
A Fungal Versatile GH10 Endoxylanase and Its Glycosynthase Variant: Synthesis of Xylooligosaccharides and Glycosides of Bioactive Phenolic Compounds

 , , , , , , and
, , , , , , and
Abstract
1. Introduction
2. Results and Discussion
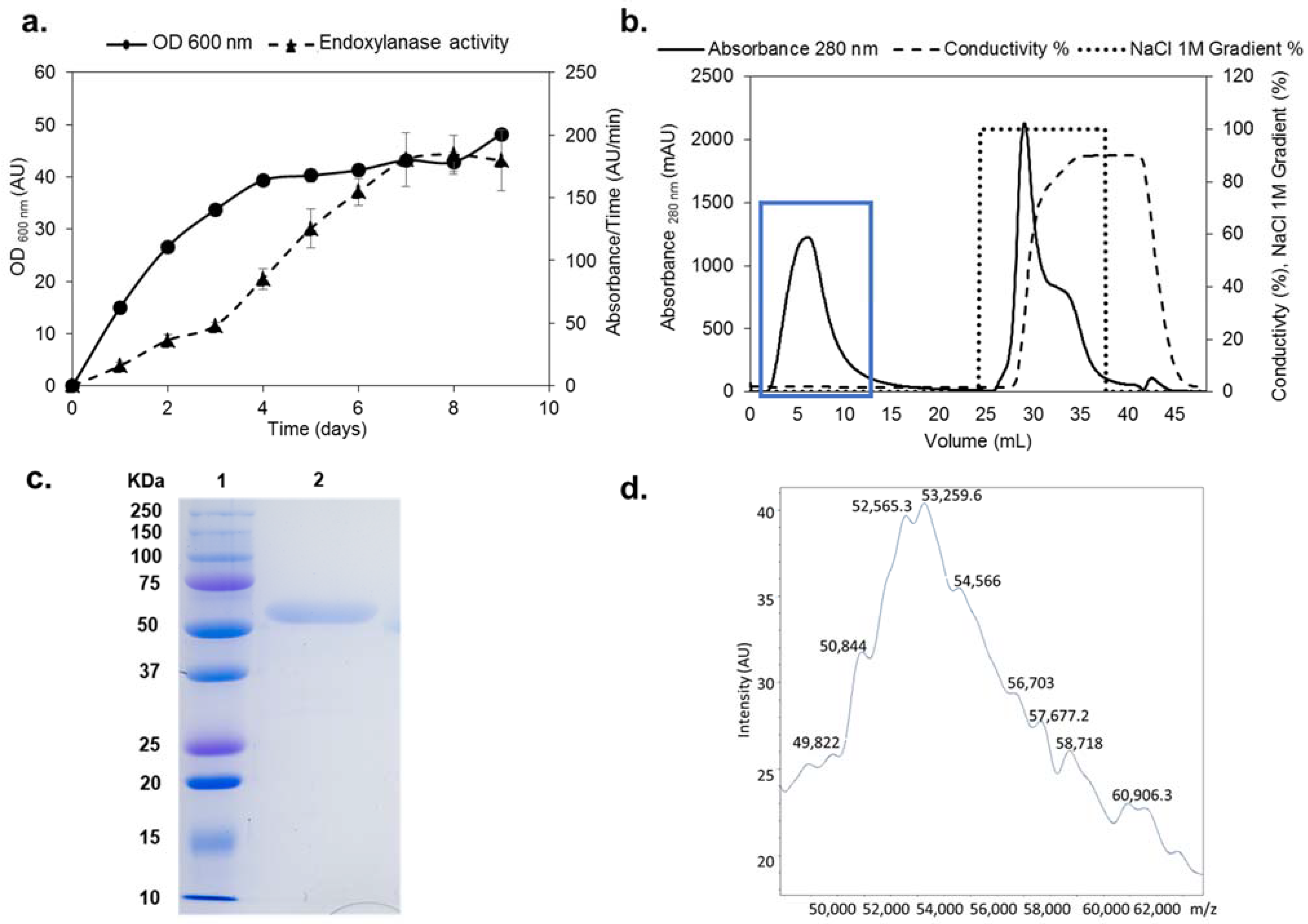
2.1. Cloning, Production, and Purification of rXynSOS
2.2. Physicochemical Properties of rXynSOS
2.3. Substrate Specificity and Kinetics of rXynSOS
2.4. Transglycosylation Potential of rXynSOS
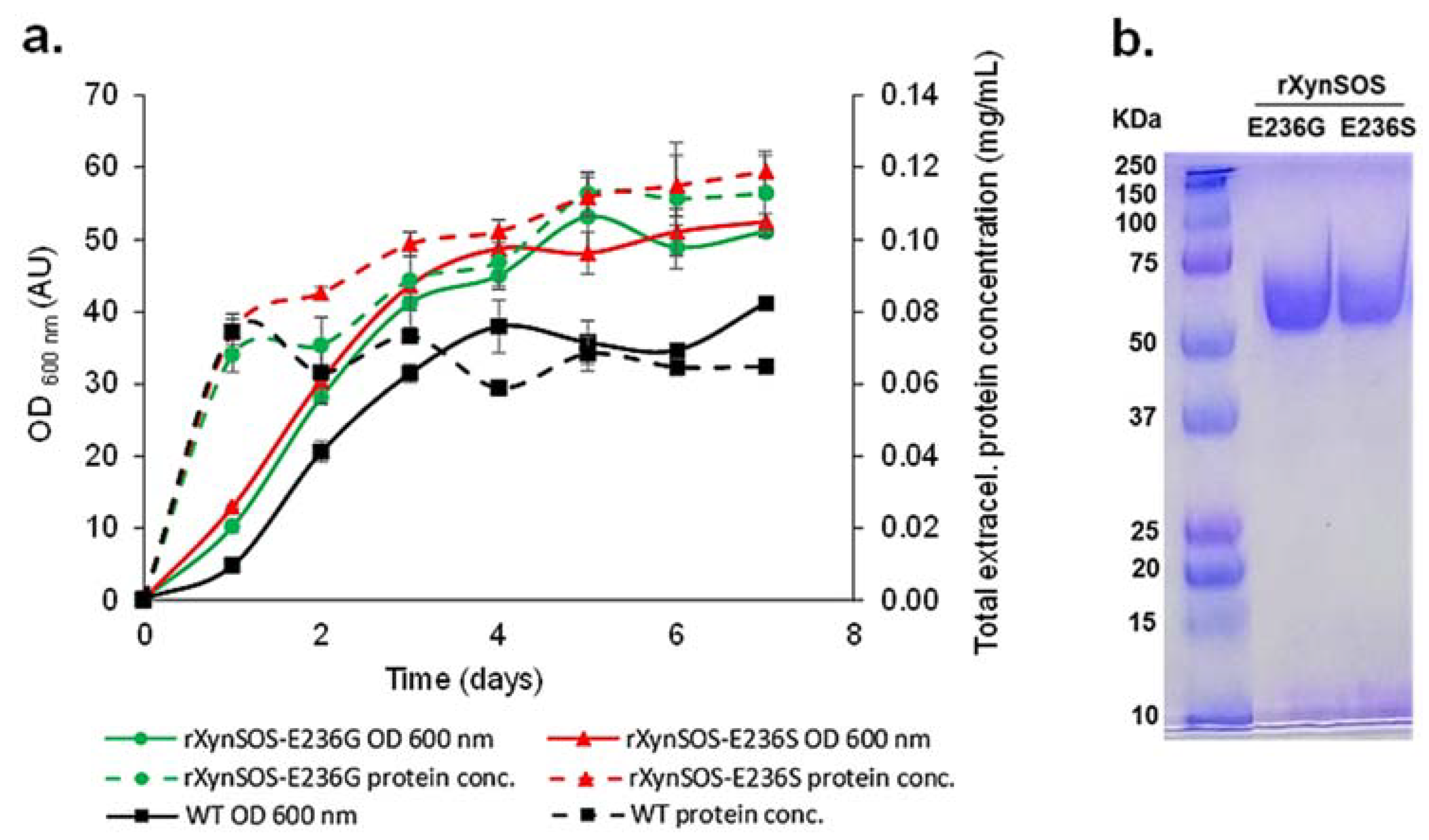
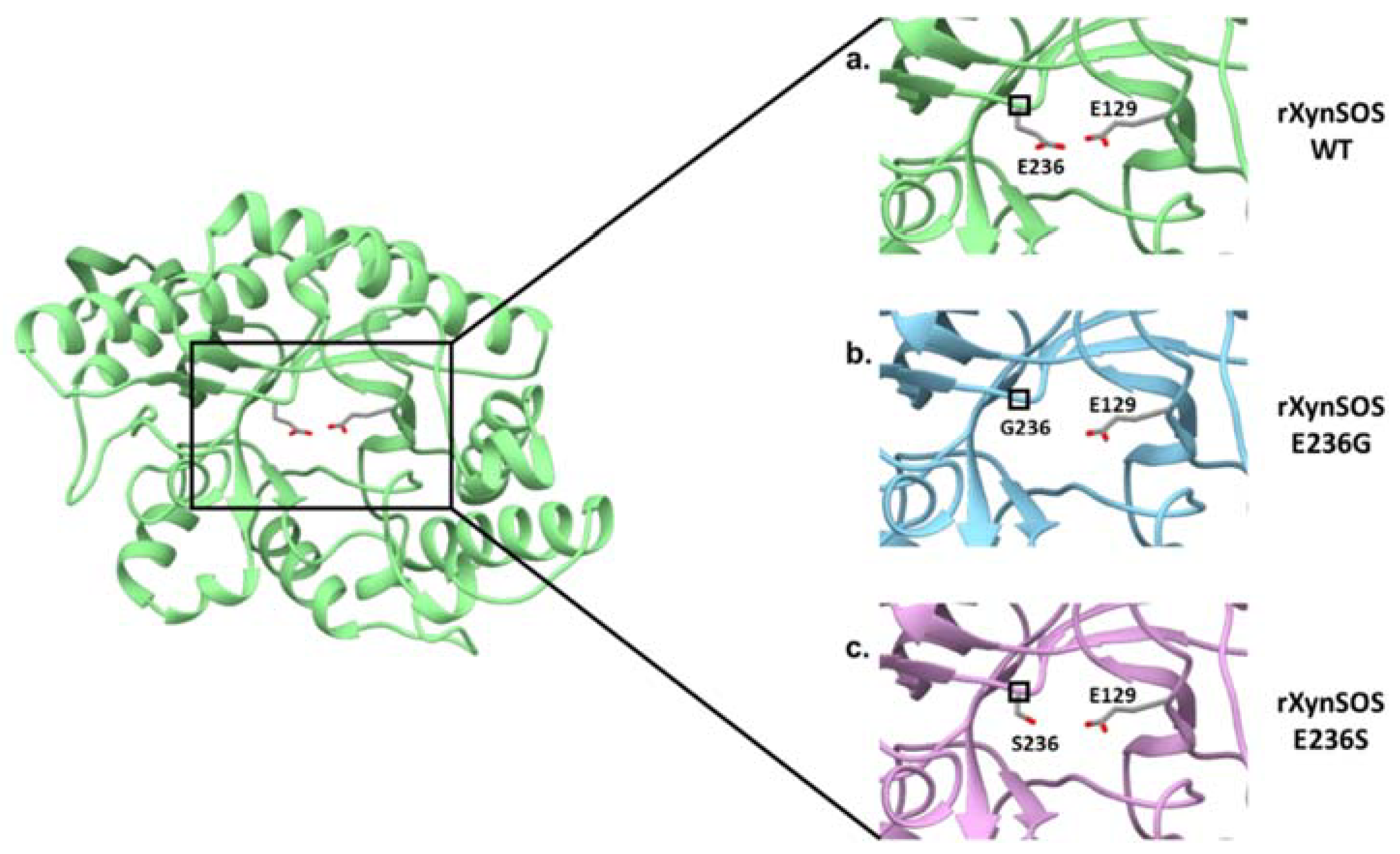
2.5. Conversion of rXynSOS into Its Glycosynthase Variants
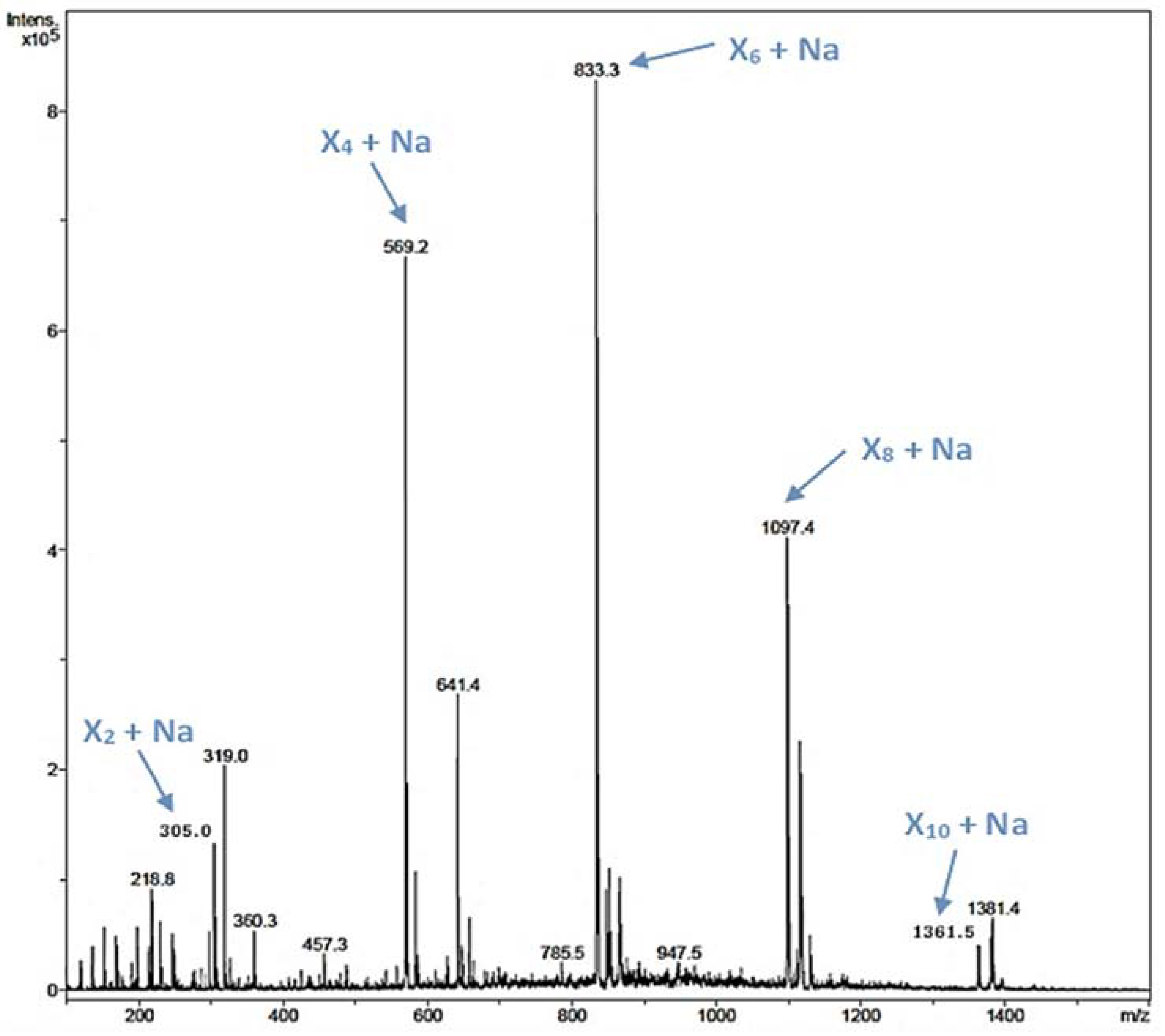
2.6. Oligosaccharides Synthesis by the Glycosynthase Variant rXynSOS-E236G
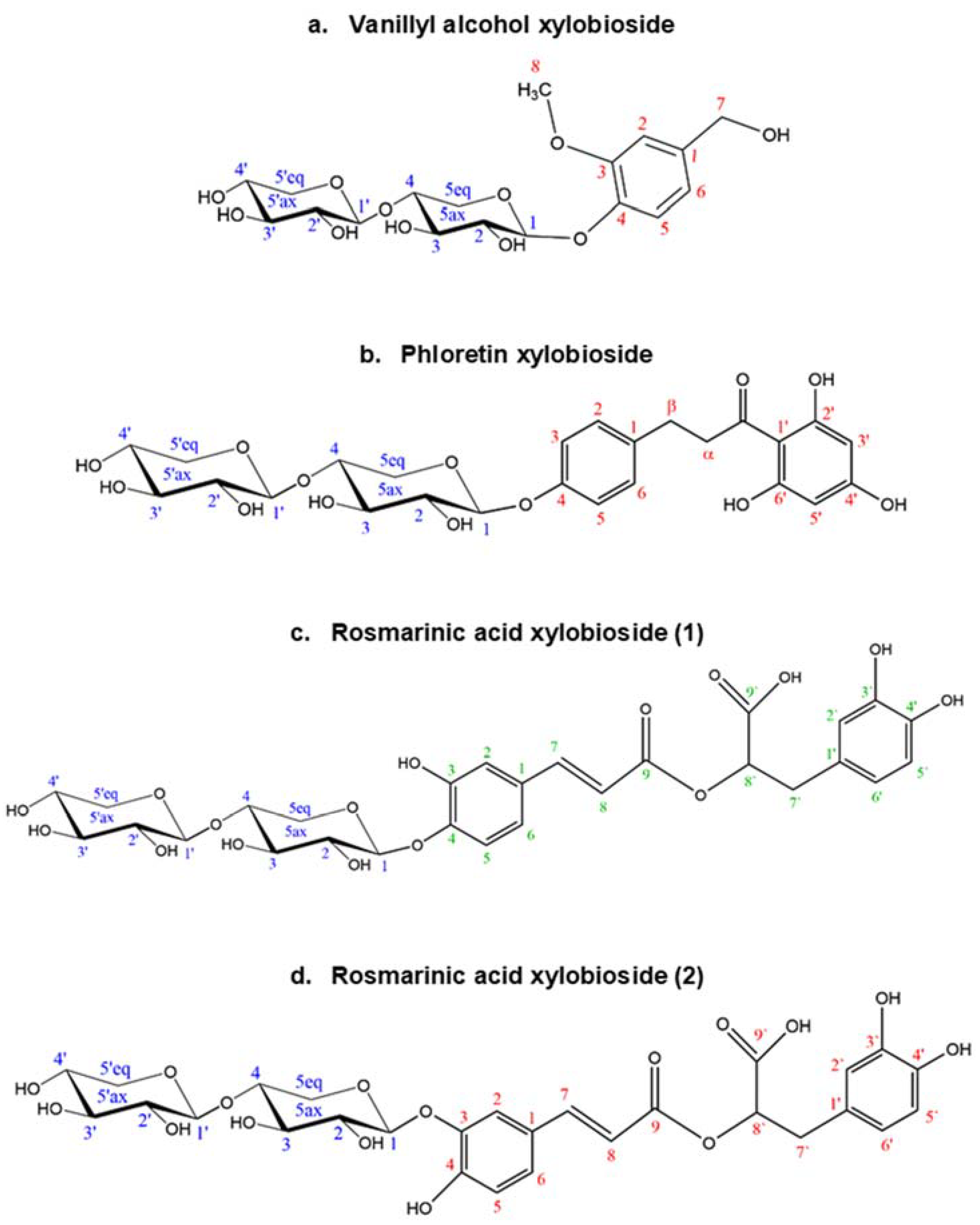
2.7. Glycosylation Profile of the Glycosynthase Variant rXynSOS-E236G
3. Materials and Methods
3.1. Strains and Media
3.2. Nucleic Acid Isolation and Cloning of rXynSOS
3.3. Conversion of rXynSOS into Its Glycosynthase Variants
3.4. Production and Purification of rXynSOS and Its Glycosynthase Variants
3.5. Determination of the Physicochemical Properties of rXynSOS
3.6. Substrate Specificity and Kinetic Assays of rXynSOS
3.7. Transglycosylation Assays
3.8. Purification and NMR Analysis of Glycosylated Products
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liguori, R.; Faraco, V. Biological processes for advancing lignocellulosic waste biorefinery by advocating circular economy. Bioresour. Technol. 2016, 215, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.J.; Wang, H.-T.; York, W.S.; Peña, M.J.; Urbanowicz, B.R. Designer biomass for next-generation biorefineries: Leveraging recent insights into xylan structure and biosynthesis. Biotechnol. Biofuels 2017, 10, 286. [Google Scholar] [CrossRef] [PubMed]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef] [PubMed]
- Méndez-Líter, J.A.; de Eugenio, L.I.; Nieto-Domínguez, M.; Prieto, A.; Martínez, M.J. Hemicellulases from Penicillium and Talaromyces for lignocellulosic biomass valorization: A review. Bioresour. Technol. 2021, 324, 124623. [Google Scholar] [CrossRef] [PubMed]
- Polizeli, M.L.T.M.; Rizzatti, A.C.S.; Monti, R.; Terenzi, H.F.; Jorge, J.A.; Amorim, D.S. Xylanases from fungi: Properties and industrial applications. Appl. Microbiol. Biotechnol. 2005, 67, 577–591. [Google Scholar] [CrossRef]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef]
- Linares-Pasten, J.A.; Aronsson, A.; Karlsson, E.N. Structural Considerations on the Use of Endo-Xylanases for the Production of prebiotic Xylooligosaccharides from Biomass. Curr. Protein Pept. Sci. 2017, 19, 48–67. [Google Scholar] [CrossRef] [PubMed]
- Collins, T.; Gerday, C.; Feller, G. Xylanases, xylanase families and extremophilic xylanases. FEMS Microbiol. Rev. 2005, 29, 3–23. [Google Scholar] [CrossRef]
