
Biologically Active Peptides from Venoms: Applications in Antibiotic Resistance, Cancer, and Beyond


Abstract
1. Introduction—Drug Discovery in Venoms
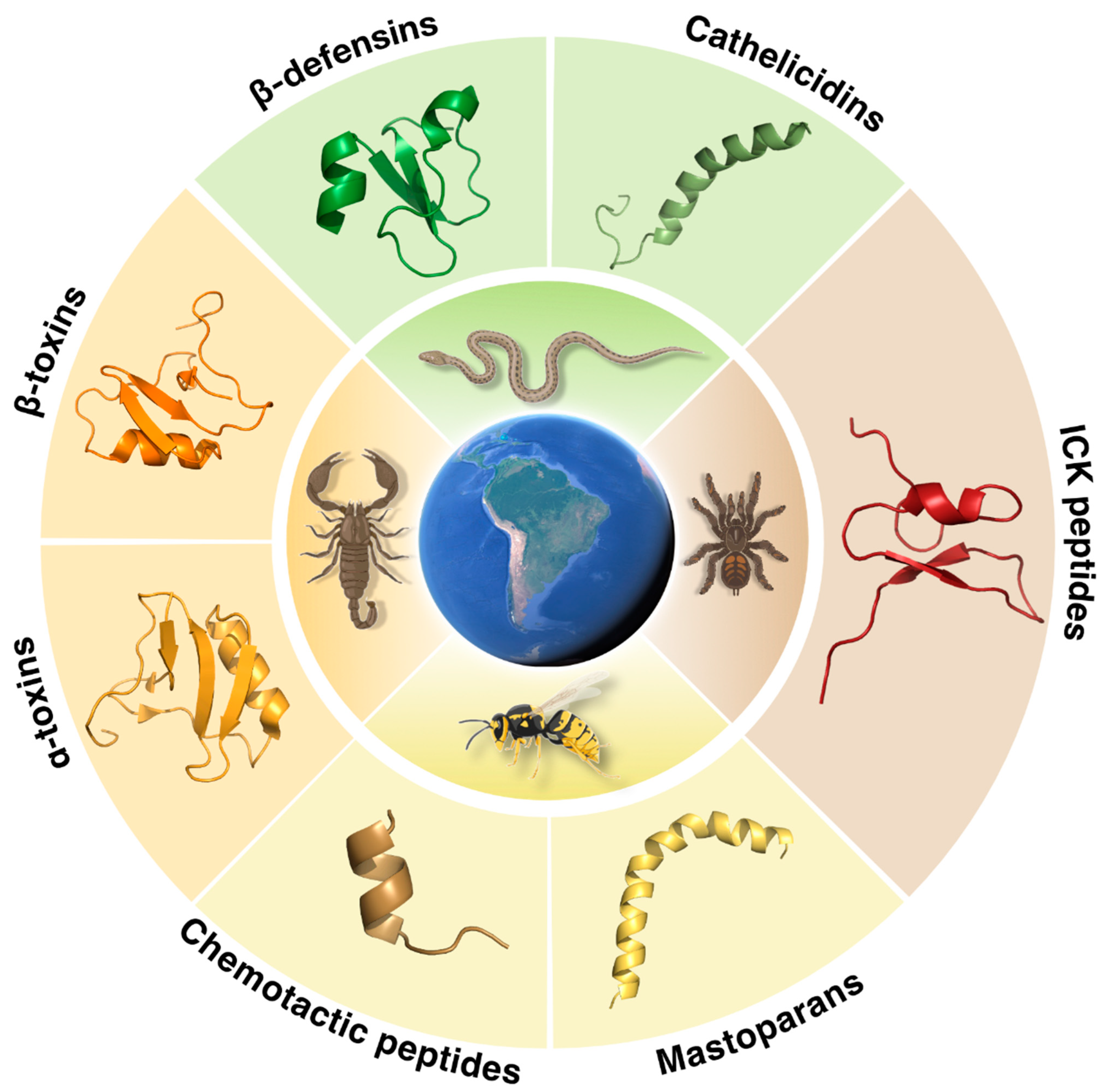
2. Venom-Derived Peptides
2.1. Snake Venoms
2.2. Spider Venoms
2.3. Scorpion Venoms
2.4. Wasp Venoms
2.5. Venom-Derived Peptides in the Clinic
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lewis, R.J.; Garcia, M.L. Therapeutic Potential of Venom Peptides. Nat. Rev. Drug Discov. 2003, 2, 790–802. [Google Scholar] [CrossRef] [PubMed]
- Pennington, M.W.; Czerwinski, A.; Norton, R.S. Peptide Therapeutics from Venom: Current Status and Potential. Bioorg. Med. Chem. 2018, 26, 2738–2758. [Google Scholar] [CrossRef] [PubMed]
- El-Aziz, T.M.A.; Soares, A.G.; Stockand, J.D. Snake Venoms in Drug Discovery: Valuable Therapeutic Tools for Life Saving. Toxins 2019, 11, 564. [Google Scholar] [CrossRef] [PubMed]
- King, G.F. Venoms as a Platform for Human Drugs: Translating Toxins into Therapeutics. Expert Opin. Biol. Ther. 2011, 11, 1469–1484. [Google Scholar] [CrossRef] [PubMed]
- Prashanth, J.R.; Hasaballah, N.; Vetter, I. Pharmacological Screening Technologies for Venom Peptide Discovery. Neuropharmacology 2017, 127, 4–19. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.D.T.; de la Fuente-Nunez, C. Toward Computer-Made Artificial Antibiotics. Curr. Opin. Microbiol. 2019, 51, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Porto, W.F.; Irazazabal, L.; Alves, E.S.F.; Ribeiro, S.M.; Matos, C.O.; Pires, Á.S.; Fensterseifer, I.C.M.; Miranda, V.J.; Haney, E.F.; Humblot, V.; et al. In Silico Optimization of a Guava Antimicrobial Peptide Enables Combinatorial Exploration for Peptide Design. Nat. Commun. 2018, 9, 1490. [Google Scholar] [CrossRef]
- Pane, K.; Cafaro, V.; Avitabile, A.; Torres, M.D.T.; Vollaro, A.; De Gregorio, E.; Catania, M.R.; Di Maro, A.; Bosso, A.; Gallo, G.; et al. Identification of Novel Cryptic Multifunctional Antimicrobial Peptides from the Human Stomach Enabled by a Computational-Experimental Platform. ACS Synth. Biol. 2018, 7, 2105–2115. [Google Scholar] [CrossRef]
- Torres, M.D.T.; Sothiselvam, S.; Lu, T.K.; de la Fuente-Nunez, C. Peptide Design Principles for Antimicrobial Applications. J. Mol. Biol. 2019, 431, 3547–3567. [Google Scholar] [CrossRef]
- Torres, M.D.T.; Cao, J.; Franco, O.L.; Lu, T.K.; de la Fuente-Nunez, C. Synthetic Biology and Computer-Based Frameworks for Antimicrobial Peptide Discovery. ACS Nano 2021, 15, 2143–2164. [Google Scholar] [CrossRef] [PubMed]
- Palmer, N.; Maasch, J.R.M.A.; Torres, M.D.T.; de la Fuente-Nunez, C. Molecular Dynamics for Antimicrobial Peptide Discovery. Infect. Immun. 2021, 89, e00703-20. [Google Scholar] [CrossRef]
- Der Torossian Torres, M.; De La Fuente-Nunez, C. Reprogramming Biological Peptides to Combat Infectious Diseases. Chem. Commun. 2019, 55, 15020–15032. [Google Scholar] [CrossRef] [PubMed]
- Silveira, G.G.O.S.; Torres, M.D.T.; Ribeiro, C.F.A.; Meneguetti, B.T.; Carvalho, C.M.E.; de la Fuente-Nunez, C.; Franco, O.L.; Cardoso, M.H. Antibiofilm Peptides: Relevant Preclinical Animal Infection Models and Translational Potential. ACS Pharmacol. Transl. Sci. 2021, 4, 55–73. [Google Scholar] [CrossRef]
- Liberio, M.; Joanitti, G.; Fontes, W.; Castro, M.S. Anticancer Peptides and Proteins: A Panoramic View. Protein Pept. Lett. 2012, 20, 380–391. [Google Scholar] [CrossRef]
- Pfalzgraff, A.; Brandenburg, K.; Weindl, G. Antimicrobial Peptides and Their Therapeutic Potential for Bacterial Skin Infections and Wounds. Front. Pharmacol. 2018, 9, 281. [Google Scholar] [CrossRef] [PubMed]
- de la Fuente-Nunez, C.; Torres, M.D.; Mojica, F.J.; Lu, T.K. Next-Generation Precision Antimicrobials: Towards Personalized Treatment of Infectious Diseases. Curr. Opin. Microbiol. 2017, 37, 95–102. [Google Scholar] [CrossRef] [PubMed]
- European Commission. A European One Health Action Plan against Antimicrobial Resistance; European Commission: Brussels, Belgium, 2017.
- World Health Organization. Global Cancer Facts & Figures, 4th ed.; American Cancer Society: Atlanta, GA, USA, 2018. [Google Scholar]
