

Derivatives of Trimethoxybenzoic Acid and Gallic Acid as Potential Efflux Pump Inhibitors: In Silico and In Vitro Studies
 , , , , ,
, , , , ,  ,
,  and
and 
Abstract
1. Introduction
2. Results and Discussion
2.1. Chemistry
2.2. Molecular Docking
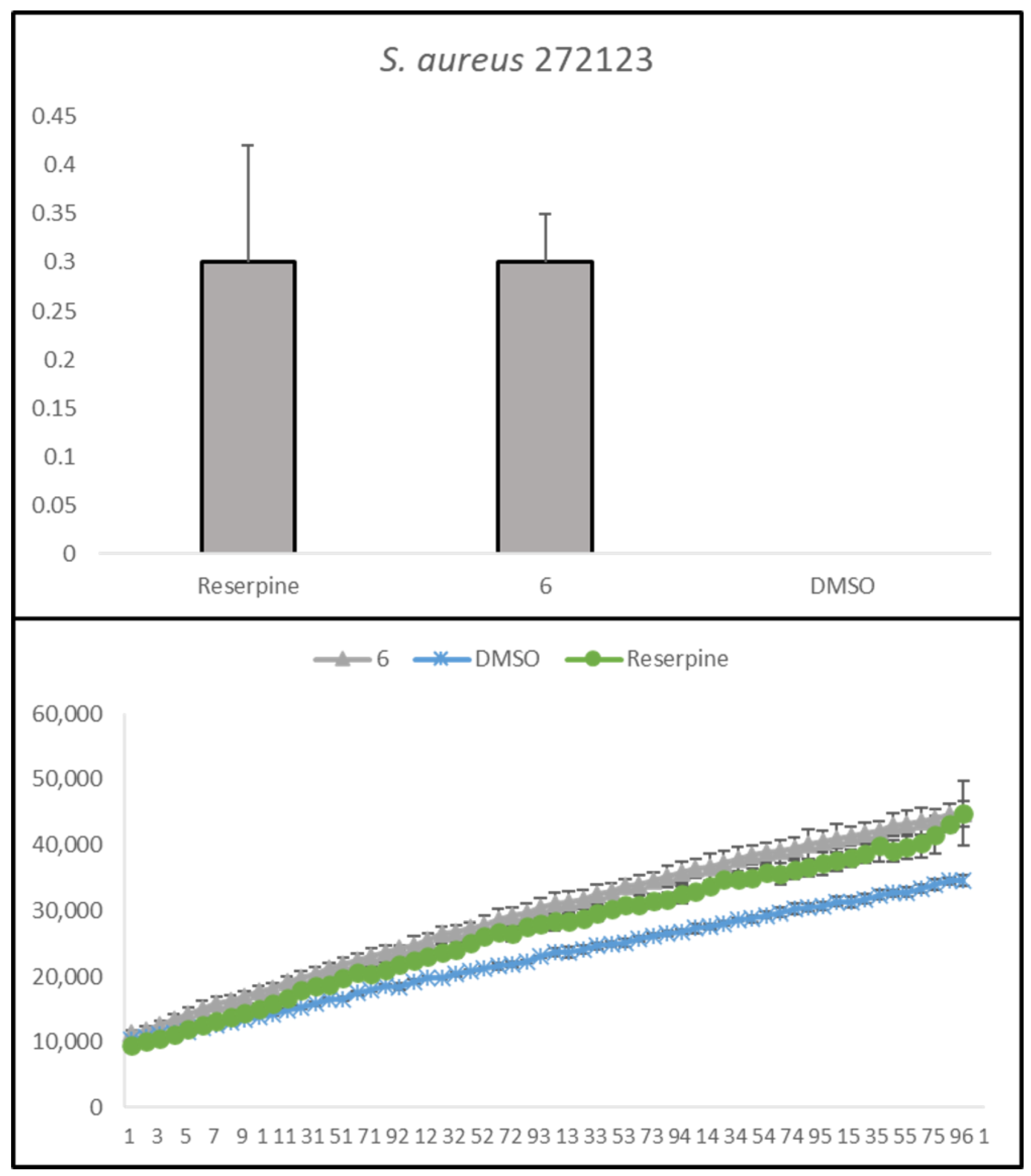
2.3. Antibacterial Activity and Antibiotic Potentiation
2.4. Efflux Pump Inhibition
2.5. Inhibition of Biofilm Formation and Quorum-Sensing Assays
2.6. Antifungal Activity
2.7. In Silico ADME Properties
3. Materials and Methods
3.1. Chemistry
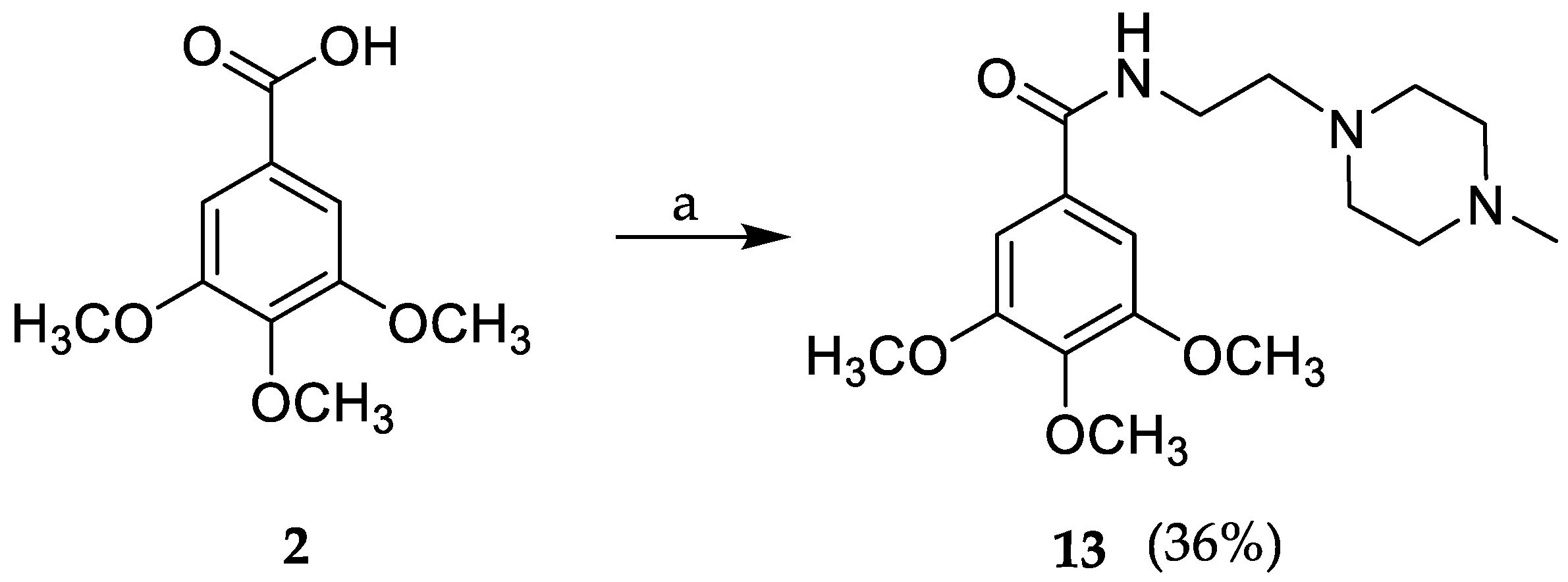
3.1.1. General Procedure for the Synthesis of Compounds 3–13
3,4,5-Trimethoxy-N-(2-(4-methylpiperazin-1-yl)ethyl)benzamide (13)
3.2. Molecular Docking
3.3. Culture Media and Chemicals
3.4. Microorganisms
3.5. Antibacterial and Antibiotic Potentiation Assays
3.6. Efflux Pump Inhibition
3.7. Inhibition of Biofilm Formation
3.8. Quorum-Sensing Inhibition
3.9. Antifungal Assays
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaynes, R.P. The Discovery of Penicillin—New Insights After More Than 75 Years of Clinical Use. Emerg. Infect. Dis. 2017, 23, 849–853. [Google Scholar] [CrossRef]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Andrei, S.; Droc, G.; Stefan, G. FDA approved antibacterial drugs: 2018-2019. Discoveries 2019, 7, e102. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, P.S. Antibacterial Discovery: 21st Century Challenges. Antibiotics 2020, 9, 213. [Google Scholar] [CrossRef] [PubMed]
- Donadio, S.; Maffioli, S.; Monciardini, P.; Sosio, M.; Jabes, D. Antibiotic discovery in the twenty-first century: Current trends and future perspectives. J. Antibiot. 2010, 63, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Durães, F.; Pinto, M.; Sousa, E. Medicinal Chemistry Updates on Bacterial Efflux Pump Modulators. Curr. Med. Chem. 2018, 25, 6030–6069. [Google Scholar] [CrossRef]
- Neves, A.R.; Boas, C.V.; Gonçalves, C.; Vasconcelos, V.; Pinto, M.; Silva, E.R.; Sousa, E.; Almeida, J.R.; Correia-da-Silva, M. Gallic acid derivatives as inhibitors of mussel (Mytilus galloprovincialis) larval settlement: Lead optimization, biological evaluation and use in antifouling coatings. Bioorganic Chem. 2022, 126, 105911. [Google Scholar] [CrossRef]
