
Perinatal Stem Cell Therapy to Treat Type 1 Diabetes Mellitus: A Never-Say-Die Story of Differentiation and Immunomodulation

 ,
,  ,
,  ,
,  ,
,  and
and
Abstract
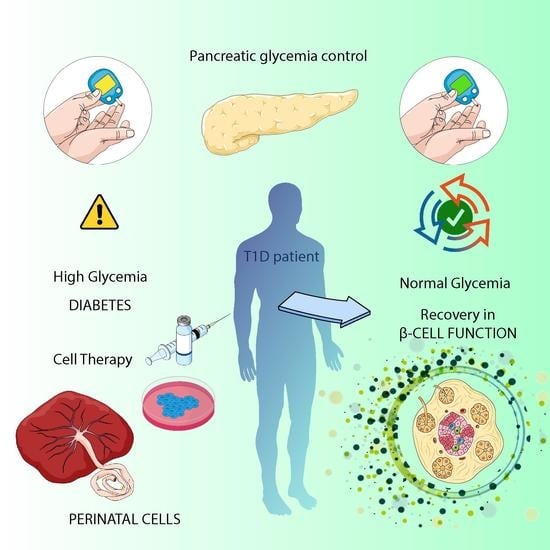
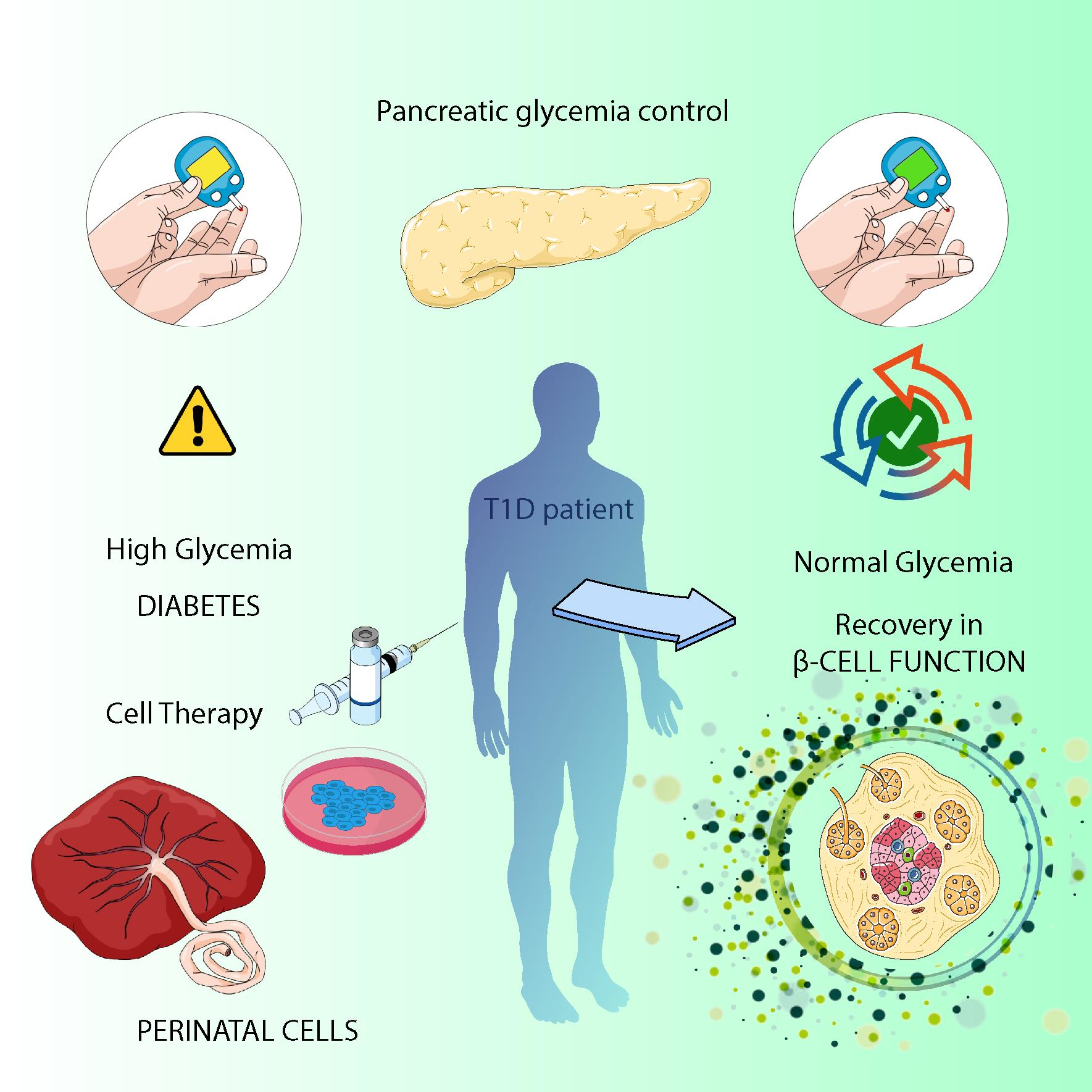
1. Introduction
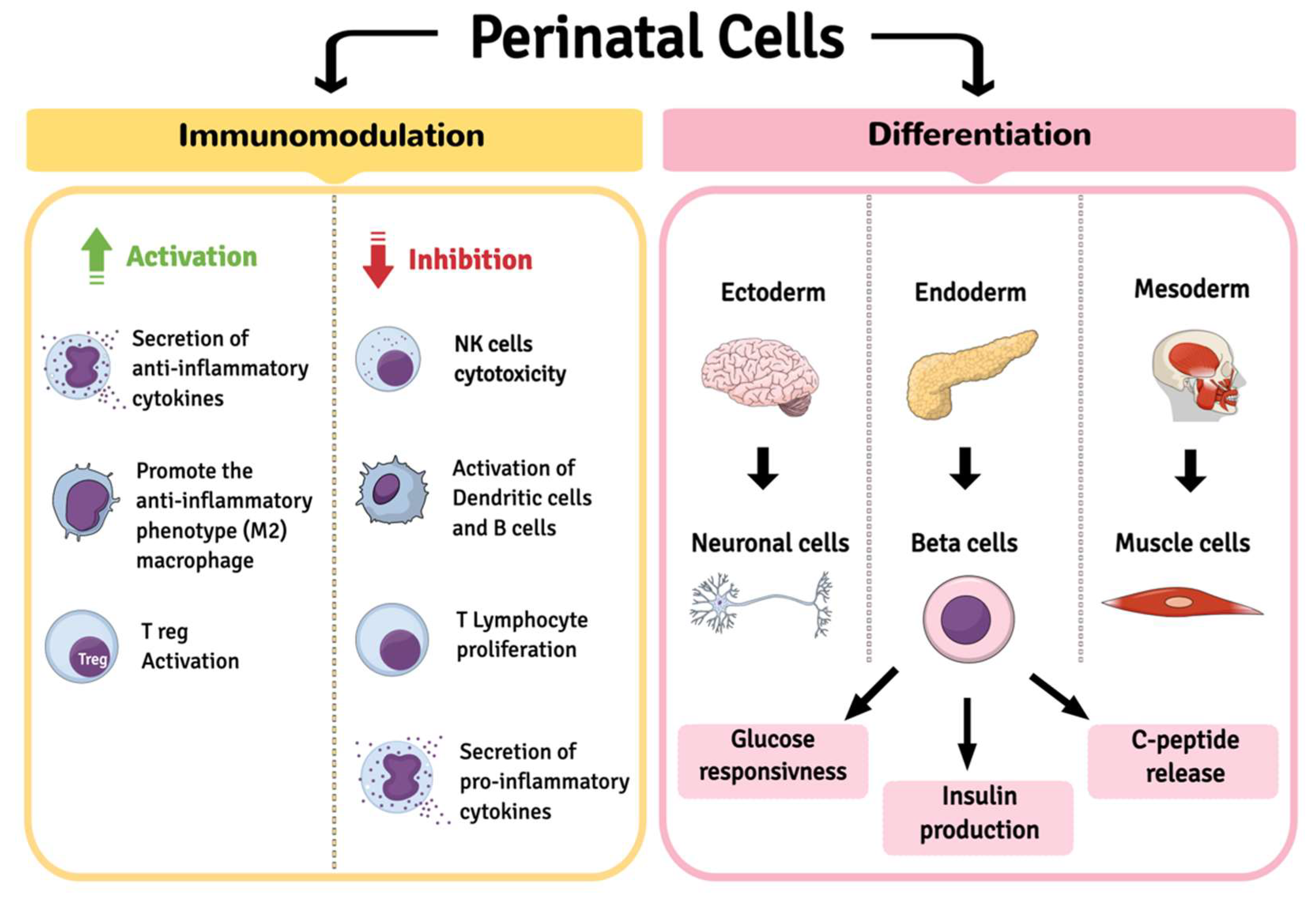
2. Perinatal Cells
2.1. Amniotic Epithelial Cells (AECs)
2.1.1. Differentiating Potential of AECs
2.1.2. Immunomodulatory Capacity of AECs
2.2. Wharton’s Jelly Mesenchymal Stem/Stromal Cells (WJ-MSCs)
2.2.1. Differentiating Potential of WJ-MSCs
2.2.2. Immunomodulatory Capacity of WJ-MSCs
2.2.3. Co-Culture Strategies Based on WJ-MSCs
2.3. Other Sources of Perinatal Cells
2.3.1. Amniotic Fluid Stem Cells (AFSCs)
2.3.2. Amniotic Membrane Mesenchymal Stem Cells (AM-MSCs)
3. Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marchetti, P.; Bugliani, M.; De Tata, V.; Suleiman, M.; Marselli, L. Pancreatic Beta Cell Identity in Humans and the Role of Type 2 Diabetes. Front. Cell Dev. Biol. 2017, 5, 55. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association. Diagnosis and Classification of Diabetes Mellitus. Diabetes Care 2014, 37 (Suppl. S1), S81–S90. [Google Scholar] [CrossRef]
- Maahs, D.M.; West, N.A.; Lawrence, J.M.; Mayer-Davis, E.J. Chapter 1: Epidemiology of Type 1 Diabetes. Endocrinol. Metab. Clin. N. Am. 2010, 39, 481–497. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, M.A.; Eisenbarth, G.S.; Michels, A.W. Type 1 Diabetes. Lancet 2014, 383, 69–82. [Google Scholar] [CrossRef]
- Desai, S.; Deshmukh, A. Mapping of Type 1 Diabetes Mellitus. Curr. Diabetes Rev. 2020, 16, 438–441. [Google Scholar] [CrossRef]
- Pepper, A.R.; Bruni, A.; Shapiro, A.M.J. Clinical Islet Transplantation: Is the Future Finally Now? Curr. Opin. Organ Transpl. 2018, 23, 428–439. [Google Scholar] [CrossRef]
- Skyler, J.S. Hope vs Hype: Where Are We in Type 1 Diabetes? Diabetologija 2018, 61, 509–516. [Google Scholar] [CrossRef]
- Sneddon, J.B.; Tang, Q.; Stock, P.; Bluestone, J.A.; Roy, S.; Desai, T.; Hebrok, M. Stem Cell Therapies for Treating Diabetes: Progress and Remaining Challenges. Cell Stem Cell 2018, 22, 810–823. [Google Scholar] [CrossRef]
- Shapiro, A.M.J.; Pokrywczynska, M.; Ricordi, C. Clinical Pancreatic Islet Transplantation. Nat. Rev. Endocrinol. 2017, 13, 268–277. [Google Scholar] [CrossRef] [PubMed]
- DiMeglio, L.A.; Evans-Molina, C.; Oram, R.A. Type 1 Diabetes. Lancet 2018, 391, 2449–2462. [Google Scholar] [CrossRef]
- Zakrzewski, W.; Dobrzyński, M.; Szymonowicz, M.; Rybak, Z. Stem Cells: Past, Present, and Future. Stem Cell Res. Ther. 2019, 10, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, K.G.; Millman, J.R. Applications of IPSC-Derived Beta Cells from Patients with Diabetes. Cell Rep. Med. 2021, 2, 100238. [Google Scholar] [CrossRef]
- Kroon, E.; Martinson, L.A.; Kadoya, K.; Bang, A.G.; Kelly, O.G.; Eliazer, S.; Young, H.; Richardson, M.; Smart, N.G.; Cunningham, J.; et al. Pancreatic Endoderm Derived from Human Embryonic Stem Cells Generates Glucose-Responsive Insulin-Secreting Cells In Vivo. Nat. Biotechnol. 2008, 26, 443–452. [Google Scholar] [CrossRef]
- Cito, M.; Pellegrini, S.; Piemonti, L.; Sordi, V. The Potential and Challenges of Alternative Sources of β Cells for the Cure of Type 1 Diabetes. Endocr. Connect. 2018, 7, R114–R125. [Google Scholar] [CrossRef]
- Suman, S.; Domingues, A.; Ratajczak, J.; Ratajczak, M.Z. Potential Clinical Applications of Stem Cells in Regenerative Medicine. Adv. Exp. Med. Biol. 2019, 1201, 1–22. [Google Scholar] [CrossRef]
- Hyun, I. The Bioethics of Stem Cell Research and Therapy. J. Clin. Investig. 2010, 120, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Bhonde, R.R.; Sheshadri, P.; Sharma, S.; Kumar, A. Making Surrogate β-Cells from Mesenchymal Stromal Cells: Perspectives and Future Endeavors. Int. J. Biochem. Cell Biol. 2014, 46, 90–102. [Google Scholar] [CrossRef]
- Murray, H.E.; Zafar, A.; Qureshi, K.M.; Paget, M.B.; Bailey, C.J.; Downing, R. The Potential Role of Multifunctional Human Amniotic Epithelial Cells in Pancreatic Islet Transplantation. J. Tissue Eng. Regen. Med. 2021, 15, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Parolini, O.; Alviano, F.; Bagnara, G.P.; Bilic, G.; Bühring, H.-J.; Evangelista, M.; Hennerbichler, S.; Liu, B.; Magatti, M.; Mao, N.; et al. Concise Review: Isolation and Characterization of Cells from Human Term Placenta: Outcome of the First International Workshop on Placenta Derived Stem Cells. Stem Cells 2008, 26, 300–311. [Google Scholar] [CrossRef]
- Silini, A.R.; Masserdotti, A.; Papait, A.; Parolini, O. Shaping the Future of Perinatal Cells: Lessons from the Past and Interpretations of the Present. Front. Bioeng. Biotechnol. 2019, 7, 75. [Google Scholar] [CrossRef]
- Okere, B.; Alviano, F.; Costa, R.; Quaglino, D.; Ricci, F.; Dominici, M.; Paolucci, P.; Bonsi, L.; Iughetti, L. In Vitro Differentiation of Human Amniotic Epithelial Cells into Insulin-Producing 3D Spheroids. Int. J. Immunopathol. Pharm. 2015, 28, 390–402. [Google Scholar] [CrossRef] [PubMed]
- Silini, A.R.; Di Pietro, R.; Lang-Olip, I.; Alviano, F.; Banerjee, A.; Basile, M.; Borutinskaite, V.; Eissner, G.; Gellhaus, A.; Giebel, B.; et al. Perinatal Derivatives: Where Do We Stand? A Roadmap of the Human Placenta and Consensus for Tissue and Cell Nomenclature. Front. Bioeng. Biotechnol. 2020, 8, 1438. [Google Scholar] [CrossRef]
- Cross, J.C. Formation of the Placenta and Extraembryonic Membranes. Ann. N. Y. Acad. Sci. 1998, 857, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Bourne, G.L. The Microscopic Anatomy of the Human Amnion and Chorion. Am. J. Obstet. Gynecol. 1960, 79, 1070–1073. [Google Scholar] [CrossRef]