- Akef, H.M. Anticancer and Antimicrobial Activities of Scorpion Venoms and Their Peptides. Toxin Rev. 2019, 38, 41–53. [Google Scholar] [CrossRef]
- Torres, M.D.T.; Silva, A.F.; Andrade, G.P.; Pedron, C.N.; Cerchiaro, G.; Ribeiro, A.O.; Oliveira, V.X.; de la Fuente-Nunez, C. The Wasp Venom Antimicrobial Peptide Polybia-CP and Its Synthetic Derivatives Display Antiplasmodial and Anticancer Properties. Bioeng. Transl. Med. 2020, 5, e10167. [Google Scholar] [CrossRef]
- Koehbach, J.; Craik, D.J. The Vast Structural Diversity of Antimicrobial Peptides. Trends Pharmacol. Sci. 2019, 40, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Perumal Samy, R.; Stiles, B.G.; Franco, O.L.; Sethi, G.; Lim, L.H.K. Animal Venoms as Antimicrobial Agents. Biochem. Pharmacol. 2017, 134, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite Envenoming. Nat. Rev. Dis. Prim. 2017, 3, 17063. [Google Scholar] [CrossRef]
- Ferraz, C.R.; Arrahman, A.; Xie, C.; Casewell, N.R.; Lewis, R.J.; Kool, J.; Cardoso, F.C. Multifunctional Toxins in Snake Venoms and Therapeutic Implications: From Pain to Hemorrhage and Necrosis. Front. Ecol. Evol. 2019, 7, 218. [Google Scholar] [CrossRef]
- Kerkis, I.; Hayashi, M.A.F.; Prieto Da Silva, A.R.B.; Pereira, A.; De Sá Júnior, P.L.; Zaharenko, A.J.; Rádis-Baptista, G.; Kerkis, A.; Yamane, T. State of the Art in the Studies on Crotamine, a Cell Penetrating Peptide from South American Rattlesnake. Biomed. Res. Int. 2014, 2014, 675985. [Google Scholar] [CrossRef] [PubMed]
- de Lima, D.C.; Alvarez Abreu, P.; de Freitas, C.C.; Santos, D.O.; Borges, R.O.; Dos Santos, T.C.; Mendes Cabral, L.; Rodrigues, C.R.; Castro, H.C. Snake Venom: Any Clue for Antibiotics and CAM? Evid. Based Complement. Alternat. Med. 2005, 2, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Peinado, C.; Defaus, S.; Andreu, D. Hitchhiking with Nature: Snake Venom Peptides to Fight Cancer and Superbugs. Toxins 2020, 12, 255. [Google Scholar] [CrossRef]
- Mercer, D.K.; Torres, M.D.T.; Duay, S.S.; Lovie, E.; Simpson, L.; von Köckritz-Blickwede, M.; de la Fuente-Nunez, C.; O’Neil, D.A.; Angeles-Boza, A.M. Antimicrobial Susceptibility Testing of Antimicrobial Peptides to Better Predict Efficacy. Front. Cell. Infect. Microbiol. 2020, 10, 326. [Google Scholar] [CrossRef] [PubMed]
- Lehrer, R.I.; Bevins, C.L.; Ganz, T. Defensins and Other Antimicrobial Peptides and Proteins. Mucosal Immunol. 2005, 95–110. [Google Scholar] [CrossRef]
- Ganz, T. Defensins: Antimicrobial Peptides of Innate Immunity. Nat. Rev. Immunol. 2003, 3, 710–720. [Google Scholar] [CrossRef]
- Meade, K.G.; O’Farrelly, C. β-Defensins: Farming the Microbiome for Homeostasis and Health. Front. Immunol. 2019, 9, 3072. [Google Scholar] [CrossRef]
- Kerkis, I.; Silva, F.D.S.; Pereira, A.; Kerkis, A.; Rádis-Baptista, G. Biological Versatility of Crotamine a Cationic Peptide from the Venom of a South American Rattlesnake. Expert Opin. Investig. Drugs 2010, 19, 1515–1525. [Google Scholar] [CrossRef] [PubMed]
- Yamane, E.S.; Bizerra, F.C.; Oliveira, E.B.; Moreira, J.T.; Rajabi, M.; Nunes, G.L.C.; de Souza, A.O.; da Silva, I.D.C.G.; Yamane, T.; Karpel, R.L.; et al. Unraveling the Antifungal Activity of a South American Rattlesnake Toxin Crotamine. Biochimie 2013, 95, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.; Kerkis, A.; Hayashi, M.A.F.; Pereira, A.S.P.; Silva, F.S.; Oliveira, E.B.; Prieto da Silva, A.R.B.; Yamane, T.; Rádis-Baptista, G.; Kerkis, I. Crotamine Toxicity and Efficacy in Mouse Models of Melanoma. Expert Opin. Investig. Drugs 2011, 20, 1189–1200. [Google Scholar] [CrossRef] [PubMed]
- Falcao, C.B.; De La Torre, B.G.; Pérez-Peinado, C.; Barron, A.E.; Andreu, D.; Rádis-Baptista, G. Vipericidins: A Novel Family of Cathelicidin-Related Peptides from the Venom Gland of South American Pit Vipers. Amino Acids 2014, 46, 2561–2571. [Google Scholar] [CrossRef] [PubMed]
