
Impact of High Glucose on Ocular Surface Glycocalyx Components: Implications for Diabetes-Associated Ocular Surface Damage


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
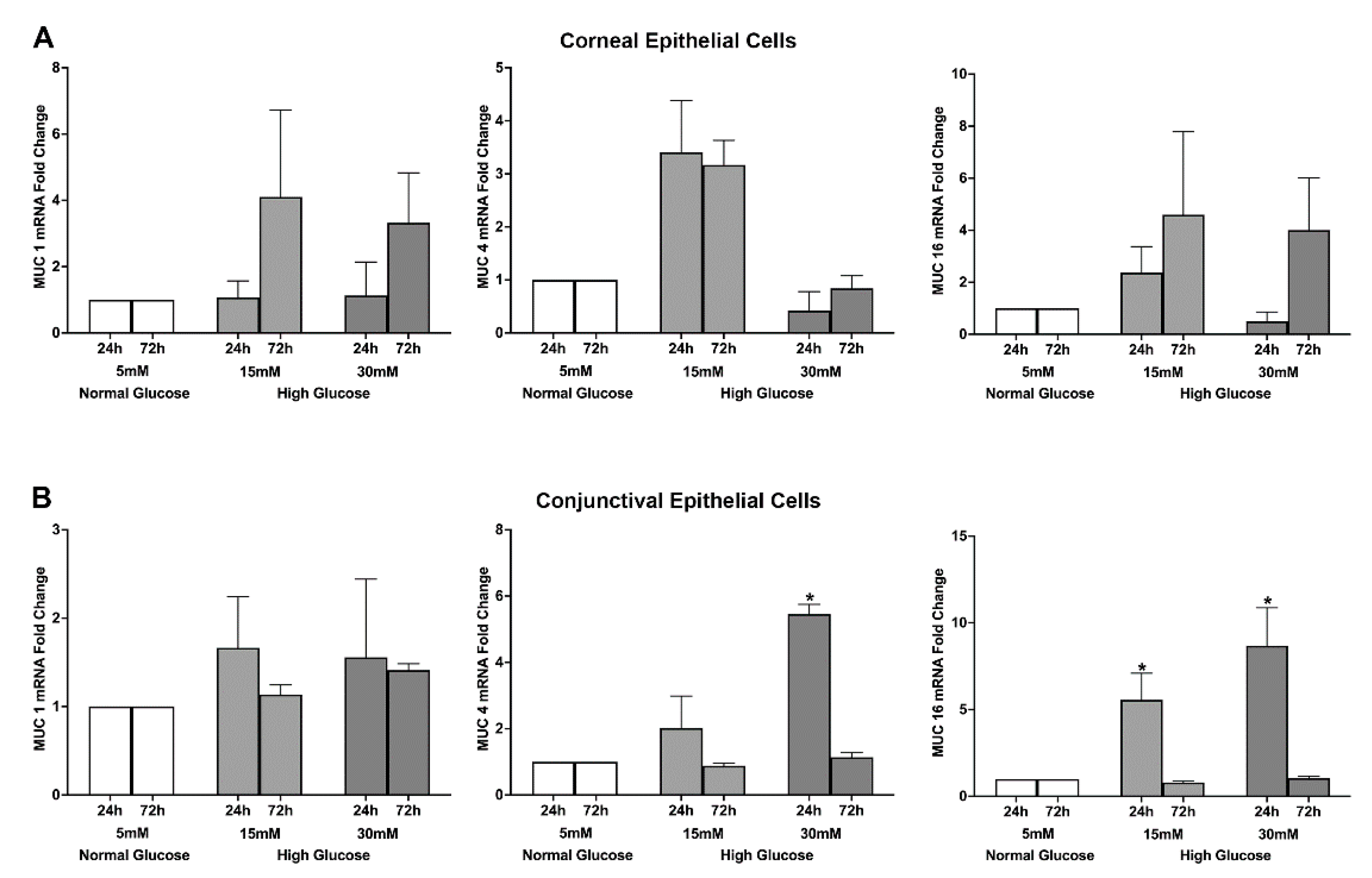
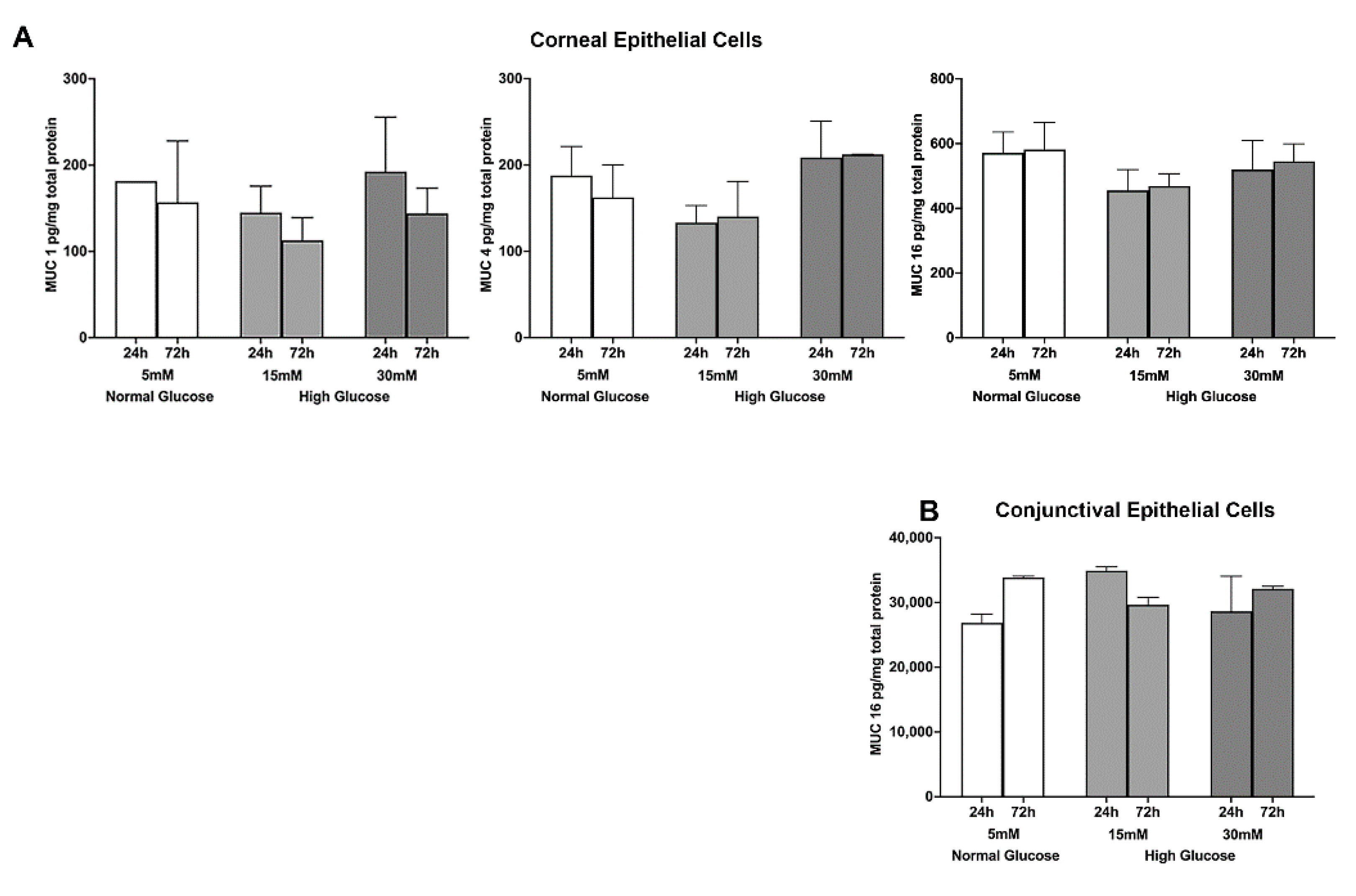
2.1. Effect of a High Glucose Level on Membrane-Tethered Mucins’ Gene and Protein Expression in Stratified Human Corneal and Conjunctival Epithelial Cells
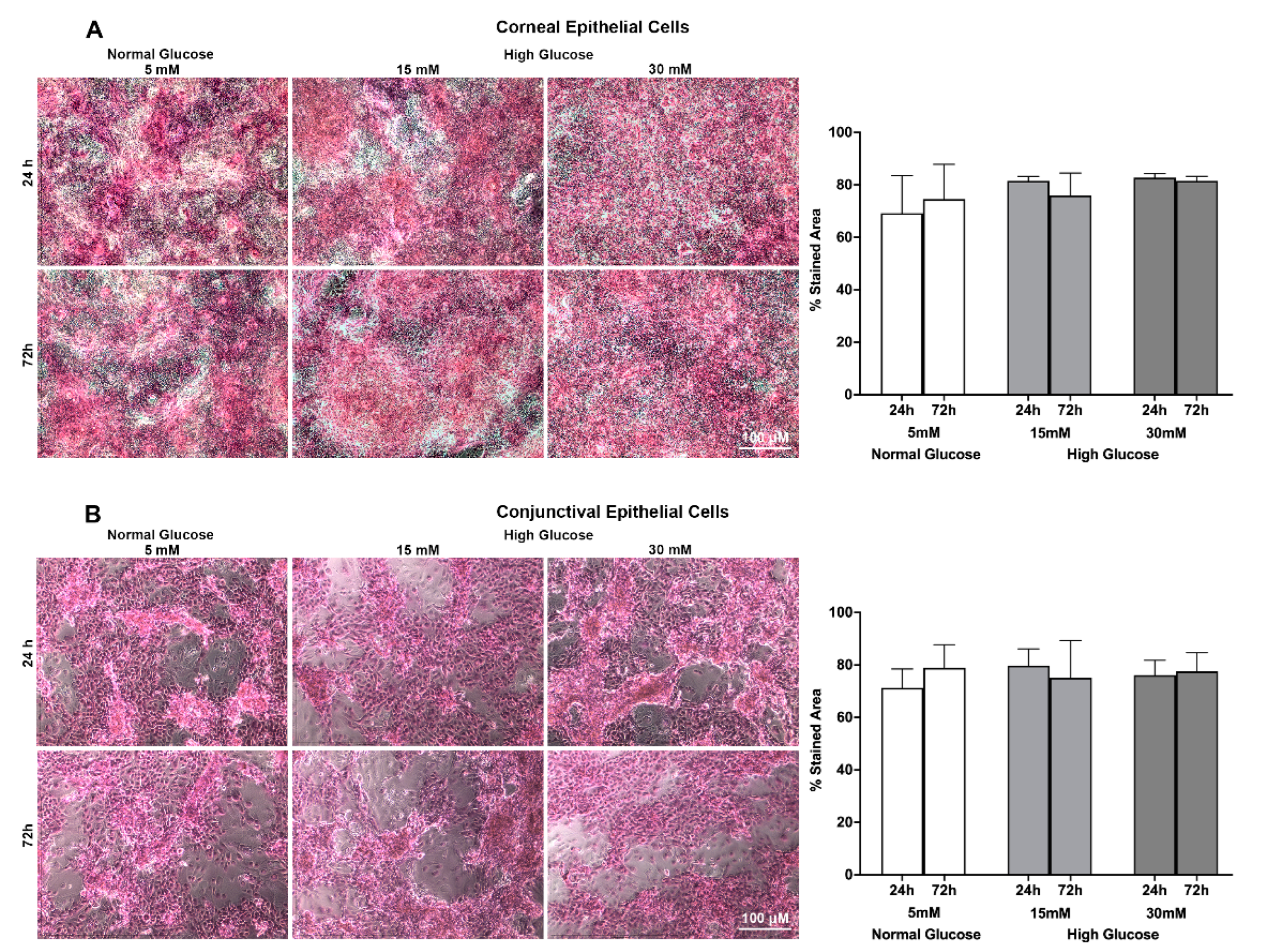
2.2. Effect of a High Glucose Level on MUC 16 Barrier Function of Stratified Human Corneal and Conjunctival Epithelial Cell Using Rose Bengal Exclusion Assay
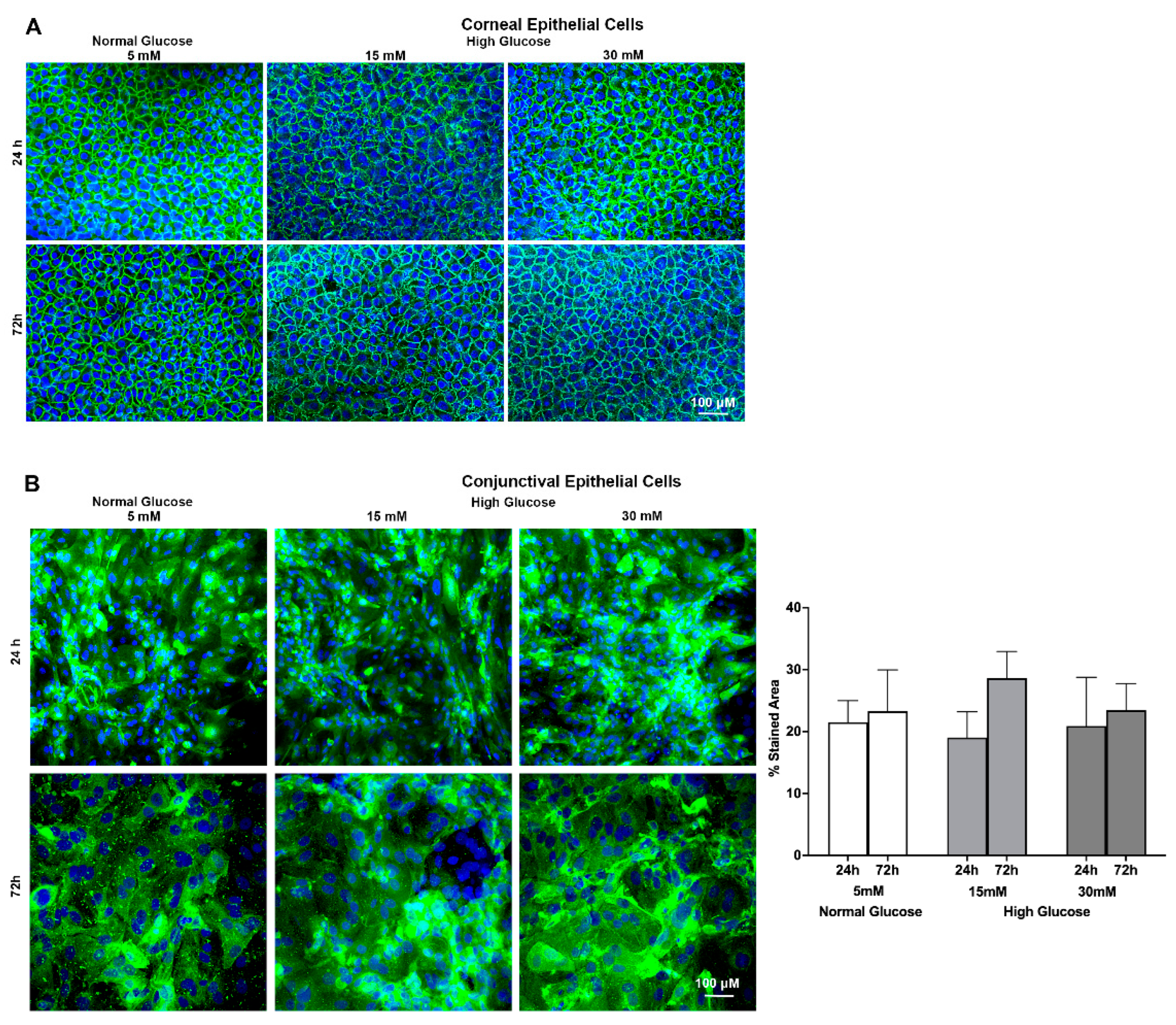
2.3. Effect of a High Glucose Level on O-glycosyl Side Chains in Stratified Human Corneal and Conjunctival Epithelial Cells Using Jacalin Staining
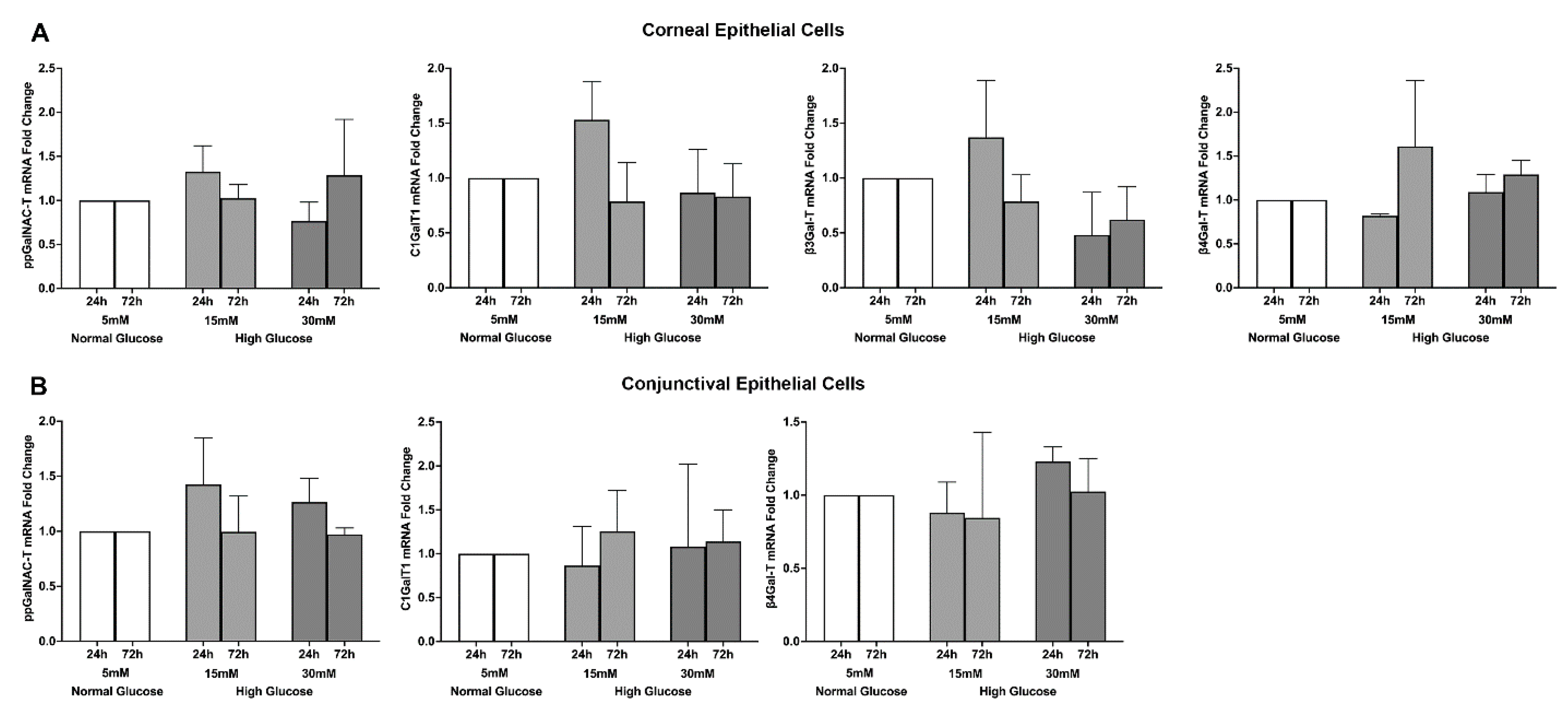
2.4. Effect of a High Glucose Level on Glycosyltransferases’ Gene Expression in Stratified Human Corneal and Conjunctival Epithelial Cells
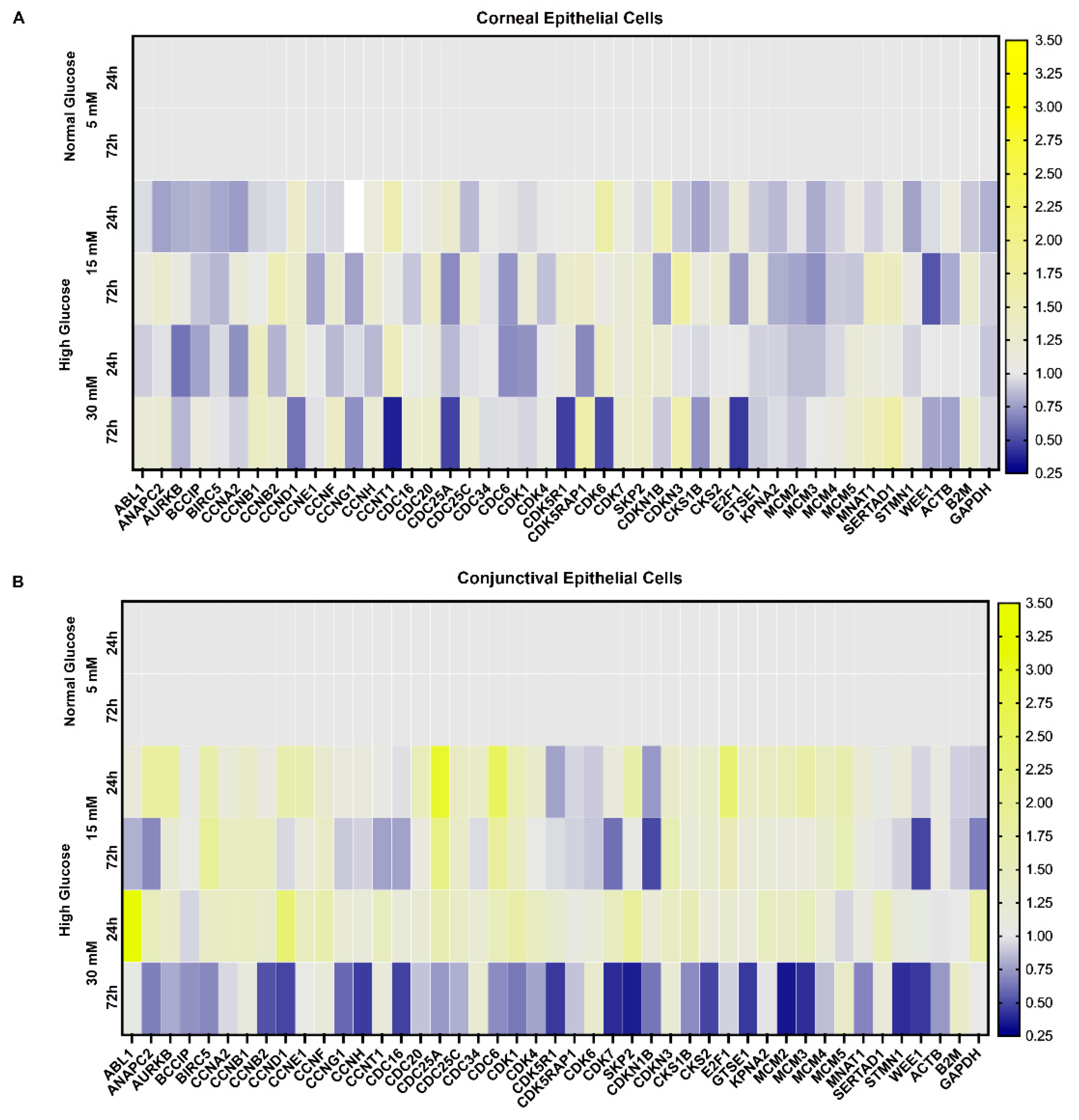
2.5. Effect of a High Glucose Level on Expression of Genes Involved in Cell Cycle in Stratified Human Corneal and Conjunctival Epithelial Cells
3. Discussion
4. Materials and Methods
4.1. Stratified Human Corneal Epithelial and Conjunctival Epithelial Cell Cultures
4.2. Glucose Treatments
