Z.; Milman, N.; Schroeder, A.; Ben David, G.; Shlomi, T.; Gil, Z. Transfer of miRNA in Macrophage-Derived Exosomes Induces Drug Resistance in Pancreatic Adenocarcinoma. Cancer Res. 2018, 78, 5287–5299. [Google Scholar] [CrossRef] [Green Version]
- Han, M.; Hu, J.; Lu, P.; Cao, H.; Yu, C.; Li, X.; Qian, X.; Yang, X.; Yang, Y.; Han, N.; et al. Exosome-Transmitted miR-567 reverses trastuzumab resistance by inhibiting ATG5 in breast cancer. Cell Death Dis. 2020, 11, 43. [Google Scholar] [CrossRef]
- Ren, J.; Ding, L.; Zhang, D.; Shi, G.; Xu, Q.; Shen, S.; Wang, Y.; Wang, T.; Hou, Y. Carcinoma-associated fibroblasts promote the stemness and chemoresistance of colorectal cancer by transferring exosomal lncRNA H19. Theranostics 2018, 8, 3932–3948. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Ruan, H.; Zhang, X.; Xu, X.; Zhu, Y.; Peng, H.; Zhang, X.; Kong, F.; Guan, M. Long noncoding RNA CCAL transferred from fibroblasts by exosomes promotes chemoresistance of colorectal cancer cells. Int. J. Cancer 2020, 146, 1700–1716. [Google Scholar] [CrossRef]
- Xu, J.; Xiao, Y.; Liu, B.; Pan, S.; Liu, Q.; Shan, Y.; Li, S.; Qi, Y.; Huang, Y.; Jia, L. Exosomal MALAT1 sponges miR-26a/26b to promote the invasion and metastasis of colorectal cancer via FUT4 enhanced fucosylation and PI3K/Akt pathway. J. Exp. Clin. Cancer Res. 2020, 39, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, C.; Yi, X.; Wu, X.; Bu, X.; Wang, D.; Wu, Z.; Zhang, G.; Gu, J.; Kang, D. Exosome-mediated transfer of circRNA CircNFIX enhances temozolomide resistance in glioma. Cancer Lett. 2020, 479, 1–12. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, H.; Yang, H.; Bai, M.; Ning, T.; Deng, T.; Liu, R.; Fan, Q.; Zhu, K.; Li, J.; et al. Exosome-Delivered circRNA promotes glycolysis to induce chemoresistance through the miR-122-PKM2 axis in colorectal cancer. Mol. Oncol. 2020, 14, 539–555. [Google Scholar] [CrossRef]
- Choi, D.-S.; Park, J.O.; Jang, S.C.; Yoon, Y.J.; Jung, J.W.; Choi, D.Y.; Kim, J.W.; Kang, J.S.; Park, J.; Hwang, D.; et al. Proteomic analysis of microvesicles derived from human colorectal cancer ascites. Proteomics 2011, 11, 2745–2751. [Google Scholar] [CrossRef]
- Xie, Y.; Dang, W.; Zhang, S.; Yue, W.; Yang, L.; Zhai, X.; Yan, Q.; Lu, J. The role of exosomal noncoding RNAs in cancer. Mol. Cancer. 2019, 18, 37. [Google Scholar] [CrossRef] [Green Version]
- He, Q.; Ye, A.; Ye, W.; Liao, X.; Qin, G.; Xu, Y.; Yin, Y.; Luo, H.; Yi, M.; Xian, L.; et al. Cancer-Secreted exosomal miR-21-5p induces angiogenesis and vascular permeability by targeting KRIT1. Cell Death Dis. 2021, 12, 576. [Google Scholar] [CrossRef]
- Simon, T.; Pinioti, S.; Schellenberger, P.; Rajeeve, V.; Wendler, F.; Cutillas, P.R.; King, A.; Stebbing, J.; Giamas, G. Shedding of bevacizumab in tumour cells-derived extracellular vesicles as a new therapeutic escape mechanism in glioblastoma. Mol. Cancer. 2018, 17, 132. [Google Scholar] [CrossRef]
- Huber, V.; Fais, S.; Iero, M.; Lugini, L.; Canese, P.; Squarcina, P.; Zaccheddu, A.; Colone, M.; Arancia, G.; Gentile, M.; et al. Human Colorectal Cancer Cells Induce T-Cell Death Through Release of Proapoptotic Microvesicles: Role in Immune Escape. Gastroenterology 2005, 128, 1796–1804. [Google Scholar] [CrossRef]
- Taylor, D.; Gerçel-Taylor, C.; Lyons, K.; Stanson, J.; Whiteside, T. T-Cell Apoptosis and Suppression of T-Cell Receptor/Cd3-Zeta by Fas Ligand-Containing Membrane Vesicles Shed From Ovarian Tumors. Clin. Cancer Res. 2003, 9, 5113–5119. [Google Scholar] [PubMed]