- Alviano, F.; Fossati, V.; Marchionni, C.; Arpinati, M.; Bonsi, L.; Franchina, M.; Lanzoni, G.; Cantoni, S.; Cavallini, C.; Bianchi, F.; et al. Term Amniotic Membrane Is a High Throughput Source for Multipotent Mesenchymal Stem Cells with the Ability to Differentiate into Endothelial Cells In Vitro. BMC Dev. Biol. 2007, 7, 11. [Google Scholar] [CrossRef]
- Stefańska, K.; Ożegowska, K.; Hutchings, G.; Popis, M.; Moncrieff, L.; Dompe, C.; Janowicz, K.; Pieńkowski, W.; Gutaj, P.; Shibli, J.A.; et al. Human Wharton’s Jelly—Cellular Specificity, Stemness Potency, Animal Models, and Current Application in Human Clinical Trials. J. Clin. Med. 2020, 9, 1102. [Google Scholar] [CrossRef]
- McElreavey, K.D.; Irvine, A.I.; Ennis, K.T.; McLean, W.H. Isolation, Culture and Characterisation of Fibroblast-like Cells Derived from the Wharton’s Jelly Portion of Human Umbilical Cord. Biochem. Soc. Trans. 1991, 19, 29S. [Google Scholar] [CrossRef]
- Magatti, M.; Vertua, E.; Cargnoni, A.; Silini, A.; Parolini, O. The Immunomodulatory Properties of Amniotic Cells: The Two Sides of the Coin. Cell Transpl. 2018, 27, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Motedayyen, H.; Esmaeil, N.; Tajik, N.; Khadem, F.; Ghotloo, S.; Khani, B.; Rezaei, A. Method and Key Points for Isolation of Human Amniotic Epithelial Cells with High Yield, Viability and Purity. BMC Res. Notes 2017, 10, 552. [Google Scholar] [CrossRef]
- Trosan, P.; Smeringaiova, I.; Brejchova, K.; Bednar, J.; Benada, O.; Kofronova, O.; Jirsova, K. The Enzymatic De-Epithelialization Technique Determines Denuded Amniotic Membrane Integrity and Viability of Harvested Epithelial Cells. PLoS ONE 2018, 13, e0194820. [Google Scholar] [CrossRef]
- Centurione, L.; Passaretta, F.; Centurione, M.A.; De Munari, S.; Vertua, E.; Silini, A.; Liberati, M.; Parolini, O.; Di Pietro, R. Mapping of the Human Placenta. Cell Transpl. 2018, 27, 12–22. [Google Scholar] [CrossRef]
- Fatimah, S.S.; Tan, G.C.; Chua, K.H.; Tan, A.E.; Hayati, A.R. Effects of Epidermal Growth Factor on the Proliferation and Cell Cycle Regulation of Cultured Human Amnion Epithelial Cells. J. Biosci. Bioeng. 2012, 114, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Pratama, G.; Vaghjiani, V.; Tee, J.Y.; Liu, Y.H.; Chan, J.; Tan, C.; Murthi, P.; Gargett, C.; Manuelpillai, U. Changes in Culture Expanded Human Amniotic Epithelial Cells: Implications for Potential Therapeutic Applications. PLoS ONE 2011, 6, e26136. [Google Scholar] [CrossRef]
- Miki, T.; Lehmann, T.; Cai, H.; Stolz, D.B.; Strom, S.C. Stem Cell Characteristics of Amniotic Epithelial Cells. Stem Cells 2005, 23, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Koike, N.; Sugimoto, J.; Okabe, M.; Arai, K.; Nogami, M.; Okudera, H.; Yoshida, T. Distribution of Amniotic Stem Cells in Human Term Amnion Membrane. Microscopy 2022, 71, 66–76. [Google Scholar] [CrossRef]
- Tabatabaei, M.; Mosaffa, N.; Nikoo, S.; Bozorgmehr, M.; Ghods, R.; Kazemnejad, S.; Rezania, S.; Keshavarzi, B.; Arefi, S.; Ramezani-Tehrani, F.; et al. Isolation and Partial Characterization of Human Amniotic Epithelial Cells: The Effect of Trypsin. Avicenna J. Med. Biotechnol. 2014, 6, 10. [Google Scholar]
- Miki, T. Stem Cell Characteristics and the Therapeutic Potential of Amniotic Epithelial Cells. Am. J. Reprod. Immunol. 2018, 80, e13003. [Google Scholar] [CrossRef]
- Jiawen, S.; Jianjun, Z.; Jiewen, D.; Dedong, Y.; Hongbo, Y.; Jun, S.; Xudong, W.; Shen, S.G.F.; Lihe, G. Osteogenic Differentiation of Human Amniotic Epithelial Cells and Its Application in Alveolar Defect Restoration. Steam Cells Transl. Med. 2014, 3, 1504–1513. [Google Scholar] [CrossRef]
- Zhang, Q.; Lai, D. Application of Human Amniotic Epithelial Cells in Regenerative Medicine: A Systematic Review. Stem Cell Res. Ther. 2020, 11, 439. [Google Scholar] [CrossRef] [PubMed]
- Marongiu, F.; Gramignoli, R.; Dorko, K.; Miki, T.; Ranade, A.R.; Paola Serra, M.; Doratiotto, S.; Sini, M.; Sharma, S.; Mitamura, K.; et al. Hepatic Differentiation of Amniotic Epithelial Cells. Hepatology 2011, 53, 1719–1729. [Google Scholar] [CrossRef]
- Yao, M.; Chen, J.; Yang, X.-X.; Zhang, X.-L.; Ji, Q.-S.; Zhou, Q.; Xu, J.-T. Differentiation of Human Amniotic Epithelial Cells into Corneal Epithelial-like Cells in Vitro. Int. J. Ophthalmol. 2013, 6, 564–572. [Google Scholar] [CrossRef]
- Niknejad, H.; Peirovi, H.; Ahmadiani, A.; Ghanavi, J.; Jorjani, M. Differentiation Factors That Influence Neuronal Markers Expression in Vitro from Human Amniotic Epithelial Cells. Eur. Cell Mater. 2010, 19, 22–29. [Google Scholar] [CrossRef]
- Bhandari, D.R.; Seo, K.-W.; Sun, B.; Seo, M.-S.; Kim, H.-S.; Seo, Y.-J.; Marcin, J.; Forraz, N.; Roy, H.L.; Larry, D.; et al. The Simplest Method for in Vitro β-Cell Production from Human Adult Stem Cells. Differentiation 2011, 82, 144–152. [Google Scholar] [CrossRef]
- Wei, J.P.; Zhang, T.S.; Kawa, S.; Aizawa, T.; Ota, M.; Akaike, T.; Kato, K.; Konishi, I.; Nikaido, T. Human Amnion-Isolated Cells Normalize Blood Glucose in Streptozotocin-Induced Diabetic Mice. Cell Transpl. 2003, 12, 545–552. [Google Scholar] [CrossRef]
- Hou, Y.; Huang, Q.; Liu, T.; Guo, L. Human Amnion Epithelial Cells Can Be Induced to Differentiate into Functional Insulin-Producing Cells. Acta Biochim. Biophys. Sin. 2008, 40, 830–839. [Google Scholar] [CrossRef][Green Version]
- Liu, X.; Qin, J.; Chang, M.; Wang, S.; Li, Y.; Pei, X.; Wang, Y. Enhanced Differentiation of Human Pluripotent Stem Cells into Pancreatic Endocrine Cells in 3D Culture by Inhibition of Focal Adhesion Kinase. Stem Cell Res. Ther. 2020, 11, 488. [Google Scholar] [CrossRef]
- D’Amour, K.A.; Bang, A.G.; Eliazer, S.; Kelly, O.G.; Agulnick, A.D.; Smart, N.G.; Moorman, M.A.; Kroon, E.; Carpenter, M.K.; Baetge, E.E. Production of Pancreatic Hormone–Expressing Endocrine Cells from Human Embryonic Stem Cells. Nat. Biotechnol. 2006, 24, 1392–1401. [Google Scholar] [CrossRef]
