

Chronic Intermittent Hypoxia during Sleep Causes Browning of Interscapular Adipose Tissue Accompanied by Local Insulin Resistance in Mice


 , and
, and
Abstract
1. Introduction
2. Results
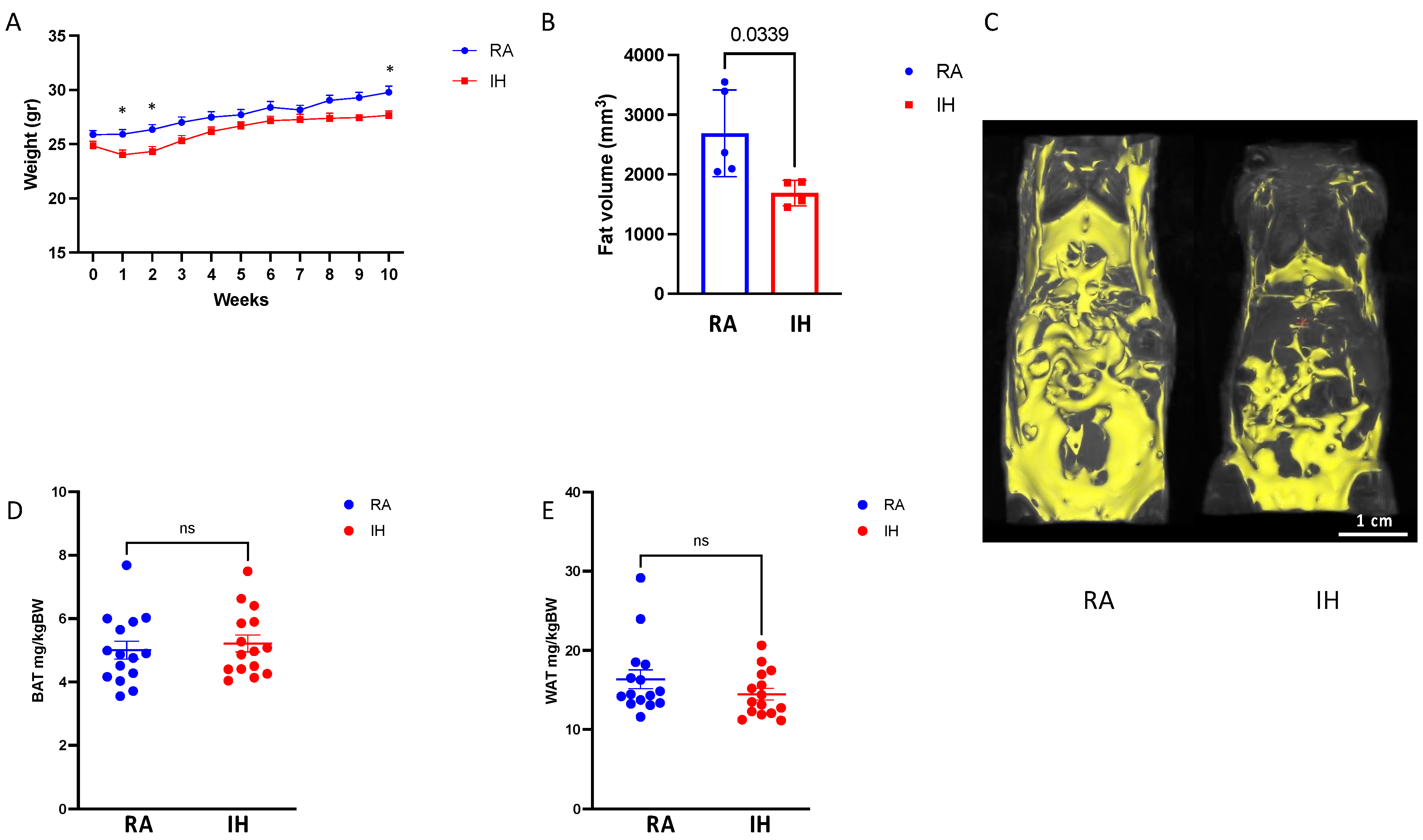
2.1. Body Composition and Food Consumption
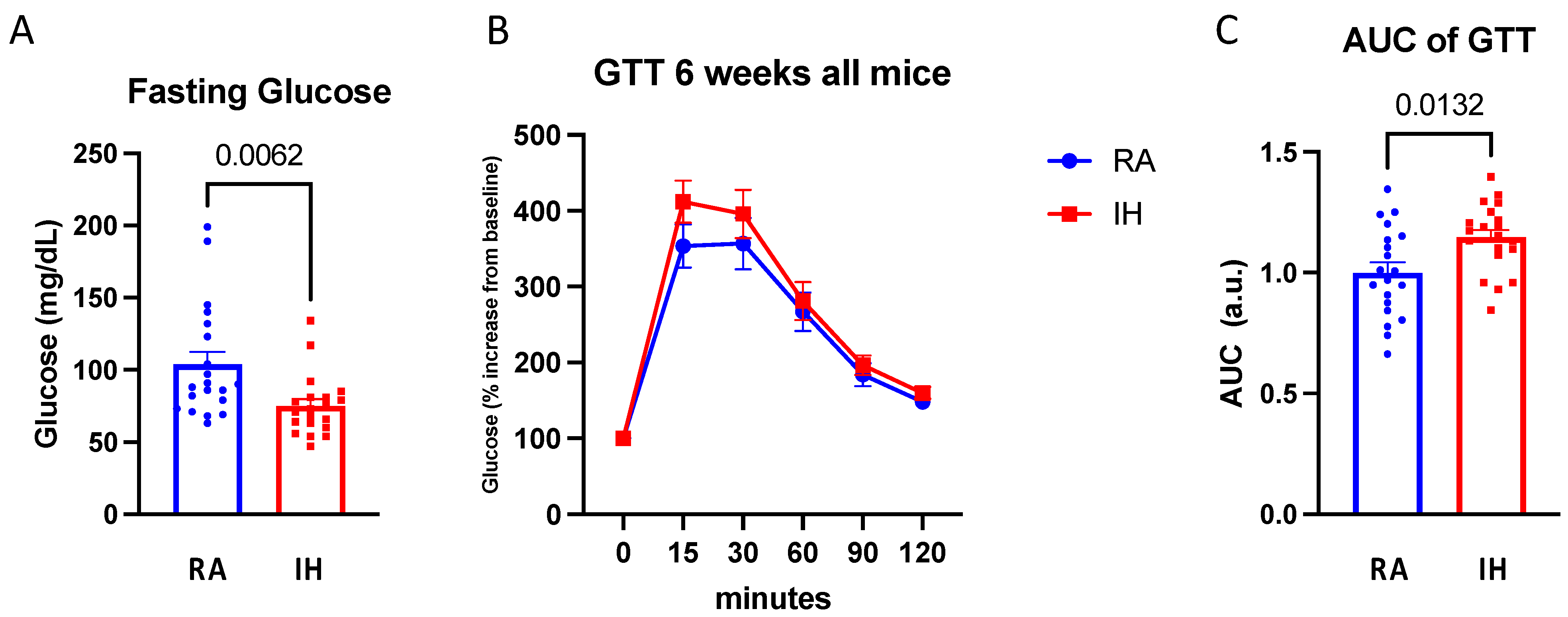
2.2. Glucose Tolerance Testing
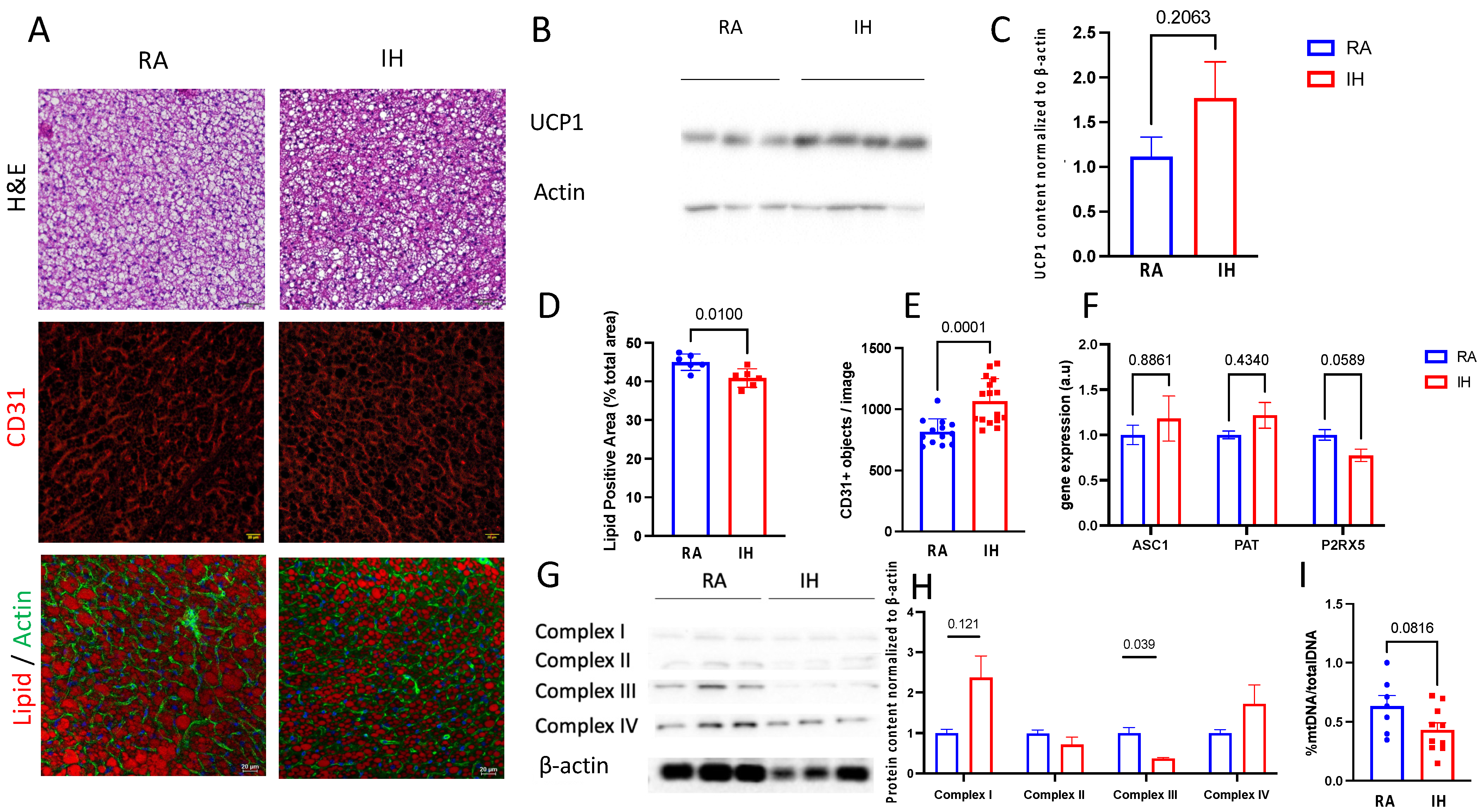
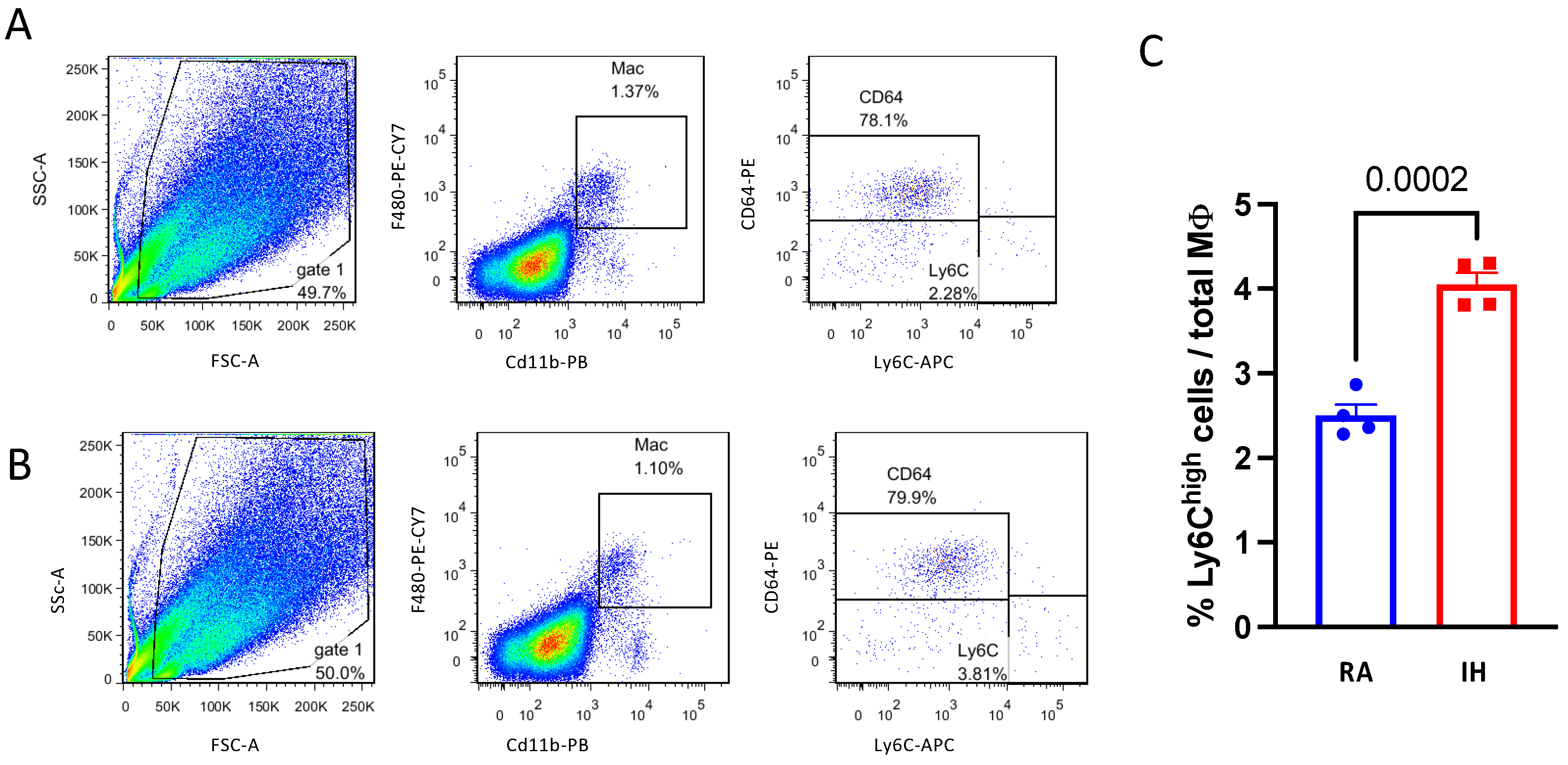
2.3. Brown Adipose Tissue Composition
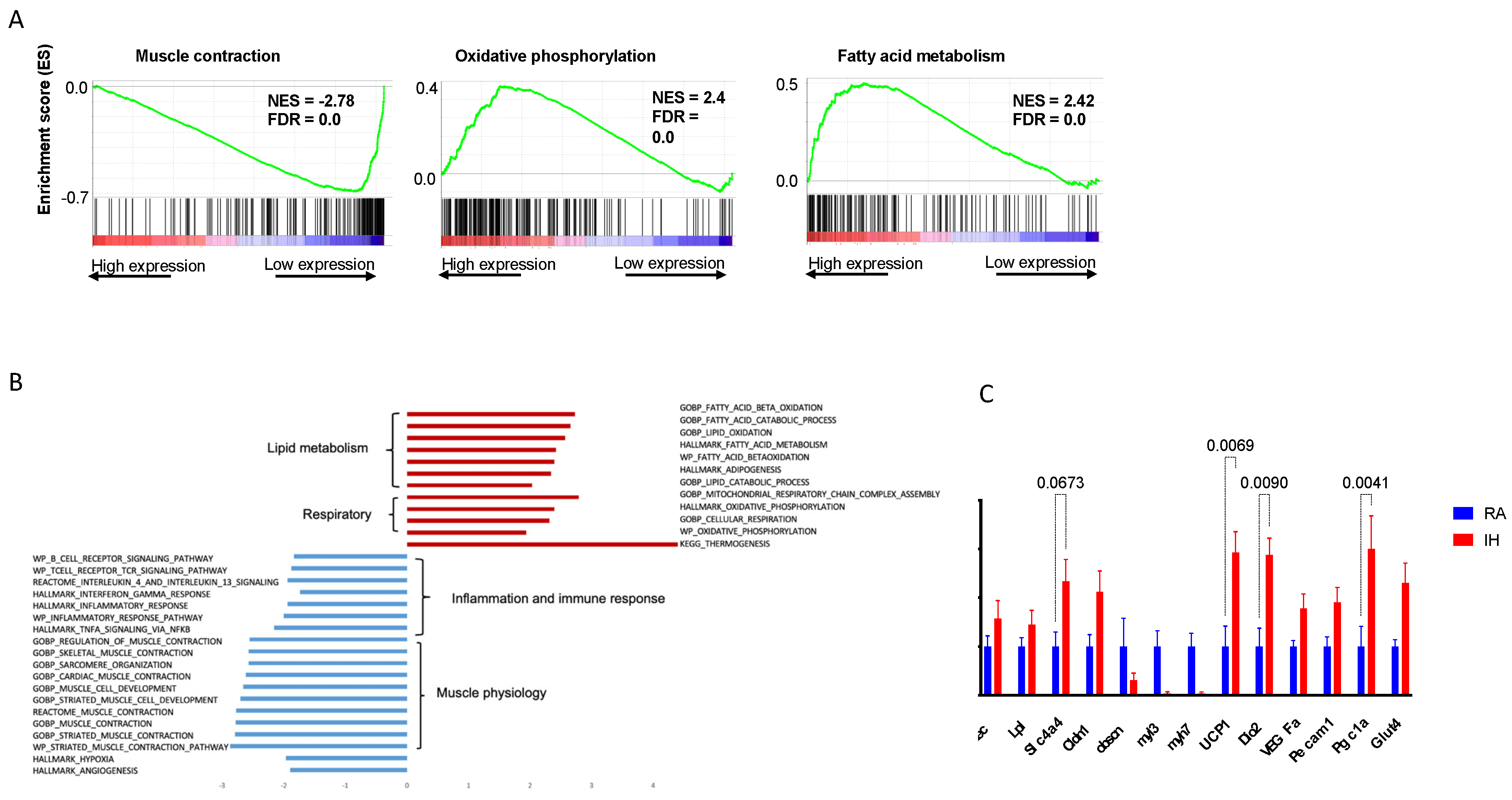
2.4. Brown Adipose Tissue Transcriptional Profile
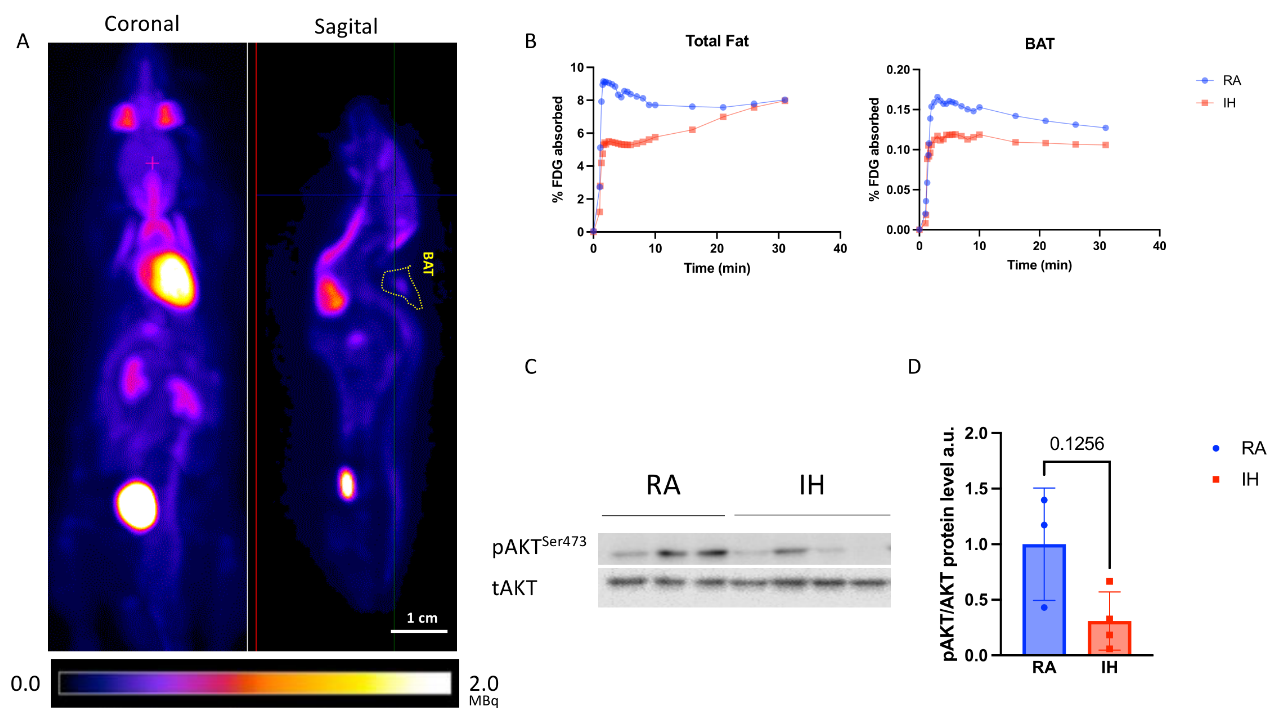
2.5. Brown Adipose Tissue Glucose Metabolism
3. Discussion
Limitations
4. Methods and Materials
4.1. Animals and Hypoxic Exposures
4.2. In Vivo Micro-PET–MRI Scanning
4.3. PET/MRI Image Processing
4.4. Glucose and Insulin Tolerance Tests
4.5. Immunohistochemistry
4.6. Western Blot Analysis
4.7. Mitochondrial DNA Quantification
4.8. RNA-Seq and Analysis
4.9. Real-Time PCR
4.10. Isolation of Stromal Vascular Fraction (SVF) and Flow Cytometry Analysis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Senaratna, C.V.; Perret, J.L.; Lodge, C.J.; Lowe, A.J.; Campbell, B.E.; Matheson, M.C.; Hamilton, G.S.; Dharmage, S.C. Prevalence of Obstructive Sleep Apnea in the General Population: A Systematic Review. Sleep Med. Rev. 2017, 34, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Goldbart, A.D.; Krishna, J.; Li, R.C.; Serpero, L.D.; Gozal, D. Inflammatory Mediators in Exhaled Breath Condensate of Children with Obstructive Sleep Apnea Syndrome. Chest 2006, 130, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Nachalon, Y.; Lowenthal, N.; Greenberg-Dotan, S.; Goldbart, A.D. Inflammation and Growth in Young Children with Obstructive Sleep Apnea Syndrome before and after Adenotonsillectomy. Mediators Inflamm. 2014, 2014, 146893. [Google Scholar] [CrossRef]
- Jun, J.C.; Drager, L.F.; Najjar, S.S.; Gottlieb, S.S.; Brown, C.D.; Smith, P.L.; Schwartz, A.R.; Polotsky, V.Y. Effects of Sleep Apnea on Nocturnal Free Fatty Acids in Subjects with Heart Failure. Sleep 2011, 34, 1207–1213. [Google Scholar] [CrossRef] [PubMed]
- Jun, J.C.; Chopra, S.; Schwartz, A.R. Sleep Apnoea. Eur. Respir. Rev. Off. J. Eur. Respir. Soc. 2016, 25, 12–18. [Google Scholar] [CrossRef]
- Gileles-Hillel, A.; Almendros, I.; Khalyfa, A.; Zhang, S.X.; Wang, Y.; Gozal, D. Early Intermittent Hypoxia Induces Proatherogenic Changes in Aortic Wall Macrophages in a Murine Model of Obstructive Sleep Apnea. Am. J. Respir. Crit. Care Med. 2014, 190, 958–961. [Google Scholar] [CrossRef]
- Kheirandish-Gozal, L.; Gileles-Hillel, A.; Alonso-Álvarez, M.L.; Peris, E.; Bhattacharjee, R.; Terán-Santos, J.; Duran-Cantolla, J.; Gozal, D. Effects of Adenotonsillectomy on Plasma Inflammatory Biomarkers in Obese Children with Obstructive Sleep Apnea: A Community-Based Study. Int. J. Obes. 2015, 39, 1094–1100. [Google Scholar] [CrossRef]