- Clayton, A.; Mitchell, J.; Court, J.; Mason, M.; Tabi, Z. Human Tumor-Derived Exosomes Selectively Impair Lymphocyte Responses to Interleukin-2. Cancer Res. 2007, 67, 7458–7466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Huang, A.C.; Zhang, W.; Zhang, G.; Wu, M.; Xu, W.; Yu, Z.; Yang, J.; Wang, B.; Sun, H.; et al. Exosomal PD-L1 contributes to immunosuppression and is associated with anti-PD-1 response. Nature 2018, 560, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Byrne, A.; Savas, P.; Sant, S.; Li, R.; Virassamy, B.; Luen, S.J.; Beavis, P.A.; Mackay, L.K.; Neeson, P.J.; Loi, S. Tissue-Resident memory T cells in breast cancer control and immunotherapy responses. Nat. Rev. Clin. Oncol. 2020, 17, 341–348. [Google Scholar] [CrossRef]
- Chen, S.W.; Zhu, S.Q.; Pei, X.; Qiu, B.Q.; Xiong, D.; Long, X.; Lin, K.; Lu, F.; Xu, J.J.; Wu, Y.B. Cancer cell-derived exosomal circUSP7 induces CD8+ T cell dysfunction and anti-PD1 resistance by regulating the miR-934/SHP2 axis in NSCLC. Mol. Cancer 2021, 20, 144. [Google Scholar] [CrossRef]
- Zhao, X.; Yuan, C.; Wangmo, D.; Subramanian, S. Tumor-Secreted Extracellular Vesicles Regulate T-Cell Costimulation and Can Be Manipulated To Induce Tumor-Specific T-Cell Responses. Gastroenterology 2021, 161, 560–574.e11. [Google Scholar] [CrossRef]
- Rezaei, R.; Baghaei, K.; Hashemi, S.M.; Zali, M.R.; Ghanbarian, H.; Amani, D. Tumor-Derived Exosomes Enriched by miRNA-124 Promote Anti-tumor Immune Response in CT-26 Tumor-Bearing Mice. Front. Med. 2021, 8, 619939. [Google Scholar] [CrossRef]
- Moradi Marjaneh, R.; Khazaei, M.; Ferns, G.A.; Avan, A.; Aghaee-Bakhtiari, S.H. MicroRNAs as potential therapeutic targets to predict responses to oxaliplatin in colorectal cancer: From basic evidence to therapeutic implication. IUBMB Life 2019, 71, 1428–1441. [Google Scholar] [CrossRef]
- Zhang, Y.; Bi, J.Y.; Huang, J.Y.; Tang, Y.N.; Du, S.Y.; Li, P.Y. Exosome: A review of its classification, isolation techniques, storage, diagnostic and targeted therapy applications. Int. J. Nanomed. 2020, 15, 6917–6934. [Google Scholar] [CrossRef]
- Mashouri, L.; Yousefi, H.; Aref, A.R.; Ahadi, A.M.; Molaei, F.; Alahari, S.K. Exosomes: Composition, biogenesis, and mechanisms in cancer metastasis and drug resistance. Mol. Cancer 2019, 18, 75. [Google Scholar] [CrossRef]
- Akao, Y.; Khoo, F.; Kumazaki, M.; Shinohara, H.; Miki, K.; Yamada, N. Extracellular Disposal of Tumor-Suppressor miRs-145 and -34a via Microvesicles and 5-FU Resistance of Human Colon Cancer Cells. Int. J. Mol. Sci. 2014, 15, 1392–1401. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.H.; Tian, D.; Yang, Z.C.; Li, J.L. Exosomal miR-21 promotes proliferation, invasion and therapy resistance of colon adenocarcinoma cells through its target PDCD4. Sci. Rep. 2020, 10, 8271. [Google Scholar] [CrossRef]
- Liang, G.; Zhu, Y.; Ali, D.J.; Tian, T.; Xu, H.; Si, K.; Sun, B.; Chen, B.; Xiao, Z. Engineered exosomes for targeted co-delivery of miR-21 inhibitor and chemotherapeutics to reverse drug resistance in colon cancer. J. Nanobiotechnol. 2020, 18, 10. [Google Scholar] [CrossRef]
- Yao, S.; Yin, Y.; Jin, G.; Li, D.; Li, M.; Hu, Y.; Feng, Y.; Liu, Y.; Bian, Z.; Wang, X.; et al. Exosome-mediated delivery of miR-204-5p inhibits tumor growth and chemoresistance. Cancer Med. 2020, 9, 5989–5998. [Google Scholar] [CrossRef]
- Senfter, D.; Holzner, S.; Kalipciyan, M.; Staribacher, A.; Walzl, A.; Huttary, N.; Krieger, S.; Brenner, S.; Jäger, W.; Krupitza, G.; et al. Loss of miR-200 family in 5-fluorouracil resistant colon cancer drives lymphendothelial invasiveness in vitro. Hum. Mol. Genet. 2015, 24, 3689–3698. [Google Scholar] [CrossRef] [Green Version]
- Holzner, S.; Senfter, D.; Stadler, S.; Staribacher, A.; Nguyen, C.H.; Gaggl, A.; Geleff, S.; Huttary, N.; Krieger, S.; Jäger, W.; et al. Colorectal cancer cell-derived microRNA200 modulates the resistance of adjacent blood endothelial barriers in vitro. Oncol. Rep. 2016, 36, 3065–3071. [Google Scholar] [CrossRef] [Green Version]