- Luo, Y.; Cheng, Y.-W.; Yu, C.-Y.; Liu, R.-M.; Zhao, Y.-J.; Chen, D.-X.; Zhong, J.-J.; Xiao, J.-H. Effects of Hyaluronic Acid on Differentiation of Human Amniotic Epithelial Cells and Cell-Replacement Therapy in Type 1 Diabetic Mice. Exp. Cell Res. 2019, 384, 111642. [Google Scholar] [CrossRef]
- Jensen, C.; Teng, Y. Is It Time to Start Transitioning From 2D to 3D Cell Culture? Front. Mol. Biosci. 2020, 7, 33. [Google Scholar] [CrossRef]
- Lebreton, F.; Lavallard, V.; Bellofatto, K.; Bonnet, R.; Wassmer, C.H.; Perez, L.; Kalandadze, V.; Follenzi, A.; Boulvain, M.; Kerr-Conte, J.; et al. Insulin-Producing Organoids Engineered from Islet and Amniotic Epithelial Cells to Treat Diabetes. Nat. Commun. 2019, 10, 4491. [Google Scholar] [CrossRef]
- Lebreton, F.; Bellofatto, K.; Wassmer, C.H.; Perez, L.; Lavallard, V.; Parnaud, G.; Cottet-Dumoulin, D.; Kerr-Conte, J.; Pattou, F.; Bosco, D.; et al. Shielding Islets with Human Amniotic Epithelial Cells Enhances Islet Engraftment and Revascularization in a Murine Diabetes Model. Am. J. Transplant. 2020, 20, 1551–1561. [Google Scholar] [CrossRef]
- Wassmer, C.-H.; Lebreton, F.; Bellofatto, K.; Perez, L.; Cottet-Dumoulin, D.; Andres, A.; Bosco, D.; Berney, T.; Othenin-Girard, V.; Martinez De Tejada, B.; et al. Bio-Engineering of Pre-Vascularized Islet Organoids for the Treatment of Type 1 Diabetes. Transpl. Int. 2022, 35, 10214. [Google Scholar] [CrossRef]
- Jiang, W.; Xu, J. Immune Modulation by Mesenchymal Stem Cells. Cell Prolif. 2020, 53, e12712. [Google Scholar] [CrossRef]
- Parolini, O.; Caruso, M. Review: Preclinical Studies on Placenta-Derived Cells and Amniotic Membrane: An Update. Placenta 2011, 32 (Suppl. S2), S186–S195. [Google Scholar] [CrossRef]
- Wassmer, C.-H.; Lebreton, F.; Bellofatto, K.; Bosco, D.; Berney, T.; Berishvili, E. Generation of Insulin-Secreting Organoids: A Step toward Engineering and Transplanting the Bioartificial Pancreas. Transpl Int. 2020, 33, 1577–1588. [Google Scholar] [CrossRef]
- Moodley, Y.; Ilancheran, S.; Samuel, C.; Vaghjiani, V.; Atienza, D.; Williams, E.D.; Jenkin, G.; Wallace, E.; Trounson, A.; Manuelpillai, U. Human Amnion Epithelial Cell Transplantation Abrogates Lung Fibrosis and Augments Repair. Am. J. Respir. Crit. Care Med. 2010, 182, 643–651. [Google Scholar] [CrossRef]
- Ilancheran, S.; Michalska, A.; Peh, G.; Wallace, E.M.; Pera, M.; Manuelpillai, U. Stem Cells Derived from Human Fetal Membranes Display Multilineage Differentiation Potential. Biol. Reprod. 2007, 77, 577–588. [Google Scholar] [CrossRef]
- Morandi, F.; Marimpietri, D.; Görgens, A.; Gallo, A.; Srinivasan, R.C.; El-Andaloussi, S.; Gramignoli, R. Human Amnion Epithelial Cells Impair T Cell Proliferation: The Role of HLA-G and HLA-E Molecules. Cells 2020, 9, 2123. [Google Scholar] [CrossRef]
- Braud, V.M.; Allan, D.S.; McMichael, A.J. Functions of Nonclassical MHC and Non-MHC-Encoded Class I Molecules. Curr. Opin. Immunol. 1999, 11, 100–108. [Google Scholar] [CrossRef]
- Rizzo, R.; Campioni, D.; Stignani, M.; Melchiorri, L.; Bagnara, G.P.; Bonsi, L.; Alviano, F.; Lanzoni, G.; Moretti, S.; Cuneo, A.; et al. A Functional Role for Soluble HLA-G Antigens in Immune Modulation Mediated by Mesenchymal Stromal Cells. Cytotherapy 2008, 10, 364–375. [Google Scholar] [CrossRef]
- Zhu, D.; Tan, J.; Maleken, A.S.; Muljadi, R.; Chan, S.T.; Lau, S.N.; Elgass, K.; Leaw, B.; Mockler, J.; Chambers, D.; et al. Human Amnion Cells Reverse Acute and Chronic Pulmonary Damage in Experimental Neonatal Lung Injury. Stem Cell Res Ther. 2017, 8, 257. [Google Scholar] [CrossRef]
- Li, H.; Niederkorn, J.Y.; Neelam, S.; Mayhew, E.; Word, R.A.; McCulley, J.P.; Alizadeh, H. Immunosuppressive Factors Secreted by Human Amniotic Epithelial Cells. Investig. Ophthalmol. Vis. Sci. 2005, 46, 900–907. [Google Scholar] [CrossRef]
- Tan, J.L.; Chan, S.T.; Lo, C.Y.; Deane, J.A.; McDonald, C.A.; Bernard, C.C.; Wallace, E.M.; Lim, R. Amnion Cell-Mediated Immune Modulation Following Bleomycin Challenge: Controlling the Regulatory T Cell Response. Stem Cell Res. Ther. 2015, 6, 8. [Google Scholar] [CrossRef]
- Fathi, I.; Miki, T. Human Amniotic Epithelial Cells Secretome: Components, Bioactivity, and Challenges. Front. Med. 2022, 8. [Google Scholar] [CrossRef]
- Tan, J.L.; Lau, S.N.; Leaw, B.; Nguyen, H.P.T.; Salamonsen, L.A.; Saad, M.I.; Chan, S.T.; Zhu, D.; Krause, M.; Kim, C.; et al. Amnion Epithelial Cell-Derived Exosomes Restrict Lung Injury and Enhance Endogenous Lung Repair. Stem Cells Transl. Med. 2018, 7, 180–196. [Google Scholar] [CrossRef]
- Cargnoni, A.; Farigu, S.; Cotti Piccinelli, E.; Bonassi Signoroni, P.; Romele, P.; Vanosi, G.; Toschi, I.; Cesari, V.; Barros Sant’Anna, L.; Magatti, M.; et al. Effect of Human Amniotic Epithelial Cells on Pro-Fibrogenic Resident Hepatic Cells in a Rat Model of Liver Fibrosis. J. Cell Mol. Med. 2018, 22, 1202–1213. [Google Scholar] [CrossRef]
- Song, Y.-S.; Joo, H.-W.; Park, I.-H.; Shen, G.-Y.; Lee, Y.; Shin, J.H.; Kim, H.; Shin, I.-S.; Kim, K.-S. Transplanted Human Amniotic Epithelial Cells Secrete Paracrine Proangiogenic Cytokines in Rat Model of Myocardial Infarctio. Cell Transpl. 2015, 24, 2055–2064. [Google Scholar] [CrossRef]
- Vosdoganes, P.; Wallace, E.M.; Chan, S.T.; Acharya, R.; Moss, T.J.M.; Lim, R. Human Amnion Epithelial Cells Repair Established Lung Injury. 2013. Available online: https://journals.sagepub.com/doi/10.3727/096368912X657657 (accessed on 25 September 2022).
- Hodge, A.; Andrewartha, N.; Lourensz, D.; Strauss, R.; Correia, J.; Goonetilleke, M.; Yeoh, G.; Lim, R.; Sievert, W. Human Amnion Epithelial Cells Produce Soluble Factors That Enhance Liver Repair by Reducing Fibrosis While Maintaining Regeneration in a Model of Chronic Liver Injury. 2020. Available online: https://journals.sagepub.com/doi/10.1177/0963689720950221 (accessed on 25 September 2022).