4.3. Isolation of mRNA Preparation of cDNA
4.4. Mucin and Glycosyltransferase Gene Expression Quantification
4.5. Mucin Enzyme-Linked Immunosorbent Assays
4.6. Jacalin Staining
4.7. Rose Bengal Staining
4.8. PCR Array for Quantification of Genes Involved in Cell Cycle
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shih, K.C.; Lam, K.S.; Tong, L. A systematic review on the impact of diabetes mellitus on the ocular surface. Nutr. Diabetes 2017, 7, e251. [Google Scholar] [CrossRef] [PubMed]
- Misra, S.L.; Braatvedt, G.D.; Patel, D.V. Impact of diabetes mellitus on the ocular surface: A review. Clin. Exp. Ophthalmol. 2016, 44, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Ljubimov, A.V. Diabetic complications in the cornea. Vision Res. 2017, 139, 138–152. [Google Scholar] [CrossRef] [PubMed]
- Sayin, N.; Kara, N.; Pekel, G. Ocular complications of diabetes mellitus. World J. Diabetes 2015, 6, 92–108. [Google Scholar] [CrossRef]
- Priyadarsini, S.; Whelchel, A.; Nicholas, S.; Sharif, R.; Riaz, K.; Karamichos, D. Diabetic keratopathy: Insights and challenges. Surv. Ophthalmol. 2020, 65, 513–529. [Google Scholar] [CrossRef]
- Mantelli, F.; Mauris, J.; Argüeso, P. The ocular surface epithelial barrier and other mechanisms of mucosal protection: From allergy to infectious diseases. Curr. Opin. Allergy Clin. Immunol. 2013, 13, 563–568. [Google Scholar] [CrossRef]