- Ren, D.; Lin, B.; Zhang, X.; Peng, Y.; Ye, Z.; Ma, Y.; Liang, Y.; Cao, L.; Li, X.; Li, R.; et al. Maintenance of cancer stemness by miR-196b-5p contributes to chemoresistance of colorectal cancer cells via activating STAT3 signaling pathway. Oncotarget 2017, 8, 49807–49823. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, X.; Du, L.; Wang, Y.; Liu, X.; Tian, H.; Wang, L.; Li, P.; Zhao, Y.; Duan, W.; et al. Exosome-Transmitted miR-128-3p increase chemosensitivity of oxaliplatin-resistant colorectal cancer. Mol. Cancer 2019, 18, 43. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Zhu, M. Novel exosomal miR-46146 transfer oxaliplatin chemoresistance in colorectal cancer. Clin. Transl. Oncol. 2020, 22, 1105–1116. [Google Scholar] [CrossRef]
- Ning, T.; Li, J.; He, Y.; Zhang, H.; Wang, X.; Deng, T.; Liu, R.; Li, H.; Bai, M.; Fan, Q.; et al. Exosomal miR-208b related with oxaliplatin resistance promotes Treg expansion in colorectal cancer. Mol. Ther. 2021, 29, 2723–2736. [Google Scholar] [CrossRef]
- Gu, Y.; Yu, J.; Zhang, J.; Wang, C. Suppressing the secretion of exosomal miR-19b by gw4869 could regulate oxaliplatin sensitivity in colorectal cancer. Neoplasma 2019, 66, 39–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Z.; Liu, Y.; Li, Q.; Liu, Q.; Liu, Y.; Luo, Y.; Wei, S. EVs delivery of miR-1915-3p improves the chemotherapeutic efficacy of oxaliplatin in colorectal cancer. Cancer Chemother. Pharmacol. 2021, 88, 1021–1031. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Sun, W.; Liu, R.; Zhou, Z.; Zhang, H.; Chen, X.; Ba, Y. Plasma Exosomal miRNA Expression Profile as Oxaliplatin-Based Chemoresistant Biomarkers in Colorectal Adenocarcinoma. Front. Oncol. 2020, 10, 1495. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Hosokawa, M.; Matsumura, J.; Matsubara, E.; Kobori, A.; Ueda, K.; Iwakawa, S. Effects of Zebularine on Invasion Activity and Intracellular Expression Level of let-7b in Colorectal Cancer Cells. Biol. Pharm. Bull. 2017, 40, 1320–1325. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, S.; Hosokawa, M.; Ueda, K.; Iwakawa, S. Effects of Decitabine on Invasion and Exosomal Expression of miR-200c and miR-141 in Oxaliplatin-Resistant Colorectal Cancer Cells. Biol. Pharm. Bull. 2015, 38, 1272–1279. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, S.; Hosokawa, M.; Miyamoto, T.; Nakagawa, A.; Haruna, M.; Ueda, K.; Iwakawa, S.; Ogawara, K.I. miR-33a-5p in small extracellular vesicles as non-invasive biomarker for oxaliplatin sensitivity in human colorectal cancer cells. Biochem. Biophys. Rep. 2021, 26, 100996. [Google Scholar] [CrossRef]
- Jin, G.; Liu, Y.; Zhang, J.; Bian, Z.; Yao, S.; Fei, B.; Zhou, L.; Yin, Y.; Huang, Z. A panel of serum exosomal microRNAs as predictive markers for chemoresistance in advanced colorectal cancer. Cancer Chemother Pharmacol. 2019, 84, 315–325. [Google Scholar] [CrossRef]
- Bigagli, E.; Luceri, C.; Guasti, D.; Cinci, L. Exosomes secreted from human colon cancer cells influence the adhesion of neighboring metastatic cells: Role of microRNA-210. Cancer Biol. Ther. 2016, 17, 1062–1069. [Google Scholar] [CrossRef] [Green Version]
- Yagi, T.; Iinuma, H.; Hayama, T.; Matsuda, K.; Nozawa, K.; Tsukamoto, M.; Shimada, R.; Akahane, T.; Tsuchiya, T.; Ozawa, T.; et al. Plasma exosomal microRNA-125b as a monitoring biomarker of resistance to mFOLFOX6-based chemotherapy in advanced and recurrent colorectal cancer patients. Mol. Clin. Oncol. 2019, 11, 416–442. [Google Scholar] [CrossRef] [Green Version]
