

Receptor-Independent Anti-Ferroptotic Activity of TrkB Modulators

 ,
,  ,
, 
 and
and
Abstract
1. Introduction
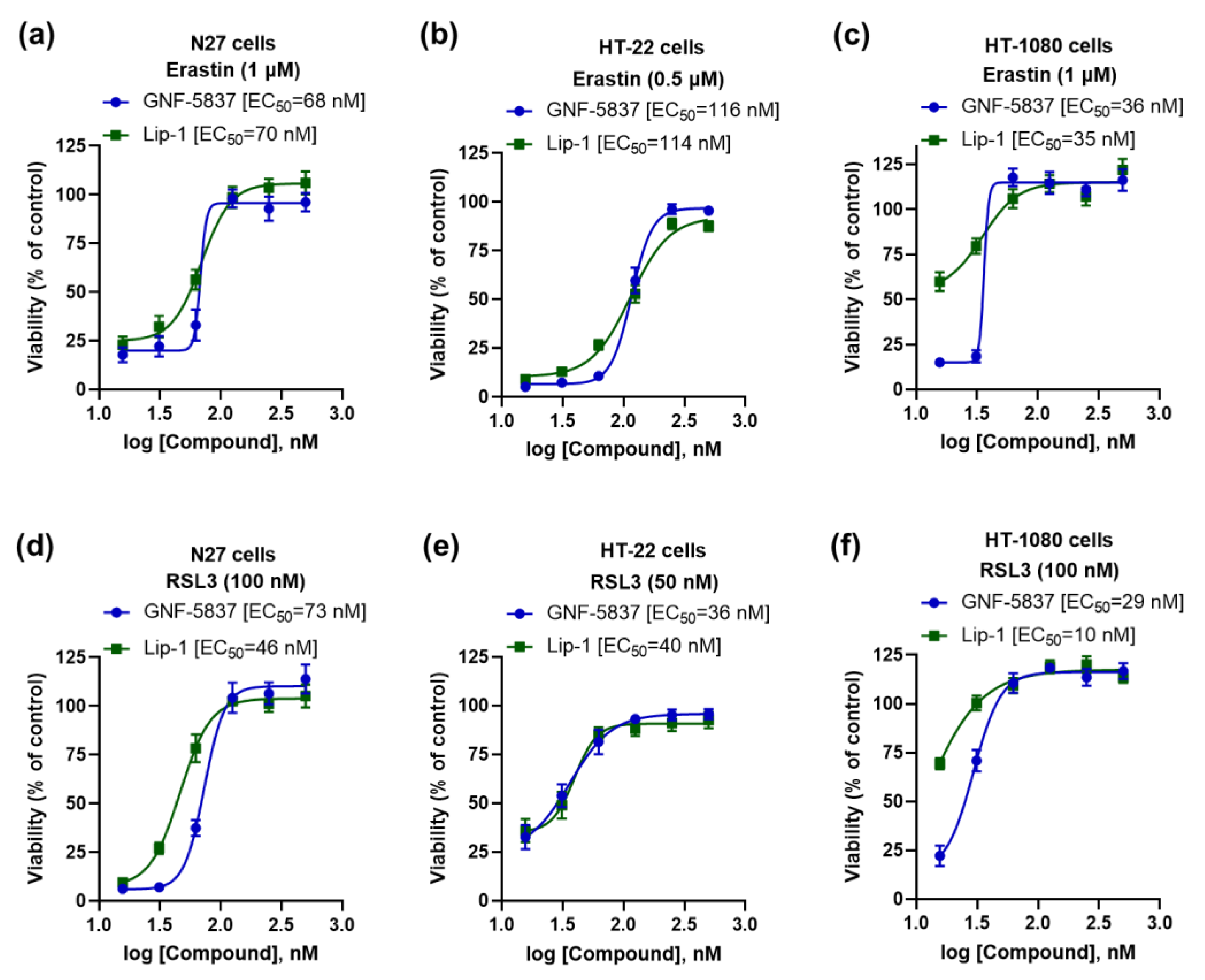
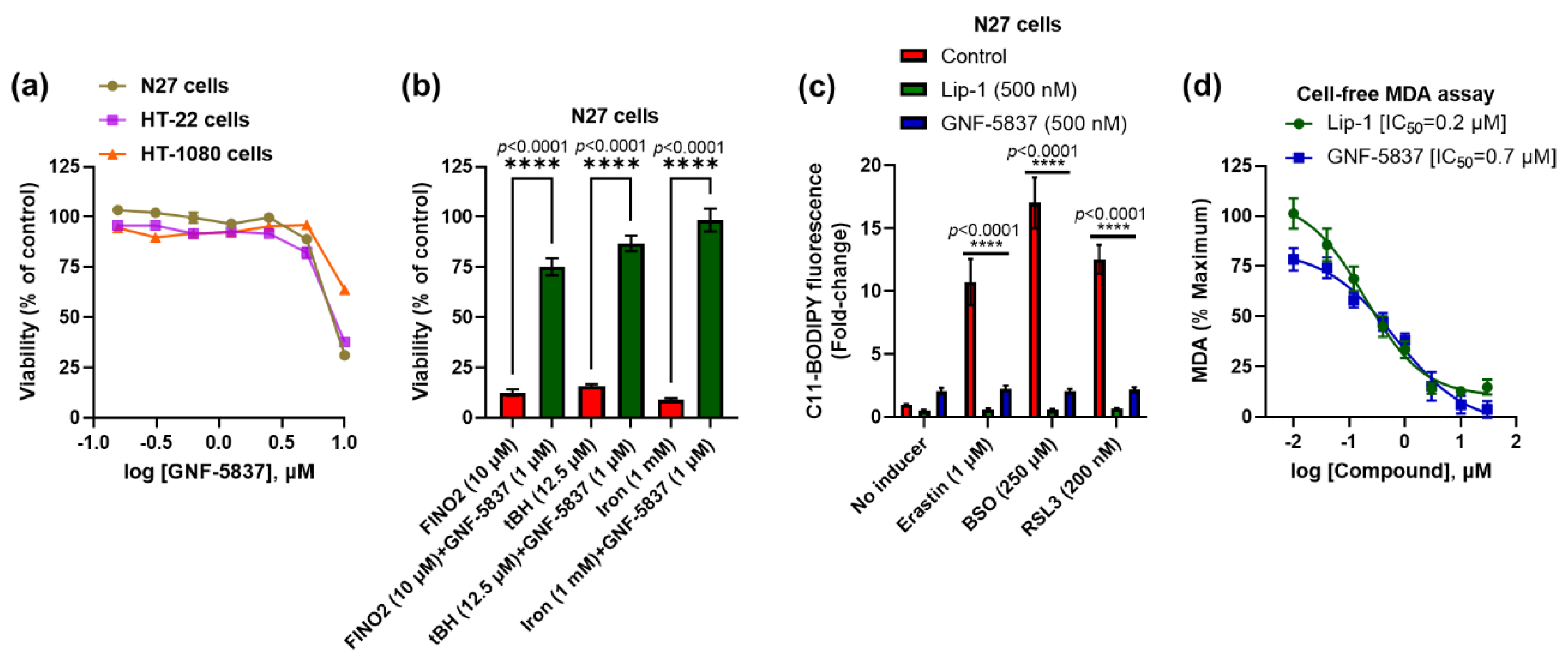
2. Results
3. Discussion
4. Materials and Methods
4.1. Drugs and Chemicals
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Radical-Trapping Antioxidant Assay
4.5. Lipid Peroxidation Measurement
4.6. Western Blotting Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 7,8-DHF | 7,8-dihydroxyflavone |
| ABTS | 2,2’-Azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt |
| Aβ | amyloid-beta |
| BBB | blood–brain barrier |
| BDNF | brain-derived neurotrophic factor |
| BSO | L-buthionine-sulfoximine |
| DFP | deferiprone |
| GPX4 | glutathione peroxidase-4 |
| Lip-1 | liproxstatin-1 |
| MDA | malondialdehyde |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| tBH | tert-Butyl hydroperoxide |
| TrkB | tropomyosin receptor kinase B |
References
- Minichiello, L. Trkb signalling pathways in ltp and learning. Nat. Rev. Neurosci. 2009, 10, 850–860. [Google Scholar] [CrossRef]
- Luberg, K.; Wong, J.; Weickert, C.S.; Timmusk, T. Human trkb gene: Novel alternative transcripts, protein isoforms and expression pattern in the prefrontal cerebral cortex during postnatal development. J. Neurochem. 2010, 113, 952–964. [Google Scholar] [CrossRef]
- Jerónimo-Santos, A.; Vaz, S.H.; Parreira, S.; Rapaz-Lérias, S.; Caetano, A.P.; Buée-Scherrer, V.; Castrén, E.; Valente, C.A.; Blum, D.; Sebastião, A.M.; et al. Dysregulation of trkb receptors and bdnf function by amyloid-β peptide is mediated by calpain. Cereb. Cortex 2014, 25, 3107–3121. [Google Scholar] [CrossRef]
- Xia, Y.; Wang, Z.-H.; Liu, P.; Edgington-Mitchell, L.; Liu, X.; Wang, X.-C.; Ye, K. Trkb receptor cleavage by delta-secretase abolishes its phosphorylation of app, aggravating Alzheimer’s disease pathologies. Mol. Psychiatry 2021, 26, 2943–2963. [Google Scholar] [CrossRef]
- Xiang, J.; Wang, Z.H.; Ahn, E.H.; Liu, X.; Yu, S.P.; Manfredsson, F.P.; Sandoval, I.M.; Ju, G.; Wu, S.; Ye, K. Delta-secretase-cleaved tau antagonizes trkb neurotrophic signalings, mediating Alzheimer’s disease pathologies. Proc. Natl. Acad. Sci. USA 2019, 116, 9094–9102. [Google Scholar] [CrossRef]