- Alarcon, I.; Tam, C.; Mun, J.J.; LeDue, J.; Evans, D.J.; Fleiszig, S.M. Factors impacting corneal epithelial barrier function against Pseudomonas aeruginosa traversal. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1368–1377. [Google Scholar] [CrossRef]
- Leong, Y.Y.; Tong, L. Barrier function in the ocular surface: From conventional paradigms to new opportunities. Ocul. Surf. 2015, 13, 103–109. [Google Scholar] [CrossRef]
- Ansari, A.S.; de Lusignan, S.; Hinton, W.; Munro, N.; McGovern, A. The association between diabetes, level of glycaemic control and eye infection: Cohort database study. Prim. Care Diabetes 2017, 11, 421–429. [Google Scholar] [CrossRef]
- Chang, Y.S.; Tai, M.C.; Ho, C.H.; Chu, C.C.; Wang, J.J.; Tseng, S.H.; Jan, R.L. Risk of Corneal Ulcer in Patients with Diabetes Mellitus: A Retrospective Large-Scale Cohort Study. Sci. Rep. 2020, 10, 7388. [Google Scholar] [CrossRef]
- Kruse, A.; Thomsen, R.W.; Hundborg, H.H.; Knudsen, L.L.; Sørensen, H.T.; Schønheyder, H.C. Diabetes and risk of acute infectious conjunctivitis-a population-based case-control study. Diabet. Med. 2006, 23, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Yang, S.; Zhai, H.L.; Zhang, Y.Y.; Cui, C.X.; Wang, J.Y.; Xie, L.X. A comparative study of risk factors for corneal infection in diabetic and non-diabetic patients. Int. J. Ophthalmol. 2018, 11, 43–47. [Google Scholar] [PubMed]