- De Miguel Pérez, D.; Rodriguez Martínez, A.; Ortigosa Palomo, A.; Delgado Ureña, M.; Garcia Puche, J.L.; Robles Remacho, A.; Exposito Hernandez, J.; Lorente Acosta, J.A.; Ortega Sánchez, F.G.; Serrano, M.J. Extracellular vesicle-miRNAs as liquid biopsy biomarkers for disease identification and prognosis in metastatic colorectal cancer patients. Sci. Rep. 2020, 10, 3974. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.W.; Shi, Y.; Liu, J.B.; Wang, H.M.; Wang, P.Y.; Wu, Z.J.; Li, L.; Gu, L.P.; Cao, P.S.; Wang, G.R.; et al. Cancer-Associated fibroblast-derived exosomal microRNA-24-3p enhances colon cancer cell resistance to MTX by down-regulating CDX2/HEPH axis. J. Cell Mol. Med. 2021, 25, 3699–3713. [Google Scholar] [CrossRef]
- Sun, F.; Liang, W.; Qian, J. The identification of CRNDE, H19, UCA1 and HOTAIR as the key lncRNAs involved in oxaliplatin or irinotecan resistance in the chemotherapy of colorectal cancer based on integrative bioinformatics analysis. Mol. Med. Rep. 2019, 20, 3583–3596. [Google Scholar] [CrossRef] [Green Version]
- Hui, B.; Lu, C.; Wang, J.; Xu, Y.; Yang, Y.; Ji, H.; Li, X.; Xu, L.; Wang, J.; Tang, W.; et al. Engineered exosomes for co-delivery of PGM5-AS1 and oxaliplatin to reverse drug resistance in colon cancer. J. Cell Physiol. 2021, 1–23. [Google Scholar] [CrossRef]
- Yang, Y.N.; Zhang, R.; Du, J.W.; Yuan, H.H.; Li, Y.J.; Wei, X.L.; Du, X.X.; Jiang, S.L.; Han, Y. Predictive role of UCA1-containing exosomes in cetuximab-resistant colorectal cancer. Cancer Cell Int. 2018, 18, 164. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Liu, Y.; Zhang, Q.; Liu, B.; Cheng, Y.; Zhang, Y.; Sun, Y.; Liu, J.; Gen, H. Exosomal Long Non-coding RNA HOTTIP Increases Resistance of Colorectal Cancer Cells to Mitomycin via Impairing MiR-214-Mediated Degradation of KPNA3. Front. Cell Dev. Biol. 2021, 8, 582723. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, Q.; Bao, C.; Li, S.; Guo, W.; Zhao, J.; Chen, D.; Gu, J.; He, X.; Huang, S. Circular RNA is enriched and stable in exosomes: A promising biomarker for cancer diagnosis. Cell Res. 2015, 25, 981–984. [Google Scholar] [CrossRef] [Green Version]
- Seimiya, T.; Otsuka, M.; Iwata, T.; Shibata, C.; Tanaka, E.; Suzuki, T.; Koike, K. Emerging Roles of Exosomal Circular RNAs in Cancer. Front. Cell Dev. Biol. 2020, 8, 568366. [Google Scholar] [CrossRef]
- Xu, Y.; Qiu, A.; Peng, F.; Tan, X.; Wang, J.; Gong, X. Exosomal transfer of circular RNA FBXW7 ameliorates the chemoresistance to oxaliplatin in colorectal cancer by sponging miR-18b-5p. Neoplasma 2021, 68, 108–118. [Google Scholar] [CrossRef]
- Zhao, K.; Cheng, X.; Ye, Z.; Li, Y.; Peng, W.; Wu, Y.; Xing, C. Exosome-Mediated Transfer of circ_0000338 Enhances 5-Fluorouracil Resistance in Colorectal Cancer through Regulating MicroRNA 217 (miR-217) and miR-485-3p. Mol. Cell Biol. 2021, 41, e00517–e00520. [Google Scholar] [CrossRef]
- Hon, K.W.; Ab-Mutalib, N.S.; Abdullah, N.M.A.; Jamal, R.; Abu, N. Extracellular Vesicle-derived circular RNAs confers chemoresistance in Colorectal cancer. Sci. Rep. 2019, 9, 16497. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Tan, X.; Lu, Y. Exosomal transfer of circ_0006174 contributes to the chemoresistance of doxorubicin in colorectal cancer by depending on the miR-1205/CCND2 axis. J. Physiol. Biochem. 2021. Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Mulkeen, A.; Silva, T.; Schmitz, J.; Uchio, E.; Chu, E.; Cha, C. siRNA-mediated gene silencing in colorectal cancer: A novel anti-angiogenic targeted therapy. J. Surg. Res. 2004, 121, 279–280. [Google Scholar] [CrossRef]
- Aghamiri, S.; Jafarpour, A.; Malekshahi, Z.V.; Mahmoudi Gomari, M.; Negahdari, B. Targeting siRNA in colorectal cancer therapy: Nanotechnology comes into view. J. Cell Physiol. 2019, 234, 14818–14827. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Liu, R.X.; Chan, K.W.; Hu, J.; Zhang, J.; Wei, L.; Tan, H.; Yang, X.; Liu, H. Exosomal transfer of p-STAT3 promotes acquired 5-FU resistance in colorectal cancer cells. J. Exp. Clin. Cancer Res. 2019, 38, 320. [Google Scholar] [CrossRef]
