


Heart Failure in Menopause: Treatment and New Approaches

 ,
,  ,
,
Abstract
1. Introduction
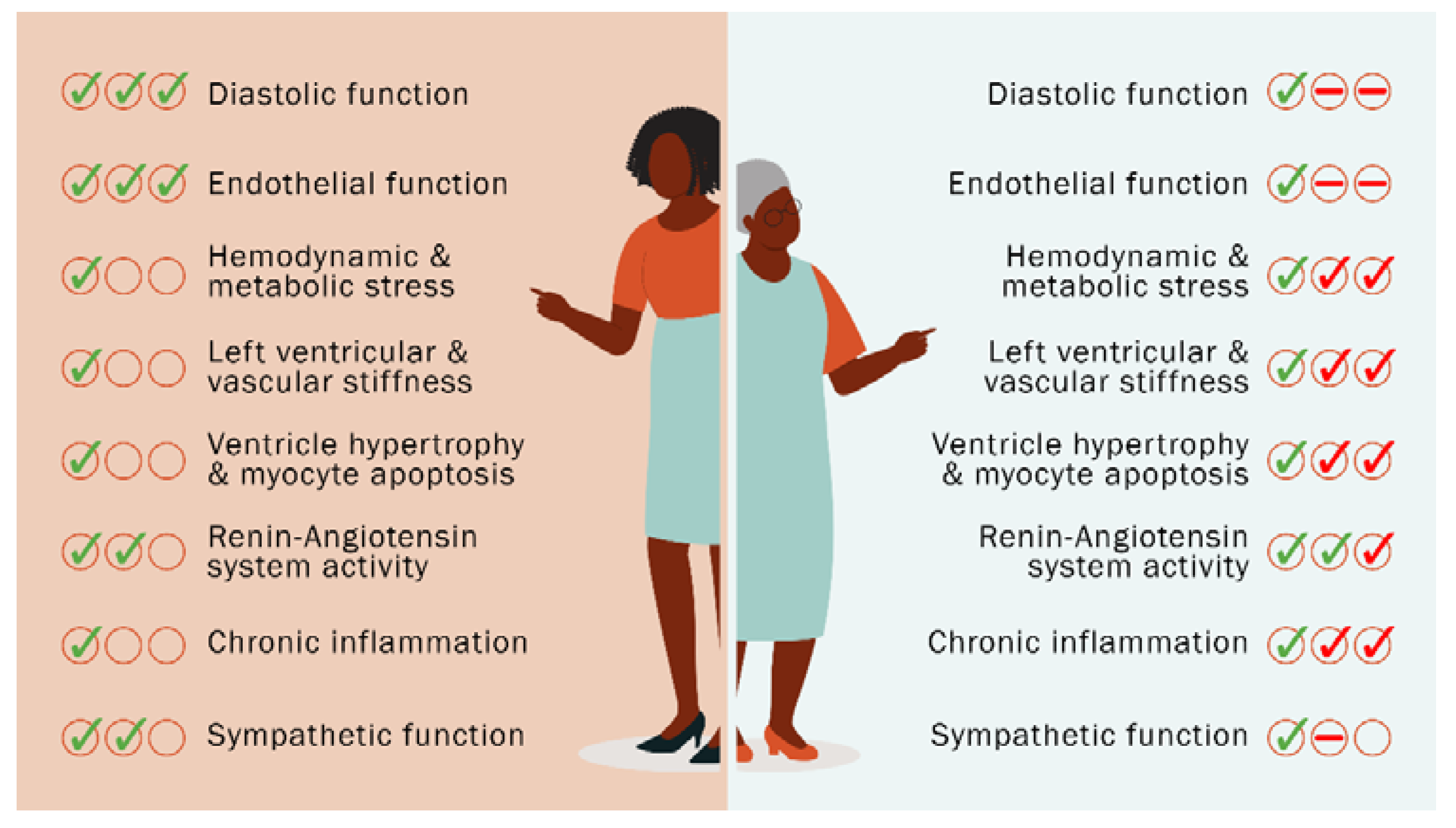
Heart Failure in Menopause and Therapeutic Targets
2. Mechanism of Menopause-Induced HFpEF and Targets for Treatment
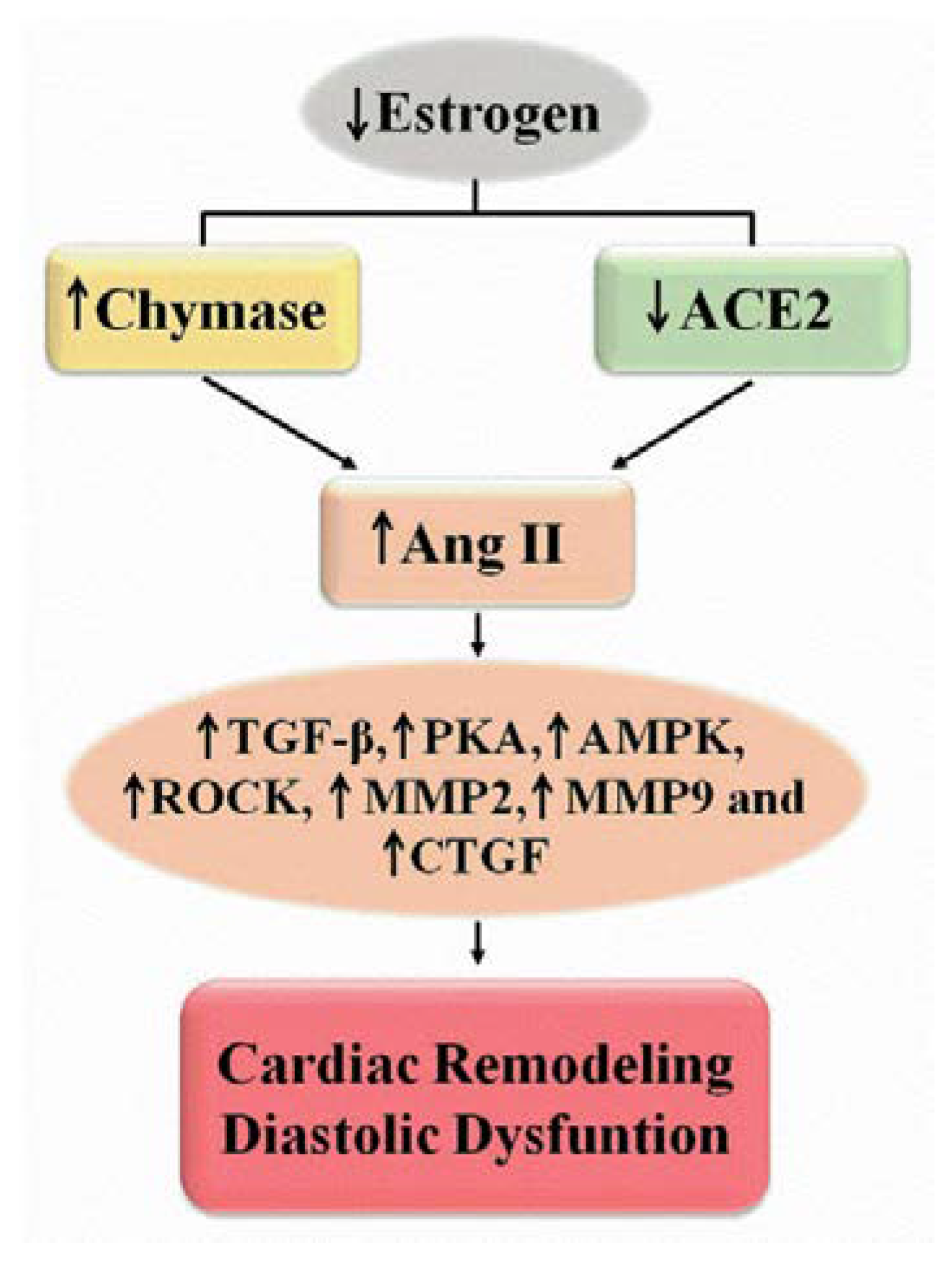
2.1. Renin-Angiotensin System
2.2. Inflammation, Metabolic Syndrome and Diabetes
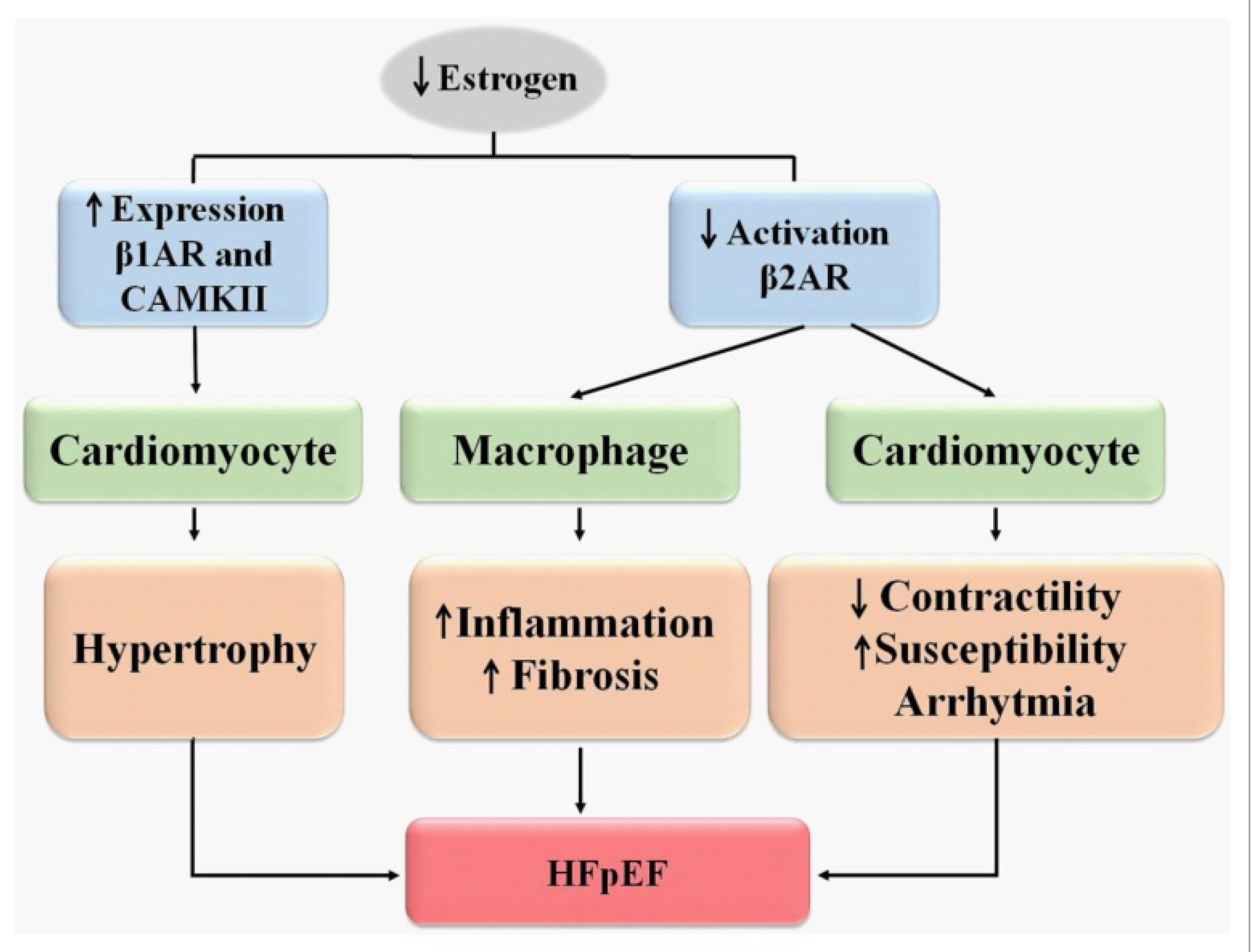
Sympathetic Nervous System
3. Treatment of Women with Menopausal-Induced HF
3.1. Pharmacological Therapy
3.2. Hormone Replacement Therapy
3.3. Phytoestrogen
3.4. Lifestyle Change and Healthy Diet
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hay, S.I.; Abajobir, A.A.; Abate, K.H.; Abbafati, C.; Abbas, K.M.; Abd-Allah, F.; Abdulkader, R.S.; Abdulle, A.M.; Abebo, T.A.; Abera, S.F.; et al. GBD 2016 DALYs and HALE Collaborators. Global, regional, and national disability-adjusted life-years (DALYs) for 333 diseases and injuries and healthy life expectancy (HALE) for 195 countries and territories, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet 2017, 390, 1260–1344. [Google Scholar] [CrossRef]
- Dagenais, G.R.; Leong, D.P.; Rangarajan, S.; Lanas, F.; Lopez-Jaramillo, P.; Gupta, R.; Diaz, R.; Avezum, A.; Oliveira, G.B.F.; Wielgosz, A.; et al. Variations in common diseases, hospital admissions, and deaths in middle-aged adults in 21 countries from five continents (PURE): A prospective cohort study. Lancet 2020, 395, 785–794. [Google Scholar] [CrossRef]
- Heidenreich, P.A.; Bozkurt, B.; Aguilar, D.; Allen, L.A.; Byun, J.J.; Colvin, M.M.; Deswal, A.; Drazner, M.H.; Dunlay, S.M.; Evers, L.R.; et al. 2022 AHA/ACC/HFSA Guideline for the Management of Heart Failure: A Report of the American College of Cardiology/American Heart Association Joint Committee on Clinical Practice Guidelines. J. Am. Coll. Cardiol. 2022, 79, e263–e421. [Google Scholar] [PubMed]
- Mozaffarian, D.; Benjamin, E.J.; Go, A.S.; Arnett, D.K.; Blaha, M.J.; Cushman, M.; Das, S.R.; De Ferranti, S.; Després, J.P.; Fullerton, H.J.; et al. Heart disease and stroke statistics-2016 update a report from the American Heart Association. Circulation 2016, 133, e38–e360. [Google Scholar] [PubMed]
- Benjamin, E.J.; Muntner, P.; Alonso, A.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Das, S.R.; et al. Heart disease and stroke statistics—2019 update: A report from the American heart association. Circulation 2019, 139, e56–e528. [Google Scholar] [CrossRef]
- Tibrewala, A.; Yancy, C.W. Heart Failure with Preserved Ejection Fraction in Women. Heart Fail. Clin. 2019, 15, 9–18. [Google Scholar] [CrossRef]
- Dunlay, S.M.; Roger, V.L.; Redfield, M.M. Epidemiology of heart failure with preserved ejection fraction. Nat. Rev. Cardiol. 2017, 14, 591–602. [Google Scholar] [CrossRef]
- Yoon, S.; Eom, G.H. Heart failure with preserved ejection fraction: Present status and future directions. Exp. Mol. Med. 2019, 51, 1–9. [Google Scholar] [CrossRef]
- Dinh, Q.N.; Vinh, A.; Arumugam, T.V.; Drummond, G.R.; Sobey, C.G. G protein-coupled estrogen receptor 1: A novel target to treat cardiovascular disease in a sex-specific manner? J. Cereb. Blood Flow Metab. 2021, 178, 3849–3863. [Google Scholar] [CrossRef]
- McDonagh, T.A.; Metra, M.; Adamo, M.; Gardner, R.S.; Baumbach, A.; Böhm, M.; Burri, H.; Butler, J.; Čelutkienė, J.; Chioncel, O.; et al. 2021 ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure. Eur. Heart J. 2021, 42, 3599–3726. [Google Scholar] [CrossRef]
- Koh, A.S.; Tay, W.T.; Teng, T.H.K.; Vedin, O.; Benson, L.; Dahlstrom, U.; Savarese, G.; Lam, C.S.; Lund, L.H. A comprehensive population-based characterization of heart failure with mid-range ejection fraction. Eur. J. Heart Fail. 2017, 19, 1624–1634. [Google Scholar] [CrossRef] [PubMed]
- Vedin, O.; Lam, C.S.; Koh, A.S.; Benson, L.; Teng, T.H.K.; Tay, W.T.; Braun, O.; Savarese, G.; Dahlström, U.; Lund, L.H. Significance of Ischemic Heart Disease in Patients With Heart Failure and Preserved, Midrange, and Reduced Ejection Fraction: A Nationwide Cohort Study. Circ. Heart Fail. 2017, 10. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, J.R.; Kapoor, R.; Ju, C.; Heidenreich, P.A.; Eapen, Z.J.; Hernandez, A.F.; Butler, J.; Yancy, C.W.; Fonarow, G.C. Precipitating Clinical Factors, Heart Failure Characterization, and Outcomes in Patients Hospitalized With Heart Failure With Reduced, Borderline, and Preserved Ejection Fraction. JACC Heart Fail. 2016, 4, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Borlaug, B.A. Evaluation and management of heart failure with preserved ejection fraction. Nat. Rev. Cardiol. 2020, 17, 559–573. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.; Mulvagh, S.L.; Merz, C.N.B.; Buring, J.E.; Manson, J.E. Cardiovascular Disease in Women. Circ. Res. 2016, 118, 1273–1293. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsova, T. Sex Differences in Epidemiology of Cardiac and Vascular Disease; Advances in Experimental Medicine and Biology; Springer: Cham, Switzerland, 2018; Volume 1065, pp. 61–70. [Google Scholar]