- Wang, Z.H.; Xiang, J.; Liu, X.; Yu, S.P.; Manfredsson, F.P.; Sandoval, I.M.; Wu, S.; Wang, J.Z.; Ye, K. Deficiency in bdnf/trkb neurotrophic activity stimulates δ-secretase by upregulating c/ebpβ in Alzheimer’s disease. Cell Rep. 2019, 28, 655–669.e655. [Google Scholar] [CrossRef]
- Ferrer, I.; Marín, C.; Rey, M.J.; Ribalta, T.; Goutan, E.; Blanco, R.; Tolosa, E.; Martí, E. Bdnf and full-length and truncated trkb expression in Alzheimer disease. Implications in therapeutic strategies. J. Neuropathol. Exp. Neurol. 1999, 58, 729–739. [Google Scholar] [CrossRef]
- Boots, E.A.; Schultz, S.A.; Clark, L.R.; Racine, A.M.; Darst, B.F.; Koscik, R.L.; Carlsson, C.M.; Gallagher, C.L.; Hogan, K.J.; Bendlin, B.B.; et al. Bdnf val66met predicts cognitive decline in the wisconsin registry for alzheimer’s prevention. Neurology 2017, 88, 2098–2106. [Google Scholar] [CrossRef]
- Lim, Y.Y.; Hassenstab, J.; Cruchaga, C.; Goate, A.; Fagan, A.M.; Benzinger, T.L.; Maruff, P.; Snyder, P.J.; Masters, C.L.; Allegri, R.; et al. Bdnf val66met moderates memory impairment, hippocampal function and tau in preclinical autosomal dominant Alzheimer’s disease. Brain J. Neurol. 2016, 139, 2766–2777. [Google Scholar] [CrossRef]
- Tuo, Q.-Z.; Masaldan, S.; Southon, A.; Mawal, C.; Ayton, S.; Bush, A.I.; Lei, P.; Belaidi, A.A. Characterization of selenium compounds for anti-ferroptotic activity in neuronal cells and after cerebral ischemia–reperfusion injury. Neurotherapeutics 2021, 18, 2682–2691. [Google Scholar] [CrossRef]
- Southon, A.; Szostak, K.; Acevedo, K.M.; Dent, K.A.; Volitakis, I.; Belaidi, A.A.; Barnham, K.J.; Crouch, P.J.; Ayton, S.; Donnelly, P.S.; et al. Cu(ii) (atsm) inhibits ferroptosis: Implications for treatment of neurodegenerative disease. Br. J. Pharmacol. 2020, 177, 656–667. [Google Scholar] [CrossRef] [PubMed]
- Jakaria, M.; Belaidi, A.A.; Bush, A.I.; Ayton, S. Ferroptosis as a mechanism of neurodegeneration in Alzheimer’s disease. J. Neurochem. 2021, 159, 804–825. [Google Scholar] [CrossRef] [PubMed]
