
Aldehyde Accumulation in Aged Alcoholic Beer: Addressing Acetaldehyde Impacts on Upper Aerodigestive Tract Cancer Risks


 , ,
, ,  and
and 
Abstract
1. Introduction
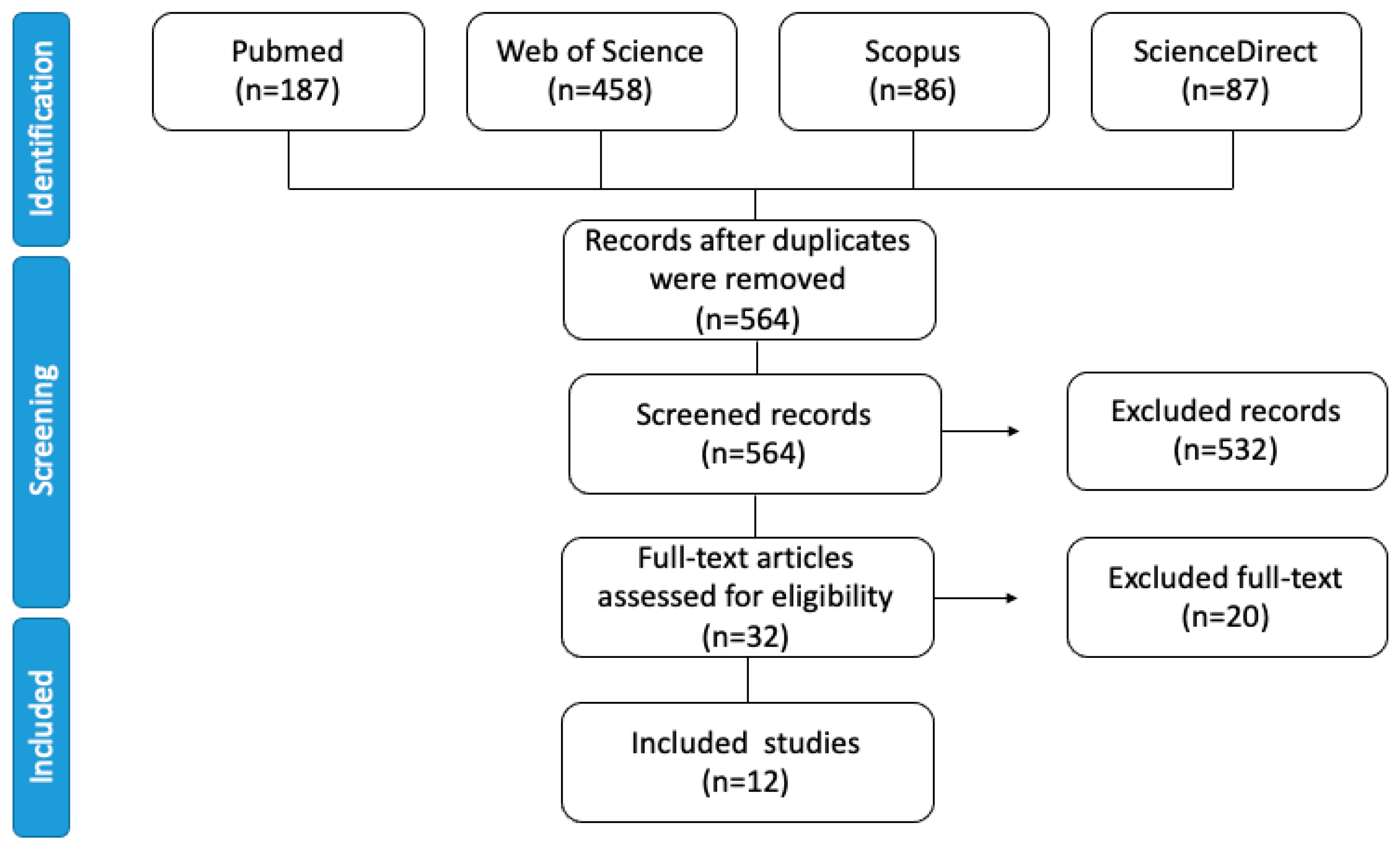
2. Systematic Literature Search Methods
2.1. Focus Questions
2.2. Research Strategy
2.3. Selection Criteria
3. Results
3.1. First Visual Approaches to the Dataset
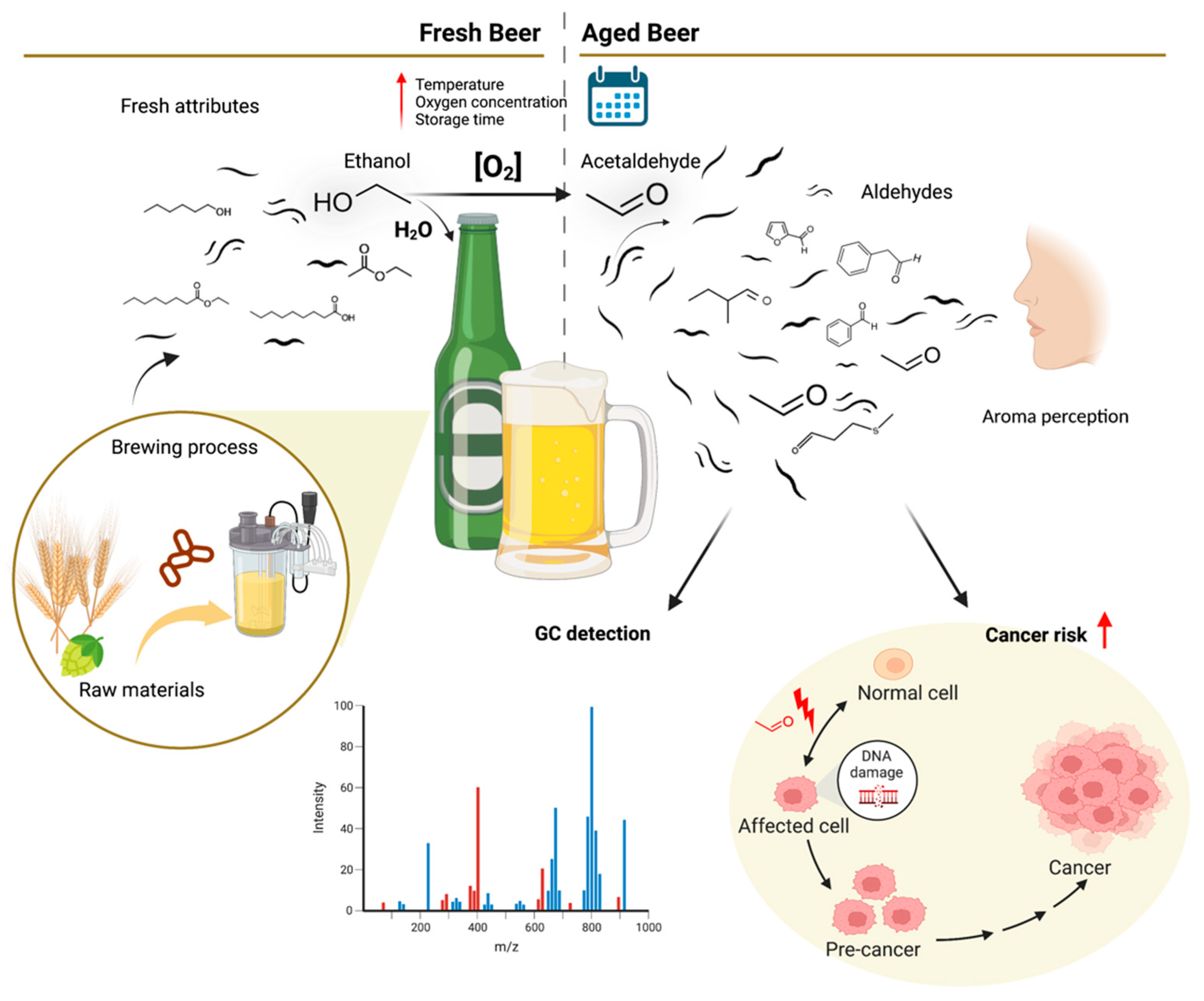
3.2. Chemical Composition of Fresh Beers
3.2.1. Non-Volatile Compounds
3.2.2. Volatile Compounds

{kind=link}
{kind=link}
{kind=link}
| Compound Class | Compound | Molecular Structure | Flavor Threshold (mg·L−1) | Boiling Point (°C) | Flavor/Aroma Description | Reference | |
| Alcohols | Hexan-1-ol |  | - | 157 | Herbaceous, greasy | [26,27] | |
| 3-Methyl-1-butanol |  | 70 | 132 | Alcohol, banana | [24,26] | ||
| 2-Methyl-1-propanol |  | 200 | 108 | Alcohol, solvent | [24,26] | ||
| n-Propanol |  | 800 | 97.2 | Alcohol, sweet | [24,26] | ||
| Esters | Ethyl acetate |  | 21–30 | 77 | Fruity, solvent like | [25] | |
| Ethyl octanoate |  | - | 206–208 | Apple, banana, pineapple | [30] | ||
| Ethyl decanoate |  | - | 241–245 | Waxy, apple, grape | [30] | ||
| Isoamyl acetate |  | 0.6–1.2 | 142 | Banana, pear | [25] | ||
| Ethyl caproate |  | 0.17–0.21 | 166–168 | Apple, anise seed | [25] | ||
| Phenylethyl acetate |  | 3.8 | 238–239 | Roses, honey, sweet | [25] | ||
| Aldehydes | - | Acetaldehyde |  | 10–20 | 20.8 | Grassy | [26] |
| Fatty Acid Oxidation Products | Hexanal |  | 0.088–0.35 | 130 | Bitter, winey | [26] | |
| (E)-2-nonenal |  | 0.0001 | 188–190 | Cardboard, papery, cucumber | [21,26] | ||
| Maillard Reaction Products | Furfural |  | 15.157 | 162 | Caramel, bready, cooked meat | [26] | |
| 5-Hydroxymethylfurfural |  | 1.174 | 114–116 | Bready, caramel | [26] | ||
| Strecker Degradation Products | 2-Methylpropanal |  | 0.086 | 64 | Grainy, varnish, fruity | [26] | |
| 2-Methylbutanal |  | 0.045 | 90–93 | Almond, apple-like, malty | [26] | ||
| 3-Methylbutanal |  | 0.056 | 92.5 | Malty, chocolate, cherry, almond | [26] | ||
| Methional |  | 0.0042 | 165 | Cooked potatoes, worty | [26] | ||
