
MPP+-Induced Changes in Cellular Impedance as a Measure for Organic Cation Transporter (SLC22A1-3) Activity and Inhibition

 , , and
, , and
Abstract
1. Introduction
2. Results
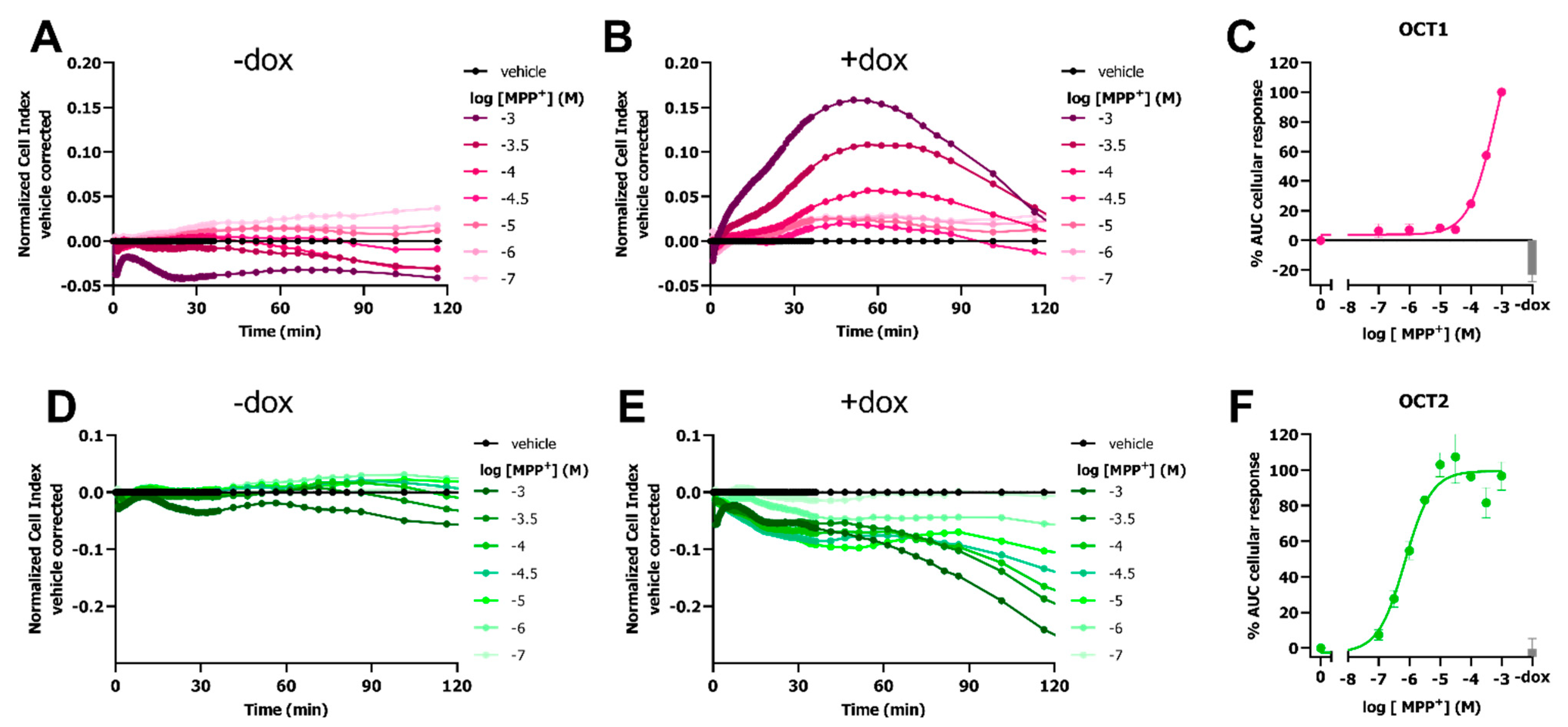
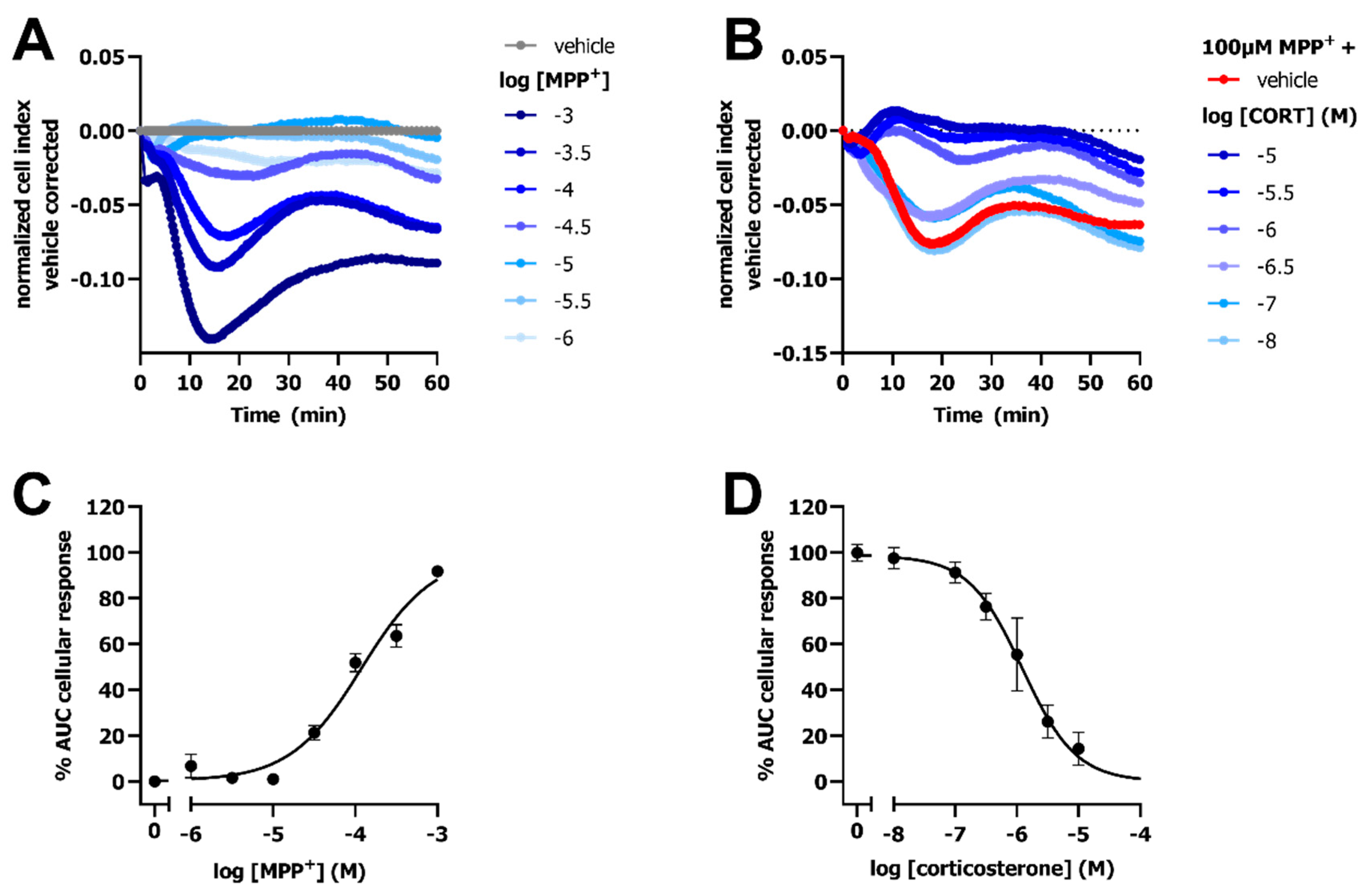
2.1. OCT3-Mediated Transport of MPP+ Induces a Peak Response in an Impedance-Based Cytotoxicity Assay
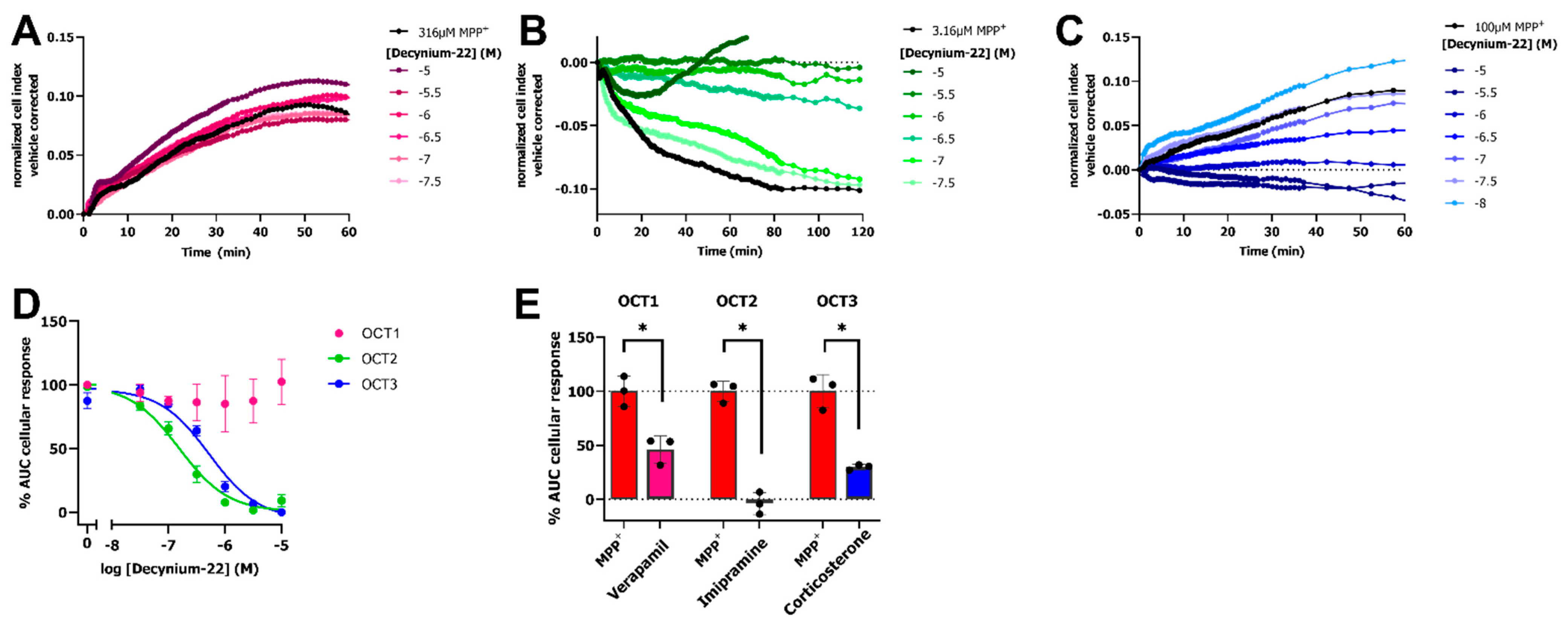
2.2. OCT Inhibitors Block the MPP+-Induced Cellular Response
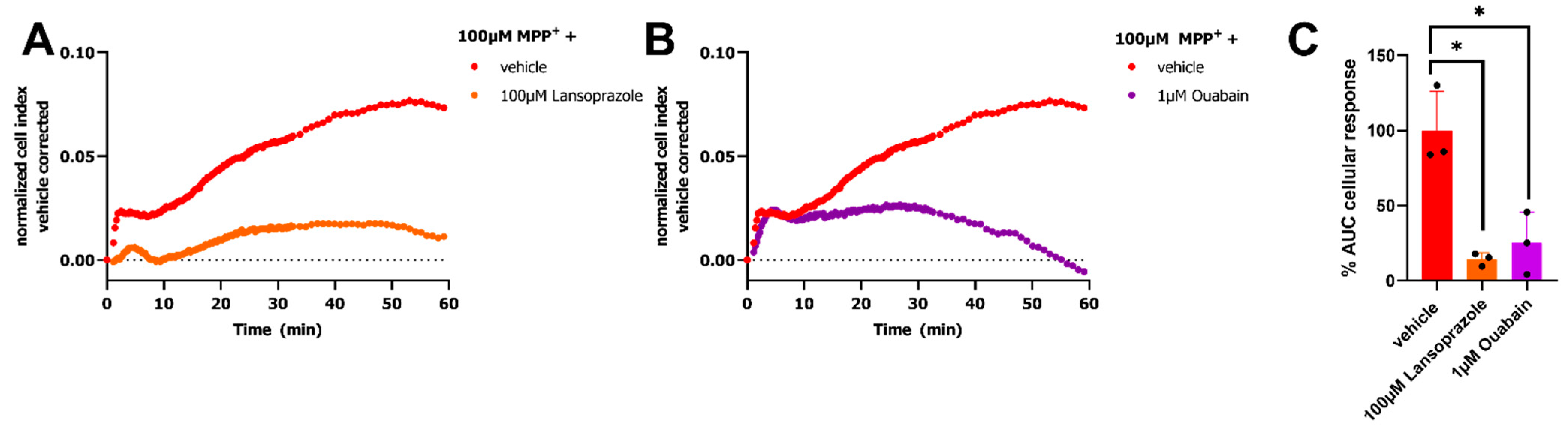
2.3. Maintenance of Electrochemical Gradient Is Fundamental for MPP+ Response
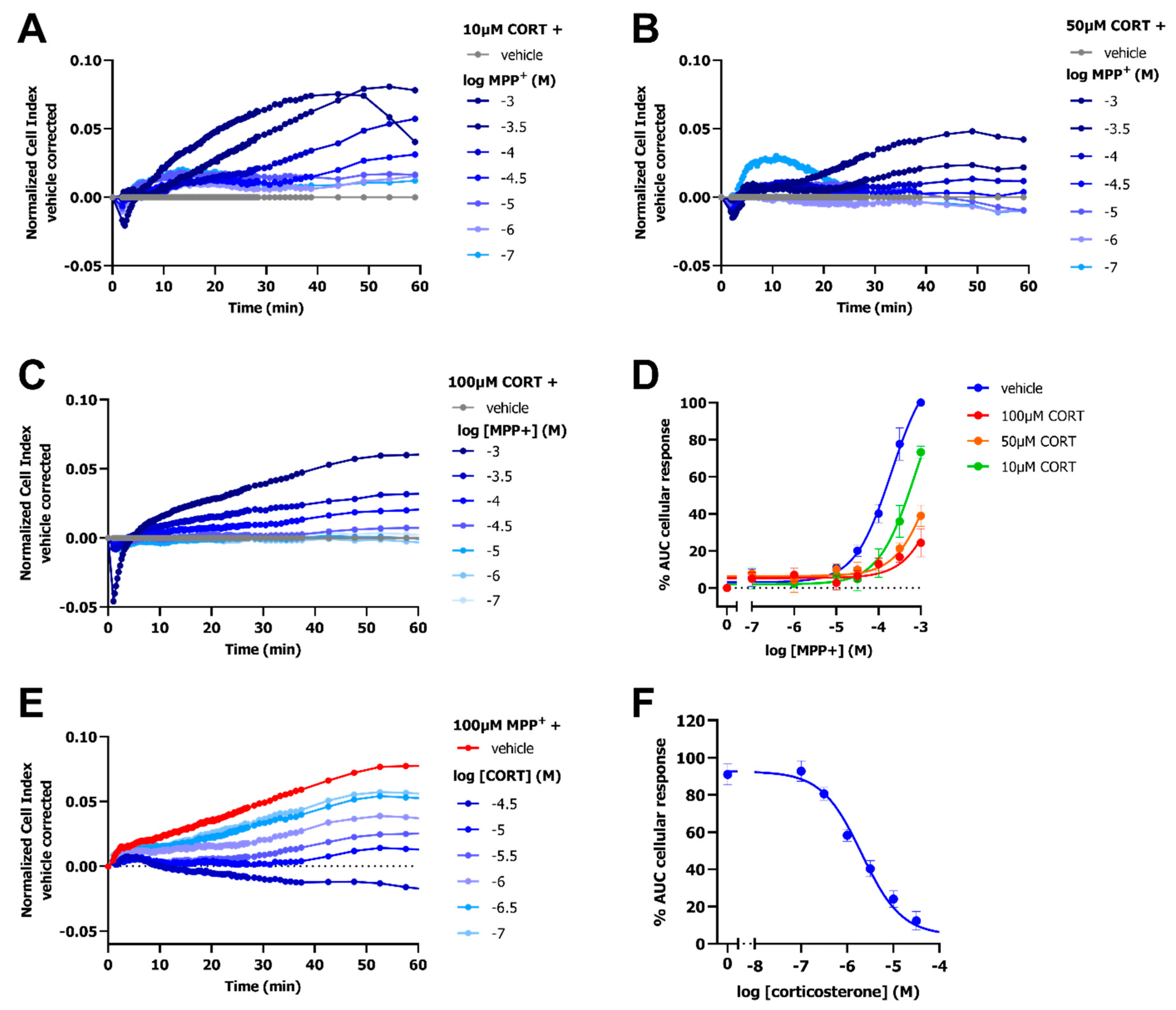
2.4. Inhibition of OCT3 by Corticosterone Abolishes the MPP+-Induced Response
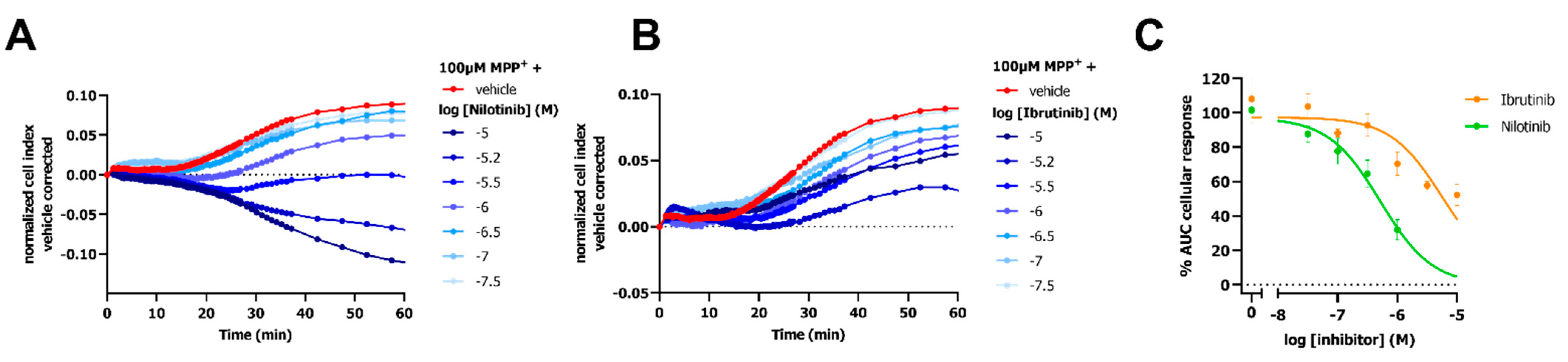
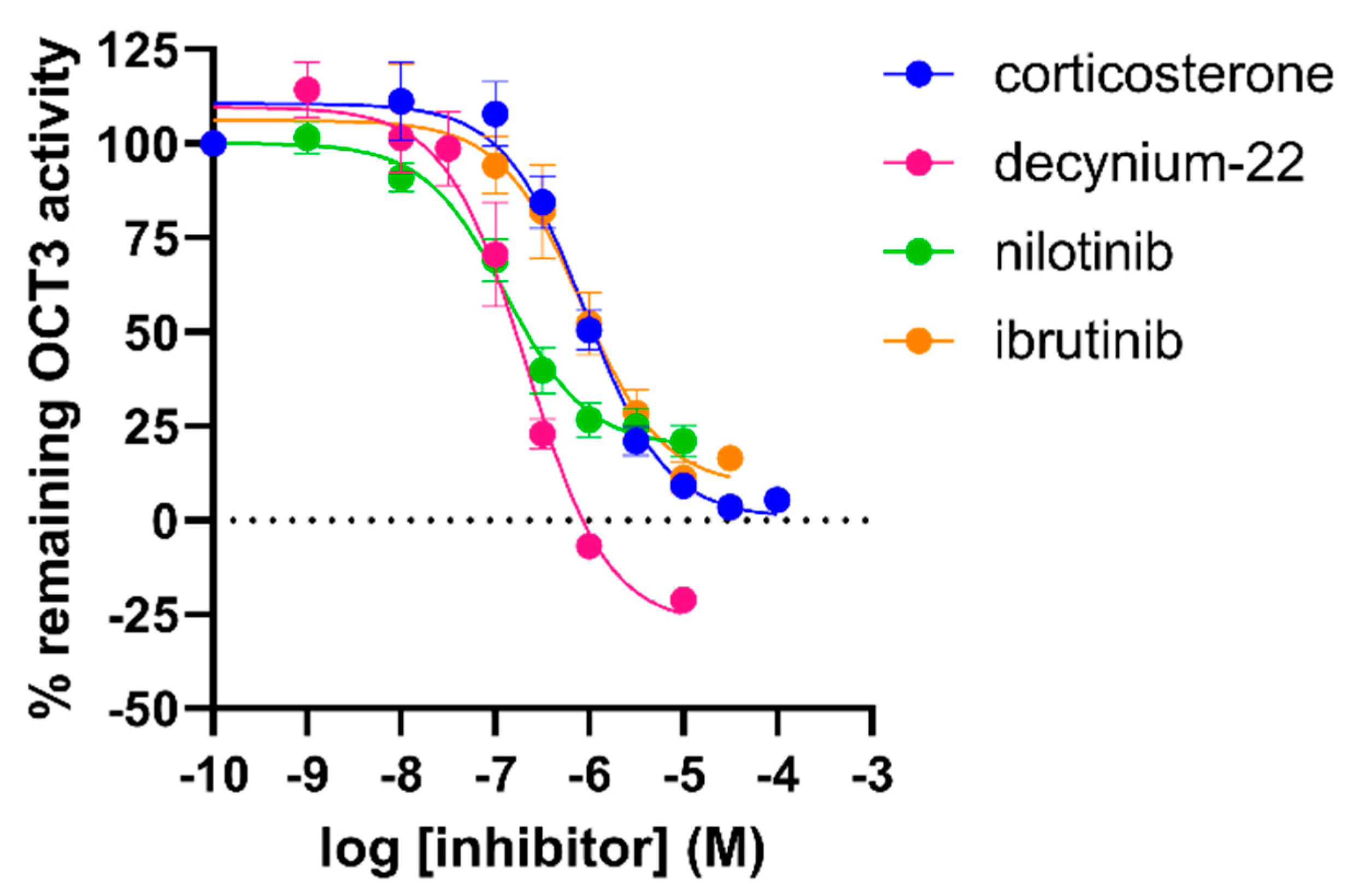
2.5. Attenuation of OCT3 Activity by TKI-Inhibitors
2.6. Inhibitory Potencies from the Impedance-Based Assay Are Comparable to Potencies in an Orthogonal Fluorescent Uptake Assay
2.7. Detection of OCT3-Mediated Transport of MPP+ and Inhibition by Corticosterone in Hela Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Impedance-Based Transport Assay (xCELLigence)
4.4. Fluorescent Substrate Uptake Assay
4.5. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AUC | Area under the curve |
| DAT | Dopamine transporter |
| DDIs | Drug-drug interactions |
| DiASP | 4-(4-diethylaminostyryl)-1-methyl-pyridinium iodide |
| dox | Doxycycline |
| ENT1 | Equilibrative nucleoside transporter 1 |
| Emax | Maximum response |
| GPCR | G protein-coupled receptor |
| HEK | Human embryonic kidney |
| MATE | Multidrug and toxic extrusion |
| MPP+ | 1-Methyl-4-phenylpyridinium iodide |
| NET | Norepinephrine transporter |
| OCT | Organic cation transporter |
| pEC50 | −log (EC50) where EC50 is the concentration required to obtain half maximal response by a compound |
| pIC50 | −log (IC50) where IC50 is concentration inhibitor required to reduce the response by 50% |
| ROS | Reactive oxygen species |
| SLC | Solute carrier protein |
| TKI | Tyrosine kinase inhibitor |
| TRACT | Transport activity through receptor activation |
References