- Belaidi, A.A.; Masaldan, S.; Southon, A.; Kalinowski, P.; Acevedo, K.; Appukuttan, A.T.; Portbury, S.; Lei, P.; Agarwal, P.; Leurgans, S.E.; et al. Apolipoprotein e potently inhibits ferroptosis by blocking ferritinophagy. Mol. Psychiatry 2022. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.-B.; Kim, H.; El Touny, L.; Simeonov, A.; Gerhold, D. Luhmes dopaminergic neurons are uniquely susceptible to ferroptosis. Neurotox. Res. 2022, 40, 1526–1536. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Stockwell, B.R.; Conrad, M. Ferroptosis: Mechanisms, biology and role in disease. Nat. Rev. Mol. Cell Biol. 2021, 22, 266–282. [Google Scholar] [CrossRef]
- Xu, D.; Lian, D.; Wu, J.; Liu, Y.; Zhu, M.; Sun, J.; He, D.; Li, L. Brain-derived neurotrophic factor reduces inflammation and hippocampal apoptosis in experimental streptococcus pneumoniae meningitis. J. Neuroinflammation 2017, 14, 156. [Google Scholar] [CrossRef]
- Wu, S.-Y.; Pan, B.-S.; Tsai, S.-F.; Chiang, Y.-T.; Huang, B.-M.; Mo, F.-E.; Kuo, Y.-M. Bdnf reverses aging-related microglial activation. J. Neuroinflammation 2020, 17, 210. [Google Scholar] [CrossRef]
- Chan, S.H.; Wu, C.W.; Chang, A.Y.; Hsu, K.S.; Chan, J.Y. Transcriptional upregulation of brain-derived neurotrophic factor in rostral ventrolateral medulla by angiotensin ii: Significance in superoxide homeostasis and neural regulation of arterial pressure. Circ. Res. 2010, 107, 1127–1139. [Google Scholar] [CrossRef]
- Jiang, J.M.; Zhou, C.F.; Gao, S.L.; Tian, Y.; Wang, C.Y.; Wang, L.; Gu, H.F.; Tang, X.Q. Bdnf-trkb pathway mediates neuroprotection of hydrogen sulfide against formaldehyde-induced toxicity to pc12 cells. PLoS ONE 2015, 10, e0119478. [Google Scholar] [CrossRef]
- Almeida, R.D.; Manadas, B.J.; Melo, C.V.; Gomes, J.R.; Mendes, C.S.; Grãos, M.M.; Carvalho, R.F.; Carvalho, A.P.; Duarte, C.B. Neuroprotection by bdnf against glutamate-induced apoptotic cell death is mediated by erk and pi3-kinase pathways. Cell Death Differ. 2005, 12, 1329–1343. [Google Scholar] [CrossRef]
- Melo, C.V.; Okumoto, S.; Gomes, J.R.; Baptista, M.S.; Bahr, B.A.; Frommer, W.B.; Duarte, C.B. Spatiotemporal resolution of bdnf neuroprotection against glutamate excitotoxicity in cultured hippocampal neurons. Neuroscience 2013, 237, 66–86. [Google Scholar] [CrossRef]