- Jirsova, K.; Jones, G.L.A. Amniotic Membrane in Ophthalmology: Properties, Preparation, Storage and Indications for Grafting—A Review. Cell Tissue Bank 2017, 18, 193–204. [Google Scholar] [CrossRef]
- Castillo-Melendez, M.; Yawno, T.; Jenkin, G.; Miller, S. Stem Cell Therapy to Protect and Repair the Developing Brain: A Review of Mechanisms of Action of Cord Blood and Amnion Epithelial Derived Cells. Front. Neurosci. 2013, 7, 194. [Google Scholar] [CrossRef]
- Podestà, M.A.; Remuzzi, G.; Casiraghi, F. Mesenchymal Stromal Cells for Transplant Tolerance. Front. Immunol. 2019, 10, 1287. [Google Scholar] [CrossRef]
- Jun, Y.; Kang, A.R.; Lee, J.S.; Park, S.-J.; Lee, D.Y.; Moon, S.-H.; Lee, S.-H. Microchip-Based Engineering of Super-Pancreatic Islets Supported by Adipose-Derived Stem Cells. Biomaterials 2014, 35, 4815–4826. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, K.M.; Oliver, R.J.; Paget, M.B.; Murray, H.E.; Bailey, C.J.; Downing, R. Human Amniotic Epithelial Cells Induce Localized Cell-Mediated Immune Privilege in Vitro: Implications for Pancreatic Islet Transplantation. Cell Transpl. 2011, 20, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Zafar, A.; Lee, J.; Yesmin, S.; Paget, M.B.; Bailey, C.J.; Murray, H.E.; Downing, R. Rotational Culture and Integration with Amniotic Stem Cells Reduce Porcine Islet Immunoreactivity in Vitro and Slow Xeno-Rejection in a Murine Model of Islet Transplantation. Xenotransplantation 2019, 26, e12508. [Google Scholar] [CrossRef]
- Cui, W.; Khan, K.M.; Ma, X.; Chen, G.; Desai, C.S. Human Amniotic Epithelial Cells and Human Amniotic Membrane as a Vehicle for Islet Cell Transplantation. Transpl. Proc. 2020, 52, 982–986. [Google Scholar] [CrossRef] [PubMed]
- Lebreton, F.; Hanna, R.; Wassmer, C.H.; Bellofatto, K.; Perez, L.; Othenin-Girard, V.; de Tejada, B.M.; Cohen, M.; Berishvili, E. Mechanisms of Immunomodulation and Cytoprotection Conferred to Pancreatic Islet by Human Amniotic Epithelial Cells. Stem Cell Rev. Rep. 2022, 18, 346–359. [Google Scholar] [CrossRef]
- Kamal, M.M.; Kassem, D.H. Therapeutic Potential of Wharton’s Jelly Mesenchymal Stem Cells for Diabetes: Achievements and Challenges. Front. Cell Dev. Biol. 2020, 8, 16. [Google Scholar] [CrossRef] [PubMed]
- Bongso, A.; Fong, C.-Y.; Gauthaman, K. Taking Stem Cells to the Clinic: Major Challenges. J. Cell Biochem. Suppl. 2008, 105, 1352–1360. [Google Scholar] [CrossRef]
- Bai, C.; Gao, Y.; Li, Q.; Feng, Y.; Yu, Y.; Meng, G.; Zhang, M.; Guan, W. Differentiation of Chicken Umbilical Cord Mesenchymal Stem Cells into Beta-like Pancreatic Islet Cells. Artif. Cells Nanomed. Biotechnol. 2015, 43, 106–111. [Google Scholar] [CrossRef]
- Weiss, M.L.; Anderson, C.; Medicetty, S.; Seshareddy, K.B.; Weiss, R.J.; VanderWerff, I.; Troyer, D.; McIntosh, K.R. Immune Properties of Human Umbilical Cord Wharton’s Jelly-Derived Cells. Stem Cells 2008, 26, 2865–2874. [Google Scholar] [CrossRef]
- El Omar, R.; Beroud, J.; Stoltz, J.-F.; Menu, P.; Velot, E.; Decot, V. Umbilical Cord Mesenchymal Stem Cells: The New Gold Standard for Mesenchymal Stem Cell-Based Therapies? Tissue Eng. Part B Rev. 2014, 20, 523–544. [Google Scholar] [CrossRef]
- La Rocca, G.; Anzalone, R.; Corrao, S.; Magno, F.; Loria, T.; Lo Iacono, M.; Di Stefano, A.; Giannuzzi, P.; Marasà, L.; Cappello, F.; et al. Isolation and Characterization of Oct-4+/HLA-G+ Mesenchymal Stem Cells from Human Umbilical Cord Matrix: Differentiation Potential and Detection of New Markers. Histochem. Cell Biol. 2009, 131, 267–282. [Google Scholar] [CrossRef] [PubMed]
- Anzalone, R.; Lo Iacono, M.; Loria, T.; Di Stefano, A.; Giannuzzi, P.; Farina, F.; La Rocca, G. Wharton’s Jelly Mesenchymal Stem Cells as Candidates for Beta Cells Regeneration: Extending the Differentiative and Immunomodulatory Benefits of Adult Mesenchymal Stem Cells for the Treatment of Type 1 Diabetes. Stem Cell Rev. Rep. 2011, 7, 342–363. [Google Scholar] [CrossRef]
- Huang, Q.; Huang, Y.; Liu, J. Mesenchymal Stem Cells: An Excellent Candidate for the Treatment of Diabetes Mellitus. Int. J. Endocrinol. 2021, 2021, 9938658. [Google Scholar] [CrossRef]
- Johnson, C.L.; Soeder, Y.; Dahlke, M.H. Concise Review: Mesenchymal Stromal Cell-Based Approaches for the Treatment of Acute Respiratory Distress and Sepsis Syndromes. Stem Cells Transl. Med. 2017, 6, 1141–1151. [Google Scholar] [CrossRef]
- Xu, Z.; Huang, Y.; Zhou, J.; Deng, X.; He, W.; Liu, X.; Li, Y.; Zhong, N.; Sang, L. Current Status of Cell-Based Therapies for COVID-19: Evidence From Mesenchymal Stromal Cells in Sepsis and ARDS. Front. Immunol. 2021, 12, 738697. [Google Scholar] [CrossRef] [PubMed]
- Gorman, E.; Millar, J.; McAuley, D.; O’Kane, C. Mesenchymal Stromal Cells for Acute Respiratory Distress Syndrome (ARDS), Sepsis, and COVID-19 Infection: Optimizing the Therapeutic Potential. Expert Rev. Respir. Med. 2021, 15, 301–324. [Google Scholar] [CrossRef] [PubMed]
- Lanzoni, G.; Linetsky, E.; Correa, D.; Messinger Cayetano, S.; Alvarez, R.A.; Kouroupis, D.; Alvarez Gil, A.; Poggioli, R.; Ruiz, P.; Marttos, A.C.; et al. Umbilical Cord Mesenchymal Stem Cells for COVID-19 Acute Respiratory Distress Syndrome: A Double-Blind, Phase 1/2a, Randomized Controlled Trial. Stem Cells Transl. Med. 2021, 10, 660–673. [Google Scholar] [CrossRef]
- Kouroupis, D.; Lanzoni, G.; Linetsky, E.; Messinger Cayetano, S.; Wishnek Metalonis, S.; Leñero, C.; Stone, L.D.; Ruiz, P.; Correa, D.; Ricordi, C. Umbilical Cord-Derived Mesenchymal Stem Cells Modulate TNF and Soluble TNF Receptor 2 (STNFR2) in COVID-19 ARDS Patients. Eur. Rev. Med. Pharm. Sci. 2021, 25, 4435–4438. [Google Scholar] [CrossRef]
- Ciccocioppo, R.; Gibellini, D.; Astori, G.; Bernardi, M.; Bozza, A.; Chieregato, K.; Elice, F.; Ugel, S.; Caligola, S.; de Sanctis, F.; et al. The Immune Modulatory Effects of Umbilical Cord-Derived Mesenchymal Stromal Cells in Severe COVID-19 Pneumonia. Stem Cell Res. Ther. 2021, 12, 1–10. [Google Scholar] [CrossRef]
