
Fine Mapping and Candidate Gene Prediction of Tuber Shape Controlling Ro Locus Based on Integrating Genetic and Transcriptomic Analyses in Potato

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
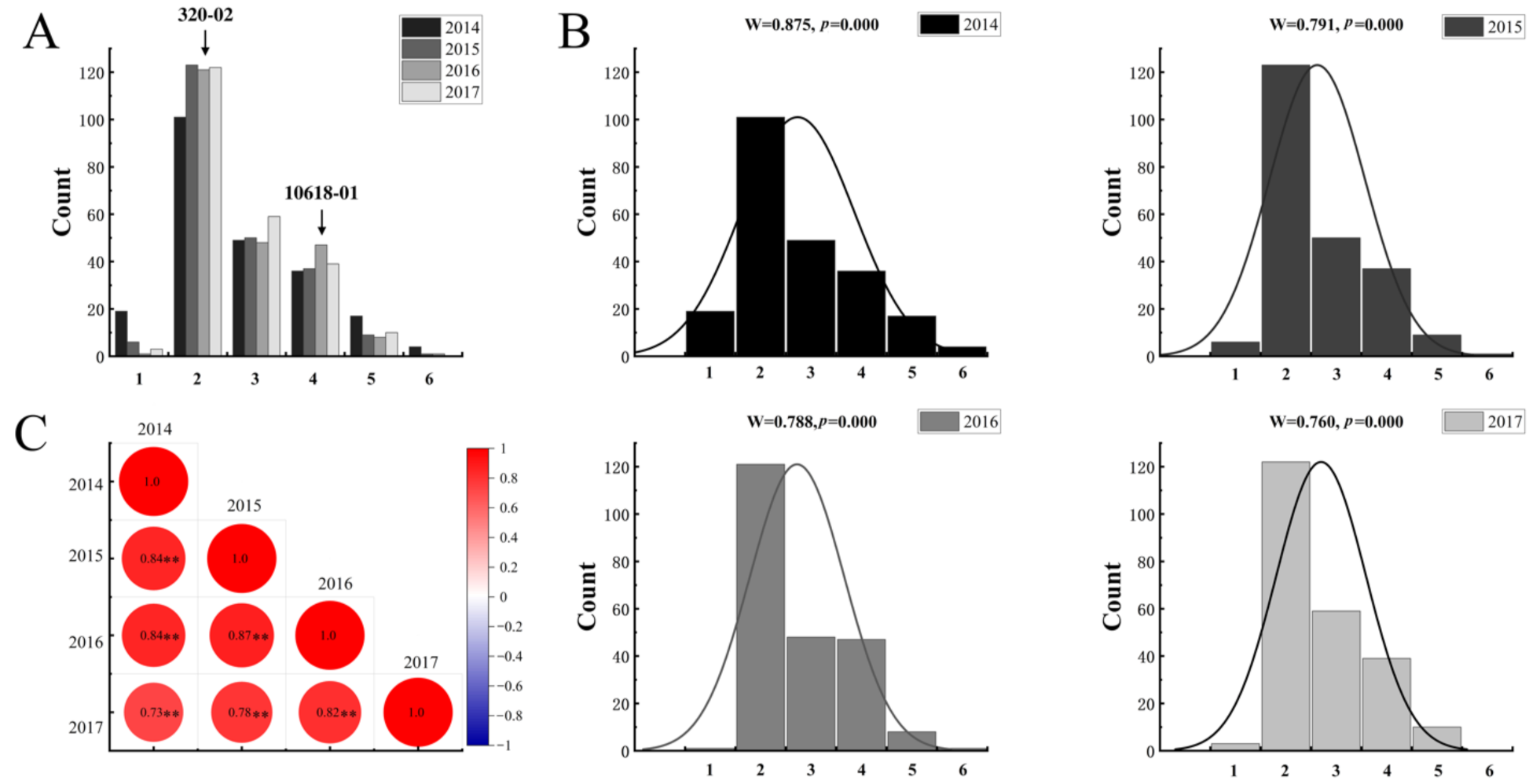
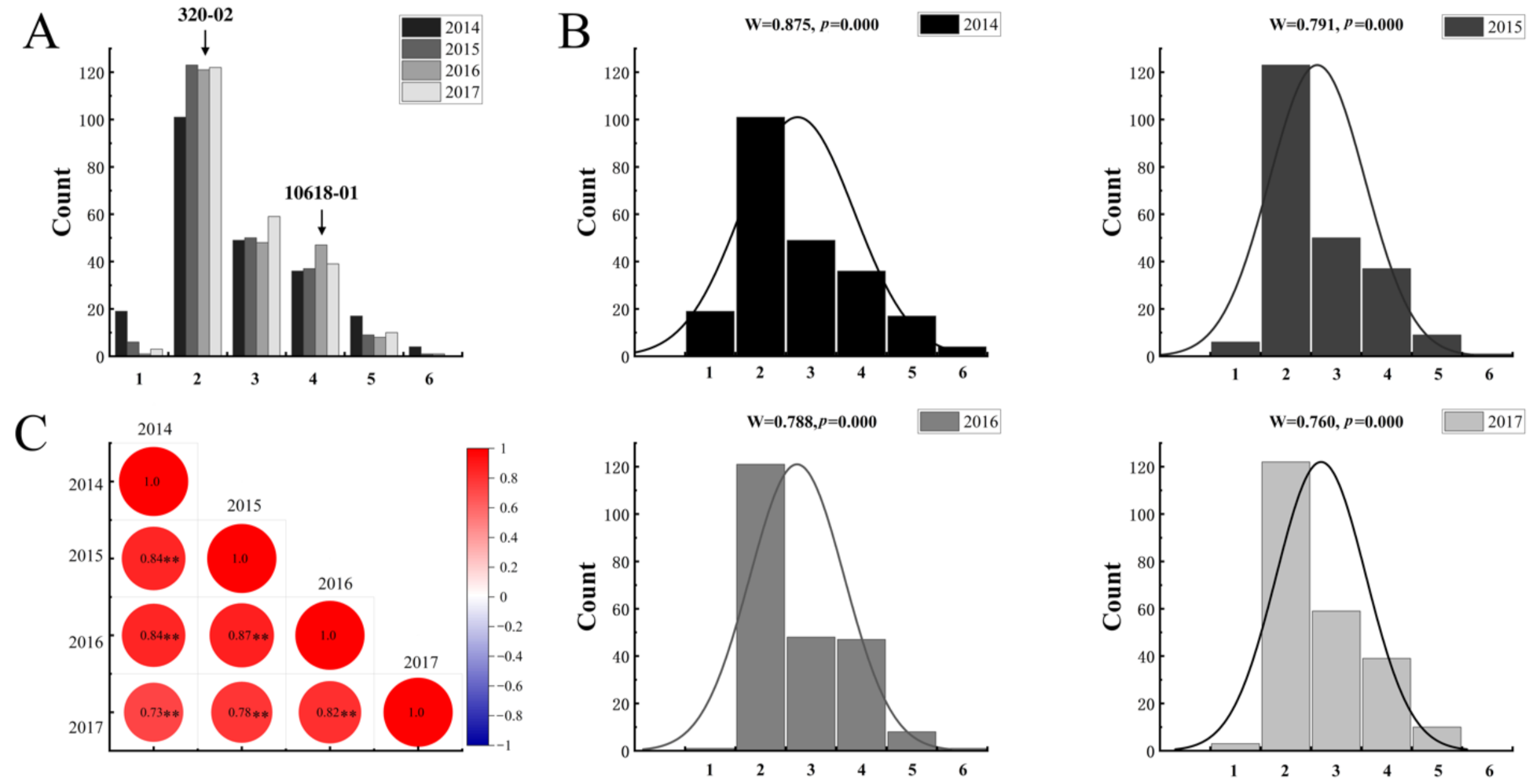
2.1. Segregation of Tuber Shape in the Mapping Population
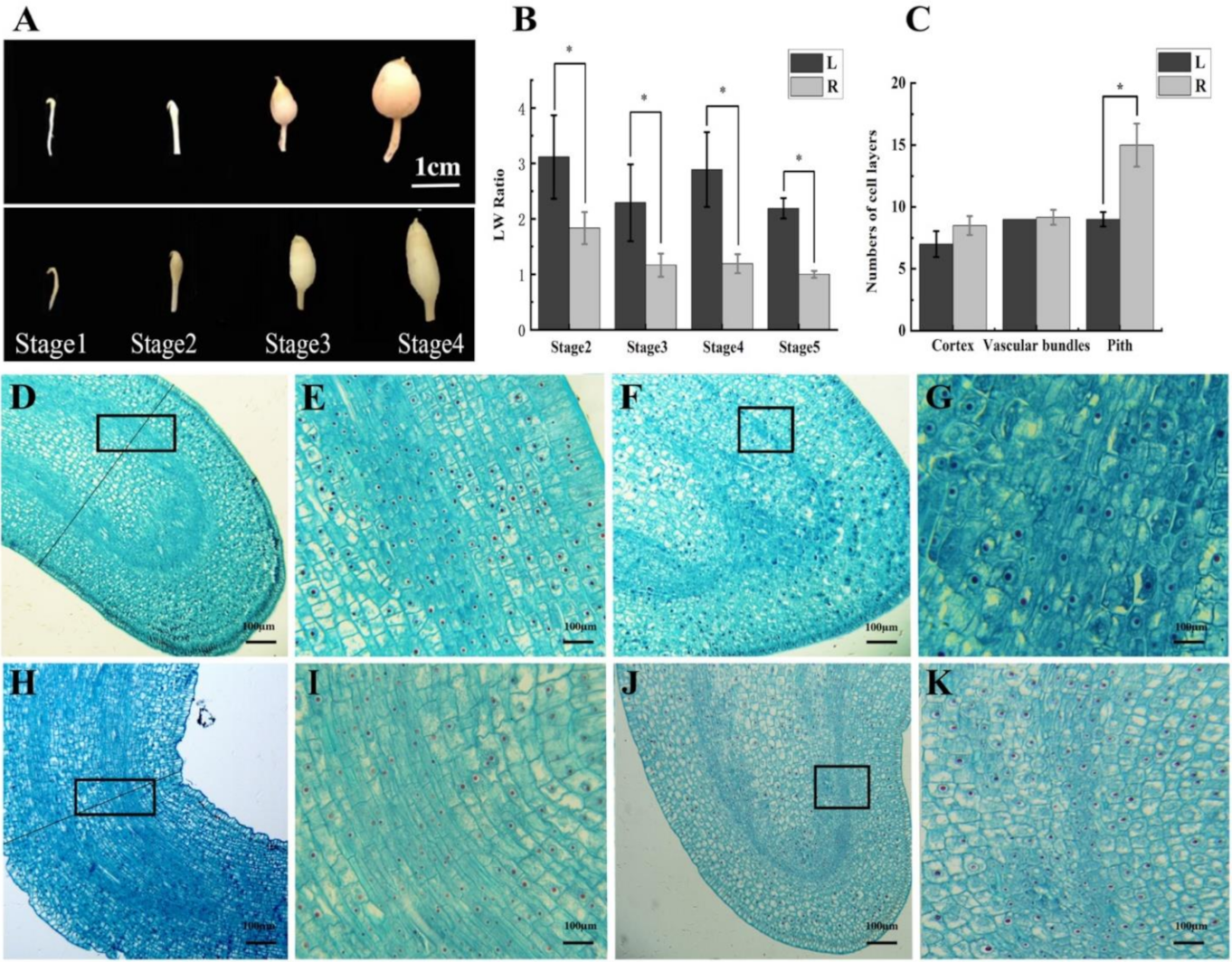
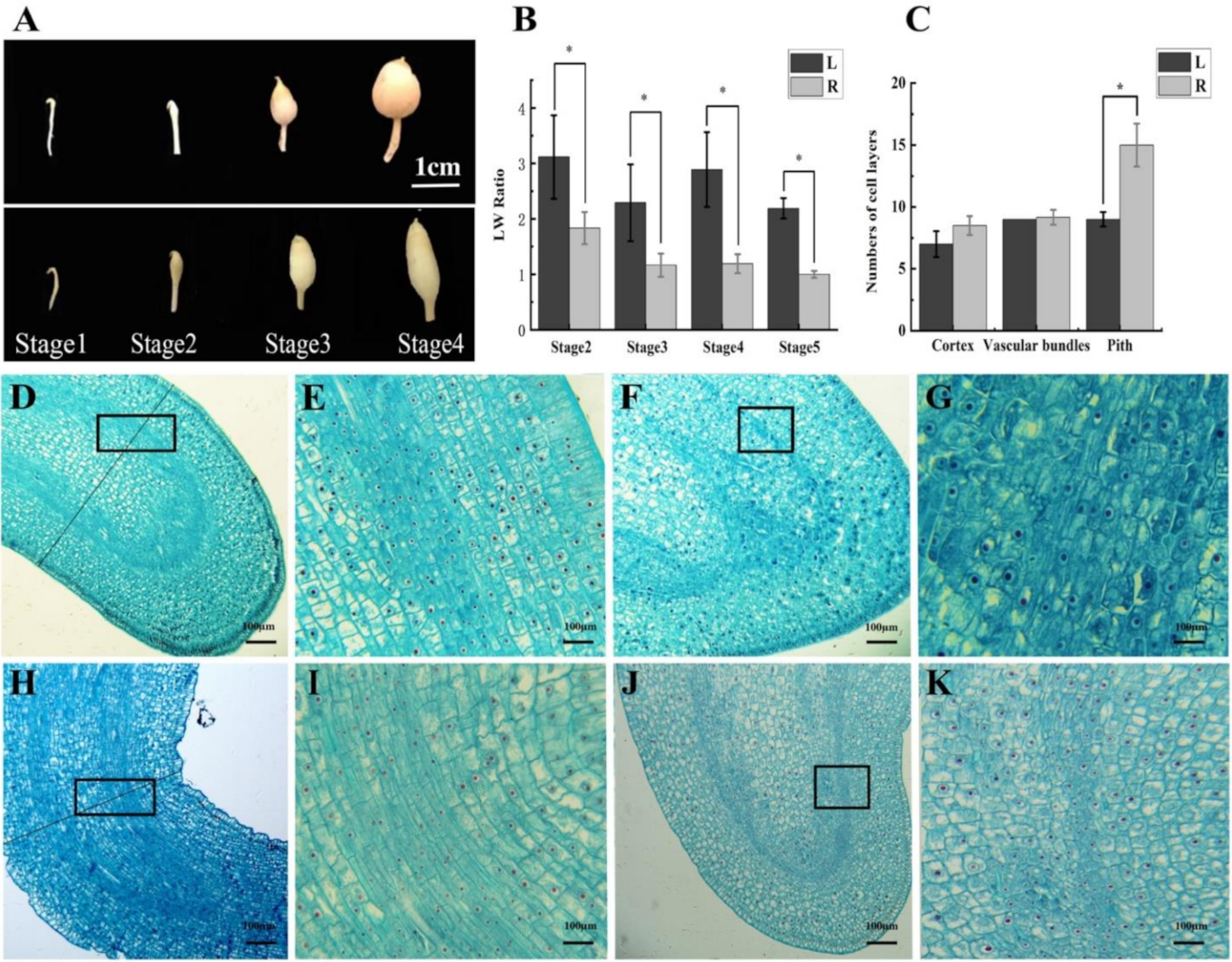
2.2. The Cytological Study Demonstrates the Association between the Width of Tuber and Narrowness of the Pith
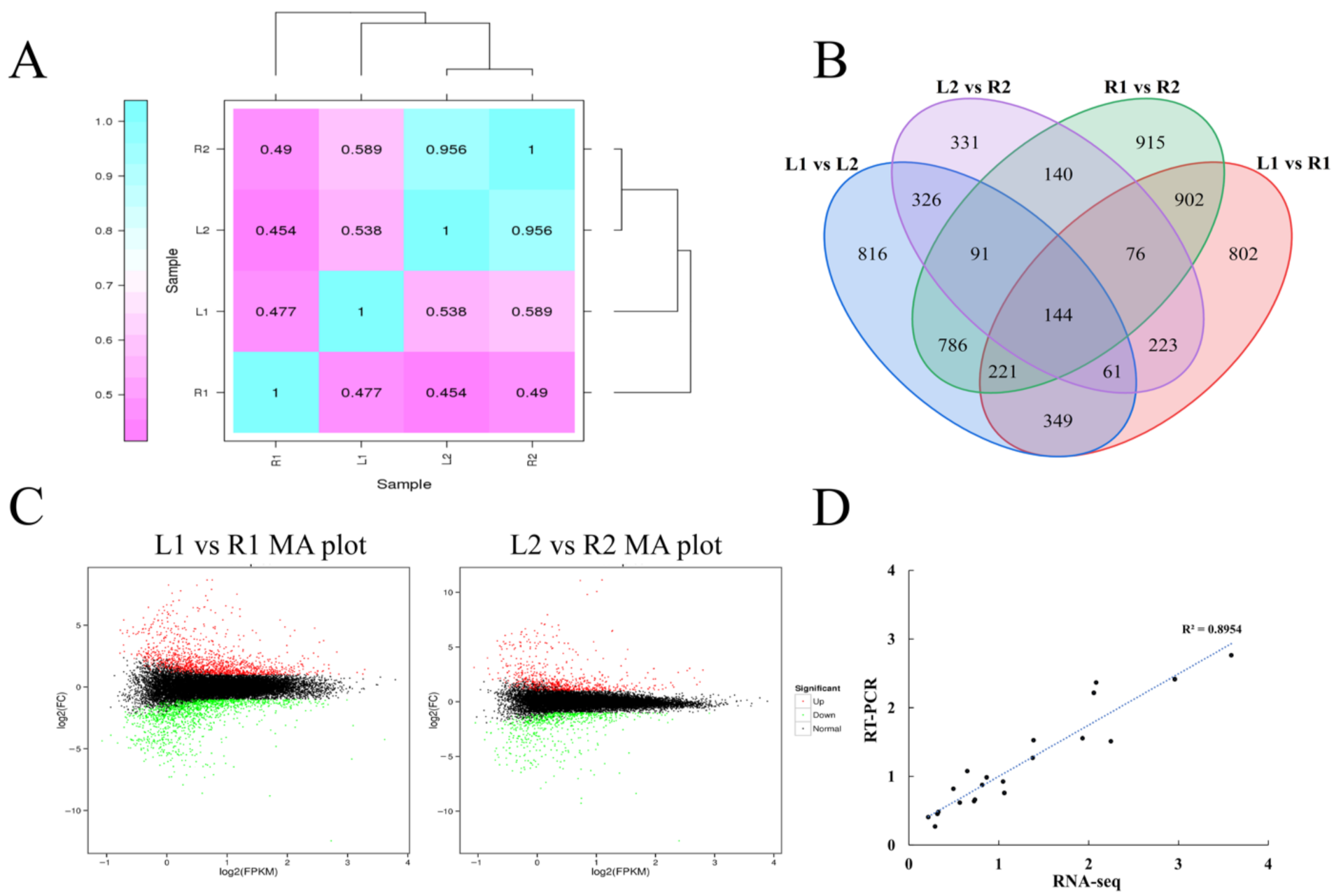
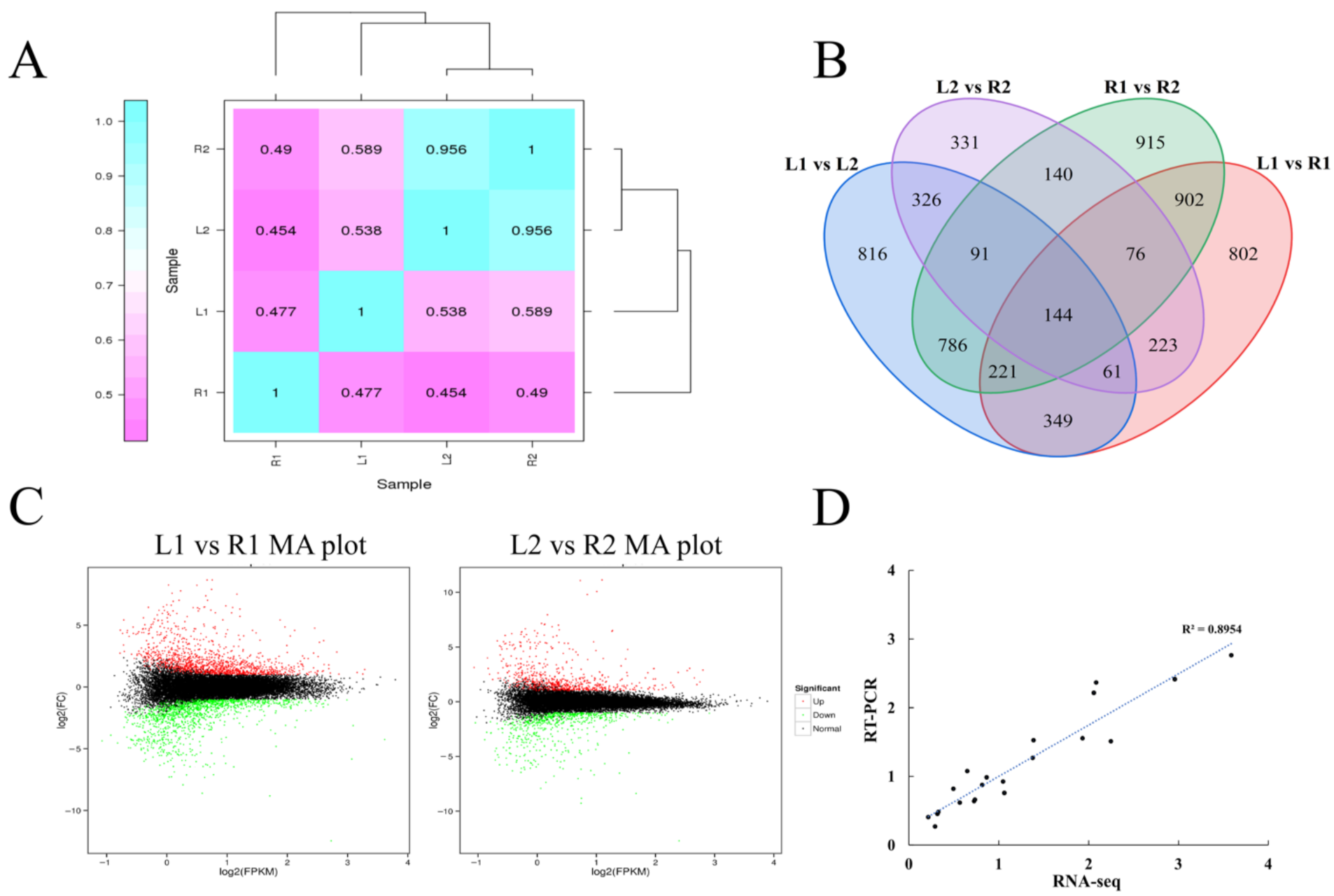
2.3. Patterns of Differential Gene Expression Reveal That Tuber Shape Determination Occurred in the Early Stage of Tuber Development
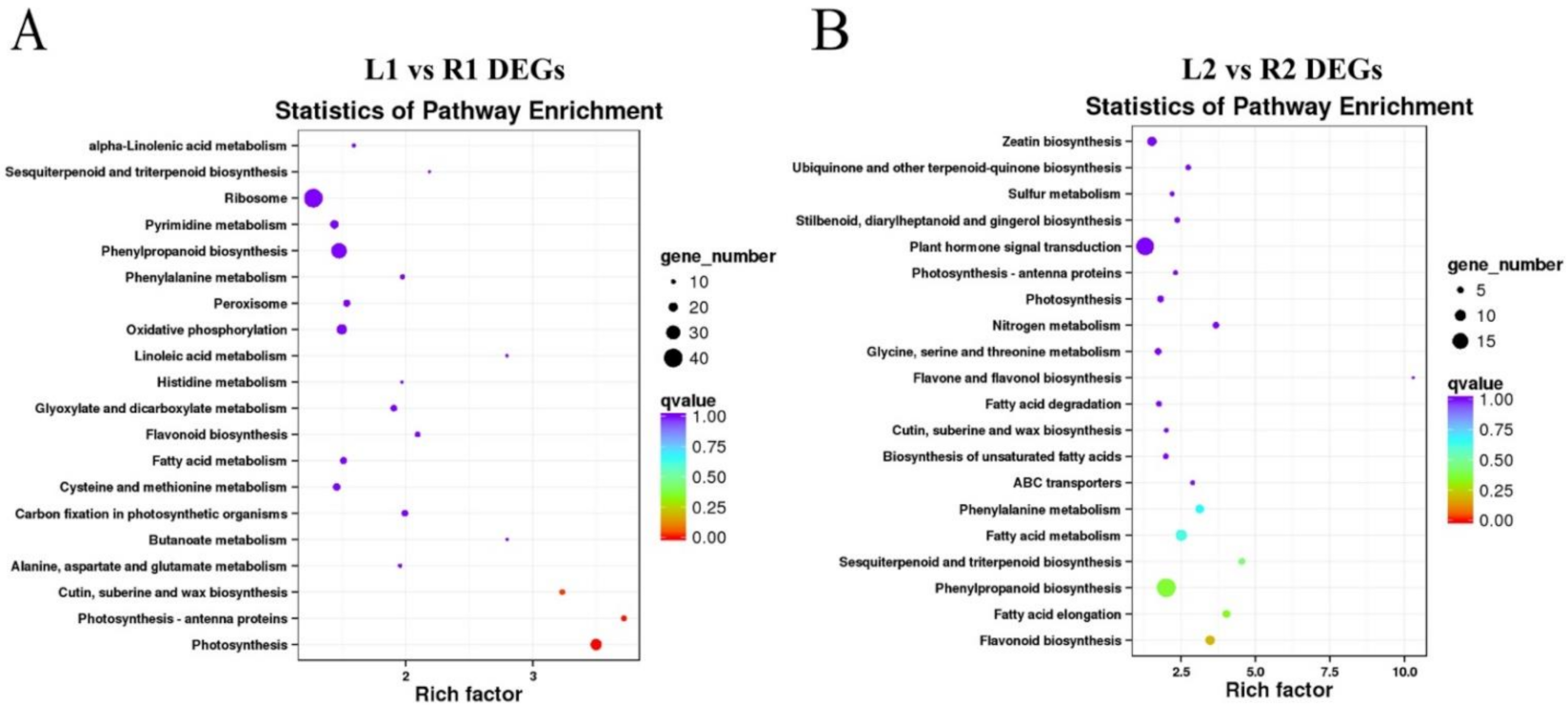
2.4. Functional Classification of DEGs Showed That Tuber Shape May Be Related to Auxin and Cell Wall Formation Pathways
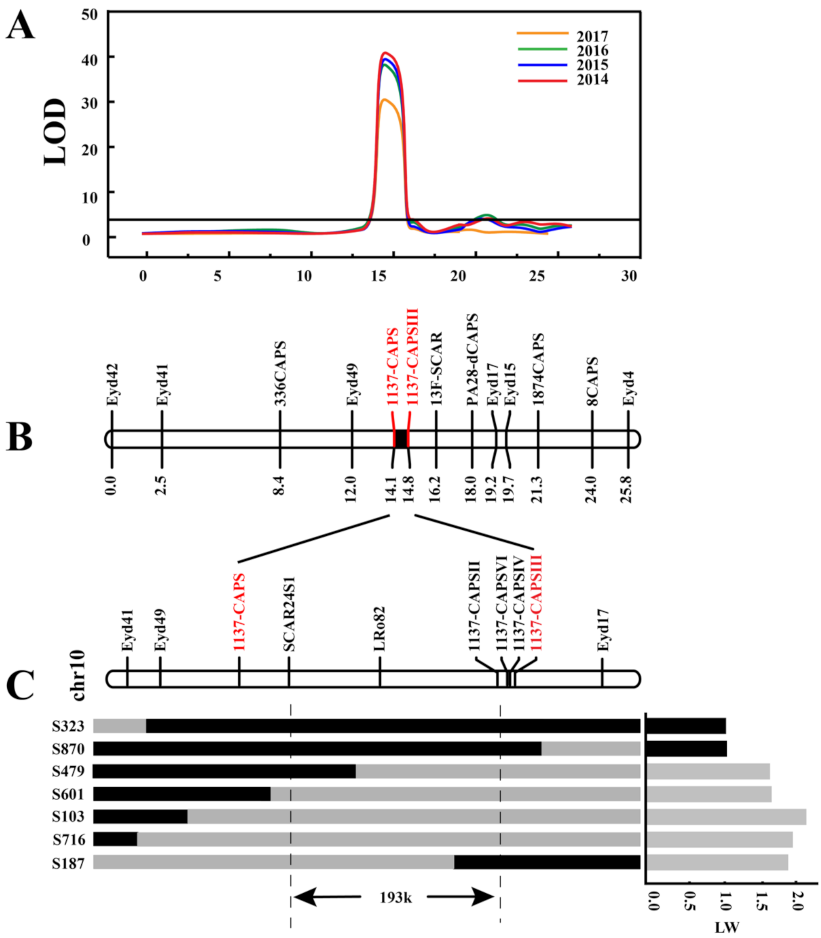
2.5. Fine Mapping of Ro in the Defined Genomic Region
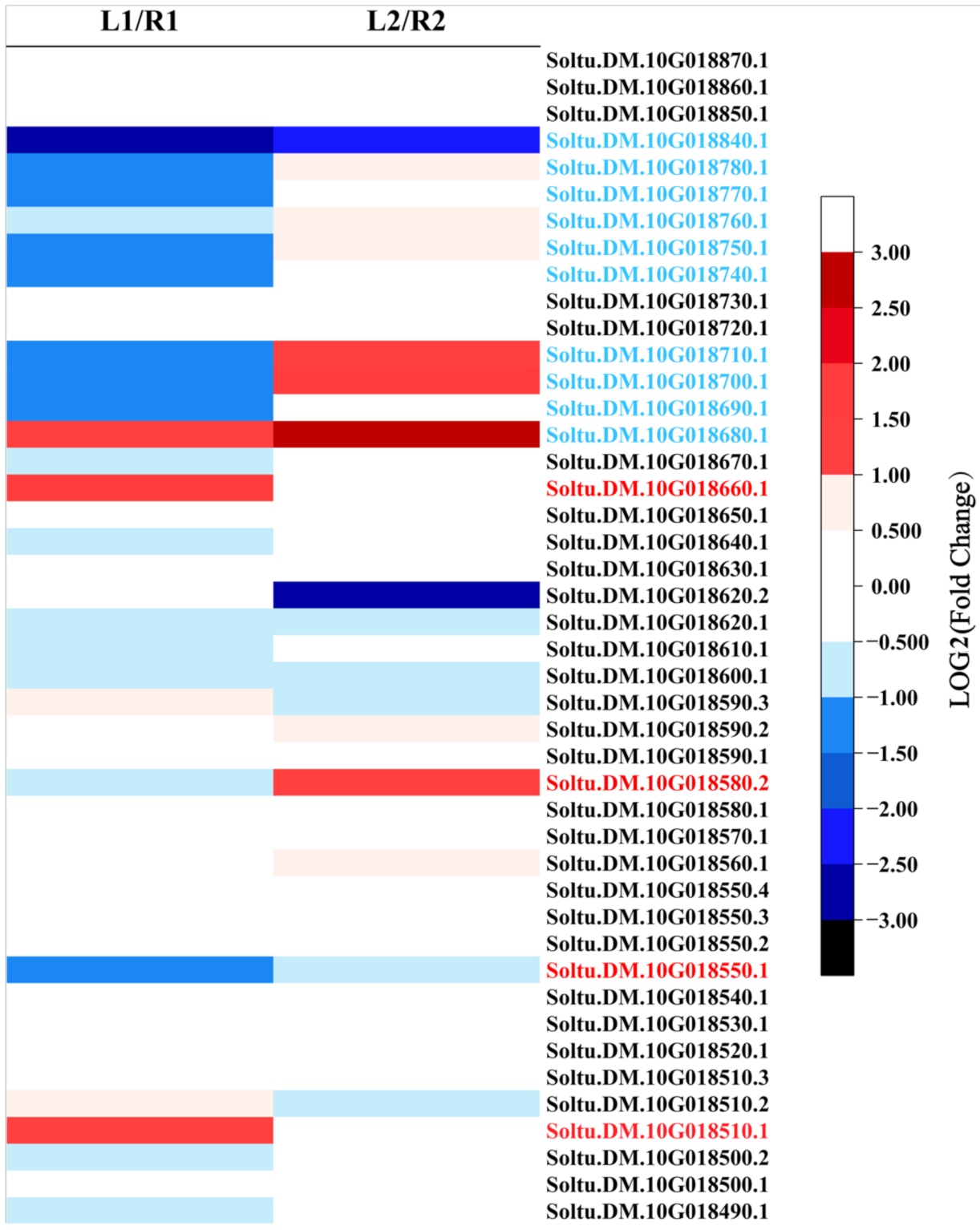
2.6. Five Differentially Expressed Genes Are Pinpointed as Candidate Genes Involved in Tuber Shape Formation
3. Discussion
3.1. Characterization of the Mapping Population
3.2. Mapping of the Ro Gene
3.3. Candidate Gene Prediction
4. Materials and Methods
4.1. Plant Material
4.2. Potato Shape Evaluation and Analysis
4.3. Tissue and Cell Observation
4.4. RNA-seq Analysis