- Neves, A.R.; Gomes, L.C.; Faria, S.I.; Sousa, J.; Ruivo, R.; Páscoa, I.; Pinto, M.; Sousa, E.; Santos, M.M.; Silva, E.R.; et al. Antifouling Marine Coatings with a Potentially Safer and Sustainable Synthetic Polyphenolic Derivative. Mar. Drugs 2022, 20, 507. [Google Scholar] [CrossRef]
- Perron, Y.G. Therapeutic Agents. US2870145A, 20 January 1959. [Google Scholar]
- Zimmermann, S.; Klinger-Strobel, M.; Bohnert, J.A.; Wendler, S.; Rödel, J.; Pletz, M.W.; Löffler, B.; Tuchscherr, L. Clinically Approved Drugs Inhibit the Staphylococcus aureus Multidrug NorA Efflux Pump and Reduce Biofilm Formation. Front. Microbiol. 2019, 10, 2762. [Google Scholar] [CrossRef]
- Shaheen, A.; Afridi, W.A.; Mahboob, S.; Sana, M.; Zeeshan, N.; Ismat, F.; Mirza, O.; Iqbal, M.; Rahman, M. Reserpine Is the New Addition into the Repertoire of AcrB Efflux Pump Inhibitors. Mol. Biol. 2019, 53, 596–605. [Google Scholar] [CrossRef]
- Shamon, S.D.; Perez, M.I. Blood pressure-lowering efficacy of reserpine for primary hypertension. Cochrane Database Syst. Rev. 2016, 12, Cd007655. [Google Scholar] [CrossRef] [PubMed]
- Hoenders, H.J.R.; Bartels-Velthuis, A.A.; Vollbehr, N.K.; Bruggeman, R.; Knegtering, H.; de Jong, J. Natural Medicines for Psychotic Disorders: A Systematic Review. J. Nerv. Ment. Dis. 2018, 206, 81–101. [Google Scholar] [CrossRef]
- Marcus, S.M.; Borzelleca, J.F. Observations on reserpine-induced bradycardia. Arch. Int. Pharmacodyn. Ther. 1968, 174, 12–16. [Google Scholar] [PubMed]
- Mladěnka, P.; Applová, L.; Patočka, J.; Costa, V.M.; Remiao, F.; Pourová, J.; Mladěnka, A.; Karlíčková, J.; Jahodář, L.; Vopršalová, M.; et al. Comprehensive review of cardiovascular toxicity of drugs and related agents. Med. Res. Rev. 2018, 38, 1332–1403. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Chen, M.; Yu, Z.; Bell, J.M.; Wang, H.; Forrester, I.; Villarreal, H.; Jakana, J.; Du, D.; Luisi, B.F.; et al. In situ structure and assembly of the multidrug efflux pump AcrAB-TolC. Nat. Commun. 2019, 10, 2635. [Google Scholar] [CrossRef]