- Lin, D.; Zhang, H.; Liu, R.; Deng, T.; Ning, T.; Bai, M.; Yang, Y.; Zhu, K.; Wang, J.; Duan, J.; et al. iRGD-Modified exosomes effectively deliver CPT1A siRNA to colon cancer cells, reversing oxaliplatin resistance by regulating fatty acid oxidation. Mol. Oncol. 2021, 15, 3430–3446. [Google Scholar] [CrossRef]
- Santos, P.; Almeida, F. Role of Exosomal miRNAs and the Tumor Microenvironment in Drug Resistance. Cells. 2020, 9, 1450. [Google Scholar] [CrossRef]
- Garcia-Mayea, Y.; Mir, C.; Masson, F.; Paciucci, R.; LLeonart, M.E. Insights into new mechanisms and models of cancer stem cell multidrug resistance. Semin. Cancer Biol. 2020, 60, 166–180. [Google Scholar] [CrossRef]
- Liu, C.; Liu, R.; Wang, B.; Lian, J.; Yao, Y.; Sun, H.; Zhang, C.; Fang, L.; Guan, X.; Shi, J.; et al. Blocking IL-17A enhances tumor response to anti-PD-1 immunotherapy in microsatellite stable colorectal cancer. J. Immunother Cancer 2021, 9, e001895. [Google Scholar] [CrossRef]
- Jiang, W.; Pan, S.; Chen, X.; Wang, Z.W.; Zhu, X. The role of lncRNAs and circRNAs in the PD-1/PD-L1 pathway in cancer immunotherapy. Mol. Cancer 2021, 20, 116. [Google Scholar] [CrossRef]
- Dong, H.; Wang, W.; Chen, R.; Zhang, Y.; Zou, K.; Ye, M.; He, X.; Zhang, F.; Han, J. Exosome-mediated transfer of lncRNA SNHG14 promotes trastuzumab chemoresistance in breast cancer. Int. J. Oncol. 2018, 53, 1013–1026. [Google Scholar] [CrossRef] [Green Version]
- Jin, K.T.; Yao, J.Y.; Fang, X.L.; Di, H.; Ma, Y.Y. Roles of lncRNAs in cancer: Focusing on angiogenesis. Life Sci. 2020, 252, 117647. [Google Scholar] [CrossRef]
- Sun, T.; Yin, Y.F.; Jin, H.G.; Liu, H.R.; Tian, W.C. Exosomal microRNA-19b targets FBXW7 to promote colorectal cancer stem cell stemness and induce resistance to radiotherapy. Kaohsiung J. Med. Sci. 2021. Online ahead of print. [Google Scholar] [CrossRef]
- Chen, X.; Liu, J.; Zhang, Q.; Liu, B.; Cheng, Y.; Zhang, Y.; Sun, Y.; Ge, H.; Liu, Y. Exosome-mediated transfer of miR-93-5p from cancer-associated fibroblasts confer radioresistance in colorectal cancer cells by downregulating FOXA1 and upregulating TGFB3. J. Exp. Clin. Cancer Res. 2020, 39, 65. [Google Scholar] [CrossRef]
- Rufino-Ramos, D.; Albuquerque, P.R.; Carmona, V.; Perfeito, R.; Nobre, R.J.; de Almeida, L.P. Extracellular vesicles: Novel promising delivery systems for therapy of brain diseases. J. Control. Release 2017, 262, 247–258. [Google Scholar] [CrossRef]
- Li, Z.; Niu, H.; Qin, Q.; Yang, S.; Wang, Q.; Yu, C.; Wei, Z.; Jin, Z.; Wang, X.; Yang, A.; et al. lncRNA UCA1 Mediates Resistance to Cisplatin by Regulating the miR-143/FOSL2-Signaling Pathway in Ovarian Cancer. Mol. Ther. Nucleic Acids 2019, 17, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Yin, K.; Wang, S.; Zhao, R.C. Exosomes from mesenchymal stem/stromal cells: A new therapeutic paradigm. Biomark Res. 2019, 7, 8. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
| EV/Exosome Content | Expression | Type of EVs | EV Source | Major Findings | Potential Clinical Application | Drug | Ref. |
|---|---|---|---|---|---|---|---|
| miR-92a-3p | ↑ | exosomes | CAFs isolated from tissues and serum samples | (i) CAF-derived exosomes transfer miR-92a-3p enhancing cell stemness, EMT, metastasis and chemoresistance (ii) Overexpression of exosomal miR-92a-3p in serum is highly linked with metastasis and chemoresistance | Prognostic value/Therapy monitoring biomarker | 5-FU/OX | [3] |