- Gileles-Hillel, A.; Alonso-Álvarez, M.L.; Kheirandish-Gozal, L.; Peris, E.; Cordero-Guevara, J.A.; Terán-Santos, J.; Martinez, M.G.; Jurado-Luque, M.J.; Corral-Peñafiel, J.; Duran-Cantolla, J.; et al. Inflammatory Markers and Obstructive Sleep Apnea in Obese Children: The NANOS Study. Mediators Inflamm. 2014, 2014, 605280. [Google Scholar] [CrossRef]
- Tauman, R.; O’Brien, L.M.; Gozal, D. Hypoxemia and Obesity Modulate Plasma C-Reactive Protein and Interleukin-6 Levels in Sleep-Disordered Breathing. Sleep Breath. Schlaf Atm. 2007, 11, 77–84. [Google Scholar] [CrossRef]
- Gileles-Hillel, A.; Kheirandish-Gozal, L.; Gozal, D. Biological Plausibility Linking Sleep Apnoea and Metabolic Dysfunction. Nat. Rev. Endocrinol. 2016, 12, 290–298. [Google Scholar] [CrossRef]
- Hakim, F.; Gozal, D.; Kheirandish-Gozal, L. Sympathetic and Catecholaminergic Alterations in Sleep Apnea with Particular Emphasis on Children. Front. Neurol. 2012, 3, 7. [Google Scholar] [CrossRef] [PubMed]
- Berger, S.; Polotsky, V.Y. Leptin and Leptin Resistance in the Pathogenesis of Obstructive Sleep Apnea: A Possible Link to Oxidative Stress and Cardiovascular Complications. Oxid. Med. Cell Longev. 2018, 2018, 5137947. [Google Scholar] [CrossRef] [PubMed]
- Pasarica, M.; Rood, J.; Ravussin, E.; Schwarz, J.-M.; Smith, S.R.; Redman, L.M. Reduced Oxygenation in Human Obese Adipose Tissue Is Associated with Impaired Insulin Suppression of Lipolysis. J. Clin. Endocrinol. Metab. 2010, 95, 4052–4055. [Google Scholar] [CrossRef] [PubMed]
- Pasarica, M.; Sereda, O.R.; Redman, L.M.; Albarado, D.C.; Hymel, D.T.; Roan, L.E.; Rood, J.C.; Burk, D.H.; Smith, S.R. Reduced Adipose Tissue Oxygenation in Human Obesity: Evidence for Rarefaction, Macrophage Chemotaxis, and Inflammation without an Angiogenic Response. Diabetes 2009, 58, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Ye, J. Emerging Role of Adipose Tissue Hypoxia in Obesity and Insulin Resistance. Int. J. Obes. 2005 2009, 33, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Trayhurn, P. Hypoxia and Adipose Tissue Function and Dysfunction in Obesity. Physiol. Rev. 2013, 93, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Goossens, G.H.; Bizzarri, A.; Venteclef, N.; Essers, Y.; Cleutjens, J.P.; Konings, E.; Jocken, J.W.E.; Cajlakovic, M.; Ribitsch, V.; Clément, K.; et al. Increased Adipose Tissue Oxygen Tension in Obese Compared with Lean Men Is Accompanied by Insulin Resistance, Impaired Adipose Tissue Capillarization, and Inflammation. Circulation 2011, 124, 67–76. [Google Scholar] [CrossRef]