- Yan, N. Structural advances for the major facilitator superfamily (MFS) transporters. Trends Biochem. Sci. 2013, 38, 151–159. [Google Scholar] [CrossRef]
- Thai, K.-M.; Ngo, T.-D.; Phan, T.-V.; Tran, T.-D.; Nguyen, N.-V.; Nguyen, T.-H.; Le, M.-T. Virtual Screening for Novel Staphylococcus Aureus NorA Efflux Pump Inhibitors From Natural Products. Med. Chem. 2015, 11, 135–155. [Google Scholar] [CrossRef]
- Yin, Y.; He, X.; Szewczyk, P.; Nguyen, T.; Chang, G. Structure of the Multidrug Transporter EmrD from Escherichia coli. Science 2006, 312, 741–744. [Google Scholar] [CrossRef]
- Alav, I.; Sutton, J.M.; Rahman, K.M. Role of bacterial efflux pumps in biofilm formation. J. Antimicrob. Chemother. 2018, 73, 2003–2020. [Google Scholar] [CrossRef]
- Costa, S.S.; Sobkowiak, B.; Parreira, R.; Edgeworth, J.D.; Viveiros, M.; Clark, T.G.; Couto, I. Genetic Diversity of norA, Coding for a Main Efflux Pump of Staphylococcus aureus. Front. Genet. 2019, 9. [Google Scholar] [CrossRef]
- Bruce, S.A.; Smith, J.T.; Mydosh, J.L.; Ball, J.; Needle, D.B.; Gibson, R.; Andam, C.P. Shared antibiotic resistance and virulence genes in Staphylococcus aureus from diverse animal hosts. Sci. Rep. 2022, 12, 4413. [Google Scholar] [CrossRef] [PubMed]
- Durães, F.; Resende, D.I.S.P.; Palmeira, A.; Szemerédi, N.; Pinto, M.M.M.; Spengler, G.; Sousa, E. Xanthones Active against Multidrug Resistance and Virulence Mechanisms of Bacteria. Antibiotics 2021, 10, 600. [Google Scholar] [CrossRef] [PubMed]
- Parai, D.; Banerjee, M.; Dey, P.; Mukherjee, S.K. Reserpine attenuates biofilm formation and virulence of Staphylococcus aureus. Microb. Pathog. 2020, 138, 103790. [Google Scholar] [CrossRef] [PubMed]
- Benomar, S.; Evans, K.C.; Unckless, R.L.; Chandler, J.R. Efflux Pumps in Chromobacterium Species Increase Antibiotic Resistance and Promote Survival in a Coculture Competition Model. Appl. Environ. Microbiol. 2019, 85, e00908–e00919. [Google Scholar] [CrossRef]