- Lau, D.; Bengtson, C.P.; Buchthal, B.; Bading, H. Bdnf reduces toxic extrasynaptic nmda receptor signaling via synaptic nmda receptors and nuclear-calcium-induced transcription of inhba/activin a. Cell Rep. 2015, 12, 1353–1366. [Google Scholar] [CrossRef]
- Nguyen, N.; Lee, S.B.; Lee, Y.S.; Lee, K.H.; Ahn, J.Y. Neuroprotection by ngf and bdnf against neurotoxin-exerted apoptotic death in neural stem cells are mediated through trk receptors, activating pi3-kinase and mapk pathways. Neurochem. Res. 2009, 34, 942–951. [Google Scholar] [CrossRef]
- Nguyen, T.L.X.; Kim, C.K.; Cho, J.-H.; Lee, K.-H.; Ahn, J.-Y. Neuroprotection signaling pathway of nerve growth factor and brain-derived neurotrophic factor against staurosporine induced apoptosis in hippocampal h19-7 cells. Exp. Mol. Med. 2010, 42, 583–595. [Google Scholar] [CrossRef]
- Suzuki, S.; Kiyosue, K.; Hazama, S.; Ogura, A.; Kashihara, M.; Hara, T.; Koshimizu, H.; Kojima, M. Brain-derived neurotrophic factor regulates cholesterol metabolism for synapse development. J. Neurosci. Off. J. Soc. Neurosci. 2007, 27, 6417–6427. [Google Scholar] [CrossRef]
- Casarotto, P.C.; Girych, M.; Fred, S.M.; Kovaleva, V.; Moliner, R.; Enkavi, G.; Biojone, C.; Cannarozzo, C.; Sahu, M.P.; Kaurinkoski, K.; et al. Antidepressant drugs act by directly binding to trkb neurotrophin receptors. Cell 2021, 184, 1299–1313.e1219. [Google Scholar] [CrossRef]
- Pereira, D.B.; Chao, M.V. The tyrosine kinase fyn determines the localization of trkb receptors in lipid rafts. J. Neurosci. Off. J. Soc. Neurosci. 2007, 27, 4859–4869. [Google Scholar] [CrossRef]
- Martin, M.G.; Perga, S.; Trovò, L.; Rasola, A.; Holm, P.; Rantamäki, T.; Harkany, T.; Castrén, E.; Chiara, F.; Dotti, C.G. Cholesterol loss enhances trkb signaling in hippocampal neurons aging in vitro. Mol. Biol. Cell 2008, 19, 2101–2112. [Google Scholar] [CrossRef]
- Castello, N.A.; Nguyen, M.H.; Tran, J.D.; Cheng, D.; Green, K.N.; LaFerla, F.M. 7,8-dihydroxyflavone, a small molecule trkb agonist, improves spatial memory and increases thin spine density in a mouse model of Alzheimer disease-like neuronal loss. PLoS ONE 2014, 9, e91453. [Google Scholar] [CrossRef]
- Gao, L.; Tian, M.; Zhao, H.Y.; Xu, Q.Q.; Huang, Y.M.; Si, Q.C.; Tian, Q.; Wu, Q.M.; Hu, X.M.; Sun, L.B.; et al. Trkb activation by 7, 8-dihydroxyflavone increases synapse ampa subunits and ameliorates spatial memory deficits in a mouse model of Alzheimer’s disease. J. Neurochem. 2016, 136, 620–636. [Google Scholar] [CrossRef]