- Lawler, H.M.; Underkofler, C.M.; Kern, P.A.; Erickson, C.; Bredbeck, B.; Rasouli, N. Adipose Tissue Hypoxia, Inflammation, and Fibrosis in Obese Insulin-Sensitive and Obese Insulin-Resistant Subjects. J. Clin. Endocrinol. Metab. 2016, 101, 1422–1428. [Google Scholar] [CrossRef]
- Trayhurn, P.; Alomar, S.Y. Oxygen Deprivation and the Cellular Response to Hypoxia in Adipocytes—Perspectives on White and Brown Adipose Tissues in Obesity. Front. Endocrinol. 2015, 6, 19. [Google Scholar] [CrossRef]
- Cao, Y. Angiogenesis and Vascular Functions in Modulation of Obesity, Adipose Metabolism, and Insulin Sensitivity. Cell Metab. 2013, 18, 478–489. [Google Scholar] [CrossRef]
- Wander, K.; Su, M.; Mattison, P.M.; Sum, C.-Y.; Witt, C.C.; Shenk, M.K.; Blumenfield, T.; Li, H.; Mattison, S.M. High-Altitude Adaptations Mitigate Risk for Hypertension and Diabetes-Associated Anemia. Am. J. Phys. Anthropol. 2020, 172, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Burtscher, M.; Millet, G.P.; Klimont, J.; Burtscher, J. Differences in the Prevalence of Physical Activity and Cardiovascular Risk Factors between People Living at Low (<1001 m) Compared to Moderate (1001–2000 m) Altitude. AIMS Public Health 2021, 8, 624–635. [Google Scholar] [CrossRef] [PubMed]
- Brito, J.; Siques, P.; López, R.; Romero, R.; León-Velarde, F.; Flores, K.; Lüneburg, N.; Hannemann, J.; Böger, R.H. Long-Term Intermittent Work at High Altitude: Right Heart Functional and Morphological Status and Associated Cardiometabolic Factors. Front. Physiol. 2018, 9, 248. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.M.; Thomas, A.; Crinion, S.J.; Kent, B.D.; Tambuwala, M.M.; Fabre, A.; Pepin, J.-L.; Roche, H.M.; Arnaud, C.; Ryan, S. Intermittent Hypoxia in Obstructive Sleep Apnoea Mediates Insulin Resistance through Adipose Tissue Inflammation. Eur. Respir. J. 2017, 49, 1601731. [Google Scholar] [CrossRef]
- Cypess, A.M.; Lehman, S.; Williams, G.; Tal, I.; Rodman, D.; Goldfine, A.B.; Kuo, F.C.; Palmer, E.L.; Tseng, Y.-H.; Doria, A.; et al. Identification and Importance of Brown Adipose Tissue in Adult Humans. N. Engl. J. Med. 2009, 360, 1509–1517. [Google Scholar] [CrossRef]
- Stanford, K.I.; Middelbeek, R.J.W.; Townsend, K.L.; An, D.; Nygaard, E.B.; Hitchcox, K.M.; Markan, K.R.; Nakano, K.; Hirshman, M.F.; Tseng, Y.-H.; et al. Brown Adipose Tissue Regulates Glucose Homeostasis and Insulin Sensitivity. J. Clin. Investig. 2013, 123, 215–223. [Google Scholar] [CrossRef]
- Becher, T.; Palanisamy, S.; Kramer, D.J.; Eljalby, M.; Marx, S.J.; Wibmer, A.G.; Butler, S.D.; Jiang, C.S.; Vaughan, R.; Schöder, H.; et al. Brown Adipose Tissue Is Associated with Cardiometabolic Health. Nat. Med. 2021, 27, 58–65. [Google Scholar] [CrossRef]