- Paës, G.; Berrin, J.-G.; Beaugrand, J. GH11 xylanases: Structure/function/properties relationships and applications. Biotechnol. Adv. 2012, 30, 564–592. [Google Scholar] [CrossRef] [PubMed]
- Biely, P.; Vršanská, M.; Tenkanen, M.; Kluepfel, D. Endo-β-1,4-xylanase families: Differences in catalytic properties. J. Biotechnol. 1997, 57, 151–166. [Google Scholar] [CrossRef]
- Bhardwaj, N.; Kumar, B.; Verma, P. A detailed overview of xylanases: An emerging biomolecule for current and future prospective. Bioresour. Bioprocess. 2019, 6, 40. [Google Scholar] [CrossRef]
- Nieto-Domínguez, M.; de Eugenio, L.I.; York-Durán, M.J.; Rodríguez-Colinas, B.; Plou, F.J.; Chenoll, E.; Pardo, E.; Codoñer, F.; Jesús Martínez, M. Prebiotic effect of xylooligosaccharides produced from birchwood xylan by a novel fungal GH11 xylanase. Food Chem. 2017, 232, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Walia, A.; Guleria, S.; Mehta, P.; Chauhan, A.; Parkash, J. Microbial xylanases and their industrial application in pulp and paper biobleaching: A review. 3 Biotech 2017, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- Gangwar, A.K.; Prakash, N.T.; Prakash, R. Applicability of Microbial Xylanases in Paper Pulp Bleaching: A Review. BioResources 2014, 9, 3733–3754. [Google Scholar] [CrossRef]
- Juturu, V.; Wu, J.C. Microbial xylanases: Engineering, production and industrial applications. Biotechnol. Adv. 2012, 30, 1219–1227. [Google Scholar] [CrossRef] [PubMed]
- Danby, P.M.; Withers, S.G. Advances in Enzymatic Glycoside Synthesis. ACS Chem. Biol. 2016, 11, 1784–1794. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Qi, T.; Xu, L.; Lu, L.; Xiao, M. Recent progress in the enzymatic glycosylation of phenolic compounds. J. Carbohydr. Chem. 2016, 35, 1–23. [Google Scholar] [CrossRef]
- Perugino, G. Oligosaccharide synthesis by glycosynthases. Trends Biotechnol. 2004, 22, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Hayes, M.; Pietruszka, J. Synthesis of Glycosides by Glycosynthases. Molecules 2017, 22, 1434. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Edmunds, G.; Gibbons, C.; Zhang, J.; Gadi, M.R.; Zhu, H.; Fang, J.; Liu, X.; Kong, Y.; Wang, P.G. Toward Automated Enzymatic Synthesis of Oligosaccharides. Chem. Rev. 2018, 118, 8151–8187. [Google Scholar] [CrossRef] [PubMed]
- Jahn, M.; Withers, S.G. New Approaches to Enzymatic Oligosaccharide Synthesis: Glycosynthases and Thioglycoligases. Biocatal. Biotransform. 2003, 21, 159–166. [Google Scholar] [CrossRef]
- Madhavan, A.; Arun, K.; Sindhu, R.; Alphonsa Jose, A.; Pugazhendhi, A.; Binod, P.; Sirohi, R.; Reshmy, R.; Kumar Awasthi, M. Engineering interventions in industrial filamentous fungal cell factories for biomass valorization. Bioresour. Technol. 2022, 344, 126209. [Google Scholar] [CrossRef]