- Leening, M.J.G.; Ferket, B.; Steyerberg, E.W.; Kavousi, M.; Deckers, J.W.; Nieboer, D.; Heeringa, J.; Portegies, M.L.P.; Hofman, A.; Ikram, M.A.; et al. Sex differences in lifetime risk and first manifestation of cardiovascular disease: Prospective population based cohort study. BMJ 2014, 349, g5992. [Google Scholar] [CrossRef]
- Louis, X.L.; Raj, P.; Chan, L.; Zieroth, S.; Netticadan, T.; Wigle, J.T. Are the cardioprotective effects of the phytoestrogen resveratrol sex-dependent? Can. J. Physiol. Pharmacol. 2019, 97, 503–514. [Google Scholar] [CrossRef]
- Da Silva, J.S.; Montagnoli, T.L.; Rocha, B.S.; Tacco, M.L.C.A.; Marinho, S.C.P.; Zapata-Sudo, G. Estrogen Receptors: Therapeutic Perspectives for the Treatment of Cardiac Dysfunction after Myocardial Infarction. Int. J. Mol. Sci. 2021, 22, 525. [Google Scholar] [CrossRef]
- Feng, Y.; Madungwe, N.; Junho, C.V.C.; Bopassa, J.C. Activation of G protein-coupled oestrogen receptor 1 at the onset of reperfusion protects the myocardium against ischemia/reperfusion injury by reducing mitochondrial dysfunction and mitophagy. Br. J. Pharmacol. 2017, 174, 4329–4344. [Google Scholar] [CrossRef]
- Hewitt, S.C.; Korach, K.S. Estrogen Receptors: New Directions in the New Millennium. Endocr. Rev. 2018, 39, 664–675. [Google Scholar] [CrossRef]
- Ueda, K.; Adachi, Y.; Liu, P.; Fukuma, N.; Takimoto, E. Regulatory Actions of Estrogen Receptor Signaling in the Cardiovascular System. Front. Endocrinol. 2020, 10, 909. [Google Scholar] [CrossRef]
- Shi, D.; Zhao, P.; Cui, L.; Li, H.; Sun, L.; Niu, J.; Chen, M. Inhibition of PI3K/AKT molecular pathway mediated by membrane estrogen receptor GPER accounts for cryptotanshinone induced antiproliferative effect on breast cancer SKBR-3 cells. BMC Pharmacol. Toxicol. 2020, 21, 32. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, M.; Suzuki, T.; Mizukami, Y.; Ikeda, T. The membrane-type estrogen receptor G-protein-coupled estrogen receptor suppresses lipopolysaccharide-induced interleukin 6 via inhibition of nuclear factor-kappa B pathway in murine macrophage cells. Anim. Sci. J. 2017, 88, 1870–1879. [Google Scholar] [CrossRef]
- Machuki, J.O.; Zhang, H.-Y.; Geng, J.; Fu, L.; Adzika, G.K.; Wu, L.; Shang, W.; Wu, J.; Kexue, L.; Zhao, Z.; et al. Estrogen regulation of cardiac cAMP-L-type Ca2+ channel pathway modulates sex differences in basal contraction and responses to β2AR-mediated stress in left ventricular apical myocytes. Cell Commun. Signal. 2019, 17, 34. [Google Scholar] [CrossRef] [PubMed]
- Lam, C.S.P.; Roger, V.L.; Rodeheffer, R.J.; Bursi, F.; Borlaug, B.A.; Ommen, S.R.; Kass, D.A.; Redfield, M.M. Cardiac Structure and Ventricular–Vascular Function in Persons With Heart Failure and Preserved Ejection Fraction From Olmsted County, Minnesota. Circulation 2007, 115, 1982–1990. [Google Scholar] [CrossRef] [PubMed]
- DeBerge, M.; Shah, S.J.; Wilsbacher, L.; Thorp, E.B. Macrophages in Heart Failure with Reduced versus Preserved Ejection Fraction. Trends Mol. Med. 2019, 25, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Gori, M.; Lam, C.S.P.; Gupta, D.K.; Santos, A.B.S.; Cheng, S.; Shah, A.M.; Claggett, B.; Zile, M.; Kraigher-Krainer, E.; Pieske, B.; et al. Sex-specific cardiovascular structure and function in heart failure with preserved ejection fraction. Eur. J. Heart Fail. 2014, 16, 535–542. [Google Scholar] [CrossRef]
- Merz, A.; Cheng, S. Sex differences in cardiovascular ageing. Heart 2016, 102, 825–831. [Google Scholar] [CrossRef]
- Bhuiyan, S.; Fukunaga, K. Characterization of an animal model of postmenopausal cardiac hypertrophy and novel mechanisms responsible for cardiac decompensation using ovariectomized pressure-overloaded rats. Menopause 2010, 17, 213–221. [Google Scholar] [CrossRef]
- Wang, H.; Jessup, J.A.; Zhao, Z.; Da Silva, J.; Lin, M.; MacNamara, L.M.; Ahmad, S.; Chappell, M.C.; Ferrario, C.M.; Groban, L. Characterization of the Cardiac Renin Angiotensin System in Oophorectomized and Estrogen-Replete mRen2. Lewis Rats. PLoS ONE 2013, 8, e76992. [Google Scholar] [CrossRef]
- Ahmad, S.; Sun, X.; Lin, M.; Varagic, J.; Zapata-Sudo, G.; Ferrario, C.M.; Groban, L.; Wang, H. Blunting of estrogen modulation of cardiac cellular chymase/RAS activity and function in SHR. J. Cell. Physiol. 2018, 233, 3330–3342. [Google Scholar] [CrossRef]
- da Silva, J.S.; Gabriel-Costa, D.; Wang, H.; Ahmad, S.; Sun, X.; Varagic, J.; Sudo, R.T.; Ferrario, C.M.; Italia, L.J.D.; Zapata-Sudo, G.; et al. Blunting of cardioprotective actions of estrogen in female rodent heart linked to altered expression of cardiac tissue chymase and ACE2. J. Renin-Angiotensin-Aldosterone Syst. 2017, 18, 1470320317722270. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Sun, X.; Ahmad, S.; Su, J.; Ferrario, C.M.; Groban, L. Estrogen modulates the differential expression of cardiac myocyte chymase isoforms and diastolic function. Mol. Cell. Biochem. 2019, 456, 85–93. [Google Scholar] [CrossRef] [PubMed]