- Chang, S.W.; Hsu, H.C.; Hu, F.R.; Chen, M.S. Corneal autofluorescence and epithelial barrier function in diabetic patients. Ophthalmic Res. 1995, 27, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Gekka, M.; Miyata, K.; Nagai, Y.; Nemoto, S.; Sameshima, T.; Tanabe, T.; Maruoka, S.; Nakahara, M.; Kato, S.; Amano, S. Corneal epithelial barrier function in diabetic patients. Cornea 2004, 23, 35–37. [Google Scholar] [CrossRef] [PubMed]
- Alfuraih, S.; Barbarino, A.; Ross, C.; Shamloo, K.; Jhanji, V.; Zhang, M.; Sharma, A. Effect of High Glucose on Ocular Surface Epithelial Cell Barrier and Tight Junction Proteins. Invest. Ophthalmol. Vis. Sci. 2020, 61, 3. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.W.; Kaili, D.; Freeman, J.; Lei, C.Y.; Geng, B.C.; Tan, T.; He, J.F.; Shi, Z.; Ma, J.J.; Luo, Y.H.; et al. Diabetes inhibits corneal epithelial cell migration and tight junction formation in mice and human via increasing ROS and impairing Akt signaling. Acta Pharmacol. Sin. 2019, 40, 1205–1211. [Google Scholar] [CrossRef]
- Uchino, Y. The Ocular Surface Glycocalyx and its Alteration in Dry Eye Disease: A Review. Investig. Ophthalmol. Vis. Sci. 2018, 59, DES157–DES162. [Google Scholar] [CrossRef]