4.5. Molecular Marker Development
4.6. Genetic Linkage Map Construction and QTL Mapping
4.7. Identification of the Candidate Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hirsch, C.N.; Hirsch, C.D.; Felcher, K.; Coombs, J.; Zarka, D.; Deynze, A.V.; Jong, W.D.; Veilleux, R.E.; Jansky, S.; Bethke, P. Retrospective View of North American Potato (Solanum tuberosum L.) Breeding in the 20th and 21st Centuries. G3 Genes Genomes Genet. 2013, 3, 1003–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, Y.; Sankaran, S.; Knowles, N.R.; Pavek, M.J. Potato Tuber Length-Width Ratio Assessment Using Image Analysis. Am. J. Potato Res. 2016, 94, 88–93. [Google Scholar] [CrossRef]
- Van Eck, H.J.; Jacobs, J.; Stam, P.; Ton, J.; Stiekema, W.J.; Jacobsen, E. Multiple alleles for tuber shape in diploid potato detected by qualitative and quantitative genetic analysis using RFLPs. Genetics 1994, 137, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Tang, D.; Huang, W.; Yang, Z.; Huang, S. Haplotype-resolved genome analyses of a heterozygous diploid potato. Nat. Genet. 2020, 52, 1018–1023. [Google Scholar] [CrossRef] [PubMed]
- Jong, H.D.; Burns, V.J. Inheritance of tuber shape in cultivated diploid potatoes. Am. Potato J. 1993, 70, 267–284. [Google Scholar] [CrossRef]
- Spooner, D.M.; Ghislain, M.; Simon, R.; Jansky, S.H.; Gavrilenko, T. Systematics, Diversity, Genetics, and Evolution of Wild and Cultivated Potatoes. Bot. Rev. 2014, 80, 283–383. [Google Scholar] [CrossRef]