- Diniz, M.R.V.; Paiva, A.L.B.; Guerra-Duarte, C.; Nishiyama, M.Y., Jr.; Mudadu, M.A.; de Oliveira, U.; Borges, M.H.; Yates, J.R.; Junqueira-de-Azevedo, I.d.L. An Overview of Phoneutria Nigriventer Spider Venom Using Combined Transcriptomic and Proteomic Approaches. PLoS ONE 2018, 13, e0200628. [Google Scholar] [CrossRef] [PubMed]
- Peigneur, S.; de Lima, M.E.; Tytgat, J. Phoneutria Nigriventer Venom: A Pharmacological Treasure. Toxicon 2018, 151, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Escoubas, P.; De Weille, J.R.; Lecoq, A.; Diochot, S.; Waldmann, R.; Champigny, G.; Moinier, D.; Ménez, A.; Lazdunski, M. Isolation of a Tarantula Toxin Specific for a Class of Proton-Gated Na+ Channels. J. Biol. Chem. 2000, 275, 25116–25121. [Google Scholar] [CrossRef] [PubMed]
- Bubien, J.K.; Ji, H.-L.; Gillespie, G.Y.; Fuller, C.M.; Markert, J.M.; Mapstone, T.B.; Benos, D.J. Cation Selectivity and Inhibition of Malignant Glioma Na+ Channels by Psalmotoxin 1. Am. J. Physiol. Cell Physiol. 2004, 287, C1282–C1291. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Segura-Ramírez, P.J.; Silva Júnior, P.I. Loxosceles Gaucho Spider Venom: An Untapped Source of Antimicrobial Agents. Toxins 2018, 10, 522. [Google Scholar] [CrossRef] [PubMed]
- Cologna, C.; Marcussi, S.; Giglio, J.; Soares, A.; Arantes, E. Tityus Serrulatus Scorpion Venom and Toxins: An Overview. Protein Pept. Lett. 2009, 16, 920–932. [Google Scholar] [CrossRef] [PubMed]
- Cologna, C.T.; Peigneur, S.; Rustiguel, J.K.; Nonato, M.C.; Tytgat, J.; Arantes, E.C. Investigation of the Relationship between the Structure and Function of Ts2, a Neurotoxin from Tityus Serrulatus Venom. FEBS J. 2012, 279, 1495–1504. [Google Scholar] [CrossRef] [PubMed]
- Zoccal, K.F.; da Silva Bitencourt, C.; Sorgi, C.A.; de Castro Figueiredo Bordon, K.; Sampaio, S.V.; Arantes, E.C.; Faccioli, L.H. Ts6 and Ts2 from Tityus Serrulatus Venom Induce Inflammation by Mechanisms Dependent on Lipid Mediators and Cytokine Production. Toxicon 2013, 61, 1–10. [Google Scholar] [CrossRef]
- Dang, B.; Kubota, T.; Mandal, K.; Correa, A.M.; Bezanilla, F.; Kent, S.B.H. Elucidation of the Covalent and Tertiary Structures of Biologically Active Ts3 Toxin. Angew. Chem. Int. Ed. Engl. 2016, 55, 8639–8642. [Google Scholar] [CrossRef]
- Arantes, E.C.; Riccioppo Neto, F.; Sampaio, S.V.; Vieira, C.A.; Giglio, J. Isolation and Characterization of TsTX-V, a New Neurotoxin from Tityus Serrulatus Scorpion Venom Which Delays the Inactivation of NA+ Channels. Biochim. Biophys. Acta (BBA) Gen. Subj. 1994, 1199, 69–75. [Google Scholar] [CrossRef]
- Silva, J.C.; Neto, L.M.; Neves, R.C.; Gonçalves, J.C.; Trentini, M.M.; Mucury-Filho, R.; Smidt, K.S.; Fensterseifer, I.C.; Silva, O.N.; Lima, L.D.; et al. Evaluation of the Antimicrobial Activity of the Mastoparan Polybia-MPII Isolated from Venom of the Social Wasp Pseudopolybia Vespiceps Testacea (Vespidae, Hymenoptera). Int. J. Antimicrob. Agents 2017, 49, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Souza, B.M.; Mendes, M.A.; Santos, L.D.; Marques, M.R.; César, L.M.M.; Almeida, R.N.A.; Pagnocca, F.C.; Konno, K.; Palma, M.S. Structural and Functional Characterization of Two Novel Peptide Toxins Isolated from the Venom of the Social Wasp Polybia Paulista. Peptides 2005, 26, 2157–2164. [Google Scholar] [CrossRef]
- Gao, W.; Xing, L.; Qu, P.; Tan, T.; Yang, N.; Li, D.; Chen, H.; Feng, X. Identification of a Novel Cathelicidin Antimicrobial Peptide from Ducks and Determination of Its Functional Activity and Antibacterial Mechanism. Sci. Rep. 2015, 5, 17260. [Google Scholar] [CrossRef]
- Zanetti, M.; Gennaro, R.; Romeo, D. Cathelicidins: A Novel Protein Family with a Common Proregion and a Variable C-Terminal Antimicrobial Domain. FEBS Lett. 1995, 374, 1–5. [Google Scholar] [CrossRef]
- De Barros, E.; Gonçalves, R.M.; Cardoso, M.H.; Santos, N.C.; Franco, O.L.; Cândido, E.S. Snake Venom Cathelicidins as Natural Antimicrobial Peptides. Front. Pharmacol. 2019, 10, 1415. [Google Scholar] [CrossRef]
- Zanetti, M. The Role of Cathelicidins in the Innate Host Defenses of Mammals. Curr. Issues Mol. Biol. 2005, 7, 179–196. [Google Scholar]