- Gajdács, M.; Spengler, G. The Role of Drug Repurposing in the Development of Novel Antimicrobial Drugs: Non-Antibiotic Pharmacological Agents as Quorum Sensing-Inhibitors. Antibiotics 2019, 8, 270. [Google Scholar] [CrossRef]
- Mardanova, A.M.; Bogomol’naya, L.M.; Romanova, Y.D.; Sharipova, M.R. Efflux systems in Serratia marcescens. Microbiology 2013, 82, 668–679. [Google Scholar] [CrossRef]
- Wang, S.; Dong, G.; Sheng, C. Structural simplification: An efficient strategy in lead optimization. Acta Pharm. Sin. B 2019, 9, 880–901. [Google Scholar] [CrossRef]
- Murakami, S.; Nakashima, R.; Yamashita, E.; Matsumoto, T.; Yamaguchi, A. Crystal structures of a multidrug transporter reveal a functionally rotating mechanism. Nature 2006, 443, 173–179. [Google Scholar] [CrossRef]
- Mikolosko, J.; Bobyk, K.; Zgurskaya, H.I.; Ghosh, P. Conformational flexibility in the multidrug efflux system protein AcrA. Structure 2006, 14, 577–587. [Google Scholar] [CrossRef]
- Koronakis, V.; Sharff, A.; Koronakis, E.; Luisi, B.; Hughes, C. Crystal structure of the bacterial membrane protein TolC central to multidrug efflux and protein export. Nature 2000, 405, 914–919. [Google Scholar] [CrossRef]
- Sussman, J.L.; Lin, D.; Jiang, J.; Manning, N.O.; Prilusky, J.; Ritter, O.; Abola, E.E. Protein Data Bank (PDB): Database of Three-Dimensional Structural Information of Biological Macromolecules. Acta Crystallogr. Sect. D 1998, 54, 1078–1084. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Apweiler, R.; Bairoch, A.; Wu, C.H.; Barker, W.C.; Boeckmann, B.; Ferro, S.; Gasteiger, E.; Huang, H.; Lopez, R.; Magrane, M.; et al. UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2017, 45, D158–D169. [Google Scholar] [CrossRef] [PubMed]
- Zárate, S.G.; Morales, P.; Świderek, K.; Bolanos-Garcia, V.M.; Bastida, A. A Molecular Modeling Approach to Identify Novel Inhibitors of the Major Facilitator Superfamily of Efflux Pump Transporters. Antibiotics 2019, 8, 25. [Google Scholar] [CrossRef]