- De Jong, H.; Rowe, P. Genetic markers in inbred clones of cultivated diploid potatoes. Potato Res. 1972, 15, 200–208. [Google Scholar] [CrossRef]
- Gebhardt, C.; Ritter, E.; Debener, T.; Schachtschabel, U.; Walkemeier, B.; Uhrig, H.; Salamini, F. RFLP analysis and linkage mapping in Solanum tuberosum. Theor. Appl. Genet. 1989, 78, 65–75. [Google Scholar] [CrossRef]
- Li, X.Q.; Jong, H.D.; Jong, D.M.D.; Jong, W.S.D. Inheritance and genetic mapping of tuber eye depth in cultivated diploid potatoes. Theor. Appl. Genet. 2005, 110, 1068–1073. [Google Scholar] [CrossRef]
- Śliwka, J.; Wasilewicz-Flis, I.; Jakuczun, H.; Gebhardt, C. Tagging quantitative trait loci for dormancy, tuber shape, regularity of tuber shape, eye depth and flesh colour in diploid potato originated from six Solanum species. Plant Breed. 2008, 127, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Prashar, A.; Hornyik, C.; Young, V.; McLean, K.; Sharma, S.K.; Dale, M.F.B.; Bryan, G.J. Construction of a dense SNP map of a highly heterozygous diploid potato population and QTL analysis of tuber shape and eye depth. Theor. Appl. Genet. 2014, 127, 2159–2171. [Google Scholar] [CrossRef] [PubMed]