- Estrada-Gomez, S.; Cardoso, F.C.; Vargas-Muñoz, L.J.; Quintana-Castillo, J.C.; Arenas Gómez, C.M.; Pineda, S.S.; Saldarriaga-Cordoba, M.M. Venomic, Transcriptomic, and Bioactivity Analyses of Pamphobeteus Verdolaga Venom Reveal Complex Disulfide-Rich Peptides That Modulate Calcium Channels. Toxins 2019, 11, 496. [Google Scholar] [CrossRef] [PubMed]
- Escoubas, P.; Sollod, B.; King, G.F. Venom Landscapes: Mining the Complexity of Spider Venoms via a Combined CDNA and Mass Spectrometric Approach. Toxicon 2006, 47, 650–663. [Google Scholar] [CrossRef] [PubMed]
- Garcia, F.; Villegas, E.; Espino-Solis, G.P.; Rodriguez, A.; Paniagua-Solis, J.F.; Sandoval-Lopez, G.; Possani, L.D.; Corzo, G. Antimicrobial Peptides from Arachnid Venoms and Their Microbicidal Activity in the Presence of Commercial Antibiotics. J. Antibiot. 2013, 66, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Langenegger, N.; Nentwig, W.; Kuhn-Nentwig, L. Spider Venom: Components, Modes of Action, and Novel Strategies in Transcriptomic and Proteomic Analyses. Toxins 2019, 11, 611. [Google Scholar] [CrossRef]
- Saez, N.J.; Senff, S.; Jensen, J.E.; Er, S.Y.; Herzig, V.; Rash, L.D.; King, G.F. Spider-Venom Peptides as Therapeutics. Toxins 2010, 2, 2851–2871. [Google Scholar] [CrossRef] [PubMed]
- Lauria, P.S.S.; Villarreal, C.F.; Casais-e-Silva, L.L. Pain Modulatory Properties of Phoneutria Nigriventer Crude Venom and Derived Peptides: A Double-Edged Sword. Toxicon 2020, 185, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Gomez, M.V.; Kalapothakis, E.; Guatimosim, C.; Prado, M.A.M. Phoneutria Nigriventer Venom: A Cocktail of Toxins That Affect Ion Channels. Cell. Mol. Neurobiol. 2002, 22, 579–588. [Google Scholar] [CrossRef]
- Furtado, A.; Silva, A.; Silva-Júnior, A.; Fernandes-Pedrosa, M. Biology, Venom Composition, and Scorpionism Induced by Brazilian Scorpion Tityus Stigmurus (Thorell, 1876) (Scorpiones: Buthidae): A Mini-Review. Toxicon 2020, 185, 36–45. [Google Scholar] [CrossRef]
- Martin-Eauclaire, M.-F.; Bougis, P.E.; de Lima, M.E. Ts1 from the Brazilian Scorpion Tityus Serrulatus: A Half-Century of Studies on a Multifunctional Beta like-Toxin. Toxicon 2018, 152, 106–120. [Google Scholar] [CrossRef]
- Lee, S.H.; Baek, J.H.; Yoon, K.A. Differential Properties of Venom Peptides and Proteins in Solitary vs. Social Hunting Wasps. Toxins 2016, 8, 32. [Google Scholar] [CrossRef]
- Konno, K.; Kazuma, K.; Rangel, M.; Stolarz-de-Oliveira, J.; Fontana, R.; Kawano, M.; Fuchino, H.; Hide, I.; Yasuhara, T.; Nakata, Y. New Mastoparan Peptides in the Venom of the Solitary Eumenine Wasp Eumenes Micado. Toxins 2019, 11, 155. [Google Scholar] [CrossRef]
- Freire, K.A.; Torres, M.D.T.; Lima, D.B.; Monteiro, M.L.; Bezerra de Menezes, R.R.P.P.; Martins, A.M.C.; Oliveira, V.X., Jr. Wasp Venom Peptide as a New Antichagasic Agent. Toxicon 2020, 181, 71–78. [Google Scholar] [CrossRef] [PubMed]
- de Souza, B.M.; da Silva, A.V.R.; Resende, V.M.F.; Arcuri, H.A.; dos Santos Cabrera, M.P.; Ruggiero Neto, J.; Palma, M.S. Characterization of Two Novel Polyfunctional Mastoparan Peptides from the Venom of the Social Wasp Polybia Paulista. Peptides 2009, 30, 1387–1395. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.D.T.; Pedron, C.N.; Higashikuni, Y.; Kramer, R.M.; Cardoso, M.H.; Oshiro, K.G.N.; Franco, O.L.; Silva Junior, P.I.; Silva, F.D.; Oliveira Junior, V.X.; et al. Structure-Function-Guided Exploration of the Antimicrobial Peptide Polybia-CP Identifies Activity Determinants and Generates Synthetic Therapeutic Candidates. Commun. Biol. 2018, 1, 221. [Google Scholar] [CrossRef]
- Arqué, X.; Torres, M.D.T.; Patiño, T.; Boaro, A.; Sánchez, S.; de la Fuente-Nunez, C. Autonomous Treatment of Bacterial Infections in Vivo Using Antimicrobial Micro- and Nanomotors. ACS Nano 2022, 16, 7547–7558. [Google Scholar] [CrossRef] [PubMed]
- de la Torre, B.G.; Albericio, F. Peptide Therapeutics 2.0. Molecules 2020, 25, 2293. [Google Scholar] [CrossRef] [PubMed]