- Cai, J.; Wu, Z.; Xu, X.; Liao, L.; Chen, J.; Huang, L.; Wu, W.; Luo, F.; Wu, C.; Pugliese, A.; et al. Umbilical Cord Mesenchymal Stromal Cell With Autologous Bone Marrow Cell Transplantation in Established Type 1 Diabetes: A Pilot Randomized Controlled Open-Label Clinical Study to Assess Safety and Impact on Insulin Secretion. Diabetes Care 2016, 39, 149–157. [Google Scholar] [CrossRef]
- Hu, J.; Yu, X.; Wang, Z.; Wang, F.; Wang, L.; Gao, H.; Chen, Y.; Zhao, W.; Jia, Z.; Yan, S.; et al. Long Term Effects of the Implantation of Wharton’s Jelly-Derived Mesenchymal Stem Cells from the Umbilical Cord for Newly-Onset Type 1 Diabetes Mellitus. Endocr. J. 2013, 60, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, F.; Liang, H.; Tang, D.; Huang, M.; Zhao, J.; Yang, X.; Liu, Y.; Shu, L.; Wang, J.; et al. Efficacy of Mesenchymal Stem Cell Transplantation Therapy for Type 1 and Type 2 Diabetes Mellitus: A Meta-Analysis. Stem Cell Res. Ther. 2021, 12, 273. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Shen, S.-M.; Ling, Q.; Wang, B.; Li, L.-R.; Zhang, W.; Qu, D.-D.; Bi, Y.; Zhu, D.-L. One Repeated Transplantation of Allogeneic Umbilical Cord Mesenchymal Stromal Cells in Type 1 Diabetes: An Open Parallel Controlled Clinical Study. Stem Cell Res. 2021, 12, 340. [Google Scholar] [CrossRef] [PubMed]
- Nilforoushzadeh, M.A.; Afzali, H.; Raoofi, A.; Nouri, M.; Naser, R.; Gholami, O.; Nasiry, D.; Mohammadnia, A.; Razzaghi, Z.; Alimohammadi, A.; et al. Topical Spray of Wharton’s Jelly Mesenchymal Stem Cells Derived from Umbilical Cord Accelerates Diabetic Wound Healing. J. Cosmet. Derm. 2022, 10, 5156–5167. [Google Scholar] [CrossRef] [PubMed]
- El-Demerdash, R.F.; Hammad, L.N.; Kamal, M.M.; El Mesallamy, H.O. A Comparison of Wharton’s Jelly and Cord Blood as a Source of Mesenchymal Stem Cells for Diabetes Cell Therapy. Regen. Med. 2015, 10, 841–855. [Google Scholar] [CrossRef] [PubMed]
- Kassem, D.H.; Kamal, M.M. Therapeutic Efficacy of Umbilical Cord-Derived Stem Cells for Diabetes Mellitus: A Meta-Analysis Study. Stem Cell Res. 2020, 11, 484. [Google Scholar] [CrossRef] [PubMed]
- Tsai, P.-J.; Wang, H.-S.; Lin, G.-J.; Chou, S.-C.; Chu, T.-H.; Chuan, W.-T.; Lu, Y.-J.; Weng, Z.-C.; Su, C.-H.; Hsieh, P.-S.; et al. Undifferentiated Wharton’s Jelly Mesenchymal Stem Cell Transplantation Induces Insulin-Producing Cell Differentiation and Suppression of T-Cell-Mediated Autoimmunity in Nonobese Diabetic Mice. Cell Transpl. 2015, 24, 1555–1570. [Google Scholar] [CrossRef]
- Kassem, D.H.; Kamal, M.M.; El-Kholy, A.E.-L.G.; El-Mesallamy, H.O. Association of Expression Levels of Pluripotency/Stem Cell Markers with the Differentiation Outcome of Wharton’s Jelly Mesenchymal Stem Cells into Insulin Producing Cells. Biochimie 2016, 127, 187–195. [Google Scholar] [CrossRef]
- Kassem, D.H.; Kamal, M.M.; El-Kholy, A.E.-L.G.; El-Mesallamy, H.O. Exendin-4 Enhances the Differentiation of Wharton’s Jelly Mesenchymal Stem Cells into Insulin-Producing Cells through Activation of Various β-Cell Markers. Stem Cell Res. 2016, 7, 108. [Google Scholar] [CrossRef]
- Ranjbaran, H.; Abediankenari, S.; Khalilian, A.; Rahmani, Z.; Momeninezhad Amiri, M.; Hosseini Khah, Z. Differentiation of Wharton’s Jelly Derived Mesenchymal Stem Cells into Insulin Producing Cells. Int. J. Hematol Oncol. Stem Cell Res. 2018, 12, 220–229. [Google Scholar]
- Ren, H.; Sang, Y.; Zhang, F.; Liu, Z.; Qi, N.; Chen, Y. Comparative Analysis of Human Mesenchymal Stem Cells from Umbilical Cord, Dental Pulp, and Menstrual Blood as Sources for Cell Therapy. Stem Cells Int. 2016, 2016, 3516574. [Google Scholar] [CrossRef]
- Gao, L.R.; Zhang, N.K.; Zhang, Y.; Chen, Y.; Wang, L.; Zhu, Y.; Tang, H.H. Overexpression of Apelin in Wharton’ Jelly Mesenchymal Stem Cell Reverses Insulin Resistance and Promotes Pancreatic β Cell Proliferation in Type 2 Diabetic Rats. Stem Cell Res. 2018, 9, 339. [Google Scholar] [CrossRef] [PubMed]
- Beikmohammadi, L.; Bandehpour, M.; Hashemi, S.M.; Kazemi, B. Generation of Insulin-Producing Hepatocyte-like Cells from Human Wharton’s Jelly Mesenchymal Stem Cells as an Alternative Source of Islet Cells. J. Cell Physiol. 2019, 234, 17326–17336. [Google Scholar] [CrossRef]
- Hsiao, C.-Y.; Chen, T.-H.; Huang, B.-S.; Chen, P.-H.; Su, C.-H.; Shyu, J.-F.; Tsai, P.-J. Comparison between the Therapeutic Effects of Differentiated and Undifferentiated Wharton’s Jelly Mesenchymal Stem Cells in Rats with Streptozotocin-Induced Diabetes. World J. Stem Cells 2020, 12, 139–151. [Google Scholar] [CrossRef]
- Aierken, A.; Li, B.; Liu, P.; Cheng, X.; Kou, Z.; Tan, N.; Zhang, M.; Yu, S.; Shen, Q.; Du, X.; et al. Melatonin Treatment Improves Human Umbilical Cord Mesenchymal Stem Cell Therapy in a Mouse Model of Type II Diabetes Mellitus via the PI3K/AKT Signaling Pathway. Stem Cell Res. 2022, 13, 164. [Google Scholar] [CrossRef] [PubMed]
- Azarbarz, N.; Khorsandi, L.; Nejaddehbashi, F.; Neisi, N.; Nejad, D.B. Decellularized Wharton’s Jelly Scaffold Enhances Differentiation of Mesenchymal Stem Cells to Insulin-Secreting Cells. Tissue Cell. 2022, 79, 101938. [Google Scholar] [CrossRef] [PubMed]
- Waterman, R.S.; Tomchuck, S.L.; Henkle, S.L.; Betancourt, A.M. A New Mesenchymal Stem Cell (MSC) Paradigm: Polarization into a Pro-Inflammatory MSC1 or an Immunosuppressive MSC2 Phenotype. PLoS ONE 2010, 5, e10088. [Google Scholar] [CrossRef]
- Marrazzo, P.; Crupi, A.N.; Alviano, F.; Teodori, L.; Bonsi, L. Exploring the Roles of MSCs in Infections: Focus on Bacterial Diseases. J. Mol. Med. 2019, 97, 437–450. [Google Scholar] [CrossRef]