- Lee, C.S.; Han, E.S.; Lee, W.B. Antioxidant effect of phenelzine on mpp+-induced cell viability loss in differentiated pc12 cells. Neurochem. Res. 2003, 28, 1833–1841. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.L.; Singh, I.N.; Wang, J.A.; Hall, E.D. Effects of phenelzine administration on mitochondrial function, calcium handling, and cytoskeletal degradation after experimental traumatic brain injury. J. Neurotrauma 2019, 36, 1231–1251. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wang, Z.; Zhang, Z.; Liu, X.; Kang, S.S.; Zhang, Y.; Ye, K. The prodrug of 7,8-dihydroxyflavone development and therapeutic efficacy for treating Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2018, 115, 578–583. [Google Scholar] [CrossRef]
- Fred, S.M.; Laukkanen, L.; Brunello, C.A.; Vesa, L.; Göös, H.; Cardon, I.; Moliner, R.; Maritzen, T.; Varjosalo, M.; Casarotto, P.C.; et al. Pharmacologically diverse antidepressants facilitate trkb receptor activation by disrupting its interaction with the endocytic adaptor complex ap-2. J. Biol. Chem. 2019, 294, 18150–18161. [Google Scholar] [CrossRef]
- Rantamäki, T.; Vesa, L.; Antila, H.; Di Lieto, A.; Tammela, P.; Schmitt, A.; Lesch, K.-P.; Rios, M.; Castrén, E. Antidepressant drugs transactivate trkb neurotrophin receptors in the adult rodent brain independently of bdnf and monoamine transporter blockade. PLoS ONE 2011, 6, e20567. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, A.G.; Al-Shboul, O.; Alfaqih, M.A.; Al-Qudah, M.A.; Al-Dwairi, A.N. Phenelzine reduces the oxidative damage induced by peroxynitrite in plasma lipids and proteins. Arch. Physiol. Biochem. 2018, 124, 418–423. [Google Scholar] [CrossRef]
- Cazorla, M.; Prémont, J.; Mann, A.; Girard, N.; Kellendonk, C.; Rognan, D. Identification of a low-molecular weight trkb antagonist with anxiolytic and antidepressant activity in mice. J. Clin. Investig. 2011, 121, 1846–1857. [Google Scholar] [CrossRef]
- Tapley, P.; Lamballe, F.; Barbacid, M. K252a is a selective inhibitor of the tyrosine protein kinase activity of the trk family of oncogenes and neurotrophin receptors. Oncogene 1992, 7, 371–381. [Google Scholar]