- de la Torre, B.G.; Albericio, F. The Pharmaceutical Industry in 2019. An Analysis of FDA Drug Approvals from the Perspective of Molecules. Molecules 2020, 25, 745. [Google Scholar] [CrossRef] [PubMed]
- Al Musaimi, O.; Al Shaer, D.; Albericio, F.; de la Torre, B.G. 2020 FDA TIDES (Peptides and Oligonucleotides) Harvest. Pharmaceuticals 2021, 14, 145. [Google Scholar] [CrossRef] [PubMed]
- FDA. New Drugs at FDA: CDER’s New Molecular Entities and New Therapeutic Biological Products. Available online: https://www.fda.gov/drugs/development-approval-process-drugs/new-drugs-fda-cders-new-molecular-entities-and-new-therapeutic-biological-products (accessed on 1 October 2022).
- Wang, L.; Wang, N.; Zhang, W.; Cheng, X.; Yan, Z.; Shao, G.; Wang, X.; Wang, R.; Fu, C. Therapeutic Peptides: Current Applications and Future Directions. Signal Transduct. Target. Ther. 2022, 7, 48. [Google Scholar] [PubMed]
- Bordon, K.d.C.F.; Cologna, C.T.; Fornari-Baldo, E.C.; Pinheiro-Júnior, E.L.; Cerni, F.A.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cardoso, I.A.; et al. From Animal Poisons and Venoms to Medicines: Achievements, Challenges and Perspectives in Drug Discovery. Front. Pharmacol. 2020, 11, 1132. [Google Scholar]
- Ferreira, S.H. A Bradykinin-Potentiating Factor (BPF) Present in the Venom of Bothrops Jararaca. Br. J. Pharmacol. Chemother. 1965, 24, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Camargo, A.C.M.; Ianzer, D.; Guerreiro, J.R.; Serrano, S.M.T. Bradykinin-Potentiating Peptides: Beyond Captopril. Toxicon 2012, 59, 516–523. [Google Scholar] [CrossRef]
- Patchett, A.A. The Chemistry of Enalapril. Br. J. Clin. Pharmacol. 1984, 18, 201S–207S. [Google Scholar] [CrossRef] [PubMed]
- Biollaz, J.; Burnier, M.; Turini, G.A.; Brunner, D.B.; Porchet, M.; Gomez, H.J.; Jones, K.H.; Ferber, F.; Abrams, W.B.; Gavras, H.; et al. Three New Long-Acting Converting-Enzyme Inhibitors: Relationship between Plasma Converting-Enzyme Activity and Response to Angiotensin I. Clin. Pharmacol. Ther. 1981, 29, 665–670. [Google Scholar] [CrossRef]
- Coulter-Parkhill, A.; McClean, S.; Gault, V.A.; Irwin, N. Therapeutic Potential of Peptides Derived from Animal Venoms: Current Views and Emerging Drugs for Diabetes. Clin. Med. Insights Endocrinol. Diabetes 2021, 14. [Google Scholar] [CrossRef]
- Stocker, K.; Barlow, G.H. The Coagulant Enzyme from Bothrops Atrox Venom (Batroxobin). In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1976; Volume 45, pp. 214–223. ISBN 0076-6879. [Google Scholar]
- Funk, C.; Gmür, J.; Herold, R.; Straub, P.W. Reptilase®-R—A New Reagent in Blood Coagulation. Br. J. Haematol. 1971, 21, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Monge-Fuentes, V.; Arenas, C.; Galante, P.; Gonçalves, J.C.; Mortari, M.R.; Schwartz, E.F. Arthropod Toxins and Their Antinociceptive Properties: From Venoms to Painkillers. Pharmacol. Ther. 2018, 188, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Smallwood, T.B.; Clark, R.J. Advances in Venom Peptide Drug Discovery: Where Are We at and Where Are We Heading? Expert Opin. Drug Discov. 2021, 16, 1163–1173. [Google Scholar] [CrossRef]
- Flinspach, M.; Xu, Q.; Piekarz, A.D.; Fellows, R.; Hagan, R.; Gibbs, A.; Liu, Y.; Neff, R.A.; Freedman, J.; Eckert, W.A.; et al. Insensitivity to Pain Induced by a Potent Selective Closed-State Nav1.7 Inhibitor. Sci. Rep. 2017, 7, 39662. [Google Scholar] [CrossRef] [PubMed]
- Murray, J.K.; Long, J.; Zou, A.; Ligutti, J.; Andrews, K.L.; Poppe, L.; Biswas, K.; Moyer, B.D.; McDonough, S.I.; Miranda, L.P. Single Residue Substitutions That Confer Voltage-Gated Sodium Ion Channel Subtype Selectivity in the NaV1.7 Inhibitory Peptide GpTx-1. J. Med. Chem. 2016, 59, 2704–2717. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Hong, M.; Guo, X.; Wu, F.; Tian, C.; Wang, Y.; Xu, Z. Facile Synthesis of Macrocyclic Peptide Toxins of GpTx-1 and Its Analogue. Org. Chem. Front. 2018, 5, 2143–2147. [Google Scholar] [CrossRef]