- Argüeso, P. Glycobiology of the ocular surface: Mucins and lectins. Jpn. J. Ophthalmol. 2013, 57, 150–155. [Google Scholar] [CrossRef][Green Version]
- Guzman-Aranguez, A.; Argüeso, P. Structure and biological roles of mucin-type O-glycans at the ocular surface. Ocul. Surf. 2010, 8, 8–17. [Google Scholar] [CrossRef]
- Brockhausen, I.; Elimova, E.; Woodward, A.M.; Argüeso, P. Glycosylation pathways of human corneal and conjunctival epithelial cell mucins. Carbohydr. Res. 2018, 470, 50–56. [Google Scholar] [CrossRef]
- Argüeso, P.; Guzman-Aranguez, A.; Mantelli, F.; Cao, Z.; Ricciuto, J.; Panjwani, N. Association of cell surface mucins with galectin-3 contributes to the ocular surface epithelial barrier. J. Biol. Chem. 2009, 284, 23037–23045. [Google Scholar] [CrossRef] [PubMed]
- Jacob, B.; Alfuraih, S.; Ross, C.; Kiumars, S.; Sharma, A. Type I diabetes mellitus adversely impacts ocular surface glycocalyx and mucins. Invest. Ophthalmol. Vis. Sci. 2020, 61, 136. [Google Scholar]
- Rutter, G.A.; Tavaré, J.M.; Palmer, D.G. Regulation of Mammalian Gene Expression by Glucose. News Physiol. Sci. 2000, 15, 149–154. [Google Scholar] [CrossRef]