- LPS-Preconditioned Mesenchymal Stromal Cells Modify Macrophage Polarization for Resolution of Chronic Inflammation via Exosome-Shuttled Let-7b—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/26386558/ (accessed on 25 September 2022).
- Kouroupis, D.; Correa, D. Increased Mesenchymal Stem Cell Functionalization in Three-Dimensional Manufacturing Settings for Enhanced Therapeutic Applications. Front. Bioeng. Biotechnol. 2021, 9, 621748. [Google Scholar] [CrossRef]
- Xie, A.W.; Zacharias, N.A.; Binder, B.Y.K.; Murphy, W.L. Controlled Aggregation Enhances Immunomodulatory Potential of Mesenchymal Stromal Cell Aggregates. Stem Cells Transl. Med. 2021, 10, 1184–1201. [Google Scholar] [CrossRef]
- Bartosh, T.J.; Ylöstalo, J.H.; Mohammadipoor, A.; Bazhanov, N.; Coble, K.; Claypool, K.; Lee, R.H.; Choi, H.; Prockop, D.J. Aggregation of Human Mesenchymal Stromal Cells (MSCs) into 3D Spheroids Enhances Their Antiinflammatory Properties. Proc. Natl. Acad. Sci. USA 2010, 107, 13724–13729. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Sun, L.; Jeske, R.; Nkosi, D.; York, S.B.; Liu, Y.; Grant, S.C.; Meckes, D.G.; Li, Y. Engineering Extracellular Vesicles by Three-Dimensional Dynamic Culture of Human Mesenchymal Stem Cells. J. Extracell Vesicles 2022, 11, e12235. [Google Scholar] [CrossRef] [PubMed]
- Seyedi, F.; Farsinejad, A.; Nematollahi-Mahani, S.A.; Eslaminejad, T.; Nematollahi-Mahani, S.N. Suspension Culture Alters Insulin Secretion in Induced Human Umbilical Cord Matrix-Derived Mesenchymal Cells. Cell J. 2016, 18, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Chandravanshi, B.; Datar, S.; Bhonde, R. Response of Chick B Islets to Insulin Secretagogues Is Comparable to Those of Human Islet Equivalents. JOP 2015, 16, 318–323. [Google Scholar] [CrossRef]
- Xiang, C.; Xie, Q.-P. Protection of Mouse Pancreatic Islet Function by Co-culture with Hypoxia Pre-treated Mesenchymal Stromal Cells. Mol. Med. Rep. 2018, 18, 2589–2598. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Zhang, L.; Yang, L.; Wang, X.; Zhao, C.; Zhao, D. Protective Effect of Mesenchymal Stem Cells on Isolated Islets Survival and Against Hypoxia Associated With the HIF-1α/PFKFB3 Pathway. Cell Transpl. 2022, 31, 09636897211073127. [Google Scholar] [CrossRef] [PubMed]
- Nie, W.; Ma, X.; Yang, C.; Chen, Z.; Rong, P.; Wu, M.; Jiang, J.; Tan, M.; Yi, S.; Wang, W. Human Mesenchymal-Stem-Cells-Derived Exosomes Are Important in Enhancing Porcine Islet Resistance to Hypoxia. Xenotransplantation 2018, 25, e12405. [Google Scholar] [CrossRef]
- Wang, G.; Li, Y.; Wang, Y.; Dong, Y.; Wang, F.-S.; Ding, Y.; Kang, Y.; Xu, X. Roles of the Co-culture of Human Umbilical Cord Wharton’s Jelly-derived Mesenchymal Stem Cells with Rat Pancreatic Cells in the Treatment of Rats with Diabetes Mellitus. Exp. Med. 2014, 8, 1389–1396. [Google Scholar] [CrossRef][Green Version]
- Dietrich, I.; Girdlestone, J.; Giele, H. Differential Cytokine Expression in Direct and Indirect Co-Culture of Islets and Mesenchymal Stromal Cells. Cytokine 2022, 150, 155779. [Google Scholar] [CrossRef]
- Kim, K.-S.; Choi, Y.K.; Kim, M.J.; Hwang, J.W.; Min, K.; Jung, S.Y.; Kim, S.-K.; Choi, Y.-S.; Cho, Y.-W. Umbilical Cord-Mesenchymal Stem Cell-Conditioned Medium Improves Insulin Resistance in C2C12 Cell. Diabetes Metab. J. 2021, 45, 260–269. [Google Scholar] [CrossRef]
- Wu, Z.; Xu, X.; Cai, J.; Chen, J.; Huang, L.; Wu, W.; Pugliese, A.; Li, S.; Ricordi, C.; Tan, J. Prevention of Chronic Diabetic Complications in Type 1 Diabetes by Co-Transplantation of Umbilical Cord Mesenchymal Stromal Cells and Autologous Bone Marrow: A Pilot Randomized Controlled Open-Label Clinical Study with 8-Year Follow-Up. Cytotherapy 2022, 24, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Moschidou, D.; Mukherjee, S.; Blundell, M.P.; Jones, G.N.; Atala, A.J.; Thrasher, A.J.; Fisk, N.M.; de Coppi, P.; Guillot, P.V. Human Mid-Trimester Amniotic Fluid Stem Cells Cultured Under Embryonic Stem Cell Conditions with Valproic Acid Acquire Pluripotent Characteristics. Stem Cells Dev. 2013, 22, 444–458. [Google Scholar] [CrossRef]
- De Coppi, P.; Bartsch, G.; Siddiqui, M.M.; Xu, T.; Santos, C.C.; Perin, L.; Mostoslavsky, G.; Serre, A.C.; Snyder, E.Y.; Yoo, J.J.; et al. Isolation of Amniotic Stem Cell Lines with Potential for Therapy. Nat. Biotechnol. 2007, 25, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Moorefield, E.C.; McKee, E.E.; Solchaga, L.; Orlando, G.; Yoo, J.J.; Walker, S.; Furth, M.E.; Bishop, C.E. Cloned, CD117 Selected Human Amniotic Fluid Stem Cells Are Capable of Modulating the Immune Response. PLoS ONE 2011, 6, e26535. [Google Scholar] [CrossRef]
- Luo, C.; Jia, W.; Wang, K.; Chi, F.; Gu, Y.; Yan, X.; Zou, G.; Duan, T.; Zhou, Q. Human Amniotic Fluid Stem Cells Suppress PBMC Proliferation through IDO and IL-10-Dependent Pathways. Curr. Stem Cell Res. 2014, 9, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Subhan, B.S.; Kwong, J.; Kuhn, J.F.; Monas, A.; Sharma, S.; Rabbani, P.S. Amniotic Fluid-Derived Multipotent Stromal Cells Drive Diabetic Wound Healing through Modulation of Macrophages. J. Transl. Med. 2021, 19, 16. [Google Scholar] [CrossRef]
- Sato, Y.; Ochiai, D.; Abe, Y.; Masuda, H.; Fukutake, M.; Ikenoue, S.; Kasuga, Y.; Shimoda, M.; Kanai, Y.; Tanaka, M. Prophylactic Therapy with Human Amniotic Fluid Stem Cells Improved Survival in a Rat Model of Lipopolysaccharide-Induced Neonatal Sepsis through Immunomodulation via Aggregates with Peritoneal Macrophages. Stem Cell Res. 2020, 11, 300. [Google Scholar] [CrossRef]
- Villani, V.; Milanesi, A.; Sedrakyan, S.; Da Sacco, S.; Angelow, S.; Conconi, M.T.; Di Liddo, R.; de Filippo, R.; Perin, L. Amniotic Fluid Stem Cells Prevent β-Cell Injury. Cytotherapy 2014, 16, 41–55. [Google Scholar] [CrossRef]