- Cherki, R.S.; Kolb, E.; Langut, Y.; Tsveyer, L.; Bajayo, N.; Meir, A. Two Tarantula Venom Peptides as Potent and Differential NaV Channels Blockers. Toxicon 2014, 77, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Lamiyan, A.K.; Dalal, R.; Kumar, N.R. Venom Peptides in Association with Standard Drugs: A Novel Strategy for Combating Antibiotic Resistance – an Overview. J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, e20200001. [Google Scholar] [CrossRef] [PubMed]
- Agbale, C.M.; Sarfo, J.K.; Galyuon, I.K.; Juliano, S.A.; Silva, G.G.O.; Buccini, D.F.; Cardoso, M.H.; Torres, M.D.T.; Angeles-Boza, A.M.; de la Fuente-Nunez, C.; et al. Antimicrobial and Antibiofilm Activities of Helical Antimicrobial Peptide Sequences Incorporating Metal-Binding Motifs. Biochemistry 2019, 58, 3802–3812. [Google Scholar] [CrossRef] [PubMed]
- Nešuta, O.; Hexnerová, R.; Buděšínský, M.; Slaninová, J.; Bednárová, L.; Hadravová, R.; Straka, J.; Veverka, V.; Čeřovský, V. Antimicrobial Peptide from the Wild Bee Hylaeus Signatus Venom and Its Analogues: Structure–Activity Study and Synergistic Effect with Antibiotics. J. Nat. Prod. 2016, 79, 1073–1083. [Google Scholar] [CrossRef]
- Drugs@FDA: FDA-Approved Drugs. Available online: https://www.accessdata.fda.gov/scripts/cder/daf/index.cfm?event=BasicSearch.process (accessed on 1 October 2022).
- ClinicalTrials.Gov. The Efficacy and Safety of Batroxobin Combined with Anticoagulation in Cerebral Venous Sinus Thrombosis—Full Text View. Available online: https://clinicaltrials.gov/ct2/show/NCT04269954?term=batroxobin&draw=3&rank=1 (accessed on 1 October 2022).


{kind=link}
| Peptide | AA | Sequence | Peptide Family | Structure | Activity | Organism Producer | Reference |
|---|---|---|---|---|---|---|---|
| Snake Venom-Derived Peptides | |||||||
| Crotamine | 42 | YKQCHKKGGHCFPKEKICLPPSSDFGKMDCRWRWKCCKKGSG | β-defensin-like | β1αβ2β3 N-terminal 𝛼-helix, two stranded antiparallel β-sheets, and two β-turns. | Antifungal (MIC = 12.5–50.0 μg mL−1) Anticancer activity (5 μg mL−1) | Crotalus durissus terrificus | [25,34] |
| Crotalicidine | 34 | KRFKKFFKKVKKSVKKRLKKIFKKPMVIGVTIPF | Cathelicidin | α-helix at the N-terminal and random coil conformation at the C-terminal of the peptide. | Antibacterial (MIC < 10 μmol L−1), Anticancer (IC50 < 5 μmol L−1), Immunomodulatory activities | Crotalus durissus terrificus | [27,35] |
| Spider Venom-Derived Peptides | |||||||
| PnTx1 | 78 | AELTSCFPVGHECDGDASNCNCCGDDVYCGCGWGRWNCKCKVADQSYSYGICKDKVNCPNRHLWPAKVCKKCRRNCGG | ICK peptide | Disulfide bridge pattern | LD50 = 5.5 pmol g−1 (mice) Target: Nav channels, antagonist | Phoneutria nigriventer | [36,37] |
| PnTx2-1 | 53 | ATCAGQDKPCKETCDCCGERGECVCALSYEGKYRCICRQGNFLIAWHKLASCK | ICK peptide | n.e. | Lethal in mice model at 0.02 pmol g−1 | Phoneutria nigriventer | [36,37] |
| PnTx2-5 | 48 | ATCAGQDQTCKVTCDCCGERGECVCGGPCICRQGNFLIAWYKLASCKK | ICK peptide | n.e. | Lethal in mice model at 2.4 pmol g−1 | Phoneutria nigriventer | [36,37] |
| PnTx2-6 | 48 | ATCAGQDQPCKETCDCCGERGECVCGGPCICRQGYFWIAWYKLANCKK | ICK peptide | n.e. | Lethal in mice model at 7.5 pmol g−1 | Phoneutria nigriventer | [36,37] |
| PnTx2-9 | 32 | SFCIPFKPCKSDENCCKKFKCKTTGIVKLCRW | ICK peptide | n.e. | - | Phoneutria nigriventer | [36,37] |
| PnTx3-1 | 41 | AECAAVYERCGKGYKRCCEERPCKCNIVMDNCTCKKFISEL | ICK peptide | n.e. | Paralysis in mice at 0.07 pmol g−1 Target: agonist of K+ channels | Phoneutria nigriventer | [36,37] |
| PnTx3-2 | 46 | ACAGLYKKCGKGASPCCEDRPCKCDLAMGNCICKKKFIEFFGGGK | ICK peptide | n.e. | Antagonist of L-type CaV channels. Paralysis in mice 0.08 pmol g−1 | Phoneutria nigriventer | [36,37] |
| PnTx3-3 | 34 | GCANAYKSCNGPHTCCWGYNGYLLACICSGXNWK | ICK peptide | n.e. | Lethal to mice at 0.07 pmol g−1 Antagonist of L-, P/Q- and R-type Cav channels | Phoneutria nigriventer | [36,37] |
| PnTx3-4 | 77 | SCINVGDFCDGKKDDCQCCRDNAFCSCSVIFGYKTNCRCEVGTTATSYGICMAKHKCGRQTTCTKPCLSKRCKKNHG | ICK peptide | n.e. | Lethal to mice at 5 μg/mouse Target: antagonist of N-, P/Q- and R-type Cav channels | Phoneutria nigriventer | [36,37] |
| PnTx3-5 | 45 | GCIGRNESCKFDRHGCCWPWSCSCWNKEGQPESDVWCECSLKIGK | ICK peptide | n.e. | Paralysis in mice 0.07 pmol g−1 Target: L-type Cav channels | Phoneutria nigriventer | [36,37] |