- Amore, A.; Giacobbe, S.; Faraco, V. Regulation of Cellulase and Hemicellulase Gene Expression in Fungi. Curr. Genom. 2013, 14, 230–249. [Google Scholar] [CrossRef]
- Yilmaz, N.; Houbraken, J.; Hoekstra, E.S.; Frisvad, J.C.; Visagie, C.M.; Samson, R.A. Delimitation and characterisation of Talaromyces purpurogenus and related species. Persoonia Mol. Phylogeny Evol. Fungi 2012, 29, 39–54. [Google Scholar] [CrossRef] [PubMed]
- De Eugenio, L.I.; Méndez-Líter, J.A.; Nieto-Domínguez, M.; Alonso, L.; Gil-Muñoz, J.; Barriuso, J.; Prieto, A.; Martínez, M.J. Differential β-glucosidase expression as a function of carbon source availability in Talaromyces amestolkiae: A genomic and proteomic approach. Biotechnol. Biofuels 2017, 10, 161. [Google Scholar] [CrossRef] [PubMed]
- Prieto, A.; de Eugenio, L.; Méndez-Líter, J.A.; Nieto-Domínguez, M.; Murgiondo, C.; Barriuso, J.; Bejarano-Muñoz, L.; Martínez, M.J. Fungal glycosyl hydrolases for sustainable plant biomass valorization: Talaromyces amestolkiae as a model fungus. Int. Microbiol. 2021, 24, 545–558. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Matsuda, M.; Maeda, K.; Mikata, K. The Phylogenetic Relationships of Methanol-assimilating Yeasts Based on the Partial Sequences of 18S and 26S Ribosomal RNAs: The Proposal of Komagataella Gen. Nov. (Saccharomycetaceae). Biosci. Biotechnol. Biochem. 2014, 59, 439–444. [Google Scholar] [CrossRef]
- Karbalaei, M.; Rezaee, S.A.; Farsiani, H. Pichia pastoris: A highly successful expression system for optimal synthesis of heterologous proteins. J. Cell. Physiol. 2020, 235, 5867–5881. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Domínguez, M.; Martínez-Fernández, J.A.; de Toro, B.F.; Méndez-Líter, J.A.; Cañada, F.J.; Prieto, A.; de Eugenio, L.I.; Martínez, M.J. Exploiting xylan as sugar donor for the synthesis of an antiproliferative xyloside using an enzyme cascade. Microb. Cell Fact. 2019, 18, 174. [Google Scholar] [CrossRef]
- Miao, Y.; Li, P.; Li, G.; Liu, D.; Druzhinina, I.S.; Kubicek, C.P.; Shen, Q.; Zhang, R. Two degradation strategies for overcoming the recalcitrance of natural lignocellulosic xylan by polysaccharides-binding GH10 and GH11 xylanases of filamentous fungi. Environ. Microbiol. 2017, 19, 1054–1064. [Google Scholar] [CrossRef] [PubMed]
- Cereghino, J.L.; Cregg, J.M. Heterologous protein expression in the methylotrophic yeast Pichia pastoris. FEMS Microbiol. Rev. 2000, 24, 45–66. [Google Scholar] [CrossRef] [PubMed]