| miR-424 | ↑ | EVs | Tumor-derived EVs (human and mouse tumor samples, cell lines, human colorectal cancer organoids, and mouse models) | (i) EVs containing miR-424 were associated with immune checkpoint blockade resistance (ii) Modified tumor-secreted EVs ↓miR-424 boost the immune checkpoint blockade efficacy in late-stage disease | Therapeutic target | ICIs | [56] |
| miR-34a and miR-145 | ↑ | EVs | Cell lines (DLD-1 and DLD-1/5-FU) | Selected secretion of miR-34a and miR-145, following 5-FU exposure results in decreased intracellular levels of tumor suppressor miRNAs (such as miR-34a and miR-145) | Therapeutic potential/Therapy monitoring biomarker | 5-FU | [61] |
| miR-21 | ↑ | exosomes | Cell lines (HT29, T84, and LS174) | targeted secretion of exosomes carrying miR-21, may represent an effective anti-cancer strategy | Therapeutic target | 5-FU | [62] |
| miR-21 inhibitor | - | purified engineered exosomes | Cell lines (THLG-293T, LG-293T and GFP-293T) | miR-21i and 5-FU co-delivery reverses drug resistance and enhances cytotoxicity | Therapeutic potential | 5-FU | [63] |
| miR-204-5p | ↑ | exosomes | Cell lines (293T-GFP and 293T-miR-204 cells) | Exosomal miR-204-5p induces apoptosis, inhibits cell proliferation, and increases chemosensitivity | Therapeutic potential | 5-FU | [64] |
| miR-200 family | ↓ | exosomes | Cell line (CCL227 cells) | Loss of miR-200 family is associated with lymphendothelial invasiveness and chemoresistance | Prognostic value/Therapeutic potential | 5-FU | [65] |
| miR200 | ↓ | exosomes | Cell lines (CCL227) | Exosomes devoid of miR200 in 5-FU resistant cells are associated with the induction of the metastatic process | N/A | 5-FU | [66] |
| miR-196b-5p | ↑ | exosomes | Serum | (i) miR-196b-5p overexpression is associated with poor survival (ii) Silencing miR-196b-5p enhances 5-FU-induced apoptosis (iii) miR-196b-5p downregulation reverses chemoresistance | Prognostic value/therapy monitoring biomarker/therapeutic target | 5-FU | [67] |
| miR-128-3p | ↑ | exosomes | Cell line [normal intestinal epithelial (FHC) cells] | (i) miR-128-3p suppresses EMT and increases intracellular drug accumulation (ii) miR-128-3p downregulation is associated with poor response in advanced CRC (iii) miR-128-3p overexpression inhibits EMT and decreased drug efflux | Prognostic value/therapy monitoring biomarker/therapeutic potential | OX | [68] |
| miR-46146 | ↑ | exosomes | Cell lines | Exosomal miR-46146 promotes resistance | Therapy monitoring biomarker/therapeutic target | OX | [69] |
| miR-208b | ↑ | exosomes | Cell lines (NCM460, SW480, SW480-OXA cells) | (i) Exosomal miR-208b is overexpressed in oxaliplatin resistant cell lines (ii) Exosomal miR-208b is delivered into recipient T cells, promoting Tregs | Predictive biomarker/therapeutic target | OX | [70] |
| miR-19b | ↑ | exosomes | Cell line (SW480 cells) | Suppressing exosomal miR-19b release leads to increased sensitivity | Therapeutic potential | OX | [71] |
| miR-1915-3p | ↑ | EVs | Non-tumorigenic intestinal cell line (FHC) | Exosomal delivery of miR-1915-3p improves chemotherapeutic effect | Therapeutic potential | OX | [72] |
| miR-100, miR-92a, miR-16, miR-30e, miR-144-5p, and let-7i | ↑ | exosomes | Plasma | A panel of 6 exosomal miRNAs can significantly distinguish chemoresistant from chemosensitive patients | Therapy monitoring biomarker/therapeutic potential | OX | [73] |
| Let 7b | ↑ | exosomes | Cell lines (SW480, SW620, and SW620/OxR). | (i) Oxaliplatin resistance may inhibit the incorporation of let-7a, let-7b, and let-7g into exosomes (ii) Exosomal let-7a, let-7b, and let-7g are downregulated in resistant cells | Therapy monitoring biomarker | OX | [74] |