- Lindqvist-Kreuze, H.; Khan, A.; Salas, E.; Meiyalaghan, S.; Bonierbale, M. Tuber shape and eye depth variation in a diploid family of Andean potatoes. BMC Genet. 2015, 16, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara-Skrzypiec, A.; Sliwka, J.; Jakuczun, H.; Zimnoch-Guzowska, E. QTL for tuber morphology traits in diploid potato. J. Appl. Genet. 2018, 59, 123–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glendinning, D.R. Potato introductions and breeding up to the early 20th century. New Phytol. 1983, 94, 479–505. [Google Scholar] [CrossRef]
- Kloosterman, B.; Oortwijn, M.; Uitdewilligen, J.; America, T.; Vos, R.D.; Visser, R.G.; Bachem, C.W. From QTL to candidate gene: Genetical genomics of simple and complex traits in potato using a pooling strategy. BMC Genom. 2010, 11, 158. [Google Scholar]
- Black, W. Notes on the progenies of various potato hybrids. J. Genet. 1930, 22, 27–43. [Google Scholar] [CrossRef]
- Masson, M.F. Mapping, Combining Abilities, Heritabilities and Heterosis with 4x x 2x crosses in Potato (Haploids, Gametes, Species, Solanum, Tubers); The University of Wisconsin-Madison: Madison, WI, USA, 1985. [Google Scholar]
- D’Hoop, B.R.; Keizer, P.L.; Paulo, M.J.; Visser, R.G.; Eeuwijk, F.A.; Eck, H.J. Identification of agronomically important QTL in tetraploid potato cultivars using a marker–trait association analysis. Theor. Appl. Genet. 2014, 127, 731–748. [Google Scholar] [CrossRef]
- Tai, G.; Misener, G.C. A comparison of tuber shape and tissue composition of potato genotypes. Potato Res. 1994, 37, 353–364. [Google Scholar]