| Phenylacetaldehyde |  | 0.105 | 195 | Hyacinth, flowery, roses | [26] | ||
| Benzaldehyde |  | 0.515 | 179 | Almond, cherry stone | [26] | ||
| Organic Acids | Caprylic acid |  | 5 | 239 | Faint, fruity-acid | [32,33] | |
| Lauric acid |  | 0.5 | 298.9 | - | [33] | ||
| Capric acid |  | 2 | 268.7 | Rancid | [32,33] | ||
| Nonanoic acid |  | 5 | 254 | Fatty | [32,33] | ||
3.3. Alcoholic-Beer Physicochemical Stability and Flavor
3.3.1. Microbiota Stability
3.3.2. Physical Parameters
3.3.3. Flavor Instability
3.4. Aldehyde Beer Generation Mechanisms
3.4.1. Release from Bound-State Aldehydes
3.4.2. Strecker Degradation
3.4.3. Unsaturated Fatty Acid Degradation
3.4.4. Maillard Reactions
3.4.5. Bitter Acid Degradation
3.4.6. Aldol Condensation
3.5. Strategies for Inhibiting Aldehyde Formation and Improving Beer Stability
3.6. Comparing Aging in Different Beer Varieties
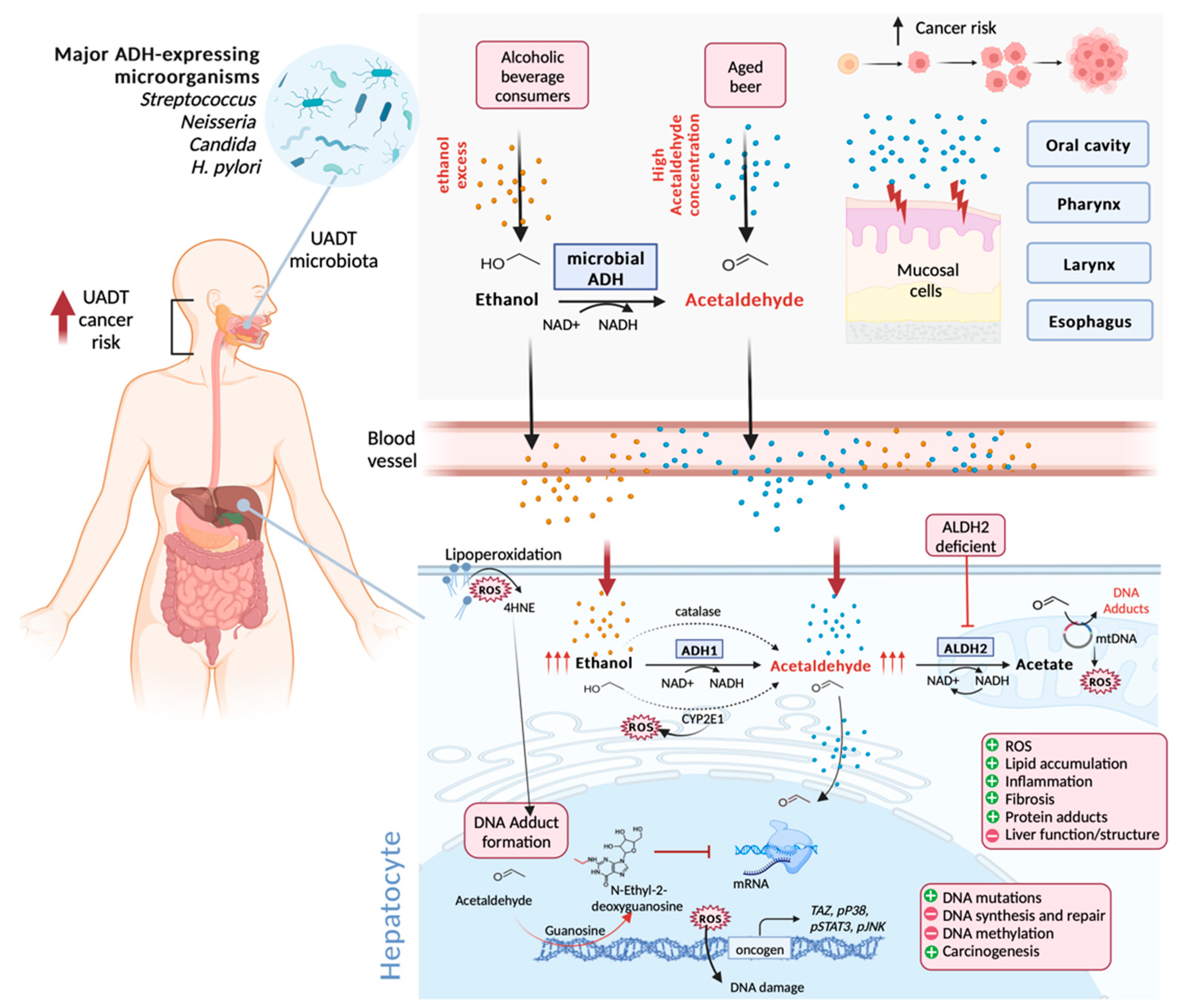
3.7. Beer Aging and Potential Health Effects
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schmelzle, A. The Beer Aroma Wheel. Brew. Sci. 2009, 62, 26. [Google Scholar]
- Baert, J.J.; De Clippeleer, J.; Hughes, P.S.; De Cooman, L.; Aerts, G. On the Origin of Free and Bound Staling Aldehydes in Beer. J. Agric. Food Chem. 2012, 60, 11449–11472. [Google Scholar] [CrossRef] [PubMed]
- Mascia, I.; Fadda, C.; Karabín, M.; Dostálek, P.; Del Caro, A. Aging of Craft Durum Wheat Beer Fermented with Sourdough Yeasts. LWT Food Sci. Technol. 2016, 65, 487–494. [Google Scholar] [CrossRef]
- Pascari, X.; Ramos, A.J.; Marín, S.; Sanchís, V. Mycotoxins and Beer. Impact of Beer Production Process on Mycotoxin Contamination. A Review. Food Res. Int. 2018, 103, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Vanderhaegen, B.; Delvaux, F.; Daenen, L.; Verachtert, H.; Delvaux, F.R. Aging Characteristics of Different Beer Types. Food Chem 2007, 103, 404–412. [Google Scholar] [CrossRef]
- Hashimoto, N. Oxidation of Higher Alcohols by Melanoidins in Beer. J. Inst. Brew. 1972, 78, 43–51. [Google Scholar] [CrossRef]
- Hashimoto, N.; Eshima, T. Composition and Pathway of Formation of Stale Aldehydes in Bottled Beer. J. Am. Soc. Brew. Chem. 1977, 35, 145–150. [Google Scholar] [CrossRef]