- da Silva, J.S.; Sun, X.; Ahmad, S.; Wang, H.; Sudo, R.T.; Varagic, J.; Ferrario, C.M.; Zapata-Sudo, G.; Groban, L. G-Protein–Coupled Estrogen Receptor Agonist G1 Improves Diastolic Function and Attenuates Cardiac Renin–Angiotensin System Activation in Estrogen-Deficient Hypertensive Rats. J. Cardiovasc. Pharmacol. 2019, 74, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Wang, H.; Hodge, H.; Wright, K.N.; Ahmad, S.; Ferrario, C.M.; Groban, L. Amplifying effect of chronic lisinopril therapy on diastolic function and the angiotensin-(1-7) Axis by the G1 agonist in ovariectomized spontaneously hypertensive rats. Transl. Res. 2021, 235, 62–76. [Google Scholar] [CrossRef]
- Santos, W.V.; Pereira, L.M.; Mandarim-De-Lacerda, C.A. The effect of enalapril on the cardiac remodelling in ovariectomized spontaneously hypertensive rats. Int. J. Exp. Pathol. 2004, 85, 287–294. [Google Scholar] [CrossRef]
- Felix, A.C.S.; Dutra, S.G.; Gastaldi, A.C.; Bonfim, P.C.; Vieira, S.; de Souza, H.C.D. Physical training promotes similar effects to the blockade of angiotensin-converting enzyme on the cardiac morphology and function in old female rats subjected to premature ovarian failure. Exp. Gerontol. 2018, 109, 90–98. [Google Scholar] [CrossRef]
- Lin, Y.-Y.; Hong, Y.; Zhou, M.-C.; Huang, H.-L.; Shyu, W.-C.; Chen, J.-S.; Ting, H.; Cheng, Y.-J.; Yang, A.-L.; Lee, S.-D. Exercise training attenuates cardiac inflammation and fibrosis in hypertensive ovariectomized rats. J. Appl. Physiol. 2020, 128, 1033–1043. [Google Scholar] [CrossRef]
- Bhuiyan, S.; Shioda, N.; Fukunaga, K. Ovariectomy augments pressure overload-induced hypertrophy associated with changes in Akt and nitric oxide synthase signaling pathways in female rats. Am. J. Physiol. Metab. 2007, 293, E1606–E1614. [Google Scholar] [CrossRef]
- Li, J.; Jubair, S.; Janicki, J.S. Estrogen Inhibits Mast Cell Chymase Release to Prevent Pressure Overload-Induced Adverse Cardiac Remodeling. Hypertension 2015, 65, 328–334. [Google Scholar] [CrossRef]
- Wattanathorn, J.; Kawvised, S.; Thukham-Mee, W. Encapsulated Mulberry Fruit Extract Alleviates Changes in an Animal Model of Menopause with Metabolic Syndrome. Oxidative Med. Cell. Longev. 2019, 2019, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Szabó, R.; Karácsonyi, Z.; Börzsei, D.; Juhasz, B.; Al-Awar, A.; Torok, S.; Berkó, A.M.; Takacs, I.; Kupai, K.; Varga, C.; et al. Role of Exercise-Induced Cardiac Remodeling in Ovariectomized Female Rats. Oxidative Med. Cell. Longev. 2018, 2018, 6709742. [Google Scholar] [CrossRef]
- Martínez-Cignoni, M.R.; González-Vicens, A.; Morán-Costoya, A.; Proenza, A.M.; Gianotti, M.; Valle, A.; Lladó, I. Estrogen Impairs Adipose Tissue Expansion and Cardiometabolic Profile in Obese-Diabetic Female Rats. Int. J. Mol. Sci. 2021, 22, 13573. [Google Scholar] [CrossRef]
- Leffler, K.E.; Abdel-Rahman, A.A. Estrogen-Dependent Disruption of Adiponectin-Connexin43 Signaling Underlies Exacerbated Myocardial Dysfunction in Diabetic Female Rats. J. Pharmacol. Exp. Ther. 2019, 368, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Azizian, H.; Khaksari, M.; Asadikaram, G.; Esmailidehaj, M.; Shahrokhi, N. Progesterone eliminates 17β-estradiol-Mediated cardioprotection against diabetic cardiovascular dysfunction in ovariectomized rats. Biomed. J. 2021, 44, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Azizian, H.; Farhadi, Z.; Khaksari, M. Selective estrogen receptor α and β antagonist aggravate cardiovascular dysfunction in type 2 diabetic ovariectomized female rats. Horm. Mol. Biol. Clin. Investig. 2022. [Google Scholar] [CrossRef]
- Wang, X.; Tan, Y.; Xu, B.; Lu, L.; Zhao, M.; Ma, J.; Liang, H.; Liu, J.; Yu, S. GPR30 Attenuates Myocardial Fibrosis in Diabetic Ovariectomized Female Rats: Role of iNOS Signaling. DNA Cell Biol. 2018, 37, 821–830. [Google Scholar] [CrossRef]
- Fouda, M.A.; Leffler, K.E.; Abdel-Rahman, A.A. Estrogen-dependent hypersensitivity to diabetes-evoked cardiac autonomic dysregulation: Role of hypothalamic neuroinflammation. Life Sci. 2020, 250, 117598. [Google Scholar] [CrossRef]
- Hou, H.; Zhao, Z.; Machuki, J.O.; Zhang, L.; Zhang, Y.; Fu, L.; Wu, J.; Liu, Y.; Harding, S.E.; Sun, H. Estrogen deficiency compromised the β2AR-Gs/Gi coupling: Implications for arrhythmia and cardiac injury. Pflugers Arch. Eur. J. Physiol. 2018, 470, 559–570. [Google Scholar] [CrossRef]
- Hou, H.; Adzika, G.K.; Wu, Q.; Ma, T.; Ma, Y.; Geng, J.; Shi, M.; Fu, L.; Rizvi, R.; Gong, Z.; et al. Estrogen Attenuates Chronic Stress-Induced Cardiomyopathy by Adaptively Regulating Macrophage Polarizations via β2-Adrenergic Receptor Modulation. Front. Cell Dev. Biol. 2021, 9, 737003. [Google Scholar] [CrossRef]
- Ferrario, C.M. Cardiac remodelling and RAS inhibition. Ther. Adv. Cardiovasc. Dis. 2016, 10, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Ferrario, C.M.; Ahmad, S.; Varagic, J.; Cheng, C.P.; Groban, L.; Wang, H.; Collawn, J.F.; dell′Italia, L.J. Intracrine angiotensin II functions originate from noncanonical pathways in the human heart. Am. J. Physiol. Circ. Physiol. 2016, 311, H404–H414. [Google Scholar] [CrossRef]