- Jang, S.-W.; Liu, X.; Yepes, M.; Shepherd, K.R.; Miller, G.W.; Liu, Y.; Wilson, W.D.; Xiao, G.; Blanchi, B.; Sun, Y.E.; et al. A selective trkb agonist with potent neurotrophic activities by 7,8-dihydroxyflavone. Proc. Natl. Acad. Sci. USA 2010, 107, 2687–2692. [Google Scholar] [CrossRef]
- Roux, P.P.; Dorval, G.; Boudreau, M.; Angers-Loustau, A.; Morris, S.J.; Makkerh, J.; Barker, P.A. K252a and cep1347 are neuroprotective compounds that inhibit mixed-lineage kinase-3 and induce activation of akt and erk. J. Biol. Chem. 2002, 277, 49473–49480. [Google Scholar] [CrossRef]
- Pan, J.; Zhang, Q.G.; Zhang, G.Y. The neuroprotective effects of k252a through inhibiting mlk3/mkk7/jnk3 signaling pathway on ischemic brain injury in rat hippocampal ca1 region. Neuroscience 2005, 131, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Hou, X.Y.; Liu, Y.; Zong, Y.Y. Different protection of k252a and n-acetyl-l-cysteine against amyloid-beta peptide-induced cortical neuron apoptosis involving inhibition of mlk3-mkk7-jnk3 signal cascades. J. Neurosci. Res. 2009, 87, 918–927. [Google Scholar] [CrossRef] [PubMed]
- Albaugh, P.; Fan, Y.; Mi, Y.; Sun, F.; Adrian, F.; Li, N.; Jia, Y.; Sarkisova, Y.; Kreusch, A.; Hood, T.; et al. Discovery of gnf-5837, a selective trk inhibitor with efficacy in rodent cancer tumor models. ACS Med. Chem. Lett. 2012, 3, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, D.; Takahashi, K.; Ohe, T.; Nakamura, S.; Mashino, T. Antioxidant activities of 5-hydroxyoxindole and its 3-hydroxy-3-phenacyl derivatives: The suppression of lipid peroxidation and intracellular oxidative stress. Bioorg. Med. Chem. 2013, 21, 7709–7714. [Google Scholar] [CrossRef] [PubMed]
- Hirata, Y.; Yamada, C.; Ito, Y.; Yamamoto, S.; Nagase, H.; Oh-Hashi, K.; Kiuchi, K.; Suzuki, H.; Sawada, M.; Furuta, K. Novel oxindole derivatives prevent oxidative stress-induced cell death in mouse hippocampal ht22 cells. Neuropharmacology 2018, 135, 242–252. [Google Scholar] [CrossRef]
- Hirata, Y.; Ito, Y.; Takashima, M.; Yagyu, K.; Oh-Hashi, K.; Suzuki, H.; Ono, K.; Furuta, K.; Sawada, M. Novel oxindole-curcumin hybrid compound for antioxidative stress and neuroprotection. ACS Chem. Neurosci. 2020, 11, 76–85. [Google Scholar] [CrossRef]