- Wu, X.; Kekuda, R.; Huang, W.; Fei, Y.J.; Leibach, F.H.; Chen, J.; Conway, S.J.; Ganapathy, V. Identity of the Organic Cation Transporter OCT3 as the Extraneuronal Monoamine Transporter (Uptake2) and Evidence for the Expression of the Transporter in the Brain. J. Biol. Chem. 1998, 273, 32776–32786. [Google Scholar] [CrossRef] [PubMed]
- Koepsell, H.; Lips, K.; Volk, C. Polyspecific Organic Cation Transporters: Structure, Function, Physiological Roles, and Biopharmaceutical Implications. Pharm. Res. 2007, 24, 1227–1251. [Google Scholar] [CrossRef] [PubMed]
- Hayer-Zillgen, M.; Brüss, M.; Bönisch, H. Expression and Pharmacological Profile of the Human Organic Cation Transporters HOCT1, HOCT2 and HOCT3. Br. J. Pharmacol. 2002, 136, 829–836. [Google Scholar] [CrossRef] [PubMed]
- Mulgaonkar, A.; Venitz, J.; Gründemann, D.; Sweet, D.H. Human Organic Cation Transporters 1 (SLC22A1), 2 (SLC22A2), and 3 (SLC22A3) as Disposition Pathways for Fluoroquinolone Antimicrobials. Antimicrob. Agents Chemother. 2013, 57, 2705–2711. [Google Scholar] [CrossRef]
- Koepsell, H. Organic Cation Transporters in Health and Disease. Pharmacol. Rev. 2020, 72, 253–319. [Google Scholar] [CrossRef]
- Zhou, S.; Zeng, S.; Shu, Y. Drug-Drug Interactions at Organic Cation Transporter 1. Front. Pharmacol. 2021, 12, 628705. [Google Scholar] [CrossRef]
- Koepsell, H. Update on Drug-Drug Interaction at Organic Cation Transporters: Mechanisms, Clinical Impact, and Proposal for Advanced in Vitro Testing. Expert Opin. Drug Metab. Toxicol. 2021, 17, 635–653. [Google Scholar] [CrossRef]
- Zamek-Gliszczynski, M.J.; Taub, M.E.; Chothe, P.P.; Chu, X.; Giacomini, K.M.; Kim, R.B.; Ray, A.S.; Stocker, S.L.; Unadkat, J.D.; Wittwer, M.B.; et al. Transporters in Drug Development: 2018 ITC Recommendations for Transporters of Emerging Clinical Importance. Clin. Pharmacol. Ther. 2018, 104, 890–899. [Google Scholar] [CrossRef]
- Huang, K.M.; Thomas, M.Z.; Magdy, T.; Eisenmann, E.D.; Uddin, M.E.; DiGiacomo, D.F.; Pan, A.; Keiser, M.; Otter, M.; Xia, S.H.; et al. Targeting OCT3 Attenuates Doxorubicin-Induced Cardiac Injury. Proc. Natl. Acad. Sci. USA 2021, 118, e2020168118. [Google Scholar] [CrossRef]
- Octavia, Y.; Tocchetti, C.G.; Gabrielson, K.L.; Janssens, S.; Crijns, H.J.; Moens, A.L. Doxorubicin-Induced Cardiomyopathy: From Molecular Mechanisms to Therapeutic Strategies. J. Mol. Cell. Cardiol. 2012, 52, 1213–1225. [Google Scholar] [CrossRef]