- Shimizu, I.; Aprahamian, T.; Kikuchi, R.; Shimizu, A.; Papanicolaou, K.N.; MacLauchlan, S.; Maruyama, S.; Walsh, K. Vascular Rarefaction Mediates Whitening of Brown Fat in Obesity. J. Clin. Investig. 2014, 124, 2099–2112. [Google Scholar] [CrossRef]
- Xue, Y.; Petrovic, N.; Cao, R.; Larsson, O.; Lim, S.; Chen, S.; Feldmann, H.M.; Liang, Z.; Zhu, Z.; Nedergaard, J.; et al. Hypoxia-Independent Angiogenesis in Adipose Tissues during Cold Acclimation. Cell Metab. 2009, 9, 99–109. [Google Scholar] [CrossRef]
- Gileles-Hillel, A.; Almendros, I.; Khalyfa, A.; Nigdelioglu, R.; Qiao, Z.; Hamanaka, R.B.; Mutlu, G.M.; Akbarpour, M.; Gozal, D. Prolonged Exposures to Intermittent Hypoxia Promote Visceral White Adipose Tissue Inflammation in a Murine Model of Severe Sleep Apnea: Effect of Normoxic Recovery. Sleep 2017, 40, zsw074. [Google Scholar] [CrossRef]
- Khalyfa, A.; Qiao, Z.; Gileles-Hillel, A.; Khalyfa, A.A.; Akbarpour, M.; Popko, B.; Gozal, D. Activation of the Integrated Stress Response and Metabolic Dysfunction in a Murine Model of Sleep Apnea. Am. J. Respir. Cell Mol. Biol. 2017, 57, 477–486. [Google Scholar] [CrossRef]
- Boroumand, P.; Prescott, D.; Mukherjee, T.; Bilan, P.J.; Wong, M.; Shen, J.; Tattoli, I.; Zhou, Y.; Li, A.; Sivasubramaniyam, T.; et al. Bone Marrow Adipocytes Drive the Development of Tissue Invasive Ly6Chigh Monocytes during Obesity. Elife 2022, 11, e65553. [Google Scholar] [CrossRef]
- Wang, C.; Liu, W.; Nie, Y.; Qaher, M.; Horton, H.E.; Yue, F.; Asakura, A.; Kuang, S. Loss of MyoD Promotes Fate Transdifferentiation of Myoblasts Into Brown Adipocytes. EBioMedicine 2017, 16, 212–223. [Google Scholar] [CrossRef]
- Gozal, D.; Gileles-Hillel, A.; Cortese, R.; Li, Y.; Almendros, I.; Qiao, Z.; Khalyfa, A.A.; Andrade, J.; Khalyfa, A. Visceral White Adipose Tissue after Chronic Intermittent and Sustained Hypoxia in Mice. Am. J. Respir. Cell Mol. Biol. 2017, 56, 477–487. [Google Scholar] [CrossRef]
- Drager, L.F.; Li, J.; Shin, M.-K.; Reinke, C.; Aggarwal, N.R.; Jun, J.C.; Bevans-Fonti, S.; Sztalryd, C.; O’Byrne, S.M.; Kroupa, O.; et al. Intermittent Hypoxia Inhibits Clearance of Triglyceride-Rich Lipoproteins and Inactivates Adipose Lipoprotein Lipase in a Mouse Model of Sleep Apnoea. Eur. Heart J. 2012, 33, 783–790. [Google Scholar] [CrossRef]
- Zhen, X.; Moya, E.A.; Gautane, M.; Zhao, H.; Lawrence, E.S.; Gu, W.; Barnes, L.A.; Yuan, J.X.-J.; Jain, P.P.; Xiong, M.; et al. Combined Intermittent and Sustained Hypoxia Is a Novel and Deleterious Cardio-Metabolic Phenotype. Sleep 2022, 45, zsab290. [Google Scholar] [CrossRef]
- Kajimura, S.; Saito, M. A New Era in Brown Adipose Tissue Biology: Molecular Control of Brown Fat Development and Energy Homeostasis. Annu. Rev. Physiol. 2014, 76, 225–249. [Google Scholar] [CrossRef]