- Hall, E.; Dekker Nitert, M.; Volkov, P.; Malmgren, S.; Mulder, H.; Bacos, K.; Ling, C. The effects of high glucose exposure on global gene expression and DNA methylation in human pancreatic islets. Mol. Cell. Endocrinol. 2018, 472, 57–67. [Google Scholar] [CrossRef]
- Meugnier, E.; Rome, S.; Vidal, H. Regulation of gene expression by glucose. Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Blalock, T.D.; Spurr-Michaud, S.J.; Tisdale, A.S.; Heimer, S.R.; Gilmore, M.S.; Ramesh, V.; Gipson, I.K. Functions of MUC16 in corneal epithelial cells. Invest. Ophthalmol. Vis. Sci. 2007, 48, 4509–4518. [Google Scholar] [CrossRef]
- Argüeso, P.; Tisdale, A.; Spurr-Michaud, S.; Sumiyoshi, M.; Gipson, I.K. Mucin characteristics of human corneal-limbal epithelial cells that exclude the rose bengal anionic dye. Invest. Ophthalmol. Vis. Sci. 2006, 47, 113–119. [Google Scholar] [CrossRef]
- Jeyaprakash, A.A.; Geetha Rani, P.; Banuprakash Reddy, G.; Banumathi, S.; Betzel, C.; Sekar, K.; Surolia, A.; Vijayan, M. Crystal structure of the jacalin-T-antigen complex and a comparative study of lectin-T-antigen complexes. J. Mol. Biol. 2002, 321, 637–645. [Google Scholar] [CrossRef]
- International Diabetes Federation. Atlas, 10th ed.; International Diabetes Federation: Brussels, Belgium, 2021. [Google Scholar]
- Govindarajan, B.; Gipson, I.K. Membrane-tethered mucins have multiple functions on the ocular surface. Exp. Eye Res. 2010, 90, 655–663. [Google Scholar] [CrossRef]