- Pianta, S.; Bonassi Signoroni, P.; Muradore, I.; Rodrigues, M.F.; Rossi, D.; Silini, A.; Parolini, O. Amniotic Membrane Mesenchymal Cells-Derived Factors Skew T Cell Polarization toward Treg and Downregulate Th1 and Th17 Cells Subsets. Stem Cell Rev. Rep. 2015, 11, 394–407. [Google Scholar] [CrossRef]
- Magaña-Guerrero, F.S.; Domínguez-López, A.; Martínez-Aboytes, P.; Buentello-Volante, B.; Garfias, Y. Human Amniotic Membrane Mesenchymal Stem Cells Inhibit Neutrophil Extracellular Traps through TSG-6. Sci. Rep. 2017, 7, 12426. [Google Scholar] [CrossRef]
- Magatti, M.; de Munari, S.; Vertua, E.; Nassauto, C.; Albertini, A.; Wengler, G.S.; Parolini, O. Amniotic Mesenchymal Tissue Cells Inhibit Dendritic Cell Differentiation of Peripheral Blood and Amnion Resident Monocytes. Cell Transpl. 2009, 18, 899–914. [Google Scholar] [CrossRef]
- Kharat, A.; Chandravanshi, B.; Gadre, S.; Patil, V.; Bhonde, R.; Dubhashi, A. IGF-1 and Somatocrinin Trigger Islet Differentiation in Human Amniotic Membrane Derived Mesenchymal Stem Cells. Life Sci. 2019, 216, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Kadam, S.S.; Sudhakar, M.; Nair, P.D.; Bhonde, R.R. Reversal of Experimental Diabetes in Mice by Transplantation of Neo-Islets Generated from Human Amnion-Derived Mesenchymal Stromal Cells Using Immuno-Isolatory Macrocapsules. Cytotherapy 2010, 12, 982–991. [Google Scholar] [CrossRef]
- Li, J.; Koike-Soko, C.; Sugimoto, J.; Yoshida, T.; Okabe, M.; Nikaido, T. Human Amnion-Derived Stem Cells Have Immunosuppressive Properties on NK Cells and Monocytes. Cell Transpl. 2015, 24, 2065–2076. [Google Scholar] [CrossRef]
- Kubo, M.; Sonoda, Y.; Muramatsu, R.; Usui, M. Immunogenicity of Human Amniotic Membrane in Experimental Xenotransplantation. Investig. Ophthalmol. Vis. Sci. 2001, 42, 1539–1546. [Google Scholar]
- Najar, M.; Fayyad-Kazan, M.; Meuleman, N.; Bron, D.; Fayyad-Kazan, H.; Lagneaux, L. Immunological Impact of Wharton’s Jelly Mesenchymal Stromal Cells and Natural Killer Cell Co-Culture. Mol. Cell Biochem. 2018, 447, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Al-Mrahleh, M.; Matar, S.; Jafar, H.; Wehaibi, S.; Aslam, N.; Awidi, A. Human Wharton’s Jelly-Derived Mesenchymal Stromal Cells Primed by Tumor Necrosis Factor-α and Interferon-γ Modulate the Innate and Adaptive Immune Cells of Type 1 Diabetic Patients. Front. Immunol. 2021, 12, 732549. [Google Scholar] [CrossRef]
- Maraldi, T.; Beretti, F.; Guida, M.; Zavatti, M.; de Pol, A. Role of Hepatocyte Growth Factor in the Immunomodulation Potential of Amniotic Fluid Stem Cells. Stem Cells Transl. Med. 2015, 4, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Full Article: Comparative Analysis of Human Mesenchymal Stem Cells from Fetal-Bone Marrow, Adipose Tissue, and Warton’s Jelly as Sources of Cell Immunomodulatory Therapy. Available online: https://www.tandfonline.com/doi/full/10.1080/21645515.2015.1030549 (accessed on 26 September 2022).
- Alipour, R.; Motedayyen, H.; Sereshki, N.; Rafiee, M.; Alsahebfosul, F.; Pourazar, A. Human Amniotic Epithelial Cells Affect the Functions of Neutrophils. Int. J. Stem Cells 2020, 13, 212–220. [Google Scholar] [CrossRef]
- Motedayyen, H.; Zarnani, A.-H.; Tajik, N.; Ghotloo, S.; Rezaei, A. Immunomodulatory Effects of Human Amniotic Epithelial Cells on Naive CD4+ T Cells from Women with Unexplained Recurrent Spontaneous Abortion. Placenta 2018, 71, 31–40. [Google Scholar] [CrossRef]
- Zheng, Y.; Zheng, S.; Fan, X.; Li, L.; Xiao, Y.; Luo, P.; Liu, Y.; Wang, L.; Cui, Z.; He, F.; et al. Amniotic Epithelial Cells Accelerate Diabetic Wound Healing by Modulating Inflammation and Promoting Neovascularization. Stem Cells Int. 2018, 2018, 1082076. [Google Scholar] [CrossRef]
- Dymowska, M.; Aksamit, A.; Zielniok, K.; Kniotek, M.; Kaleta, B.; Roszczyk, A.; Zych, M.; Dabrowski, F.; Paczek, L.; Burdzinska, A. Interaction between Macrophages and Human Mesenchymal Stromal Cells Derived from Bone Marrow and Wharton’s Jelly—A Comparative Study. Pharmaceutics 2021, 13, 1822. [Google Scholar] [CrossRef] [PubMed]
- Manuelpillai, U.; Tchongue, J.; Lourensz, D.; Vaghjiani, V.; Samuel, C.S.; Liu, A.; Williams, E.D.; Sievert, W. Transplantation of Human Amnion Epithelial Cells Reduces Hepatic Fibrosis in Immunocompetent CCl4-Treated Mice. Cell Transpl. 2010, 19, 1157–1168. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, N.J.; Inatomi, T.J.; Sotozono, C.J.; Fullwood, N.J.; Quantock, A.J.; Kinoshita, S. Growth Factor MRNA and Protein in Preserved Human Amniotic Membrane. Curr. Eye Res. 2000, 20, 173–177. [Google Scholar] [CrossRef]
- Alhomrani, M.; Correia, J.; Zavou, M.; Leaw, B.; Kuk, N.; Xu, R.; Saad, M.I.; Hodge, A.; Greening, D.W.; Lim, R.; et al. The Human Amnion Epithelial Cell Secretome Decreases Hepatic Fibrosis in Mice with Chronic Liver Fibrosis. Front. Pharm. 2017, 8, 748. [Google Scholar] [CrossRef]
- Thakur, G.; Bok, E.-Y.; Kim, S.-B.; Jo, C.-H.; Oh, S.-J.; Baek, J.-C.; Park, J.-E.; Kang, Y.-H.; Lee, S.-L.; Kumar, R.; et al. Scaffold-Free 3D Culturing Enhance Pluripotency, Immunomodulatory Factors, and Differentiation Potential of Wharton’s Jelly-Mesenchymal Stem Cells. Eur. J. Cell Biol. 2022, 101, 151245. [Google Scholar] [CrossRef]
- Joerger-Messerli, M.S.; Marx, C.; Oppliger, B.; Mueller, M.; Surbek, D.V.; Schoeberlein, A. Mesenchymal Stem Cells from Wharton’s Jelly and Amniotic Fluid. Best Pract. Res. Clin. Obs. Gynaecol. 2016, 31, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Koike, C.; Zhou, K.; Takeda, Y.; Fathy, M.; Okabe, M.; Yoshida, T.; Nakamura, Y.; Kato, Y.; Nikaido, T. Characterization of Amniotic Stem Cells. Cell. Reprogramm. 2014, 16, 298–305. [Google Scholar] [CrossRef]
- Al Madhoun, A.; Ali, H.; AlKandari, S.; Atizado, V.L.; Akhter, N.; Al-Mulla, F.; Atari, M. Defined Three-Dimensional Culture Conditions Mediate Efficient Induction of Definitive Endoderm Lineage from Human Umbilical Cord Wharton’s Jelly Mesenchymal Stem Cells. Stem Cell Res. 2016, 7, 165. [Google Scholar] [CrossRef]