- Dennenlöhr, J.; Thörner, S.; Rettberg, N. Analysis of Hop-Derived Thiols in Beer Using On-Fiber Derivatization in Combination with HS-SPME and GC-MS/MS. J. Agric. Food Chem. 2020, 68, 15036–15047. [Google Scholar] [CrossRef]
- Lehnhardt, F.; Gastl, M.; Becker, T. Forced into Aging: Analytical Prediction of the Flavor-Stability of Lager Beer. A Review. Crit. Rev. Food Sci. Nutr. 2019, 59, 2642–2653. [Google Scholar] [CrossRef]
- Guido, L.F. Sulfites in Beer: Reviewing Regulation, Analysis and Role. Sci. Agric. 2016, 73, 189–197. [Google Scholar] [CrossRef]
- Nobis, A.; Kwasnicki, M.; Lehnhardt, F.; Hellwig, M.; Henle, T.; Becker, T.; Gastl, M. A Comprehensive Evaluation of Flavor Instability of Beer (Part 2): The Influence of De Novo Formation of Aging Aldehydes. Foods 2021, 10, 2668. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization Agents Classified by the IARC Monographs, Volumes 1–132. Available online: https://www.iarc.who.int/ (accessed on 31 October 2022).
- Lachenmeier, D.W.; Kanteres, F.; Rehm, J. Carcinogenicity of Acetaldehyde in Alcoholic Beverages: Risk Assessment Outside Ethanol Metabolism. Addiction 2009, 104, 533–550. [Google Scholar] [CrossRef] [PubMed]
- Lachenmeier, D.W.; Sohnius, E.M. The Role of Acetaldehyde Outside Ethanol Metabolism in the Carcinogenicity of Alcoholic Beverages: Evidence from a Large Chemical Survey. Food Chem. Toxicol. 2008, 46, 2903–2911. [Google Scholar] [CrossRef] [PubMed]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.A.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA Statement for Reporting Systematic Reviews and Meta-Analyses of Studies That Evaluate Healthcare Interventions: Explanation and Elaboration. BMJ 2009, 339, b2700. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Shamseer, L.; Clarke, M.; Ghersi, D.; Liberati, A.; Petticrew, M.; Shekelle, P.; Stewart, L.A. Preferred Reporting Items for Systematic Review and Meta-Analysis Protocols (PRISMA-P) 2015 Statement. Syst. Rev. 2015, 4, 1. [Google Scholar] [CrossRef]
- Piggott, J.R.; Paterson, A. Understanding Natural Flavors; Springer: Berlin/Heidelberg, Germany, 1994; ISBN 9781461521433. [Google Scholar]