- Taniguchi, T.; Woodward, A.M.; Magnelli, P.; McColgan, N.M.; Lehoux, S.; Jacobo, S.M.P.; Mauris, J.; Argüeso., P. N-Glycosylation affects the stability and barrier function of the MUC16 mucin. J. Biol. Chem. 2017, 292, 11079–11090. [Google Scholar] [CrossRef]
- Fujita, H.; Morita, I.; Takase, H.; Ohno-Matsui, K.; Murota, S.; Mochizuki, M. The Effect of High Glucose on Proliferation, Adhesion and Migration of Normal Human Corneal Epithelial Cells. Investig. Ophthalmol. Vis. Sci. 2002, 43, 1635. [Google Scholar]
- Giurdanella, G.; Longo, A.; Salerno, L.; Romeo, G.; Intagliata, S.; Lupo, G.; Distefano, A.; Platania, C.B.M.; Bucolo, C.; Li Volti, G.; et al. Glucose-impaired Corneal Re-epithelialization Is Promoted by a Novel Derivate of Dimethyl Fumarate. Antioxidants 2021, 10, 831. [Google Scholar] [CrossRef] [PubMed]
- Robertson, D.M.; Li, L.; Fisher, S.; Pearce, V.P.; Shay, J.W.; Wright, W.E.; Cavanagh, H.D.; Jester, J.V. Characterization of growth and differentiation in a telomerase-immortalized human corneal epithelial cell line. Investig. Ophthalmol. Vis. Sci. 2005, 46, 470–478. [Google Scholar] [CrossRef]