- Reyes, S.; Varagic, J.; Ahmad, S.; VonCannon, J.; Kon, N.D.; Wang, H.; Groban, L.; Cheng, C.P.; Dell’Italia, L.J.; Ferrario, C.M. Novel Cardiac Intracrine Mechanisms Based on Ang-(1-12)/Chymase Axis Require a Revision of Therapeutic Approaches in Human Heart Disease. Curr. Hypertens. Rep. 2017, 19, 16. [Google Scholar] [CrossRef]
- Fu, L.; Wei, C.; Powell, P.C.; Bradley, W.E.; Ahmad, S.; Ferrario, C.M.; Collawn, J.F.; Affairs, V.; States, U.; States, U.; et al. Increased fibroblast chymase production mediates procollagen autophagic digestion in volume overload. J. Mol. Cell. Cardiol. 2016, 92, 1–9. [Google Scholar] [CrossRef][Green Version]
- Borges, C.C.; Penna-De-Carvalho, A.; Junior, J.L.M.; Aguila, M.B.; Mandarim-De-Lacerda, C.A. Ovariectomy modify local renin-angiotensin-aldosterone system gene expressions in the heart of ApoE (−/−) mice. Life Sci. 2017, 191, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lyu, L.; Wang, H.; Li, B.; Qin, Q.; Qi, L.; Nagarkatti, M.; Nagarkatti, P.; Janicki, J.S.; Wang, X.L.; Cui, T. A critical role of cardiac fibroblast-derived exosomes in activating renin angiotensin system in cardiomyocytes. J. Mol. Cell. Cardiol. 2015, 89, 268–279. [Google Scholar] [CrossRef]
- Lützelschwab, C.; Pejler, G.; Aveskogh, M.; Hellman, L. Secretory Granule Proteases in Rat Mast Cells. Cloning of 10 Different Serine Proteases and a Carboxypeptidase A from Various Rat Mast Cell Populations. J. Exp. Med. 1997, 185, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Caughey, G.H.; Raymond, W.W.; Wolters, P.J. Angiotensin II generation by mast cell α- and β-chymases. Biochim. Biophys. Acta 2000, 1480, 245–257. [Google Scholar] [CrossRef]
- Pedram, A.; Razandi, M.; Narayanan, R.; Levin, E.R. Estrogen receptor beta signals to inhibition of cardiac fibrosis. Mol. Cell. Endocrinol. 2016, 434, 57–68. [Google Scholar] [CrossRef]
- Ponnusamy, S.; Tran, Q.T.; Harvey, I.; Smallwood, H.S.; Thiyagarajan, T.; Banerjee, S.; Johnson, D.L.; Dalton, J.T.; Sullivan, R.D.; Miller, D.D.; et al. Pharmacologic activation of estrogen receptor α increases mitochondrial function, energy expenditure, and brown adipose tissue. FASEB J. 2017, 31, 266–281. [Google Scholar] [CrossRef]
- Meyers, M.; Sun, J.; Carlson, K.E.; Marriner, G.A.; Katzenellenbogen, B.S.; Katzenellenbogen, J.A. Estrogen Receptor-β Potency-Selective Ligands: Structure−Activity Relationship Studies of Diarylpropionitriles and Their Acetylene and Polar Analogues. J. Med. Chem. 2001, 44, 4230–4251. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhan, Y.; Ono, K.; Yin, Y.; Wang, L.; Wei, M.; Ji, L.; Liu, M.; Liu, G.; Zhou, X.; et al. Pharmacological activation of estrogenic receptor G protein-coupled receptor 30 attenuates angiotensin II-induced atrial fibrosis in ovariectomized mice by modulating TGF-β1/smad pathway. Mol. Biol. Rep. 2022, 49, 6341–6355. [Google Scholar] [CrossRef] [PubMed]
- Dutra, S.G.V.; Felix, A.C.S.; Gastaldi, A.C.; Facioli, T.D.P.; Vieira, S.; De Souza, H.C.D. Chronic treatment with angiotensin-converting enzyme inhibitor increases cardiac fibrosis in young rats submitted to early ovarian failure. Basic Clin. 2017, 206, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Newson, L. Menopause and cardiovascular disease. Post Reprod. Health 2018, 24, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.B. Menopause and Chronic Kidney Disease. Semin. Nephrol. 2017, 37, 404–411. [Google Scholar] [CrossRef]
- Carr, M.C. The Emergence of the Metabolic Syndrome with Menopause. J. Clin. Endocrinol. Metab. 2003, 88, 2404–2411. [Google Scholar] [CrossRef]
- Medina-Contreras, J.; Villalobos-Molina, R.; Zarain-Herzberg, A.; Balderas-Villalobos, J. Ovariectomized rodents as a menopausal metabolic syndrome model. A minireview. Mol. Cell. Biochem. 2020, 475, 261–276. [Google Scholar] [CrossRef]
- Zhang, H.; Ma, Z.F.; Luo, X.; Li, X. Effects of Mulberry Fruit (Morus alba L.) Consumption on Health Outcomes: A Mini-Review. Antioxidants 2018, 7, 69. [Google Scholar] [CrossRef]
- Jacob-Ferreira, A.L.; Kondo, M.Y.; Baral, P.K.; James, M.; Holt, A.; Fan, X.; Schulz, R. Phosphorylation Status of 72 kDa MMP-2 Determines Its Structure and Activity in Response to Peroxynitrite. PLoS ONE 2013, 8, e71794. [Google Scholar] [CrossRef]
- Liang, D.; Zhuo, Y.; Guo, Z.; He, L.; Wang, X.; He, Y.; Li, L.; Dai, H. SIRT1/PGC-1 pathway activation triggers autophagy/mitophagy and attenuates oxidative damage in intestinal epithelial cells. Biochimie 2020, 170, 10–20. [Google Scholar] [CrossRef]
- Singh, V.; Ubaid, S. Role of Silent Information Regulator 1 (SIRT1) in Regulating Oxidative Stress and Inflammation. Inflammation 2020, 43, 1589–1598. [Google Scholar] [CrossRef] [PubMed]
- Rada, P.; Pardo, V.; Mobasher, M.A.; García-Martínez, I.; Ruiz, L.; González-Rodríguez, A.; Sanchez-Ramos, C.; Muntané, J.; Alemany, S.; James, L.P.; et al. SIRT1 Controls Acetaminophen Hepatotoxicity by Modulating Inflammation and Oxidative Stress. Antioxid. Redox Signal. 2018, 28, 1187–1208. [Google Scholar] [CrossRef] [PubMed]