- Olson, R.D.; Mushlin, P.S. Doxorubicin Cardiotoxicity: Analysis of Prevailing Hypotheses. FASEB J. 1990, 4, 3076–3086. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.E.; Moseley, A. Contribution of Membrane Transporters to Chemotherapy- Induced Cardiotoxicity. Basic Clin. Pharmacol. Toxicol. 2021, 130, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Alim, K.; Moreau, A.; Bruyère, A.; Jouan, E.; Denizot, C.; Nies, A.T.; Parmentier, Y.; Fardel, O. Inhibition of Organic Cation Transporter 3 Activity by Tyrosine Kinase Inhibitors. Fundam. Clin. Pharmacol. 2021, 35, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Minematsu, T.; Giacomini, K.M. Interactions of Tyrosine Kinase Inhibitors with Organic Cation Transporters and Multidrug and Toxic Compound Extrusion Proteins. Mol. Cancer Ther. 2011, 10, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Gorboulev, V.; Rehman, S.; Albert, C.M.; Roth, U.; Meyer, M.J.; Tzvetkov, M.V.; Mueller, T.D.; Koepsell, H. Assay Conditions Influence Affinities of Rat Organic Cation Transporter 1: Analysis of Mutagenesis in the Modeled Outward-Facing Cleft by Measuring Effects of Substrates and Inhibitors on Initial Uptake. Mol. Pharmacol. 2018, 93, 402–415. [Google Scholar] [CrossRef]
- Dvorak, V.; Wiedmer, T.; Ingles-Prieto, A.; Altermatt, P.; Batoulis, H.; Bärenz, F.; Bender, E.; Digles, D.; Dürrenberger, F.; Heitman, L.H.; et al. An Overview of Cell-Based Assay Platforms for the Solute Carrier Family of Transporters. Front. Pharmacol. 2021, 12, 722889. [Google Scholar] [CrossRef]
- Sijben, H.J.; van den Berg, J.J.E.; Broekhuis, J.D.; IJzerman, A.P.; Heitman, L.H. A Study of the Dopamine Transporter Using the TRACT Assay, a Novel in Vitro Tool for Solute Carrier Drug Discovery. Sci. Rep. 2021, 11, 1312. [Google Scholar] [CrossRef]
- Sijben, H.J.; van Oostveen, W.M.; Hartog, P.B.R.; Stucchi, L.; Rossignoli, A.; Maresca, G.; Scarabottolo, L.; IJzerman, A.P.; Heitman, L.H. Label-Free High-Throughput Screening Assay for the Identification of Norepinephrine Transporter (NET/SLC6A2) Inhibitors. Sci. Rep. 2021, 11, 12290. [Google Scholar] [CrossRef]
- Vlachodimou, A.; IJzerman, A.P.; Heitman, L.H. Label-Free Detection of Transporter Activity via GPCR Signalling in Living Cells: A Case for SLC29A1, the Equilibrative Nucleoside Transporter 1. Sci. Rep. 2019, 9, 13802. [Google Scholar] [CrossRef]