- Gipson, I.K.; Spurr-Michaud, S.; Argüeso, P.; Tisdale, A.; Ng, T.F.; Russo, C.L. Mucin gene expression in immortalized human corneal-limbal and conjunctival epithelial cell lines. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2496–2506. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weng, J.; Trinh, S.; Lee, R.; Metwale, R.; Sharma, A. Impact of High Glucose on Ocular Surface Glycocalyx Components: Implications for Diabetes-Associated Ocular Surface Damage. Int. J. Mol. Sci. 2022, 23, 14289. https://doi.org/10.3390/ijms232214289
Weng J, Trinh S, Lee R, Metwale R, Sharma A. Impact of High Glucose on Ocular Surface Glycocalyx Components: Implications for Diabetes-Associated Ocular Surface Damage. International Journal of Molecular Sciences. 2022; 23(22):14289. https://doi.org/10.3390/ijms232214289
Chicago/Turabian StyleWeng, Judy, Steven Trinh, Rachel Lee, Rana Metwale, and Ajay Sharma. 2022. "Impact of High Glucose on Ocular Surface Glycocalyx Components: Implications for Diabetes-Associated Ocular Surface Damage" International Journal of Molecular Sciences 23, no. 22: 14289. https://doi.org/10.3390/ijms232214289
APA StyleWeng, J., Trinh, S., Lee, R., Metwale, R., & Sharma, A. (2022). Impact of High Glucose on Ocular Surface Glycocalyx Components: Implications for Diabetes-Associated Ocular Surface Damage. International Journal of Molecular Sciences, 23(22), 14289. https://doi.org/10.3390/ijms232214289