- Mu, X.-P.; Ren, L.-Q.; Yan, H.-W.; Zhang, X.-M.; Xu, T.-M.; Wei, A.-H.; Jiang, J.-L. Enhanced Differentiation of Human Amniotic Fluid-Derived Stem Cells into Insulin-Producing Cells In Vitro. J. Diabetes Investig. 2017, 8, 34–43. [Google Scholar] [CrossRef]
- Cargnoni, A.; Papait, A.; Masserdotti, A.; Pasotti, A.; Stefani, F.R.; Silini, A.R.; Parolini, O. Extracellular Vesicles From Perinatal Cells for Anti-Inflammatory Therapy. Front. Bioeng. Biotechnol. 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Wassmer, C.-H.; Berishvili, E. Immunomodulatory Properties of Amniotic Membrane Derivatives and Their Potential in Regenerative Medicine. Curr. Diabetes Rep. 2020, 20, 31. [Google Scholar] [CrossRef] [PubMed]
- Wei, P.; Zhong, C.; Yang, X.; Shu, F.; Xiao, S.; Gong, T.; Luo, P.; Li, L.; Chen, Z.; Zheng, Y.; et al. Exosomes Derived from Human Amniotic Epithelial Cells Accelerate Diabetic Wound Healing via PI3K-AKT-MTOR-Mediated Promotion in Angiogenesis and Fibroblast Function. Burn. Trauma 2020, 8, tkaa020. [Google Scholar] [CrossRef] [PubMed]
- Magatti, M.; Cargnoni, A.; Silini, A.R.; Parolini, O. Chapter 11—Epithelial and Mesenchymal Stromal Cells from the Amniotic Membrane: Both Potent Immunomodulators. In Perinatal Stem Cells; Atala, A., Cetrulo, K.J., Taghizadeh, R.R., Murphy, S.V., Cetrulo, C.L., Eds.; Academic Press: Boston, MA, USA, 2018; pp. 147–155. ISBN 978-0-12-812015-6. [Google Scholar] [CrossRef]
- Vieira Paladino, F.; de Moraes Rodrigues, J.; da Silva, A.; Goldberg, A.C. The Immunomodulatory Potential of Wharton’s Jelly Mesenchymal Stem/Stromal Cells. Stem Cells Int. 2019, 2019, 3548917. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Qiu, X.; Ni, P.; Qiu, X.; Lin, X.; Wu, W.; Xie, L.; Lin, L.; Min, J.; Lai, X.; et al. Immunological Characteristics of Human Umbilical Cord Mesenchymal Stem Cells and the Therapeutic Effects of Their Transplantion on Hyperglycemia in Diabetic Rats. Int. J. Mol. Med. 2014, 33, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Bailey, A.J.M.; Li, H.; Kirkham, A.M.; Tieu, A.; Maganti, H.B.; Shorr, R.; Fergusson, D.A.; Lalu, M.M.; Elomazzen, H.; Allan, D.S. MSC-Derived Extracellular Vesicles to Heal Diabetic Wounds: A Systematic Review and Meta-Analysis of Preclinical Animal Studies. Stem Cell Rev. Rep. 2022, 18, 968–979. [Google Scholar] [CrossRef]
- Silini, A.R.; Papait, A.; Cargnoni, A.; Vertua, E.; Romele, P.; Bonassi Signoroni, P.; Magatti, M.; de Munari, S.; Masserdotti, A.; Pasotti, A.; et al. CM from Intact HAM: An Easily Obtained Product with Relevant Implications for Translation in Regenerative Medicine. Stem Cell Res. 2021, 12, 540. [Google Scholar] [CrossRef]
- Zia, S.; Cavallo, C.; Vigliotta, I.; Parisi, V.; Grigolo, B.; Buda, R.; Marrazzo, P.; Alviano, F.; Bonsi, L.; Zattoni, A.; et al. Effective Label-Free Sorting of Multipotent Mesenchymal Stem Cells from Clinical Bone Marrow Samples. Bioengineering 2022, 9, 49. [Google Scholar] [CrossRef]
- Zeynaloo, E.; Stone, L.D.; Dikici, E.; Ricordi, C.; Deo, S.K.; Bachas, L.G.; Daunert, S.; Lanzoni, G. Delivery of Therapeutic Agents and Cells to Pancreatic Islets: Towards a New Era in the Treatment of Diabetes. Mol. Asp. Med. 2022, 83, 101063. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Immunomodulation Activity | AESC | WJ-MSC | AFSC | AM-MSC |
|---|---|---|---|---|
| Inhibiting the cytotoxicity of natural killer cells | [137,138] | [139] | [133] | |
| Reducing of the activation of dendritic cells | [62] | [140] | [134] | |
| Reducing B cell activation | [62] | [141] | ||
| Anti-proliferative effect on activated PBMC | [21,74] | [142] | [128] | |
| [143,144] | [142] | |||
| Promoting the anti-inflammatory phenotype (M2) of macrophage populations | [145] | [146] | [129,130] | |
| Reducing T lymphocytes proliferation | [75,138] | [81,115,140] | [133] | [133] |
| In vivo allogenic transplantation | [37,51,75,147] | [84,85,99,112] | ||
| Increased expression of: TGF-β1, IL-6, TSG-6, PGE-2 | [148] | [112,140] | ||
| Secretion of EVs | [115,149] | [115] | ||
| Secretion of: IL-10, PGE2, hyaluronic acid | [112,140] | |||
| Expression of HLA Ib | [57,58,59] | [81] | ||
| Expression of migration inhibitor factor (MIF) | [62] | |||
| Differentiation capacity | AECs | WJ-MSCs | AFSCs | AM-MSCs |
| Expression of pluripotency markers: NANOG, OCT-4, SSEA-3, SSEA-4, TRA1-60, TRA1-80 | [34] | [150] | [151] | [152] |
| Pancreatic-endodermic lineage | [18,43,45] | [95,102,106,153] | [154] | [135,136] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paris, F.; Pizzuti, V.; Marrazzo, P.; Pession, A.; Alviano, F.; Bonsi, L. Perinatal Stem Cell Therapy to Treat Type 1 Diabetes Mellitus: A Never-Say-Die Story of Differentiation and Immunomodulation. Int. J. Mol. Sci. 2022, 23, 14597. https://doi.org/10.3390/ijms232314597
Paris F, Pizzuti V, Marrazzo P, Pession A, Alviano F, Bonsi L. Perinatal Stem Cell Therapy to Treat Type 1 Diabetes Mellitus: A Never-Say-Die Story of Differentiation and Immunomodulation. International Journal of Molecular Sciences. 2022; 23(23):14597. https://doi.org/10.3390/ijms232314597
Chicago/Turabian StyleParis, Francesca, Valeria Pizzuti, Pasquale Marrazzo, Andrea Pession, Francesco Alviano, and Laura Bonsi. 2022. "Perinatal Stem Cell Therapy to Treat Type 1 Diabetes Mellitus: A Never-Say-Die Story of Differentiation and Immunomodulation" International Journal of Molecular Sciences 23, no. 23: 14597. https://doi.org/10.3390/ijms232314597
APA StyleParis, F., Pizzuti, V., Marrazzo, P., Pession, A., Alviano, F., & Bonsi, L. (2022). Perinatal Stem Cell Therapy to Treat Type 1 Diabetes Mellitus: A Never-Say-Die Story of Differentiation and Immunomodulation. International Journal of Molecular Sciences, 23(23), 14597. https://doi.org/10.3390/ijms232314597