- White, C.; Zainasheff, J. Yeast: The Practical Guide to Beer Fermentation; Brewers Publications: Milwaukee, WI, USA, 2010; ISBN 1938469062. [Google Scholar]
- Buiatti, S. Beer Composition: An Overview. Beer Health Dis. Prev. 2008, 213–225. [Google Scholar] [CrossRef]
- Anderson, H.E.; Santos, I.C.; Hildenbrand, Z.L.; Schug, K.A. A Review of the Analytical Methods Used for Beer Ingredient and Finished Product Analysis and Quality Control. Anal. Chim. Acta 2019, 1085, 1–20. [Google Scholar] [CrossRef]
- Gordon, R.; Power, A.; Chapman, J.; Chandra, S.; Cozzolino, D. A Review on the Source of Lipids and Their Interactions during Beer Fermentation That Affect Beer Quality. Fermentation 2018, 4, 89. [Google Scholar] [CrossRef]
- Vanderhaegen, B.; Neven, H.; Verachtert, H.; Derdelinckx, G. The Chemistry of Beer Aging—A Critical Review. Food Chem. 2006, 95, 357–381. [Google Scholar] [CrossRef]
- Gibson, B.; Aumala, V.; Heiniö, R.L.; Mikkelson, A.; Honkapää, K. Differential Evolution of Strecker and Non-Strecker Aldehydes during Aging of Pale and Dark Beers. J. Cereal. Sci. 2018, 83, 130–138. [Google Scholar] [CrossRef]
- Pires, E.; Brányik, T. Biochemistry of Beer Fermentation; Springer: Berlin/Heidelberg, Germany, 2015; ISBN 9783319151885. [Google Scholar]
- Verstrepen, K.J.; Derdelinckx, G.; Dufour, J.; Winderickx, J.; Thevelein, J.M.; Pretorius, I.S.; Delvaux, F.R.; Box, P.O.; Osmond, G.; Sa, A. Flavor-Active Esters: Adding Fruitiness to Beer. J. Biosci. Bioeng. 2003, 96, 110–118. [Google Scholar] [CrossRef]
- Olaniran, A.O.; Hiralal, L.; Mokoena, M.P.; Pillay, B. Flavour-Active Volatile Compounds in Beer: Production, Regulation and Control. J. Inst. Brew. 2017, 123, 13–23. [Google Scholar] [CrossRef]
- Rettberg, N.; Schubert, C.; Dennenlöhr, J.; Thörner, S.; Knoke, L.; Maxminer, J. Instability of Hop-Derived 2-Methylbutyl Isobutyrate during Aging of Commercial Pasteurized and Unpasteurized Ales. J. Am. Soc. Brew. Chem. 2020, 78, 175–184. [Google Scholar] [CrossRef]
- Parker, D.K. Beer: Production, Sensory Characteristics and Sensory Analysis; Woodhead Publishing Limited: Sawston, UK, 2012. [Google Scholar]