- Velasco, J.; Oliva, B.; Mulinari, E.J.; Quintero, L.P.; da Silva Lima, A.; Gonçalves, A.L.; Gonçalves, T.A.; Damasio, A.; Squina, F.M.; Ferreira Milagres, A.M.; et al. Heterologous expression and functional characterization of a GH10 endoxylanase from Aspergillus fumigatus var. niveus with potential biotechnological application. Biotechnol. Rep. 2019, 24, e00382. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, L.F.; De Lucas, R.C.; Vitcosque, G.L.; Ribeiro, L.F.; Ward, R.J.; Rubio, M.V.; Damásio, A.R.; Squina, F.M.; Gregory, R.C.; Walton, P.H.; et al. A novel thermostable xylanase GH10 from Malbranchea pulchella expressed in Aspergillus nidulans with potential applications in biotechnology. Biotechnol. Biofuels 2014, 7, 115. [Google Scholar] [CrossRef]
- Lafond, M.; Tauzin, A.; Desseaux, V.; Bonnin, E.; Ajandouz, E.-H.; Giardina, T. GH10 xylanase D from Penicillium funiculosum: Biochemical studies and xylooligosaccharide production. Microb. Cell Fact. 2011, 10, 20. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-L.; Zhang, M.; Zhang, D.-H.; Chen, X.-L.; Sun, C.-Y.; Zhou, B.-C.; Zhang, Y.-Z. Purification and enzymatic characterization of two β-endoxylanases from Trichoderma sp. K9301 and their actions in xylooligosaccharide production. Bioresour. Technol. 2009, 100, 5230–5236. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.; Zheng, H.; Li, S.; Wei, Z.; Mei, X.; Ma, H.; Shen, Q.; Xu, Y. Functional diversity and properties of multiple xylanases from Penicillium oxalicum GZ-2. Sci. Rep. 2015, 5, 12631. [Google Scholar] [CrossRef] [PubMed]
- Shibata, N.; Suetsugu, M.; Kakeshita, H.; Igarashi, K.; Hagihara, H.; Takimura, Y. A novel GH10 xylanase from Penicillium sp. accelerates saccharification of alkaline-pretreated bagasse by an enzyme from recombinant Trichoderma reesei expressing Aspergillus β-glucosidase. Biotechnol. Biofuels 2017, 10, 278. [Google Scholar] [CrossRef] [PubMed]
- Inoue, H.; Kishishita, S.; Kumagai, A.; Kataoka, M.; Fujii, T.; Ishikawa, K. Contribution of a family 1 carbohydrate-binding module in thermostable glycoside hydrolase 10 xylanase from Talaromyces cellulolyticus toward synergistic enzymatic hydrolysis of lignocellulose. Biotechnol. Biofuels 2015, 8, 77. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Mumper, R.J. Plant Phenolics: Extraction, Analysis and Their Antioxidant and Anticancer Properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, B.R.; Heleno, S.A.; Oliveira, M.B.P.P.; Barros, L.; Ferreira, I.C.F.R. Phenolic compounds: Current industrial applications, limitations and future challenges. Food Funct. 2021, 12, 14–29. [Google Scholar] [CrossRef] [PubMed]
- Abdul Manas, N.H.; Md. Illias, R.; Mahadi, N.M. Strategy in manipulating transglycosylation activity of glycosyl hydrolase for oligosaccharide production. Crit. Rev. Biotechnol. 2018, 38, 272–293. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-W.; Fox, D.T.; Hekmat, O.; Kantner, T.; McIntosh, L.P.; Warren, R.A.J.; Withers, S.G. Glycosynthase-based synthesis of xylo-oligosaccharides using an engineered retaining xylanase from Cellulomonas fimi. Org. Biomol. Chem. 2006, 4, 2025–2032. [Google Scholar] [CrossRef] [PubMed]