- Hillger, J.M.; Schoop, J.; Boomsma, D.I.; Eline Slagboom, P.; IJzerman, A.P.; Heitman, L.H. Whole-Cell Biosensor for Label-Free Detection of GPCR-Mediated Drug Responses in Personal Cell Lines. Biosens. Bioelectron. 2015, 74, 233–242. [Google Scholar] [CrossRef]
- Türker Şener, L.; Albenïz, G.; Dïnç, B.; Albenïz, I. ICELLigence Real-time Cell Analysis System for Examining the Cytotoxicity of Drugs to Cancer Cell Lines (Review). Exp. Ther. Med. 2017, 14, 1866–1870. [Google Scholar] [CrossRef] [PubMed]
- Shang, T.; Uihlein, A.V.; Van Asten, J.; Kalyanaraman, B.; Hillard, C.J. 1-Methyl-4-Phenylpyridinium Accumulates in Cerebellar Granule Neurons via Organic Cation Transporter 3. J. Neurochem. 2003, 85, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Massmann, V.; Edemir, B.; Schlatter, E.; Al-Monajjed, R.; Harrach, S.; Klassen, P.; Holle, S.K.; Sindic, A.; Dobrivojevic, M.; Pavenstädt, H.; et al. The Organic Cation Transporter 3 (OCT3) as Molecular Target of Psychotropic Drugs: Transport Characteristics and Acute Regulation of Cloned Murine OCT3. Pflugers Arch. Eur. J. Physiol. 2014, 466, 517–527. [Google Scholar] [CrossRef]
- Wu, X.; Wei, H.; Ganapathy, M.E.; Wang, H.; Kekuda, R.; Conway, S.J.; Leibach, F.H.; Ganapathy, V. Structure, Function, and Regional Distribution of the Organic Cation Transporter OCT3 in the Kidney. Am. J. Physiol.-Ren. Physiol. 2000, 279, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Sleijfer, D.T.; Offerman, J.J.G.; Mulder, N.H.; Verweij, M.; van der Hem, G.K.; Koops, H.S.; Meijer, S. The Protective Potential of the Combination of Verapamil and Cimetidine on Cisplatin-Induced Nephrotoxicity in Man. Cancer 1987, 60, 2823–2828. [Google Scholar] [CrossRef]
- Orrico-Sanchez, A.; Chausset-Boissarie, L.; Alves de Sousa, R.; Coutens, B.; Rezai Amin, S.; Vialou, V.; Louis, F.; Hessani, A.; Dansette, P.M.; Zornoza, T.; et al. Antidepressant Efficacy of a Selective Organic Cation Transporter Blocker in a Mouse Model of Depression. Mol. Psychiatry 2020, 25, 1245–1259. [Google Scholar] [CrossRef]
- Maier, J.; Niello, M.; Rudin, D.; Daws, L.C.; Sitte, H.H. The Interaction of Organic Cation Transporters 1–3 and PMAT with Psychoactive Substances; Springer: Berlin/Heidelberg, Germany, 2021; pp. 1–16. [Google Scholar] [CrossRef]