- Bartelt, A.; Heeren, J. Adipose Tissue Browning and Metabolic Health. Nat. Rev. Endocrinol. 2014, 10, 24–36. [Google Scholar] [CrossRef]
- Leitner, B.P.; Huang, S.; Brychta, R.J.; Duckworth, C.J.; Baskin, A.S.; McGehee, S.; Tal, I.; Dieckmann, W.; Gupta, G.; Kolodny, G.M.; et al. Mapping of Human Brown Adipose Tissue in Lean and Obese Young Men. Proc. Natl. Acad. Sci. USA 2017, 114, 8649–8654. [Google Scholar] [CrossRef]
- Virtanen, K.A.; Lidell, M.E.; Orava, J.; Heglind, M.; Westergren, R.; Niemi, T.; Taittonen, M.; Laine, J.; Savisto, N.-J.; Enerbäck, S.; et al. Functional Brown Adipose Tissue in Healthy Adults. N. Engl. J. Med. 2009, 360, 1518–1525. [Google Scholar] [CrossRef]
- Yoneshiro, T.; Aita, S.; Matsushita, M.; Kayahara, T.; Kameya, T.; Kawai, Y.; Iwanaga, T.; Saito, M. Recruited Brown Adipose Tissue as an Antiobesity Agent in Humans. J. Clin. Investig. 2013, 123, 3404–3408. [Google Scholar] [CrossRef]
- Semenza, G.L.; Prabhakar, N.R. The Role of Hypoxia-Inducible Factors in Carotid Body (Patho) Physiology. J. Physiol. 2018, 596, 2977–2983. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Ueda, S.; Kobayashi, T.; Nishiyama, A.; Fujisawa, Y.; Sugaya, T.; Shiota, S.; Takahashi, K.; Gohda, T.; Horikoshi, S.; et al. Chronic Intermittent Hypoxia-Mediated Renal Sympathetic Nerve Activation in Hypertension and Cardiovascular Disease. Sci. Rep. 2018, 8, 17926. [Google Scholar] [CrossRef] [PubMed]
- Nanduri, J.; Peng, Y.-J.; Wang, N.; Prabhakar, N.R. Neural Activation of Molecular Circuitry in Intermittent Hypoxia. Curr. Opin. Physiol. 2019, 7, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Cetin-Atalay, R.; Meliton, A.Y.; Wu, D.; Woods, P.S.; Sun, K.A.; Peng, Y.-J.; Nanduri, J.; Su, X.; Fang, Y.; Hamanaka, R.B.; et al. Intermittent Hypoxia-Induced Activation of Endothelial Cells Is Mediated via Sympathetic Activation-Dependent Catecholamine Release. Front. Physiol. 2021, 12, 701995. [Google Scholar] [CrossRef]
- Almendros, I.; Wang, Y.; Gozal, D. The Polymorphic and Contradictory Aspects of Intermittent Hypoxia. Am. J. Physiol. Lung Cell Mol. Physiol. 2014, 307, L129–L140. [Google Scholar] [CrossRef]
- Yao, Q.; Shin, M.-K.; Jun, J.C.; Hernandez, K.L.; Aggarwal, N.R.; Mock, J.R.; Gay, J.; Drager, L.F.; Polotsky, V.Y. Effect of Chronic Intermittent Hypoxia on Triglyceride Uptake in Different Tissues. J. Lipid Res. 2013, 54, 1058–1065. [Google Scholar] [CrossRef]
- Martinez, D.; Fiori, C.Z.; Baronio, D.; Carissimi, A.; Kaminski, R.S.R.; Kim, L.J.; Rosa, D.P.; Bos, Â. Brown Adipose Tissue: Is It Affected by Intermittent Hypoxia? Lipids Health Dis. 2010, 9, 121. [Google Scholar] [CrossRef]