- Sugimura, M.; Nishimoto, M.; Kitaoka, M. Characterization of Glycosynthase Mutants Derived from Glycoside Hydrolase Family 10 Xylanases. Biosci. Biotechnol. Biochem. 2006, 70, 1210–1217. [Google Scholar] [CrossRef] [PubMed]
- Aachary, A.A.; Prapulla, S.G. Xylooligosaccharides (XOS) as an Emerging Prebiotic: Microbial Synthesis, Utilization, Structural Characterization, Bioactive Properties, and Applications. Compr. Rev. Food Sci. Food Saf. 2011, 10, 2–16. [Google Scholar] [CrossRef]
- Chapla, D.; Pandit, P.; Shah, A. Production of xylooligosaccharides from corncob xylan by fungal xylanase and their utilization by probiotics. Bioresour. Technol. 2012, 115, 215–221. [Google Scholar] [CrossRef]
- Da Silva Menezes, B.; Rossi, D.M.; Squina, F.; Ayub, M.A.Z. Comparative production of xylanase and the liberation of xylooligosaccharides from lignocellulosic biomass by Aspergillus brasiliensis BLf1 and recombinant Aspergillus nidulans XynC A773. Int. J. Food Sci. Technol. 2018, 53, 2110–2118. [Google Scholar] [CrossRef]
- Driss, D.; Zouari-Ellouzi, S.; Chaari, F.; Kallel, F.; Ghazala, I.; Bouaziz, F.; Ghorbel, R.; Chaabouni, S.E. Production and in vitro evaluation of xylooligosaccharides generated from corncobs using immobilized Penicillium occitanis xylanase. J. Mol. Catal. B Enzym. 2014, 102, 146–153. [Google Scholar] [CrossRef]
- Wu, Q.; Fan, G.; Yu, T.; Sun, B.; Tang, H.; Teng, C.; Yang, R.; Li, X. Biochemical characteristics of the mutant xylanase T-XynC(122)C(166) and production of xylooligosaccharides from corncobs. Ind. Crops Prod. 2019, 142, 111848. [Google Scholar] [CrossRef]
- González-Alfonso, J.; Rodrigo-Frutos, D.; Belmonte-Reche, E.; Peñalver, P.; Poveda, A.; Jiménez-Barbero, J.; Ballesteros, A.; Hirose, Y.; Polaina, J.; Morales, J.; et al. Enzymatic Synthesis of a Novel Pterostilbene α-Glucoside by the Combination of Cyclodextrin Glucanotransferase and Amyloglucosidase. Molecules 2018, 23, 1271. [Google Scholar] [CrossRef]
- Wang, Y.; Li, D.; Lin, H.; Jiang, S.; Han, L.; Hou, S.; Lin, S.; Cheng, Z.; Bian, W.; Zhang, X.; et al. Enhanced oral bioavailability and bioefficacy of phloretin using mixed polymeric modified self-nanoemulsions. Food Sci. Nutr. 2020, 8, 3545–3558. [Google Scholar] [CrossRef]
- Bethune, S.J.; Schultheiss, N.; Henck, J.-O. Improving the Poor Aqueous Solubility of Nutraceutical Compound Pterostilbene through Cocrystal Formation. Cryst. Growth Des. 2011, 11, 2817–2823. [Google Scholar] [CrossRef]