- Tamer, S.A.; Altınoluk, T.; Emran, M.; Korkmaz, S.; Yüksel, R.G.; Baykal, Z.; Dur, Z.S.; Levent, H.N.; Ural, M.A.; Yüksel, M.; et al. Melatonin Alleviates Ovariectomy-Induced Cardiovascular Inflammation in Sedentary or Exercised Rats by Upregulating SIRT1. Inflammation 2022, 45, 2202–2222. [Google Scholar] [CrossRef]
- Murphy, E.; Amanakis, G.; Fillmore, N.; Parks, R.J.; Sun, J. Sex differences in metabolic cardiomyopathy. Cardiovasc. Res. 2017, 113, 370–377. [Google Scholar] [CrossRef]
- Ramesh, P.; Yeo, J.L.; Brady, E.M.; McCann, G.P. Role of inflammation in diabetic cardiomyopathy. Ther. Adv. Endocrinol. Metab. 2022, 13, 204201882210835. [Google Scholar] [CrossRef] [PubMed]
- Tawfik, S.H.; Mahmoud, B.F.; Saad, M.I.; Shehata, M.; Kamel, M.A.; Helmy, M.H. Similar and Additive Effects of Ovariectomy and Diabetes on Insulin Resistance and Lipid Metabolism. Biochem. Res. Int. 2015, 2015, 567945. [Google Scholar] [CrossRef] [PubMed]
- Esmailidehaj, M.; Kuchakzade, F.; Rezvani, M.E.; Farhadi, Z.; Esmaeili, H.; Azizian, H. 17β-Estradiol improves insulin signalling and insulin resistance in the aged female hearts: Role of inflammatory and anti-inflammatory cytokines. Life Sci. 2020, 253, 117673. [Google Scholar] [CrossRef]
- White, R.E.; Gerrity, R.; Barman, S.A.; Han, G. Estrogen and oxidative stress: A novel mechanism that may increase the risk for cardiovascular disease in women. Steroids 2010, 75, 788–793. [Google Scholar] [CrossRef]
- Rochette, L.; Zeller, M.; Cottin, Y.; Vergely, C. Diabetes, oxidative stress and therapeutic strategies. Biochim. Biophys. Acta 2014, 1840, 2709–2729. [Google Scholar] [CrossRef]
- Gong, H.; Chen, Z.; Zhang, X.; Li, Y.; Zhang, J.; Chen, Y.; Ding, Y.; Zhang, G.; Yang, C.; Zhu, Y.; et al. Urotensin II Protects Cardiomyocytes from Apoptosis Induced by Oxidative Stress through the CSE/H2S Pathway. Int. J. Mol. Sci. 2015, 16, 12482–12498. [Google Scholar] [CrossRef]
- Yao, F.; Abdel-Rahman, A.A. Estrogen receptor ERα plays a major role in ethanol-evoked myocardial oxidative stress and dysfunction in conscious female rats. Alcohol 2016, 50, 27–35. [Google Scholar] [CrossRef]
- Yao, F.; Abdel-Rahman, A.A. Estrogen Receptors α and β Play Major Roles in Ethanol-Evoked Myocardial Oxidative Stress and Dysfunction in Conscious Ovariectomized Rats. Alcohol. Clin. Exp. Res. 2017, 41, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Phillis, J. The effects of selective A1 and A2a adenosine receptor antagonists on cerebral ischemic injury in the gerbil. Brain Res. 1995, 705, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Stockwell, J.; Jakova, E.; Cayabyab, F.S. Adenosine A1 and A2A Receptors in the Brain: Current Research and Their Role in Neurodegeneration. Molecules 2017, 22, 676. [Google Scholar] [CrossRef] [PubMed]
- Curl, C.; Wendt, I.; Kotsanas, G. Effects of gender on intracellular [Ca2+] in rat cardiac myocytes. Pflügers Arch. 2001, 441, 709–716. [Google Scholar] [CrossRef]
- Cao, X.; Zhou, C.; Chong, J.; Fu, L.; Zhang, L.; Sun, D.; Hou, H.; Zhang, Y.; Li, D.; Sun, H. Estrogen resisted stress-induced cardiomyopathy through increasing the activity of β2AR–Gαs signal pathway in female rats. Int. J. Cardiol. 2015, 187, 377–386. [Google Scholar] [CrossRef]
- Fujita, T.; Umemura, M.; Yokoyama, U.; Okumura, S.; Ishikawa, Y. The role of Epac in the heart. Cell. Mol. Life Sci. 2017, 74, 591–606. [Google Scholar] [CrossRef]
- Parks, R.J.; Bogachev, O.; Mackasey, M.; Ray, G.; Rose, R.A.; Howlett, S.E. The impact of ovariectomy on cardiac excitation-contraction coupling is mediated through cAMP/PKA-dependent mechanisms. J. Mol. Cell. Cardiol. 2017, 111, 51–60. [Google Scholar] [CrossRef]
- Parks, R.J.; Howlett, S.E. H-89 decreases the gain of excitation–contraction coupling and attenuates calcium sparks in the absence of beta-adrenergic stimulation. Eur. J. Pharmacol. 2012, 691, 163–172. [Google Scholar] [CrossRef]
- Fares, E.; Parks, R.J.; MacDonald, J.K.; Egar, J.M.; Howlett, S.E. Ovariectomy enhances SR Ca2+ release and increases Ca2+ spark amplitudes in isolated ventricular myocytes. J. Mol. Cell. Cardiol. 2012, 52, 32–42. [Google Scholar] [CrossRef]
- Bhupathy, P.; Babu, G.J.; Ito, M.; Periasamy, M. Threonine-5 at the N-terminus can modulate sarcolipin function in cardiac myocytes. J. Mol. Cell. Cardiol. 2009, 47, 723–729. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, J.; Liu, S.; Wang, W.; Chen, X.; Jiang, H.; Li, J.; Wang, K.; Bai, W.; Zhang, H.; et al. Effects of oestrogen and Cimicifuga racemosa on the cardiac noradrenaline pathway of ovariectomized rats. Exp. Physiol. 2017, 102, 974–984. [Google Scholar] [CrossRef] [PubMed]
- Eskin, B.A.; Snyder, D.L.; Roberts, J.; Aloyo, V.J. Cardiac norepinephrine release: Modulation by ovariectomy and estrogen. Exp. Biol. Med. 2003, 228, 194–199. [Google Scholar] [CrossRef]
- Wehrwein, E.A.; Novotny, M.; Swain, G.M.; Parker, L.M.; Esfahanian, M.; Spitsbergen, J.M.; Habecker, B.A.; Kreulen, D.L. Regional changes in cardiac and stellate ganglion norepinephrine transporter in DOCA–salt hypertension. Auton. Neurosci. 2013, 179, 99–107. [Google Scholar] [CrossRef]