- Sun, S.; Wang, K.; Lei, H.; Li, L.; Tu, M.; Zeng, S.; Zhou, H.; Jiang, H. Inhibition of Organic Cation Transporter 2 and 3 May Be Involved in the Mechanism of the Antidepressant-like Action of Berberine. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 49, 1–6. [Google Scholar] [CrossRef]
- Wang, W.W.; Gallo, L.; Jadhav, A.; Hawkins, R.; Parker, C.G. The Druggability of Solute Carriers. J. Med. Chem. 2019, 63, 3834–3867. [Google Scholar] [CrossRef]
- Ciarimboli, G.; Schlatter, E. Organic Cation Transport Measurements Using Fluorescence Techniques BT—Neurotransmitter Transporters: Investigative Methods; Bönisch, H., Sitte, H.H., Eds.; Springer: New York, NY, USA, 2016; pp. 173–187. [Google Scholar] [CrossRef]
- Wong, S.H.; Gao, A.; Ward, S.; Henley, C.; Lee, P.H. Development of a Label-Free Assay for Sodium-Dependent Phosphate Transporter NaPi-IIb. J. Biomol. Screen. 2012, 17, 829–834. [Google Scholar] [CrossRef][Green Version]
- Lin, Z.; Liu, C.; Fan, E.; Zhang, Y.; Zheng, S.; Rao, Y. An Efficient and Label-Free LC-MS/MS Method for Assessing Drug’s Activity at Dopamine and Serotonin Transporters Using Transporter-Transfected HEK293T Cells. J. Psychopharmacol. 2021, 35, 693–700. [Google Scholar] [CrossRef]
- Bailey, T.L.; Nieto, A.; McDonald, P.H. A Nonradioactive High-Throughput Screening-Compatible Cell-Based Assay to Identify Inhibitors of the Monocarboxylate Transporter Protein 1. Assay Drug Dev. Technol. 2019, 17, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Martel, F.; Keating, E.; Calhau, C.; Gründemann, D.; Schömig, E.; Azevedo, I. Regulation of Human Extraneuronal Monoamine Transporter (HEMT) Expressed in HEK293 Cells by Intracellular Second Messenger Systems. Naunyn. Schmiedebergs. Arch. Pharmacol. 2001, 364, 487–495. [Google Scholar] [CrossRef]
- Gorboulev, V.; Ulzheimer, J.C.; Akhoundova, A.; Ulzheime-Teuber, I.; Karbach, U.; Quester, S.; Baumann, C.; Lang, F.; Koepsell, H. Cloning and Characterization of Two Human Polyspecific Organic Cation Transporters. DNA Cell Biol. 1997, 16, 871–881. [Google Scholar] [CrossRef] [PubMed]