- Bessa, L.J.; Barbosa-Vasconcelos, A.; Mendes, A.; Vaz-Pires, P.; Martins da Costa, P. High prevalence of multidrug-resistant Escherichia coli and Enterococcus spp. in river water, upstream and downstream of a wastewater treatment plant. J. Water Health 2014, 12, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Gajdács, M.; Spengler, G. Standard operating procedure (SOP) for disk diffusion-based quorum sensing inhibition assays. Acta Pharm. Hung. 2020, 89, 117–125. [Google Scholar] [CrossRef][Green Version]
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically, 11th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Spengler, G.; Takács, D.; Horváth, A.; Szabó, A.M.; Riedl, Z.; Hajós, G.; Molnár, J.; Burián, K. Efflux pump inhibiting properties of racemic phenothiazine derivatives and their enantiomers on the bacterial AcrAB-TolC system. In Vivo 2014, 28, 1071–1075. [Google Scholar]
- Peeters, E.; Nelis, H.J.; Coenye, T. Comparison of multiple methods for quantification of microbial biofilms grown in microtiter plates. J. Microbiol. Methods 2008, 72, 157–165. [Google Scholar] [CrossRef]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Approved standard—CLSI document M27-A3; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008. [Google Scholar]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi; Approved Standard—Second Edition. CLSI document M38-A2; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Docking Score | ||||
|---|---|---|---|---|---|
| AcrB | AcrA | TolC | NorA | ||
| HH | LD | ||||
| 1 | −5.9 | −5.1 | −4.5 | −5.5 | −4.1 |
| 2 | −5.6 | −4.3 | −3.7 | −5.5 | −4.0 |
| 3 | −6.5 | −4.9 | −4.3 | −6.0 | −4.5 |
| 4 | −6.2 | −5.5 | −4.6 | −5.9 | −4.2 |
| 5 | −7.3 | −5.7 | −4.7 | −6.4 | −7.3 |
| 6 | −7.6 | −6.4 | −5.4 | −6.7 | −6.2 |
| 7 | −6.8 | −5.0 | −6.0 | −5.7 | −4.4 |
| 8 | −6.7 | −5.0 | −5.8 | −5.9 | −4.7 |
| 9 | −8.9 | −5.8 | −6.7 | −6.5 | −5.9 |
| 10 | −9.8 | −6.5 | −7.1 | −7.3 | −6.2 |
| 11 | −6.6 | −4.7 | −5.3 | −5.7 | −3.9 |
| 12 | −7.0 | −5.0 | −5.0 | −5.9 | −4.8 |
| 13 | −7.1 | −5.0 | −5.6 | −5.9 | −4.8 |
| Reserpine | −8.1 | 5.6 | 4.6 | −7.5 | −4.1 |
| Compound | RFI ± SD | |
|---|---|---|
| S. aureus 272123 | S. Typhimurium SL1344 | |
| 3 | ND | ND |
| 4 | ND | ND |
| 5 | 0.08 ± 0.07 | 0.22 ± 0.02 |
| 6 | 0.30 ± 0.05 | 0.22 ± 0.11 |