- Chen, N.; Zhu, W.; Xu, J.; Duan, S.; Bian, C.; Hu, J.; Wang, W.; Li, G.; Jin, L. Molecular marker development and primary physical map construction for the tuber shape Ro gene locus in diploid potato (Solanum tuberosum L.). Mol. Breed. 2019, 39, 6. [Google Scholar] [CrossRef]
- Israeli, A.; Reed, J.W.; Ori, N. Genetic dissection of the auxin response network. Nat. Plants 2020, 6, 1082–1090. [Google Scholar] [CrossRef]
- Stone, S.L.; Kwong, L.W.; Yee, K.M.; Pelletier, J.; Lepiniec, L.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. LEAFY COTYLEDON2 encodes a B3 domain transcription factor that induces embryo development. Proc. Natl. Acad. Sci. USA 2001, 98, 11806–11811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieuwland, J. Lipid Transfer Proteins Enhance Cell Wall Extension in Tobacco. Plant Cell 2005, 17, 2009–2019. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, R.; Huaman, Z. Inheritance of morphological and tuber characteristics. Potato Genet. 1994, 267–283. [Google Scholar]
- Tabatabaeefar, A. Size and shape of potato tubers. Int. Agrophysics 2002, 16, 301–305. [Google Scholar]
- Luo, Z.; Hackett, C.; Bradshaw, J.; McNicol, J.; Milbourne, D. Predicting parental genotypes and gene segregation for tetrasomic inheritance. Theor. Appl. Genet. 2000, 100, 1067–1073. [Google Scholar] [CrossRef]