- Kumar, G.K.; Rai, V.; Sharma, S.D.; Ramakrishnan, D.P.; Peng, Y.-J.; Souvannakitti, D.; Prabhakar, N.R. Chronic Intermittent Hypoxia Induces Hypoxia-Evoked Catecholamine Efflux in Adult Rat Adrenal Medulla via Oxidative Stress: Induction of Hypoxic Sensitivity in Adult Rat Adrenal Medulla. J. Physiol. 2006, 575, 229–239. [Google Scholar] [CrossRef]
- Shin, M.-K.; Yao, Q.; Jun, J.C.; Bevans-Fonti, S.; Yoo, D.-Y.; Han, W.; Mesarwi, O.; Richardson, R.; Fu, Y.-Y.; Pasricha, P.J.; et al. Carotid Body Denervation Prevents Fasting Hyperglycemia during Chronic Intermittent Hypoxia. J. Appl. Physiol. 2014, 117, 765–776. [Google Scholar] [CrossRef]
- Kooijman, S.; van den Heuvel, J.K.; Rensen, P.C.N. Neuronal Control of Brown Fat Activity. Trends Endocrinol. Metab. 2015, 26, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Hurtado, E.; Lee, J.; Choi, J.; Wolfgang, M.J. Fatty Acid Oxidation Is Required for Active and Quiescent Brown Adipose Tissue Maintenance and Thermogenic Programing. Mol. Metab. 2018, 7, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zhou, J.H.; Zhang, H.; Canfran-Duque, A.; Singh, A.K.; Perry, R.J.; Shulman, G.I.; Fernandez-Hernando, C.; Min, W. Brown Adipose TRX2 Deficiency Activates MtDNA-NLRP3 to Impair Thermogenesis and Protect against Diet-Induced Insulin Resistance. J. Clin. Investig. 2022, 132, e148852. [Google Scholar] [CrossRef] [PubMed]
- Okamatsu-Ogura, Y.; Kuroda, M.; Tsutsumi, R.; Tsubota, A.; Saito, M.; Kimura, K.; Sakaue, H. UCP1-Dependent and UCP1-Independent Metabolic Changes Induced by Acute Cold Exposure in Brown Adipose Tissue of Mice. Metabolism. 2020, 113, 154396. [Google Scholar] [CrossRef]
- Hankir, M.K.; Kranz, M.; Keipert, S.; Weiner, J.; Andreasen, S.G.; Kern, M.; Patt, M.; Klöting, N.; Heiker, J.T.; Brust, P.; et al. Dissociation Between Brown Adipose Tissue 18F-FDG Uptake and Thermogenesis in Uncoupling Protein 1–Deficient Mice. J. Nucl. Med. 2017, 58, 1100–1103. [Google Scholar] [CrossRef]
- Timmons, J.A.; Wennmalm, K.; Larsson, O.; Walden, T.B.; Lassmann, T.; Petrovic, N.; Hamilton, D.L.; Gimeno, R.E.; Wahlestedt, C.; Baar, K.; et al. Myogenic Gene Expression Signature Establishes That Brown and White Adipocytes Originate from Distinct Cell Lineages. Proc. Natl. Acad. Sci. USA 2007, 104, 4401–4406. [Google Scholar] [CrossRef]
- Crewe, C.; An, Y.A.; Scherer, P.E. The Ominous Triad of Adipose Tissue Dysfunction: Inflammation, Fibrosis, and Impaired Angiogenesis. J. Clin. Investig. 2017, 127, 74–82. [Google Scholar] [CrossRef]
- Alcalá, M.; Calderon-Dominguez, M.; Bustos, E.; Ramos, P.; Casals, N.; Serra, D.; Viana, M.; Herrero, L. Increased Inflammation, Oxidative Stress and Mitochondrial Respiration in Brown Adipose Tissue from Obese Mice. Sci. Rep. 2017, 7, 16082. [Google Scholar] [CrossRef]