- Sala-Rabanal, M.; Li, D.C.; Dake, G.R.; Kurata, H.T.; Inyushin, M.; Skatchkov, S.N.; Nichols, C.G. Polyamine Transport by the Polyspecific Organic Cation Transporters OCT1, OCT2, and OCT3. Mol. Pharm. 2013, 10, 1450–1458. [Google Scholar] [CrossRef] [PubMed]
- Kitayama, S.; Dohi, T. Cellular and Molecular Aspects of Monoamine Neurotransmitter Transporters. Jpn. J. Pharmacol. 1996, 72, 195–208. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yang, S.A.; Yoon, J.; Kim, K.; Park, Y.K. Measurements of Morphological and Biophysical Alterations in Individual Neuron Cells Associated with Early Neurotoxic Effects in Parkinson’s Disease. Cytom. Part A 2017, 91, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Allani, P.K.; Sum, T.; Bhansali, S.G.; Mukherjee, S.K.; Sonee, M. A Comparative Study of the Effect of Oxidative Stress on the Cytoskeleton in Human Cortical Neurons. Toxicol. Appl. Pharmacol. 2004, 196, 29–36. [Google Scholar] [CrossRef]
- Mapa, M.S.T.; Le, V.Q.; Wimalasena, K. Characteristics of the Mitochondrial and Cellular Uptake of MPP+, as Probed by the Fluorescent Mimic, 4′I-MPP+. PLoS ONE 2018, 13, e0197946. [Google Scholar] [CrossRef]
- Schmidt, N.; Ferger, B. Neurochemical Findings in the MPTP Model of Parkinson’s Disease. J. Neural Transm. 2001, 108, 1263–1282. [Google Scholar] [CrossRef]
- Javadov, S.; Chapa-Dubocq, X.; Makarov, V. Different Approaches to Modeling Analysis of Mitochondrial Swelling. Mitochondrion 2018, 38, 58–70. [Google Scholar] [CrossRef]
- Cartelli, D.; Ronchi, C.; Maggioni, M.G.; Rodighiero, S.; Giavini, E.; Cappelletti, G. Microtubule Dysfunction Precedes Transport Impairment and Mitochondria Damage in MPP+-Induced Neurodegeneration. J. Neurochem. 2010, 115, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Weber, A.; Iturri, J.; Benitez, R.; Zemljic-Jokhadar, S.; Toca-Herrera, J.L. Microtubule Disruption Changes Endothelial Cell Mechanics and Adhesion. Sci. Rep. 2019, 9, 14903. [Google Scholar] [CrossRef] [PubMed]
- Sirinathsinghji, D.J.S.; Heavens, R.P.; McBride, C.S. Dopamine-Releasing Action of 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine (MPTP) and 1-Methyl-4-Phenylpyridine (MPP+) in the Neostriatum of the Rat as Demonstrated in Vivo by the Push-Pull Perfusion Technique: Dependence on Sodium but Not Calcium Ions. Brain Res. 1988, 443, 101–116. [Google Scholar] [CrossRef]