| miR-200c and miR-141 | ↑ | exosomes | Cell lines (SW620/OxR, SW480, SW620) | Up-regulated miR-200c and miR-141 following decitabine treatment was associated with mesenchymal-epithelial transition of oxaliplatin -resistant CRC cells | Predictive biomarker | OX | [75] |
| miR-33a-5p, miR-210–3p | ↓ | small EVs (sEVs) | Cell lines (SW620, SW480 and SW620-OxR cells) | miR-33a-5p and miR-210–3p are under-expressed in oxaliplatin-resistant cells | Therapy monitoring biomarker | OX | [76] |
| miR-21-5p, miR-1246, miR-1229-5p, miR-135b, miR-425 and miR-96-5p | ↑ | exosomes | Serum | This panel of exosomal miRNAs could significantly distinguish chemotherapy-resistant patients | Therapy monitoring biomarker/therapeutic target | 5-FU/OX | [77] |
| miR-210 | ↑ | exosomes | Cell line (HCT-8 cells) | (i) Exosomal miR-210 is associated with significantly lower chemosensitivity (ii) Exosomal miR-210 promotes EMT and metastasis | Predictive biomarker/therapeutic target | 5-FU, FOLFOX-like treatment (5FU + Oxaliplatin) | [78] |
| miR-125b | ↑ | exosomes | Plasma | (i) Exosomal miR-125b has been correlated with chemoresistance (ii) Exosomal miR-125b before chemotherapy is a predictive biomarker for PFS | Predictive biomarker/therapy monitoring biomarker | mFOLFOX6 | [79] |
| miR-92a | ↑ | EVs | Serum EVs isolated from 44 mCRC patients | (i) Overexpression of miR-92a was associated with shorter PFS (close but not statistically significant) | Further investigation is needed | Bevacizumab + FOLFOX-6m (5FU,LV,OX) | [80] |
| miRNA-24-3p | ↑ | exosomes | miR-24-3p inhibitor-treated CAFs (CRC tissues and cells) | (i) CAF-derived exosomal miR-24-3p was associated with chemoresistance (ii) CAF-derived exosomal miR-24-3p inhibitor enhanced cell apoptosis in vitro and inhibited tumor growth in vivo | Therapy monitoring biomarker/therapeutic target | MTX | [81] |
| EV/Exosome Content | Expression | Type of EVs | EV Source | Major Finding | Potential (Clinical) Application | Drug | Ref. |
|---|---|---|---|---|---|---|---|
| H19 | ↑ | exosomes | Tumor tissues, normal tissues and cell lines (HCT116 and SW480) | (i) H19 is overexpressed in CAF-derived exosomes (ii) CAFs-derived exosomal H19 promotes stemness and chemoresistance | Prognostic value/therapeutic target | OX | [41] |
| CCAL | ↑ | exosomes | CAFs isolated from human CRC tissues and NFs | CAF-derived exosomal CCAL is transferred to cancer cells inhibiting apoptosis and conferring chemoresistance | Therapeutic monitoring/therapeutic target | OX | [42] |
| CRNDE, H19, UCA1 and HOTAIR | ↑ | bioinformatic analysis | - | integrated analysis showed that several differentially expressed genes of lncRNAs are components of the extracellular exosomes | Further investigation is needed | OΧ, IRI | [82] |
| PGM5-AS1 | ↓ | exosomes | Tumor tissues | (i) PGM5-AS1 hinders proliferation, metastasis, and acquired oxaliplatin resistance of colon cancer cells (ii) Exosomes encapsulating oxaliplatin and PGM5-AS1 can reverse drug resistance | Therapeutic target | OX | [83] |
| UCA1 | ↑ | exosomes | Serum samples and cell lines (Caco2) | (i) UCA1 was overexpressed in cetuximab-resistant cancer cells and their exosomes (ii) Circulating exosomal UCA1 was significantly overexpressed in progressive disease/stable disease than in the partial response/complete response patients. (iii) Exosomal transfer of UCA1 transmits cetuximab resistance from resistant cells to sensitive ones | Therapy monitoring biomarker/prognostic biomarker/therapeutic target | cetuximab | [84] |
| HOTTIP | ↑ | EVs | Serum and cell lines (HCT116, SW620, FHC, LoVo, HT29, SW480, SW1116, and Caco2) | (i) HOTTIP was overexpressed in mitomycin-resistant CRC cells and its inhibition reduced resistance (ii) EV-transferred HOTTIP from mitomycin-resistant cells contributes to mitomycin resistance | Predictive value/therapeutic target | mitomycin | [85] |