- Choi, E.W.; Lee, M.; Song, J.W.; Kim, K.; Lee, J.; Yang, J.; Lee, S.H.; Kim, I.Y.; Choi, J.-H.; Seong, J.K. Fas Mutation Reduces Obesity by Increasing IL-4 and IL-10 Expression and Promoting White Adipose Tissue Browning. Sci. Rep. 2020, 10, 12001. [Google Scholar] [CrossRef]
- Kotzbeck, P.; Giordano, A.; Mondini, E.; Murano, I.; Severi, I.; Venema, W.; Cecchini, M.P.; Kershaw, E.E.; Barbatelli, G.; Haemmerle, G.; et al. Brown Adipose Tissue Whitening Leads to Brown Adipocyte Death and Adipose Tissue Inflammation. J. Lipid Res. 2018, 59, 784–794. [Google Scholar] [CrossRef]
- Wu, C.; Cheng, W.; Sun, Y.; Dang, Y.; Gong, F.; Zhu, H.; Li, N.; Li, F.; Zhu, Z. Activating Brown Adipose Tissue for Weight Loss and Lowering of Blood Glucose Levels: A MicroPET Study Using Obese and Diabetic Model Mice. PLoS ONE 2014, 9, e113742. [Google Scholar] [CrossRef] [PubMed]
- Farré, R.; Gozal, D.; Almendros, I. Human Experimental Models: Seeking to Enhance Multiscale Research in Sleep Apnoea. Eur. Respir. J. 2021, 58, 2101169. [Google Scholar] [CrossRef] [PubMed]
- Farré, R.; Montserrat, J.M.; Gozal, D.; Almendros, I.; Navajas, D. Intermittent Hypoxia Severity in Animal Models of Sleep Apnea. Front. Physiol. 2018, 9, 1556. [Google Scholar] [CrossRef] [PubMed]
- Courteau, A.; McGrath, J.; Walker, P.M.; Pegg, R.; Martin, G.; Garipov, R.; Doughty, P.; Cochet, A.; Brunotte, F.; Vrigneaud, J.-M. Performance Evaluation and Compatibility Studies of a Compact Preclinical Scanner for Simultaneous PET/MR Imaging at 7 Tesla. IEEE Trans. Med. Imaging 2021, 40, 205–217. [Google Scholar] [CrossRef]
- Nir, T.; Melton, D.A.; Dor, Y. Recovery from Diabetes in Mice by Beta Cell Regeneration. J. Clin. Investig. 2007, 117, 2553–2561. [Google Scholar] [CrossRef]
- Malik, A.N.; Czajka, A.; Cunningham, P. Accurate Quantification of Mouse Mitochondrial DNA without Co-Amplification of Nuclear Mitochondrial Insertion Sequences. Mitochondrion 2016, 29, 59–64. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate Alignment of Transcriptomes in the Presence of Insertions, Deletions and Gene Fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq--a Python Framework to Work with High-Throughput Sequencing Data. Bioinforma. Oxf. Engl. 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene Set Enrichment Analysis: A Knowledge-Based Approach for Interpreting Genome-Wide Expression Profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed]
- Liberzon, A.; Subramanian, A.; Pinchback, R.; Thorvaldsdóttir, H.; Tamayo, P.; Mesirov, J.P. Molecular Signatures Database (MSigDB) 3.0. Bioinforma. Oxf. Engl. 2011, 27, 1739–1740. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.W.; Zamarron, B.F.; Muir, L.A.; Singer, K.; Porsche, C.E.; DelProposto, J.B.; Geletka, L.; Meyer, K.A.; O’Rourke, R.W.; Lumeng, C.N. Adipose Tissue Dendritic Cells Are Independent Contributors to Obesity-Induced Inflammation and Insulin Resistance. J. Immunol. 2016, 197, 3650–3661. [Google Scholar] [CrossRef] [PubMed]