- Humia, B.V.; Santos, K.S.; Barbosa, A.M.; Sawata, M.; Mendonça, M.d.C.; Padilha, F.F. Beer Molecules and Its Sensory and Biological Properties: A Review. Molecules 2019, 24, 1568. [Google Scholar] [CrossRef]
- Gonzalez Viejo, C.; Fuentes, S.; Torrico, D.D.; Godbole, A.; Dunshea, F.R. Chemical Characterization of Aromas in Beer and Their Effect on Consumers Liking. Food Chem. 2019, 293, 479–485. [Google Scholar] [CrossRef]
- Hashimoto, N.; Kuroiwa, Y. Proposed Pathways for the Formation of Volatile Aldehydes during Storage of Bottled Beer. Proc. Annu. Meet. Am. Soc. Brew. Chem. 1975, 33, 104–111. [Google Scholar] [CrossRef]
- Burdock, G.A. Fenaroli’s Handbook of Flavor Ingredients, 6th ed.; CRC Press: Boca Raton, FL, USA, 2009; ISBN 9780429150838. [Google Scholar]
- Sigmund, E. Organoleptic Threshold Values of Some Organic Acids in Beer. J. Inst. Brew. 1973, 80, 162–163. [Google Scholar]
- Stewart, G.G. Beer Shelf Life and Stability. In The Stability and Shelf Life of Food; Elsevier: Amsterdam, The Netherlands, 2016; pp. 293–309. ISBN 9780081004357. [Google Scholar]
- Bekatorou, A.; Koutinas, A.A.; Psarianos, K.; Kanellaki, M. Low-Temperature Brewing by Freeze-Dried Immobilized Cells on Gluten Pellets. J. Agric. Food Chem. 2001, 49, 373–377. [Google Scholar] [CrossRef]
- Bamforth, C.W.; Lentini, A. The Flavor Instability of Beer. Beer 2009, 85–109. [Google Scholar] [CrossRef]
- Filipowska, W.; Jaskula-Goiris, B.; Ditrych, M.; Schlich, J.; De Rouck, G.; Aerts, G.; De Cooman, L. Determination of Optimal Sample Preparation for Aldehyde Extraction from Pale Malts and Their Quantification via Headspace Solid-Phase Microextraction Followed by Gas Chromatography and Mass Spectrometry. J. Chromatogr. A 2020, 1612, 460647. [Google Scholar] [CrossRef]
- Rodrigues, J.A.; Barros, A.S.; Carvalho, B.; Brandão, T.; Gil, A.M.; Ferreira, A.C.S. Evaluation of Beer Deterioration by Gas Chromatography-Mass Spectrometry/Multivariate Analysis: A Rapid Tool for Assessing Beer Composition. J. Chromatogr. A 2011, 1218, 990–996. [Google Scholar] [CrossRef]
- Guedes de Pinho, P.; Silva Ferreira, A.C. Role of Strecker Aldehydes on Beer Flavour Stability. Dev. Food Sci. 2006, 43, 529–532. [Google Scholar] [CrossRef]