| PnTx3-6 | 54 | ACIPRGEICTDDCECCGCDNQCYCPPGSSLGIFKCSCAHANKYFCNRKKEKCKK | ICK peptide | n.e. | Paralysis in mice 0.05 pmol g−1 Target: N-, P/Q- and L-type Cav channels | Phoneutria nigriventer | [36,37] |
| PnTx4-3 | 48 | CGDINAACKEDCDCCGYTTACDCYWSSSCKCREAAIVIYTAPKKKLTC | ICK peptide | n.e. | Non-toxic to mice (288.5 pmol g−1) LD50 = 192.3 pmol g−1 (house fly) | Phoneutria nigriventer | [36,37] |
| PnTx4 (5-5) | 47 | CADINGACKSDCDCCGDSVTCDCYWSDSCKCRESNFKIGMAIRKKF-C | ICK peptide | n.e. | Non-toxic to mice (290 pmol g−1) Target: NMDAR (antagonist), NaV channels (agonist) | Phoneutria nigriventer | [36,37] |
| PnTx4 (6-1) | 48 | CGDINAACKEDCDCCGYTTACDCYWSKSCKCREAAIVIYTAPKKKLTC | ICK peptide | n.e. | LD50=9.3 ng/house fly Non-toxic to mice (286.2 pmol g−1) ED50= 36.3 pmol g−1 (house fly) Target: agonist NaV channels | Phoneutria nigriventer | [36,37] |
| Psalmotoxin 1 (PcTX1) | 40 | EDCIPKWKGCVNRHGDCCEGLECWKRRRSFEVCVPKTPKT | ICK peptide | Three antiparallel ®-sheet structure with three disulfide bridges tightly folded into the “knottin” fold pattern | Antinociceptive effects IC50 = 36 pmol L−1 in glioma cells. | Psalmopoeuscambridgei | [38,39] |
| U1-SCRTX-Lg1a | 16 | VGTDFSGNDDISDVQK | Anionic antimicrobial peptide (AAMP) | Random coil conformation with a <-helix structure between the ISDV residues | Active against Gram-negative bacteria (MIC 1.5–4.6 μmol L−1) | Loxoscelesgaucho | [40] |
| Scorpion Venom-Derived Peptides | |||||||
| Ts1 | 61 | KEGYLMDHEGCKLSCFIRPSGYCGRECGIKKGSSGYCAWPACYCYGLPNWVKVWDRATNKC | ®-like neurotoxin | Three antiparallel β-strands and a α-helix bonded by disulfide bridges | Toxic against mammals and insects Intravenous LD50 = 76 ± 9 μg kg−1 Target: Na+ channels | Tityus serrulatus | [41] |
| Ts2 | 62 | KEGYAMDHEGCKFSCFIRPAGFCDGYCKTHLKASSGYCAWPACYCYGVPDHIKVWDYATNKC | ®-like neurotoxin | three β-strands and one α-helix, and is arranged in a triangular shape forming a cysteine-stabilized α-helix/ β-sheet (CSab) motif. three β-strands and one α-helix, and is arranged in a triangular shape forming a cysteine-stabilized α-helix/ β-sheet (CSab) motif. three β-strands and one α-helix, and is arranged in a triangular shape forming a cysteine-stabilized α-helix/ β-sheet (CSab) motif three β-strands and one α-helix, and is arranged in a triangular shape forming a cysteine-stabilized α-helix/β-sheet (CSab) motif Cysteine-stabilized α-helix/β-sheet (CSαβ) motif composed of three β-strands and one α-helix arranged in a triangular shape | Induction of inflammation and production of cytokines Inhibition of the rapid inactivation of some NaV channels | Tityus serrulatus | [41,42,43] |
| Ts3 | 62 | KKDGYPVEYDNCAYICWNYDNAYCDKLCKDKKADSGYCYWVHILCYCYGLPDSEPTKTNGKC | α-neurotoxin | α-helix and three-stranded antiparallel β-sheet | Inhibition of the inactivation of NaV channels Muscle relaxation | Tityus serrulatus | [41,44] |
| Ts5 | 64 | KKDGYPVEGDNCAFACFGYDNAYCDKLCKDKKADDGYCVWSPDCYCYGLPEHILKEPTKTSGRC | α-neurotoxin | Core composed of three β-strands and one α-helix | LD50 = 94 ± 7 μg kg−1 in mice Causes hypertension Target: Na+ channels | Tityus serrulatus | [41,45] |
| Ts6 | 40 | WCSTCLDLACGASRECYDPCFKAFGRAHGKCMNNKCRCYT | α-KTx (Potassium channel toxin) | α-helix and triple-stranded β-sheet stabilized by four disulfide bridges | Induction of inflammation and production of cytokines Blockage of KV channels | Tityus serrulatus | [41,43] |
| Ts7 | 37 | VFINAKCRGSPECLPKCKEAIGKAAGKCMNGKCKCYP | α-KTx (Potassium channel toxin) | n.e. | Blockage of K+ current Blockage of 86Rb efflux | Tityus serrulatus | [41] |
| Ts8 | 60 | KLVALIPNDQLRSILKAVVHKVAKTQFGCPAYEGYCNDHCNDIERKDGECHGFKCKCAKD | ®-KTx (Potassium channel toxin) | n.e. | Blockage of voltage-gated non-inactivating K+ channels from rat brain synaptosomes at IC50: 30 nmol L−1 | Tityus serrulatus | [41] |
| Ts9 | 35 | VVIGQRCYRSPDCYSACKKLVGKATGKCTNGRCDC | κ-KTx (Kappa potassium channel toxin) | Core composed of a short <-helix and a three-stranded antiparallel ®-sheet | Ligand for small-conductance apamin-sensitive calcium-activated potassium channel | Tityus serrulatus | [41] |