- Lin, K.H.; Li, C.Y.; Hsu, Y.M.; Tsai, C.H.; Tsai, F.J.; Tang, C.H.; Yang, J.S.; Wang, Z.H.; Yin, M.C. Oridonin, A Natural Diterpenoid, Protected NGF-Differentiated PC12 Cells against MPP+- and Kainic Acid-Induced Injury. Food Chem. Toxicol. 2019, 133, 110765. [Google Scholar] [CrossRef]
- Fraser-Spears, R.; Krause-Heuer, A.M.; Basiouny, M.; Mayer, F.P.; Manishimwe, R.; Wyatt, N.A.; Dobrowolski, J.C.; Roberts, M.P.; Greguric, I.; Kumar, N.; et al. Comparative Analysis of Novel Decynium-22 Analogs to Inhibit Transport by the Low-Affinity, High-Capacity Monoamine Transporters, Organic Cation Transporters 2 and 3, and Plasma Membrane Monoamine Transporter. Eur. J. Pharmacol. 2019, 842, 351–364. [Google Scholar] [CrossRef]
- Ahlin, G.; Chen, L.; Lazorova, L.; Chen, Y.; Ianculescu, A.G.; Davis, R.L.; Giacomini, K.M.; Artursson, P. Genotype-Dependent Effects of Inhibitors of the Organic Cation Transporter, OCT1: Predictions of Metformin Interactions. Pharmacogenomics J. 2011, 11, 400–411. [Google Scholar] [CrossRef]
- Koepsell, H. Multiple Binding Sites in Organic Cation Transporters Require Sophisticated Procedures to Identify Interactions of Novel Drugs. Biol. Chem. 2018, 400, 195–207. [Google Scholar] [CrossRef]
- Sandoval, P.J.; Zorn, K.M.; Clark, A.M.; Ekins, S.; Wright, S.H. Assessment of Substrate-Dependent Ligand Interactions at the Organic Cation Transporter OCT2 Using Six Model Substrates S. Mol. Pharmacol. 2018, 94, 1057–1068. [Google Scholar] [CrossRef]
- Nederpelt, I.; Vergroesen, R.D.; IJzerman, A.P.; Heitman, L.H. Persistent GnRH Receptor Activation in Pituitary AT3-1 Cells Analyzed with a Label-Free Technology. Biosens. Bioelectron. 2016, 79, 721–727. [Google Scholar] [CrossRef]
- Ke, N.; Wang, X.; Xu, X.; Abassi, Y.A. The XCELLigence System for Real-Time and Label-Free Monitoring of Cell Viability BT—Mammalian Cell Viability: Methods and Protocols; Stoddart, M.J., Ed.; Humana Press: Totowa, NJ, USA, 2011; pp. 33–43. [Google Scholar] [CrossRef]
- Peters, M.F.; Scott, C.W. Evaluating Cellular Impedance Assays for Detection of GPCR Pleiotropic Signaling and Functional Selectivity. J. Biomol. Screen. 2009, 14, 246–255. [Google Scholar] [CrossRef]