- Xu, H.; Perreau, V.M.; Dent, K.A.; Bush, A.I.; Finkelstein, D.I.; Adlard, P.A. Iron regulates apolipoprotein e expression and secretion in neurons and astrocytes. J. Alzheimers Dis. 2016, 51, 471–487. [Google Scholar] [CrossRef]
- Lee, K.J.; Oh, Y.C.; Cho, W.K.; Ma, J.Y. Antioxidant and anti-inflammatory activity determination of one hundred kinds of pure chemical compounds using offline and online screening hplc assay. Evid. Based Complement. Altern. Med. Ecam 2015, 2015, 165457. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Character |
|---|---|
| BDNF | Selective TrkB agonist |
| 7, 8-dihydroxyflavone (7,8-DHF, a flavonoid) | Non-selective TrkB agonist |
| Phenelzine sulphate (antidepressant; monoamine oxidase inhibitor) | Non-selective TrkB activator |
| ANA-12 (a synthetic agent) | Selective, non-competitive TrkB antagonist |
| K252a (an alkaloidal compound isolated from Nocardiopsis bacteria) | Tyrosine kinase inhibitor |
| GNF-5837 (a synthetic oxindole compound) | Tyrosine kinase inhibitor |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jakaria, M.; Belaidi, A.A.; Southon, A.; Dent, K.A.; Lane, D.J.R.; Bush, A.I.; Ayton, S. Receptor-Independent Anti-Ferroptotic Activity of TrkB Modulators. Int. J. Mol. Sci. 2022, 23, 16205. https://doi.org/10.3390/ijms232416205
Jakaria M, Belaidi AA, Southon A, Dent KA, Lane DJR, Bush AI, Ayton S. Receptor-Independent Anti-Ferroptotic Activity of TrkB Modulators. International Journal of Molecular Sciences. 2022; 23(24):16205. https://doi.org/10.3390/ijms232416205
Chicago/Turabian StyleJakaria, Md., Abdel A. Belaidi, Adam Southon, Krista A. Dent, Darius J. R. Lane, Ashley I. Bush, and Scott Ayton. 2022. "Receptor-Independent Anti-Ferroptotic Activity of TrkB Modulators" International Journal of Molecular Sciences 23, no. 24: 16205. https://doi.org/10.3390/ijms232416205
APA StyleJakaria, M., Belaidi, A. A., Southon, A., Dent, K. A., Lane, D. J. R., Bush, A. I., & Ayton, S. (2022). Receptor-Independent Anti-Ferroptotic Activity of TrkB Modulators. International Journal of Molecular Sciences, 23(24), 16205. https://doi.org/10.3390/ijms232416205