- Budde, H.; Hassoun, R.; Mügge, A.; Kovács, Á.; Hamdani, N. Current Understanding of Molecular Pathophysiology of Heart Failure With Preserved Ejection Fraction. Front. Physiol. 2022, 13, 928232. [Google Scholar] [CrossRef] [PubMed]
- Merrill, M.; Sweitzer, N.K.; Lindenfeld, J.; Kao, D.P. Sex Differences in Outcomes and Responses to Spironolactone in Heart Failure With Preserved Ejection Fraction: A Secondary Analysis of TOPCAT Trial. JACC Heart Fail. 2019, 7, 228–238. [Google Scholar] [CrossRef]
- Solomon, S.D.; McMurray, J.J.V.; Anand, I.S.; Junbo Ge, D.P.; Lam, C.S.P.; Maggioni, A.P.; Martinez, F.; Packer, M.; Pfeffer, M.A.; Pieske, B.; et al. Angiotensin–Neprilysin Inhibition in Heart Failure with Preserved Ejection Fraction. N. Engl. J. Med. 2019, 381, 1609–1620. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, S.; Claggett, B.L.; Böhm, M.; Anker, S.D.; Vaduganathan, M.; Zannad, F.; Pieske, B.; Lam, C.S.; Anand, I.S.; Shi, V.C.; et al. Systolic Blood Pressure in Heart Failure With Preserved Ejection Fraction Treated With Sacubitril/Valsartan. J. Am. Coll. Cardiol. 2020, 75, 1644–1656. [Google Scholar] [CrossRef]
- Cunningham, J.W.; Vaduganathan, M.; Claggett, B.L.; Zile, M.R.; Anand, I.S.; Packer, M.; Zannad, F.; Lam, C.S.; Janssens, S.; Jhund, P.S.; et al. Effects of Sacubitril/Valsartan on N-Terminal Pro-B-Type Natriuretic Peptide in Heart Failure With Preserved Ejection Fraction. JACC: Heart Fail. 2020, 8, 372–381. [Google Scholar] [CrossRef]
- Pieske, B.; Wachter, R.; Shah, S.J.; Baldridge, A.; Szeczoedy, P.; Ibram, G.; Shi, V.; Zhao, Z.; Cowie, M.R.; Prado, A.C.; et al. Effect of Sacubitril/Valsartan vs Standard Medical Therapies on Plasma NT-proBNP Concentration and Submaximal Exercise Capacity in Patients With Heart Failure and Preserved Ejection Fraction: The PARALLAX Randomized Clinical Trial. JAMA 2021, 326, 1919–1929. [Google Scholar] [CrossRef]
- Butt, J.H.; Dewan, P.; Jhund, P.S.; Anand, I.S.; Atar, D.; Ge, J.; Desai, A.S.; Echeverria, L.E.; Køber, L.; Lam, C.S.; et al. Sacubitril/Valsartan and Frailty in Patients With Heart Failure and Preserved Ejection Fraction. J. Am. Coll. Cardiol. 2022, 80, 1130–1143. [Google Scholar] [CrossRef]
- Packer, M.; Anker, S.D.; Butler, J.; Filippatos, G.; Pocock, S.J.; Carson, P.; Januzzi, J.; Verma, S.; Tsutsui, H.; Brueckmann, M.; et al. Cardiovascular and Renal Outcomes with Empagliflozin in Heart Failure. N. Engl. J. Med. 2020, 383, 1413–1424. [Google Scholar] [CrossRef]
- Solomon, S.D.; McMurray, J.J.; Claggett, B.; de Boer, R.A.; DeMets, D.; Hernandez, A.F.; Inzucchi, S.E.; Kosiborod, M.N.; Lam, C.S.; Martinez, F.; et al. Dapagliflozin in Heart Failure with Mildly Reduced or Preserved Ejection Fraction. N. Engl. J. Med. 2022, 387, 1089–1098. [Google Scholar] [CrossRef]
- Nassif, M.E.; Windsor, S.L.; Borlaug, B.A.; Kitzman, D.W.; Shah, S.J.; Tang, F.; Khariton, Y.; Malik, A.O.; Khumri, T.; Umpierrez, G.; et al. The SGLT2 inhibitor dapagliflozin in heart failure with preserved ejection fraction: A multicenter randomized trial. Nat. Med. 2021, 27, 1954–1960. [Google Scholar] [CrossRef]
- Van Tassell, B.W.; Trankle, C.R.; Canada, J.M.; Carbone, S.; Buckley, L.; Kadariya, D.; Del Buono, M.G.; Billingsley, H.; Wohlford, G.; Viscusi, M.; et al. IL-1 Blockade in Patients With Heart Failure With Preserved Ejection Fraction. Circ. Heart Fail. 2018, 11, e005036. [Google Scholar] [CrossRef]
- McMurray, J.J.; Jackson, A.M.; Lam, C.S.; Redfield, M.M.; Anand, I.S.; Ge, J.; Lefkowitz, M.P.; Maggioni, A.P.; Martinez, F.; Packer, M.; et al. Effects of Sacubitril-Valsartan Versus Valsartan in Women Compared With Men With Heart Failure and Preserved Ejection Fraction: Insights from PARAGON-HF. Circulation 2020, 141, 338–351. [Google Scholar] [CrossRef] [PubMed]
- Anker, S.D.; Butler, J.; Filippatos, G.; Ferreira, J.P.; Bocchi, E.; Böhm, M.; Brunner–La Rocca, H.-P.; Choi, D.-J.; Chopra, V.; Chuquiure-Valenzuela, E.; et al. Empagliflozin in Heart Failure with a Preserved Ejection Fraction. N. Engl. J. Med. 2021, 385, 1451–1461. [Google Scholar] [CrossRef] [PubMed]
- Requena-Ibanez, J.A.; Santos-Gallego, C.G.; Zafar, M.U.; Badimon, J.J. SGLT2-Inhibitors on HFpEF Patients. Role of Ejection Fraction. Role of Ejection Fraction. Cardiovasc. Drugs Ther. 2022, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Liang, Y.; Gu, N. Pharmacological mechanisms of sodium-glucose co-transporter 2 inhibitors in heart failure with preserved ejection fraction. BMC Cardiovasc. Disord. 2022, 22, 261. [Google Scholar] [CrossRef]
- Kolijn, D.; Pabel, S.; Tian, Y.; Lódi, M.; Herwig, M.; Carrizzo, A.; Zhazykbayeva, S.; Kovács, Á.; Fülöp, G.; Falcão-Pires, I.; et al. Empagliflozin improves endothelial and cardiomyocyte function in human heart failure with preserved ejection fraction via reduced pro-inflammatory-oxidative pathways and protein kinase Gα oxidation. Cardiovasc. Res. 2021, 117, 495–507. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Kuwahara, K. Sodium-Glucose Cotransporter-2 inhibitors are potential therapeutic agents for treatment of non-diabetic heart failure patients. J. Cardiol. 2020, 76, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Fukuta, H.; Goto, T.; Wakami, K.; Kamiya, T.; Ohte, N. Effect of beta-blockers on heart failure severity in patients with heart failure with preserved ejection fraction: A meta-analysis of randomized controlled trials. Heart Fail. Rev. 2021, 26, 165–171. [Google Scholar] [CrossRef]