- Bradshaw, J.E.; Hackett, C.A.; Lowe, R.; McLean, K.; Stewart, H.E.; Tierney, I.; Vilaro, M.D.; Bryan, G.J. Detection of a quantitative trait locus for both foliage and tuber resistance to late blight [Phytophthora infestans (Mont.) de Bary] on chromosome 4 of a dihaploid potato clone (Solanum tuberosum subsp. tuberosum). Theor. Appl. Genet. 2006, 113, 943. [Google Scholar] [CrossRef] [PubMed]
- Romanel, E.; Schrago, C.G.; Couñago, R.; Russo, C.; Alves-Ferreira, M. Evolution of the B3 DNA Binding Superfamily: New Insights into REM Family Gene Diversification. PLoS ONE 2009, 4, e5791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardtke, C.S.; Berleth, T. The Arabidopsis gene MONOPTEROS encodes a transcription factor mediating embryo axis formation and vascular development. Embo J. 2014, 17, 1405–1411. [Google Scholar] [CrossRef]
- Li, G.; Hou, M.; Liu, Y.; Pei, Y.; Ye, M.; Zhou, Y.; Huang, C.; Zhao, Y.; Ma, H. Genome-wide identification, characterization and expression analysis of the non-specific lipid transfer proteins in potato. BMC Genom. 2019, 20, 375. [Google Scholar] [CrossRef]
- Horvath, B.M.; Bachem, C.W.; Trindade, L.M.; Oortwijn, M.E.; Visser, R.G. Expression analysis of a family of nsLTP genes tissue specifically expressed throughout the plant and during potato tuber life cycle. Plant Physiol. 2002, 129, 1494–1506. [Google Scholar] [CrossRef] [Green Version]