| Wasp Venom-Derived Peptides | |||||||
| Polybia-MPI | 14 | IDWKKLLDAAKQIL | Mastoparan | 71.43% α-helix | Antifungal activity (ED50 = 8–16 μmol L−1 Antimicrobial activity against Gram-positive and Gram-negative bacteria (MIC = 4–15 μg mL−1 | Polybiapaulista | [46,47] |
| Polybia-MPII | 14 | INWLKLGKMVIDAL | Mastoparan | α-helix | Antimicrobial activity against Gram-positive bacteria (MIC = 2–5 μmol L−1 Antifungal activity (ED50 = 111–12.9 μmol L−1 Hemolytic properties (ED50 of 5 × 10 -5 mol L−1 | Pseudopolybiavespiceps Testacea, Polybia paulista | [46] |
| Polybia-CP | 12 | ILGTILGLLKSL | Chemotactic peptide | 50% random coil 50% ambiguous conformations | Antimicrobial activity against Gram-positive bacteria (MIC = 15 μg mL−1 Mast cell degranulation (10−5 mol L−1) Low hemolytic activity | Polybiapaulista | [47] |
| Peptide | Producer Organism | Description | Target | Target Indication | Company (Brand) | Clinical Trial | Reference |
|---|---|---|---|---|---|---|---|
| Captopril | Bothropsjararaca | Synthetic peptide based on bradykinin-potentiating peptides (BPP) | Angiotensin-converting enzyme (ACE) | Hypertension, cardiac failure | Bristol-Myers Squibb. (Capoten®) | Completed | [2,72,73,74,89] |
| Enalapril | Bothropsjararaca | Synthetic peptide based on bradykinin-potentiating peptides (BPP) | Angiotensin-converting enzyme (ACE) | hypertension and cardiac failure | Merck (Vasotec®) | Completed | [72,75,76,89] |
| Batroxobin | Brazilian lancehead snake (Bothrops moojeni) | Peptide isolated from venom | Cleavage of the Aα chain of fibrinogen at the [Ala]16-[Gly]17 bond | Defibrinogenating effect Anticoagulation therapy (thrombotic diseases) Diagnosis of fibrinogen levels and blood coagulation capabilities | Tobishi Pharmaceutical (Batroxobin, Reptilase, Defibrase) DSM Nutritional Products Ltd/Branch Pentapharm (Defibrase) Hanlim (Botropase) Juggat Pharma (Botropase, Botroclot) Drugs.com, (Botroclot) Plateltex S.R.O. (Plateltex-Act®) Vivostat A/S (Vivostat System). | Phase IV (Combination with anticoagulation in cerebral venous sinus thrombosis, NCT04269954) Completed in different countries | [72,77,78,79,90] |
| Psalmotoxin 1 (PcTx-1) | Psalmopoeuscambridgei | Peptide isolated from venom | Inhibitor of AISCs | Pain treatment | - | Preclinical | [72,80,81] |
| JNJ63955918 | Thrixopelmapruriens | Synthetic peptide based on the natural peptide ProTX-II | NaV1.7 channels | Pain treatment | Janssen | Preclinical | [2,81,82] |
| GpTx-1 | Grammostila porter, Grammostila rosea, and Paraphysa scrofa | Peptide isolated from the venom | NaV1.7 channels | Pain treatment | Amgen | Preclinical | [2,81,83,84,85] |
| [Ala5, Phe6, Leu26, Arg28]GpTx-1 | Grammostila porter, Grammostila rosea, and Paraphysa scrofa | Synthetic peptide based on the natural peptide GpTx-1 | NaV1.7, NaV1.5 and NaV1.4 channels | Pain treatment | Amgen | Preclinical | [2,81,83,84,85] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ageitos, L.; Torres, M.D.T.; de la Fuente-Nunez, C. Biologically Active Peptides from Venoms: Applications in Antibiotic Resistance, Cancer, and Beyond. Int. J. Mol. Sci. 2022, 23, 15437. https://doi.org/10.3390/ijms232315437
Ageitos L, Torres MDT, de la Fuente-Nunez C. Biologically Active Peptides from Venoms: Applications in Antibiotic Resistance, Cancer, and Beyond. International Journal of Molecular Sciences. 2022; 23(23):15437. https://doi.org/10.3390/ijms232315437
Chicago/Turabian StyleAgeitos, Lucía, Marcelo D. T. Torres, and Cesar de la Fuente-Nunez. 2022. "Biologically Active Peptides from Venoms: Applications in Antibiotic Resistance, Cancer, and Beyond" International Journal of Molecular Sciences 23, no. 23: 15437. https://doi.org/10.3390/ijms232315437
APA StyleAgeitos, L., Torres, M. D. T., & de la Fuente-Nunez, C. (2022). Biologically Active Peptides from Venoms: Applications in Antibiotic Resistance, Cancer, and Beyond. International Journal of Molecular Sciences, 23(23), 15437. https://doi.org/10.3390/ijms232315437