- Nieto-Domínguez, M.; Prieto, A.; Fernández de Toro, B.; Cañada, F.J.; Barriuso, J.; Armstrong, Z.; Withers, S.G.; de Eugenio, L.I.; Martínez, M.J. Enzymatic fine-tuning for 2-(6-hydroxynaphthyl) β-d-xylopyranoside synthesis catalyzed by the recombinant β-xylosidase BxTW1 from Talaromyces amestolkiae. Microb. Cell Fact. 2016, 15, 171. [Google Scholar] [CrossRef] [PubMed]
- Méndez-Líter, J.A.; Tundidor, I.; Nieto-Domínguez, M.; de Toro, B.F.; González Santana, A.; de Eugenio, L.I.; Prieto, A.; Asensio, J.L.; Cañada, F.J.; Sánchez, C.; et al. Transglycosylation products generated by Talaromyces amestolkiae GH3 β-glucosidases: Effect of hydroxytyrosol, vanillin and its glucosides on breast cancer cells. Microb. Cell Fact. 2019, 18, 97. [Google Scholar] [CrossRef] [PubMed]
- Cosme, P.; Rodríguez, A.B.; Espino, J.; Garrido, M. Plant Phenolics: Bioavailability as a Key Determinant of Their Potential Health-Promoting Applications. Antioxidants 2020, 9, 1263. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Alfonso, J.L.; Ubiparip, Z.; Jimenez-Ortega, E.; Poveda, A.; Alonso, C.; Coderch, L.; Jimenez-Barbero, J.; Sanz-Aparicio, J.; Ballesteros, A.O.; Desmet, T.; et al. Enzymatic Synthesis of Phloretin α-Glucosides Using a Sucrose Phosphorylase Mutant and its Effect on Solubility, Antioxidant Properties and Skin Absorption. Adv. Synth. Catal. 2021, 363, 3079–3089. [Google Scholar] [CrossRef]
- De Winter, K.; Dewitte, G.; Dirks-Hofmeister, M.E.; De Laet, S.; Pelantová, H.; Křen, V.; Desmet, T. Enzymatic Glycosylation of Phenolic Antioxidants: Phosphorylase-Mediated Synthesis and Characterization. J. Agric. Food Chem. 2015, 63, 10131–10139. [Google Scholar] [CrossRef]
- Nieto-Domínguez, M.; de Eugenio, L.I.; Peñalver, P.; Belmonte-Reche, E.; Morales, J.C.; Poveda, A.; Jiménez-Barbero, J.; Prieto, A.; Plou, F.J.; Martínez, M.J. Enzymatic Synthesis of a Novel Neuroprotective Hydroxytyrosyl Glycoside. J. Agric. Food Chem. 2017, 65, 10526–10533. [Google Scholar] [CrossRef]
- Nieto-Domínguez, M.; Fernández de Toro, B.; de Eugenio, L.I.; Santana, A.G.; Bejarano-Muñoz, L.; Armstrong, Z.; Méndez-Líter, J.A.; Asensio, J.L.; Prieto, A.; Withers, S.G.; et al. Thioglycoligase derived from fungal GH3 β-xylosidase is a multi-glycoligase with broad acceptor tolerance. Nat. Commun. 2020, 11, 4864. [Google Scholar] [CrossRef]
- Oode, C.; Shimada, W.; Yokota, M.; Yamada, Y.; Nihei, K. Dihydroresveratrol cellobioside and xylobioside as effective melanogenesis activators. Carbohydr. Res. 2016, 436, 45–49. [Google Scholar] [CrossRef]
- Méndez-Líter, J.A.; Nieto-Domínguez, M.; Fernández de Toro, B.; González Santana, A.; Prieto, A.; Asensio, J.L.; Cañada, F.J.; de Eugenio, L.I.; Martínez, M.J. A glucotolerant β-glucosidase from the fungus Talaromyces amestolkiae and its conversion into a glycosynthase for glycosylation of phenolic compounds. Microb. Cell Fact. 2020, 19, 127. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.; Gübitz, G.M.; Kratky, C. Xylan Binding Subsite Mapping in the Xylanase from Penicillium simplicissimum Using Xylooligosaccharides as Cryo-Protectant. Biochemistry 1999, 38, 2403–2412. [Google Scholar] [CrossRef] [PubMed]