- Bustillo Trueba, P.; Jaskula-Goiris, B.; Ditrych, M.; Filipowska, W.; De Brabanter, J.; De Rouck, G.; Aerts, G.; De Cooman, L.; De Clippeleer, J. Monitoring the Evolution of Free and Cysteinylated Aldehydes from Malt to Fresh and Forced Aged Beer. Food Res. Int. 2021, 140, 110049. [Google Scholar] [CrossRef] [PubMed]
- Atanasova, B.; Thomas-Danguin, T.; Langlois, D.; Nicklaus, S.; Chabanet, C.; Etiévant, P. Perception of Wine Fruity and Woody Notes: Influence of Peri-Threshold Odorants. Food Qual. Prefer. 2005, 16, 504–510. [Google Scholar] [CrossRef]
- Lehnhardt, F.; Steiner, J.; Gastl, M.; Becker, T. Prediction Power and Accuracy of Forced Ageing–Matching Sensory and Analytical Results for Lager Beer. BrewingScience 2018, 71, 39–48. [Google Scholar] [CrossRef]
- Baert, J.J.; De Clippeleer, J.; Bustillo Trueba, P.; Jaskula-Goiris, B.; De Rouck, G.; Aerts, G.; De Cooman, L. Exploring Aldehyde Release in Beer by 4-Vinylpyridine and the Effect of Cysteine Addition on the Beer’s Pool of Bound Aldehydes. J. Am. Soc. Brew. Chem. 2018, 76, 257–271. [Google Scholar] [CrossRef]
- Lehnhardt, F.; Nobis, A.; Skornia, A.; Becker, T.; Gastl, M. A Comprehensive Evaluation of Flavor Instability of Beer (Part 1): Influence of Release of Bound State Aldehydes. Foods 2021, 10, 2432. [Google Scholar] [CrossRef]
- Baert, J.J.; De Clippeleer, J.; De Cooman, L.; Aerts, G. Exploring the Binding Behavior of Beer Staling Aldehydes in Model Systems. J. Am. Soc. Brew. Chem. 2015, 73, 100–108. [Google Scholar] [CrossRef]
- Li, H.; Zhao, M.; Cui, C.; Sun, W.; Zhao, H. Antioxidant Activity and Typical Ageing Compounds: Their Evolutions and Relationships during the Storage of Lager Beers. Int. J. Food Sci. Technol. 2016, 51, 2026–2033. [Google Scholar] [CrossRef]
- Nagai, C.; Noda, K.; Kirihara, A.; Tomita, Y.; Murata, M. A Low-Molecular Weight Maillard Pigment from Beer Was Identified as Perlolyrine, a Maillard Reaction Product from Tryptophan. Food Sci. Technol. Res. 2019, 25, 81–88. [Google Scholar] [CrossRef]
- Ferreira, I.M.; Carvalho, D.O.; da Silva, M.G.; Guido, L.F. Gas-Diffusion Microextraction (GDME) Combined with Derivatization for Assessing Beer Staling Aldehydes: Validation and Application. Foods 2021, 10, 1704. [Google Scholar] [CrossRef] [PubMed]