- Tapia, G.; Morales-Quintana, L.; Parra, C.; Berbel, A.; Alcorta, M. Study of nsLTPs in Lotus japonicus genome reveal a specific epidermal cell member (LjLTP10) regulated by drought stress in aerial organs with a putative role in cutin formation. Plant Mol. Biol. 2013, 82, 485–501. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, B.; Zhang, S.; Yao, J.; Chai, S.; Y, V.; Athar, H.; Rahman, M.U.; Wang, L.; Wang, S. Ectopic Expression of VvFUS3, B3-Domain Transcription Factor, in Tomato Influences Seed Development via Affecting Endoreduplication and Hormones. Hortic. Plant J. 2021. [Google Scholar] [CrossRef]
- Wang, X.; Wu, Z.; Wang, L.; Wu, M.; Zhang, D.; Fang, W.; Chen, F.; Teng, N. Cytological and Molecular Characteristics of Pollen Abortion in Lily with Dysplastic Tapetum. Hortic. Plant J. 2019, 5, 281–294. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Kim, D.; Pertea, G.M.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. Erecipes Res. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Yang, Y.; Guo, X.; Jin, L.; Xiong, X.; Yang, X.; Li, G. Exogenous SA Initiated Defense Response and Multi-signaling Pathway in Tetraploid Potato SD20. Hortic. Plant J. 2020, 6, 99–110. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voorrips, R.; Van, D.; van den Heuvel, L.P.W.J.; Ooijen, J.; Van, J.W. JoinMap® 4.0: Software for the Calculation of Genetic Linkage Maps in Experimental Populations. 2006. Available online: www.biometris.wur.nl (accessed on 20 November 2021).