- Payan, F.; Leone, P.; Porciero, S.; Furniss, C.; Tahir, T.; Williamson, G.; Durand, A.; Manzanares, P.; Gilbert, H.J.; Juge, N.; et al. The Dual Nature of the Wheat Xylanase Protein Inhibitor XIP-I. J. Biol. Chem. 2004, 279, 36029–36037. [Google Scholar] [CrossRef]
- Lo Leggio, L.; Kalogiannis, S.; Eckert, K.; Teixeira, S.C.; Bhat, M.K.; Andrei, C.; Pickersgill, R.W.; Larsen, S. Substrate specificity and subsite mobility in T. aurantiacus xylanase 10A. FEBS Lett. 2001, 509, 303–308. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef]
- Gupta, R.; Brunak, S. Prediction of glycosylation across the human proteome and the correlation to protein function. Pac. Symp. Biocomput. 2002, 7, 310–322. [Google Scholar] [CrossRef]
- Steentoft, C.; Vakhrushev, S.Y.; Joshi, H.J.; Kong, Y.; Vester-Christensen, M.B.; Schjoldager, K.T.-B.G.; Lavrsen, K.; Dabelsteen, S.; Pedersen, N.B.; Marcos-Silva, L.; et al. Precision mapping of the human O-GalNAc glycoproteome through SimpleCell technology. EMBO J. 2013, 32, 1478–1488. [Google Scholar] [CrossRef]
- Nieto-Domínguez, M.; de Eugenio, L.I.; Barriuso, J.; Prieto, A.; Fernández de Toro, B.; Canales-Mayordomo, Á.; Martínez, M.J. Novel pH-Stable Glycoside Hydrolase Family 3 β-Xylosidase from Talaromyces amestolkiae: An Enzyme Displaying Regioselective Transxylosylation. Appl. Environ. Microbiol. 2015, 81, 6380–6392. [Google Scholar] [CrossRef]
- Zhang, H.; Yohe, T.; Huang, L.; Entwistle, S.; Wu, P.; Yang, Z.; Busk, P.K.; Xu, Y.; Yin, Y. dbCAN2: A meta server for automated carbohydrate-active enzyme annotation. Nucleic Acids Res. 2018, 46, W95–W101. [Google Scholar] [CrossRef] [PubMed]
- Somogyi, M. A new reagent for the determination of sugars. J. Biol. Chem. 1945, 160, 61–68. [Google Scholar] [CrossRef]
- Gonzalez-Alfonso, J.L.; Peñalver, P.; Ballesteros, A.O.; Morales, J.C.; Plou, F.J. Effect of α-Glucosylation on the Stability, Antioxidant Properties, Toxicity, and Neuroprotective Activity of (−)-Epigallocatechin Gallate. Front. Nutr. 2019, 6, 30. [Google Scholar] [CrossRef] [PubMed]
- Faijes, M.; Saura-Valls, M.; Pérez, X.; Conti, M.; Planas, A. Acceptor-dependent regioselectivity of glycosynthase reactions by Streptomyces E383A β-glucosidase. Carbohydr. Res. 2006, 341, 2055–2065. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, H.E.; Kotlyar, V.; Nudelman, A. NMR Chemical Shifts of Common Laboratory Solvents as Trace Impurities. J. Org. Chem. 1997, 62, 7512–7515. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Specific Activity (U mg−1) |
|---|---|