- Miller, V.M.; Mankad, R. Sex Steroids and Incident Cardiovascular Disease in Post-Menopausal Women. J. Am. Coll. Cardiol. 2018, 71, 2567–2569. [Google Scholar] [CrossRef]
- Gersh, F.L.; Lavie, C.J. Menopause and hormone replacement therapy in the 21st century. Heart 2020, 106, 479–481. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Klein, L.; Eaton, C.; Panjrath, G.; Martin, L.W.; Chae, C.U.; Greenland, P.; Lloyd-Jones, D.M.; Wactawski-Wende, J.; Manson, J.E. Menopausal Hormone Therapy and Risks of First Hospitalized Heart Failure and its Subtypes During the Intervention and Extended Postintervention Follow-up of the Women’s Health Initiative Randomized Trials. J. Card. Fail. 2020, 26, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Prentice, R.L.; Aragaki, A.K.; Chlebowski, R.T.; Rossouw, J.E.; Anderson, G.L.; Stefanick, M.L.; Wactawski-Wende, J.; Kuller, L.H.; Wallace, R.; Johnson, K.C.; et al. Randomized Trial Evaluation of the Benefits and Risks of Menopausal Hormone Therapy Among Women 50–59 Years of Age. Am. J. Epidemiol. 2020, 190, 365–375. [Google Scholar] [CrossRef]
- Johansen, N.; Liavaag, A.H.; Iversen, O.-E.; Dørum, A.; Braaten, T.; Michelsen, T.M. Use of hormone replacement therapy after risk-reducing salpingo-oophorectomy. Acta Obstet. Gynecol. Scand. 2017, 96, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Fait, T. Menopause hormone therapy: Latest developments and clinical practice. Drugs Context 2019, 8, 1–9. [Google Scholar] [CrossRef]
- Thomas, A.J.; Ismail, R.; Taylor-Swanson, L.; Cray, L.; Schnall, J.G.; Mitchell, E.S.; Woods, N.F. Effects of isoflavones and amino acid therapies for hot flashes and co-occurring symptoms during the menopausal transition and early postmenopause: A systematic review. Maturitas 2014, 78, 263–276. [Google Scholar] [CrossRef]
- Gencel, V.B.; Benjamin, M.; Bahou, S.N.; Khalil, R.A. Vascular effects of phytoestrogens and alternative menopausal hormone therapy in cardiovascular disease. Mini-Reviews Med. Chem. 2012, 12, 149–174. [Google Scholar] [CrossRef]
- Roberts, H.; Lethaby, A. Phytoestrogens for menopausal vasomotor symptoms: A Cochrane review summary. Maturitas 2014, 78, 79–81. [Google Scholar] [CrossRef]
- Barsky, L.; Cook-Wiens, G.; Doyle, M.; Shufelt, C.; Rogers, W.; Reis, S.; Pepine, C.J.; Merz, C.N.B. Phytoestrogen blood levels and adverse outcomes in women with suspected ischemic heart disease. Eur. J. Clin. Nutr. 2021, 75, 829–835. [Google Scholar] [CrossRef]
- Wolters, M.; Dejanovic, G.M.; Asllanaj, E.; Günther, K.; Pohlabeln, H.; Bramer, W.M.; Ahrens, J.; Nagrani, R.; Pigeot, I.; Franco, O.H.; et al. Effects of phytoestrogen supplementation on intermediate cardiovascular disease risk factors among postmenopausal women: A meta-analysis of randomized controlled trials. Menopause 2020, 27, 1081–1092. [Google Scholar] [CrossRef] [PubMed]
- Amerizadeh, A.; Asgary, S.; Vaseghi, G.; Farajzadegan, Z. Effect of Genistein Intake on Some Cardiovascular Risk Factors: An Updated Systematic Review and Meta-analysis. Curr. Probl. Cardiol. 2022, 47, 100902. [Google Scholar] [CrossRef] [PubMed]
- Błaszczuk, A.; Barańska, A.; Kanadys, W.; Malm, M.; Jach, M.E.; Religioni, U.; Wróbel, R.; Herda, J.; Polz-Dacewicz, M. Role of Phytoestrogen-Rich Bioactive Substances (Linumusitatissimum L., Glycine max L., Trifolium pratense L.) in Cardiovascular Disease Prevention in Postmenopausal Women: A Systematic Review and Meta-Analysis. Nutrients 2022, 14, 2467. [Google Scholar] [CrossRef] [PubMed]
- Campos, J.R.; Severino, P.; Ferreira, C.S.; Zielinska, A.; Santini, A.; Souto, S.B.; Souto, E.B. Linseed Essential Oil—Source of Lipids as Active Ingredients for Pharmaceuticals and Nutraceuticals. Curr. Med. Chem. 2019, 26, 4537–4558. [Google Scholar] [CrossRef]
- Lemežienė, N.; Padarauskas, A.; Butkutė, B.; Cesevičienė, J.; Taujenis, L.; Norkevičienė, E. The concentration of isoflavones in red clover (Trifolium pratense L.) at flowering stage. Zemdirb. Agric. 2015, 102, 443–448. [Google Scholar] [CrossRef]
- Moradi, M.; Daneshzad, E.; Azadbakht, L. The effects of isolated soy protein, isolated soy isoflavones and soy protein containing isoflavones on serum lipids in postmenopausal women: A systematic review and meta-analysis. Crit. Rev. Food Sci. Nutr. 2020, 60, 3414–3428. [Google Scholar] [CrossRef]
- Si, C.L.; Qin, P.P.; Lu, Y.Y.; Wu, L.; Wang, H.H.; Hui, L.F.; Liu, Z.; Ni, Y.H. GC-MS Analysis of Chemical Composition and Free Radical Scavenging Activity of Elaeagnus angustifolia Bark. Adv. Mater. Res. 2011, 183–185, 854–858. [Google Scholar] [CrossRef]