| 7 | −0.02 ± 0.01 | −0.07 ± 0.03 |
| 8 | −0.07 ± 0.03 | −0.07 ± 0.05 |
| 9 | −0.35 ± 0.01 | −0.41 ± 0.01 |
| 10 | −0.21 ± 0.03 | −0.36 ± 0.01 |
| 11 | −0.03 ± 0.03 | −0.06 ± 0.01 |
| 12 | 0.17 ± 0.03 | −0.01 ± 0.01 |
| 13 | 0.14 ± 0.02 | 0.06 ± 0.01 |
| Reserpine | 0.30 ± 0.12 | 0.04 ± 0.07 |
| CCCP | ND | 0.16 ± 0.06 |
| Compound | Molecular Weight | # Rotatable Bonds | # H-Bond Acceptors | # H-Bond Donors | ilogP |
|---|---|---|---|---|---|
| Reserpine | 608.68 | 10 | 10 | 1 | 5.16 |
| 5 | 330.38 | 9 | 4 | 2 | 3.12 |
| 6 | 355.39 | 8 | 5 | 2 | 2.46 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neves, A.R.; Durães, F.; Freitas-Silva, J.; Szemerédi, N.; Martins-da-Costa, P.; Pinto, E.; Correia-da-Silva, M.; Spengler, G.; Sousa, E. Derivatives of Trimethoxybenzoic Acid and Gallic Acid as Potential Efflux Pump Inhibitors: In Silico and In Vitro Studies. Int. J. Mol. Sci. 2022, 23, 14468. https://doi.org/10.3390/ijms232214468
Neves AR, Durães F, Freitas-Silva J, Szemerédi N, Martins-da-Costa P, Pinto E, Correia-da-Silva M, Spengler G, Sousa E. Derivatives of Trimethoxybenzoic Acid and Gallic Acid as Potential Efflux Pump Inhibitors: In Silico and In Vitro Studies. International Journal of Molecular Sciences. 2022; 23(22):14468. https://doi.org/10.3390/ijms232214468
Chicago/Turabian StyleNeves, Ana Rita, Fernando Durães, Joana Freitas-Silva, Nikoletta Szemerédi, Paulo Martins-da-Costa, Eugénia Pinto, Marta Correia-da-Silva, Gabriella Spengler, and Emília Sousa. 2022. "Derivatives of Trimethoxybenzoic Acid and Gallic Acid as Potential Efflux Pump Inhibitors: In Silico and In Vitro Studies" International Journal of Molecular Sciences 23, no. 22: 14468. https://doi.org/10.3390/ijms232214468
APA StyleNeves, A. R., Durães, F., Freitas-Silva, J., Szemerédi, N., Martins-da-Costa, P., Pinto, E., Correia-da-Silva, M., Spengler, G., & Sousa, E. (2022). Derivatives of Trimethoxybenzoic Acid and Gallic Acid as Potential Efflux Pump Inhibitors: In Silico and In Vitro Studies. International Journal of Molecular Sciences, 23(22), 14468. https://doi.org/10.3390/ijms232214468