- Ooijen, J.V.; Ooijen, J.V.; Ooijen, J.; Hoorn, J.; Duin, J.; Van, J.W. MapQTL®6. Software for the Mapping of Quantitative Trait loci in Experimental Populations of Diploid Species. 2009. Available online: www.biometris.wur.nl (accessed on 20 November 2021).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | QTL | LOD | PVE (%) | Additive Effect | Marker Interval | Position |
|---|---|---|---|---|---|---|
| 2014 | 1137CAPS | 42.00 | 72.9% | 0.433438 | 1137CAPS–137CAPSIII | chr10: 49510442–49735039 |
| 2015 | 1137CAPS | 40.29 | 71.4% | 0.364612 | 1137CAPS–1137CAPSIII | chr10: 49510442–49735039 |
| 2016 | 1137CAPS | 39.87 | 71.3% | 0.357161 | 1137CAPS–1137CAPSIII | chr10: 49510442–49735039 |
| 2017 | 1137CAPS | 31.51 | 61.7% | 0.304663 | 1137CAPS–1137CAPSIII | chr10: 49510442–49735039 |
| Name | Description |
|---|---|
| Soltu.DM.10G018490 | Leucine-rich repeat protein kinase family protein |
| Soltu.DM.10G018500 | Leucine-rich repeat protein kinase family protein |
| Soltu.DM.10G018510 | Nuclear shuttle interacting |
| Soltu.DM.10G018520 | Glycosyl hydrolase superfamily protein |
| Soltu.DM.10G018530 | Serine/threonine protein kinase |
| Soltu.DM.10G018540 | Hypothetical protein |
| Soltu.DM.10G018550 | Aminophospholipid ATPase |
| Soltu.DM.10G018560 | WRKY DNA-binding protein |
| Soltu.DM.10G018570 | Protein of unknown function, DUF599 |
| Soltu.DM.10G018580 Soltu.DM.10G018590 | Lung seven transmembrane receptor family protein Vacuolar sorting receptor homolog |
| Soltu.DM.10G018600 | Polynucleotidyl transferase, ribonuclease H-like superfamily protein |
| Soltu.DM.10G018610 | Elongation factor P (EF-P) family protein |
| Soltu.DM.10G018620 | HSI2-like |
| Soltu.DM.10G018630 Soltu.DM.10G018640 | Hypothetical protein HSI2-like |
| Soltu.DM.10G018650 | Protein kinase superfamily protein |
| Soltu.DM.10G018660 | Non-specific lipid transfer protein |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, G.; Wang, Q.; Xu, J.; Chen, N.; Zhu, W.; Duan, S.; Yang, X.; De Jong, W.S.; Guo, Y.; Jin, L.; et al. Fine Mapping and Candidate Gene Prediction of Tuber Shape Controlling Ro Locus Based on Integrating Genetic and Transcriptomic Analyses in Potato. Int. J. Mol. Sci. 2022, 23, 1470. https://doi.org/10.3390/ijms23031470
Fan G, Wang Q, Xu J, Chen N, Zhu W, Duan S, Yang X, De Jong WS, Guo Y, Jin L, et al. Fine Mapping and Candidate Gene Prediction of Tuber Shape Controlling Ro Locus Based on Integrating Genetic and Transcriptomic Analyses in Potato. International Journal of Molecular Sciences. 2022; 23(3):1470. https://doi.org/10.3390/ijms23031470
Chicago/Turabian StyleFan, Guiyan, Qianru Wang, Jianfei Xu, Na Chen, Wenwen Zhu, Shaoguang Duan, Xiaohui Yang, Walter S. De Jong, Yangdong Guo, Liping Jin, and et al. 2022. "Fine Mapping and Candidate Gene Prediction of Tuber Shape Controlling Ro Locus Based on Integrating Genetic and Transcriptomic Analyses in Potato" International Journal of Molecular Sciences 23, no. 3: 1470. https://doi.org/10.3390/ijms23031470
APA StyleFan, G., Wang, Q., Xu, J., Chen, N., Zhu, W., Duan, S., Yang, X., De Jong, W. S., Guo, Y., Jin, L., & Li, G. (2022). Fine Mapping and Candidate Gene Prediction of Tuber Shape Controlling Ro Locus Based on Integrating Genetic and Transcriptomic Analyses in Potato. International Journal of Molecular Sciences, 23(3), 1470. https://doi.org/10.3390/ijms23031470