| Beechwood xylan (0.8%) | 132.33 ± 11.6 |
| Wheat arabinoxylan (0.8%) | 149.15 ± 3.45 |
| CMC (0.8%) | 1.25 ± 0.35 |
| Microcrystalline cellulose (0.8%) | Not active |
| Cellobiose (0.1%) | Not active |
| pNPX2 (0.1%) | 56.97 ± 3.96 |
| pNPX (0.1%) | 1.81 ± 0.06 |
| pNPG2 (0.1%) | 0.82 ± 0.01 |
| pNPG (0.1%) | 0.03 ± 0.001 |
| Substrate | Km (g L−1) | kcat (s−1) | kcat/Km (s−1 g−1 L) |
|---|---|---|---|
| Beechwood xylan | 1.07 | 144.25 | 134.69 |
| Wheat arabinoxylan | 1.94 | 163.93 | 84.65 |
| CMC | 51.29 | 7.90 | 0.15 |
| pNPX2 | 0.05 g L−1 0.12 mM | 102.96 | 2181.45 s−1 g−1 L 880.04 mM−1 |
| Glycoside | Mass ESI-MS (m/z) 1 | Yield HPLC (%) |
|---|---|---|
| Vanillyl alcohol-X2 | 441.1 | 4.3 |
| 2-hydroxybenzyl alcohol-X2 | 411.0 | 7.6 |
| Hydroquinone-X2 | 397.0 | 26.0 |
| Gallic acid-X2 | 457.1 | 21.7 |
| Epigallocatechin gallate (EGCG)-X2 | 745.2 | 1.8 |
| Rosmarinic acid-X2 (two main glycosides detected) | 647.2 | 20.9 (1) 20.0 (2) |
| Phloretin-X2 | 561.1 | 7.8 |
| Pterostilbene-X2 | 543.3 | 1.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pozo-Rodríguez, A.; Méndez-Líter, J.A.; de Eugenio, L.I.; Nieto-Domínguez, M.; Calviño, E.; Cañada, F.J.; Santana, A.G.; Díez, J.; Asensio, J.L.; Barriuso, J.; et al. A Fungal Versatile GH10 Endoxylanase and Its Glycosynthase Variant: Synthesis of Xylooligosaccharides and Glycosides of Bioactive Phenolic Compounds. Int. J. Mol. Sci. 2022, 23, 1383. https://doi.org/10.3390/ijms23031383
Pozo-Rodríguez A, Méndez-Líter JA, de Eugenio LI, Nieto-Domínguez M, Calviño E, Cañada FJ, Santana AG, Díez J, Asensio JL, Barriuso J, et al. A Fungal Versatile GH10 Endoxylanase and Its Glycosynthase Variant: Synthesis of Xylooligosaccharides and Glycosides of Bioactive Phenolic Compounds. International Journal of Molecular Sciences. 2022; 23(3):1383. https://doi.org/10.3390/ijms23031383
Chicago/Turabian StylePozo-Rodríguez, Ana, Juan A. Méndez-Líter, Laura I. de Eugenio, Manuel Nieto-Domínguez, Eva Calviño, Francisco Javier Cañada, Andrés G. Santana, Jaime Díez, Juan L. Asensio, Jorge Barriuso, and et al. 2022. "A Fungal Versatile GH10 Endoxylanase and Its Glycosynthase Variant: Synthesis of Xylooligosaccharides and Glycosides of Bioactive Phenolic Compounds" International Journal of Molecular Sciences 23, no. 3: 1383. https://doi.org/10.3390/ijms23031383
APA StylePozo-Rodríguez, A., Méndez-Líter, J. A., de Eugenio, L. I., Nieto-Domínguez, M., Calviño, E., Cañada, F. J., Santana, A. G., Díez, J., Asensio, J. L., Barriuso, J., Prieto, A., & Martínez, M. J. (2022). A Fungal Versatile GH10 Endoxylanase and Its Glycosynthase Variant: Synthesis of Xylooligosaccharides and Glycosides of Bioactive Phenolic Compounds. International Journal of Molecular Sciences, 23(3), 1383. https://doi.org/10.3390/ijms23031383