- Röszer, T. Understanding the Biology of Self-Renewing Macrophages. Cells 2018, 7, 103. [Google Scholar] [CrossRef] [PubMed]
- Poroyko, V.A.; Carreras, A.; Khalyfa, A.; Khalyfa, A.A.; Leone, V.; Peris, E.; Almendros, I.; Gileles-Hillel, A.; Qiao, Z.; Hubert, N.; et al. Chronic Sleep Disruption Alters Gut Microbiota, Induces Systemic and Adipose Tissue Inflammation and Insulin Resistance in Mice. Sci. Rep. 2016, 6, 35405. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Forward | Reverse |
|---|---|---|
| GLUT4 | GTCGGGTTTCCAGCAGATC | AAACTGAAGGGAGCCAAGC |
| PGC1a | AAATCATATCCAACCAGTACA | CATCTGTCAGTGCATCAAAT |
| Myh7 | TGCTGTTTCCTTACTTGCTA | GGATTCTCAAACGTGTCTAGT |
| Cldn1 | TACAGTGCAAAGTCTTCGACT | GACACAAAGATTGCGATCAG |
| Slc4a4 | GATGAAGCTGTCCTGGACA | GACCCCAATGTAGATCGTG |
| Lpl | GTTTGGCTCCAGAGTTTGAC | CAAGTGTCCTCAGCTGTGTCT |
| Cidec | CTCACAGCTTGGAGGACCT | CAGGGCTTGGAAGTATTCTT |
| Myl3 | TGATGCCTCCAAGATTAAG | CGTATGTGATCTTCATCTCG |
| YWHAZ | AGAAGATCGAGACGGAGCT | GCCAAGTAACGGTAGTAGTCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dahan, T.; Nassar, S.; Yajuk, O.; Steinberg, E.; Benny, O.; Abudi, N.; Plaschkes, I.; Benyamini, H.; Gozal, D.; Abramovitch, R.; et al. Chronic Intermittent Hypoxia during Sleep Causes Browning of Interscapular Adipose Tissue Accompanied by Local Insulin Resistance in Mice. Int. J. Mol. Sci. 2022, 23, 15462. https://doi.org/10.3390/ijms232415462
Dahan T, Nassar S, Yajuk O, Steinberg E, Benny O, Abudi N, Plaschkes I, Benyamini H, Gozal D, Abramovitch R, et al. Chronic Intermittent Hypoxia during Sleep Causes Browning of Interscapular Adipose Tissue Accompanied by Local Insulin Resistance in Mice. International Journal of Molecular Sciences. 2022; 23(24):15462. https://doi.org/10.3390/ijms232415462
Chicago/Turabian StyleDahan, Tehila, Shahd Nassar, Olga Yajuk, Eliana Steinberg, Ofra Benny, Nathalie Abudi, Inbar Plaschkes, Hadar Benyamini, David Gozal, Rinat Abramovitch, and et al. 2022. "Chronic Intermittent Hypoxia during Sleep Causes Browning of Interscapular Adipose Tissue Accompanied by Local Insulin Resistance in Mice" International Journal of Molecular Sciences 23, no. 24: 15462. https://doi.org/10.3390/ijms232415462
APA StyleDahan, T., Nassar, S., Yajuk, O., Steinberg, E., Benny, O., Abudi, N., Plaschkes, I., Benyamini, H., Gozal, D., Abramovitch, R., & Gileles-Hillel, A. (2022). Chronic Intermittent Hypoxia during Sleep Causes Browning of Interscapular Adipose Tissue Accompanied by Local Insulin Resistance in Mice. International Journal of Molecular Sciences, 23(24), 15462. https://doi.org/10.3390/ijms232415462