- Ncube, S.; Dube, S.; Nindi, M.M. Determination of Volatile Compounds during Deterioration of African Opaque Beer Using a Stir Bar Sorptive Extraction Technique and Gas Chromatography-High Resolution Mass Spectrometry. Curr. Res. Food Sci. 2020, 3, 256–267. [Google Scholar] [CrossRef] [PubMed]
- Caballero, I.; Blanco, C.A.; Porras, M. Iso-α-Acids, Bitterness and Loss of Beer Quality during Storage. Trends Food Sci. Technol. 2012, 26, 21–30. [Google Scholar] [CrossRef]
- de Clippeleer, J.; de Rouck, G.; de Cooman, L.; Aerts, G. Influence of the Hopping Technology on the Storage-Induced Appearance of Staling Aldehydes in Beer. J. Inst. Brew. 2010, 116, 381–398. [Google Scholar] [CrossRef]
- Rizzi, G.P. The Strecker Degradation of Amino Acids: Newer Avenues for Flavor Formation. Food Rev. Int. 2008, 24, 416–435. [Google Scholar] [CrossRef]
- Aron, P.M.; Shellhammer, T.H. A Discussion of Polyphenols in Beer Physical and Flavour Stability. J. Inst. Brew. 2010, 116, 369–380. [Google Scholar] [CrossRef]
- Aguiar, D.; Pereira, A.C.; Marques, J.C. The Influence of Transport and Storage Conditions on Beer Stability—a Systematic Review. Food Bioprocess. Technol. 2022, 15, 1477–1494. [Google Scholar] [CrossRef]
- Wang, L.; Hong, K.; Agbaka, J.I.; Zhu, G.; Lv, C.; Ma, C. Application of UHPLC-Q/TOF-MS-Based Metabolomics Analysis for the Evaluation of Bitter-Tasting Krausen Metabolites during Beer Fermentation. J. Food Compos. Anal. 2021, 99, 103850. [Google Scholar] [CrossRef]
- Otter, G.E.; Taylor, L. Estimation and occurrence of acetaldehyde in beer. J. Inst. Brew. 1971, 77, 467–472. [Google Scholar] [CrossRef]
- Wietstock, P.C.; Kunz, T.; Methner, F.J. Relevance of Oxygen for the Formation of Strecker Aldehydes during Beer Production and Storage. J. Agric. Food Chem. 2016, 64, 8035–8044. [Google Scholar] [CrossRef]
- Attchelouwa, C.K.; N’guessan, F.K.; Marcotte, S.; Amoikon, T.L.S.; Charmel, M.; Djè, M.K. Characterisation of Volatile Compounds Associated to Sensory Changes during the Storage of Traditional Sorghum Beer by HS-GC/FID and SPME-GC/MS. J. Agric. Food Res. 2020, 2, 100088. [Google Scholar] [CrossRef]