- Nies, A.T.; Koepsell, H.; Winter, S.; Burk, O.; Klein, K.; Kerb, R.; Zanger, U.M.; Keppler, D.; Schwab, M.; Schaeffeler, E. Expression of Organic Cation Transporters OCT1 (SLC22A1) and OCT3 (SLC22A3) Is Affected by Genetic Factors and Cholestasis in Human Liver. Hepatology 2009, 50, 1227–1240. [Google Scholar] [CrossRef] [PubMed]
- Motohashi, H.; Nakao, Y.; Masuda, S.; Katsura, T.; Kamba, T.; Ogawa, O.; Inui, K.-I. Precise Comparison of Protein Localization among OCT, OAT, and MATE in Human Kidney. J. Pharm. Sci. 2013, 102, 3302–3308. [Google Scholar] [CrossRef] [PubMed]
- Duan, H.; Hu, T.; Foti, R.S.; Pan, Y.; Swaan, P.W.; Wang, J. Potent and Selective Inhibition of Plasma Membrane Monoamine Transporter by HIV Protease Inhibitors. Drug Metab. Dispos. 2015, 43, 1773–1780. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | pIC50 | |
|---|---|---|
| xCELLigence | Fluorescent Uptake Assay | |
| Decynium-22 | 6.4 ± 0.0 (3) | 6.7 ± 0.1 (3) |
| Corticosterone | 5.8 ± 0.1 (7) | 6.1 ± 0.1 (6) |
| Nilotinib | 6.3 ± 0.1 (3) * | 6.9 ± 0.1 (3) |
| Ibrutinib | 5.1 ± 0.2 (3) *** | 6.1 ± 0.2 (4) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mocking, T.A.M.; Sijben, H.J.; Vermeulen, Y.W.; IJzerman, A.P.; Heitman, L.H. MPP+-Induced Changes in Cellular Impedance as a Measure for Organic Cation Transporter (SLC22A1-3) Activity and Inhibition. Int. J. Mol. Sci. 2022, 23, 1203. https://doi.org/10.3390/ijms23031203
Mocking TAM, Sijben HJ, Vermeulen YW, IJzerman AP, Heitman LH. MPP+-Induced Changes in Cellular Impedance as a Measure for Organic Cation Transporter (SLC22A1-3) Activity and Inhibition. International Journal of Molecular Sciences. 2022; 23(3):1203. https://doi.org/10.3390/ijms23031203
Chicago/Turabian StyleMocking, Tamara A. M., Hubert J. Sijben, Yimé W. Vermeulen, Adriaan P. IJzerman, and Laura H. Heitman. 2022. "MPP+-Induced Changes in Cellular Impedance as a Measure for Organic Cation Transporter (SLC22A1-3) Activity and Inhibition" International Journal of Molecular Sciences 23, no. 3: 1203. https://doi.org/10.3390/ijms23031203
APA StyleMocking, T. A. M., Sijben, H. J., Vermeulen, Y. W., IJzerman, A. P., & Heitman, L. H. (2022). MPP+-Induced Changes in Cellular Impedance as a Measure for Organic Cation Transporter (SLC22A1-3) Activity and Inhibition. International Journal of Molecular Sciences, 23(3), 1203. https://doi.org/10.3390/ijms23031203