- Shabani, M.; Rezaei, A.; Badehnoosh, B.; Qorbani, M.; Yaseri, M.; Ramezani, R.; Emaminia, F. The effects of Elaeagnus angustifolia L. on lipid and glycaemic profiles and cardiovascular function in menopausal women: A double-blind, randomized, placebo-controlled study. Int. J. Clin. Pract. 2020, 75, e13812. [Google Scholar] [CrossRef]
- Howard, B.V.; Van Horn, L.; Hsia, J.; Manson, J.E.; Stefanick, M.L.; Wassertheil-Smoller, S.; Kuller, L.H.; Lacroix, A.Z.; Langer, R.D.; Lasser, N.L.; et al. Low-Fat Dietary Pattern and Risk of Cardiovascular Disease: The Women’s Health Initiative Randomized Controlled Dietary Modification Trial. JAMA 2006, 295, 655–666. [Google Scholar] [CrossRef] [PubMed]
- Ramsden, C.E.; Zamora, D.; Majchrzak-Hong, S.; Faurot, K.; Broste, S.K.; Frantz, R.; Davis, J.M.; Ringel, A.; Suchindran, C.M.; Hibbeln, J.R. Re-evaluation of the traditional diet-heart hypothesis: Analysis of recovered data from Minnesota Coronary Experiment (1968–73). BMJ 2016, 353, i1246. [Google Scholar] [CrossRef]
- Noakes, T.D. Hiding unhealthy heart outcomes in a low-fat diet trial: The Women’s Health Initiative Randomized Controlled Dietary Modification Trial finds that postmenopausal women with established coronary heart disease were at increased risk of an adverse outcome if they consumed a low-fat ‘heart-healthy’ diet. Open Heart 2021, 8, e001680. [Google Scholar] [CrossRef] [PubMed]
- Chiavaroli, L.; Nishi, S.K.; Khan, T.A.; Braunstein, C.R.; Glenn, A.J.; Mejia, S.B.; Rahelić, D.; Kahleová, H.; Salas-Salvadó, J.; Jenkins, D.J.; et al. Portfolio Dietary Pattern and Cardiovascular Disease: A Systematic Review and Meta-analysis of Controlled Trials. Prog. Cardiovasc. Dis. 2018, 61, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Glenn, A.J.; Lo, K.; Jenkins, D.J.A.; Boucher, B.A.; Hanley, A.J.; Kendall, C.W.C.; Manson, J.E.; Vitolins, M.Z.; Snetselaar, L.G.; Liu, S.; et al. Relationship Between a Plant-Based Dietary Portfolio and Risk of Cardiovascular Disease: Findings From the Women’s Health Initiative Prospective Cohort Study. J. Am. Heart Assoc. 2021, 10, 021515. [Google Scholar] [CrossRef]
- Noel, C.A.; LaMonte, M.J.; Roberts, M.B.; Pearlman, D.H.; Banack, H.; Allison, M.; Shadyab, A.H.; Haring, B.; Laddu, D.; Martin, L.W.; et al. Healthy lifestyle and risk of incident heart failure with preserved and reduced ejection fraction among post-menopausal women: The Women’s Health Initiative study. Prev. Med. 2020, 138, 106155. [Google Scholar] [CrossRef]
- Ho, J.E.; Gona, P.; Pencina, M.J.; Tu, J.V.; Austin, P.C.; Vasan, R.S.; Kannel, W.B.; D’Agostino, R.B.; Lee, D.S.; Levy, D. Discriminating clinical features of heart failure with preserved vs. reduced ejection fraction in the community. Eur. Heart J. 2012, 33, 1734–1741. [Google Scholar] [CrossRef]
- LaMonte, M.J.; Manson, J.E.; Chomistek, A.K.; Larson, J.C.; Lewis, C.E.; Bea, J.W.; Johnson, K.C.; Li, W.; Klein, L.; LaCroix, A.Z.; et al. Physical Activity and Incidence of Heart Failure in Postmenopausal Women. JACC Heart Fail. 2018, 6, 983–995. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Drugs | Response | % Women in Clinical Trial | References |
|---|---|---|---|
| Perindoprol, candesartan, irbesartan, spironolactone, sacubitril/valsartan (Sac/Val) | Mostly neutral response | - | [10,96] |
| Spironolactone | Neutral response between the sexes: ↓ All-cause mortality in women | 49.9 | [97] |
| Sac/Val | Potential benefit in women | ≈50 | [98] |
| Sac/Val | ↓ SBP more in women than in men ↓ Plasma levels of NT-proBNP No response on primary outcome | 52 | [99] |
| Sac/Val | ↓ Plasma levels of NT-proBNP in both sexes | 50 | [100,101] |
| ↓ Greater risk of hospitalization for HF in women than in men | [100] | ||
| 5Sac/Val | ↓ Primary outcome in the fragilest patient ↓↓ Secondary clinical outcomes for most fragile patients | 50 | [102] |
| Empagliflozin | ↓ Risk of cardiovascular death and HF hospitalization | 24 | [103] |
| Dapagliflozin | ↓ Risk of worsening HF events or death regardless of sex | 44 | [104] |
| Dapaglifozin | Improves primary endpoint and secondary endpoint ↓ Weight | 57 | [105] |
| Anakinra | ↓ Plasma levels of CRP and NT-proBNP | 64 | [106] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva, J.S.; Montagnoli, T.L.; de Sá, M.P.L.; Zapata-Sudo, G. Heart Failure in Menopause: Treatment and New Approaches. Int. J. Mol. Sci. 2022, 23, 15140. https://doi.org/10.3390/ijms232315140
da Silva JS, Montagnoli TL, de Sá MPL, Zapata-Sudo G. Heart Failure in Menopause: Treatment and New Approaches. International Journal of Molecular Sciences. 2022; 23(23):15140. https://doi.org/10.3390/ijms232315140
Chicago/Turabian Styleda Silva, Jaqueline S., Tadeu Lima Montagnoli, Mauro Paes Leme de Sá, and Gisele Zapata-Sudo. 2022. "Heart Failure in Menopause: Treatment and New Approaches" International Journal of Molecular Sciences 23, no. 23: 15140. https://doi.org/10.3390/ijms232315140
APA Styleda Silva, J. S., Montagnoli, T. L., de Sá, M. P. L., & Zapata-Sudo, G. (2022). Heart Failure in Menopause: Treatment and New Approaches. International Journal of Molecular Sciences, 23(23), 15140. https://doi.org/10.3390/ijms232315140