| EV/Exosome Content | Expression | Type of EVs | EV Source | Major Finding | Potential (Clinical) Application | Drug | Ref. |
|---|---|---|---|---|---|---|---|
| hsa_circ_0005963 | ↑ | exosomes | Serum and cell lines (SW480, HCT116, and HEK293) | (i) Circular RNA hsa_circ_0005963 was associated with chemoresistance (ii) Exosomal hsa_circ_0005963 from oxaliplatin-resistant cells confers resistance to sensitive ones (ii) Exosomal transport of si-ciRS-122 suppresses glycolysis and reverses resistance | Therapy monitoring biomarker/therapeutic target | OX | [45] |
| FBXW7 | ↓ | exosomes | FHC cell culture | (i) circ-FBXW7 was decreased in oxaliplatin-resistant CRC patients and cells. (ii) Exosomal circ-FBXW7 delivery can improve chemoresistance | Therapeutic potential | OX | [88] |
| circ_0000338 | ↑ | exosomes | Cell lines (SW480/5-FU and HCT116/5-FU) | (i) Circ_0000338 was upregulated in 5-FU-resistant CRC tissues and cells, and its knockdown reversed 5-FU resistance (ii) Exosome-mediated delivery of circ_0000338 confers chemoresistance to sensitive cells | Therapy monitoring biomarker/therapeutic target | 5-FU | [89] |
| hsa_circ_0032883, hsa_circ_0066629, hsa_circ_0002039, and hsa_circ_0000338 | ↑ | exosomes | FOLFOX-resistant HCT116-R cells | (i) 105 upregulated and 34 downregulated circRNAs in exosomes from FOLFOX-resistant cells (ii) Drug resistance can be conferred from resistant cells to sensitive ones via exosomes (ii) Exosomal hsa_circ_0000338 was differentially upregulated in chemoresistant cells | Therapy monitoring biomarker | 5-FU, OX | [90] |
| circ_0006174 | ↑ | exosomes | Tissue samples | (i) Circ_0006174 was overexpressed in doxorubicin-resistant cells and its downregulation reversed resistance and metastatic potential (ii) Exosomal transfer of circ_0006174 enhances chemoresistance | Therapy monitoring biomarker/therapeutic target | DOX | [91] |
| EV/Exosome Content | Expression | Type of EVs | EV Source | Major Finding | Potential (Clinical) Application | Drug | Ref. |
|---|---|---|---|---|---|---|---|
| GSTP1 and STAT3 siRNAs | - | exosomes | Cell lines (RKO/R and RKO/P) | p-STAT3 transferred by exosomes from 5-FU-resistant cells confers chemoresistance | Therapeutic potential | 5-FU | [94] |
| CPT1A siRNA | - | exosomes | Tumor Tissue | (i) Silencing CPT1A by siRNA could reverse chemoresistance and combined delivery of oxaliplatin with CPT1A inhibitor promotes apoptosis and proliferation (ii) iRGD-engineered exosomes with siCPT1A suppress fatty acid oxidation and thus reverse chemoresistance | Therapeutic potential | OX | [95] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lampropoulou, D.I.; Pliakou, E.; Aravantinos, G.; Filippou, D.; Gazouli, M. The Role of Exosomal Non-Coding RNAs in Colorectal Cancer Drug Resistance. Int. J. Mol. Sci. 2022, 23, 1473. https://doi.org/10.3390/ijms23031473
Lampropoulou DI, Pliakou E, Aravantinos G, Filippou D, Gazouli M. The Role of Exosomal Non-Coding RNAs in Colorectal Cancer Drug Resistance. International Journal of Molecular Sciences. 2022; 23(3):1473. https://doi.org/10.3390/ijms23031473
Chicago/Turabian StyleLampropoulou, Dimitra Ioanna, Evangelia Pliakou, Gerasimos Aravantinos, Dimitrios Filippou, and Maria Gazouli. 2022. "The Role of Exosomal Non-Coding RNAs in Colorectal Cancer Drug Resistance" International Journal of Molecular Sciences 23, no. 3: 1473. https://doi.org/10.3390/ijms23031473
APA StyleLampropoulou, D. I., Pliakou, E., Aravantinos, G., Filippou, D., & Gazouli, M. (2022). The Role of Exosomal Non-Coding RNAs in Colorectal Cancer Drug Resistance. International Journal of Molecular Sciences, 23(3), 1473. https://doi.org/10.3390/ijms23031473