- Schubert, C.; Lafontaine, S.; Dennenlöhr, J.; Thörner, S.; Rettberg, N. The Influence of Storage Conditions on the Chemistry and Flavor of Hoppy Ales. Food Chem. 2022, 395, 133616. [Google Scholar] [CrossRef] [PubMed]
- Spaggiari, G.; Cignarelli, A.; Sansone, A.; Baldi, M.; Santi, D. To Beer or Not to Beer: A Meta-Analysis of the Effects of Beer Consumption on Cardiovascular Health. PLoS ONE 2020, 15, e0233619. [Google Scholar] [CrossRef] [PubMed]
- de Gaetano, G.; Costanzo, S.; Di Castelnuovo, A.; Badimon, L.; Bejko, D.; Alkerwi, A.; Chiva-Blanch, G.; Estruch, R.; La Vecchia, C.; Panico, S.; et al. Effects of Moderate Beer Consumption on Health and Disease: A Consensus Document. Nutr. Metab. Cardiovasc. Dis. 2016, 26, 443–467. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, N.M.; Palmer, B.F.; Leiden, F.V.; Lee, R. Southwestern Internal Medicine Conference. Am. J. Med. Sci. 2001, 322, 88–102. [Google Scholar] [CrossRef]
- Thun, M. Alcohol Consumption and Mortality Among Middle-Aged and Elderly U.S Adults. N. Engl. J. Med. 1997, 337, 1705–1714. [Google Scholar] [CrossRef]
- Mukamal, K.J.; Kuller, L.H.; Fitzpatrick, A.L.; Longstreth, W.T.; Mittleman, M.A.; Siscovick, D.S. Prospective Study of Alcohol Consumption and Risk of Dementia in Older Adults. J. Am. Med. Assoc. 2003, 289, 1405–1413. [Google Scholar] [CrossRef]
- Das, U.N. Alcohol Consumption and Risk of Dementia. Lancet 2002, 360, 490. [Google Scholar] [CrossRef]
- Sánchez-Muniz, F.J.; Macho-González, A.; Garcimartín, A.; Santos-López, J.A.; Benedí, J.; Bastida, S.; González-Muñoz, M.J. The Nutritional Components of Beer and Its Relationship with Neurodegeneration and Alzheimer’s Disease. Nutrients 2019, 11, 1558. [Google Scholar] [CrossRef]
- Quesada-Molina, M.; Muñoz-Garach, A.; Tinahones, F.J.; Moreno-Indias, I. A New Perspective on the Health Benefits of Moderate Beer Consumption: Involvement of the Gut Microbiota. Metabolites 2019, 9, 272. [Google Scholar] [CrossRef]
- Osorio-Paz, I.; Brunauer, R.; Alavez, S. Beer and Its Non-Alcoholic Compounds in Health and Disease. Crit. Rev. Food Sci. Nutr. 2020, 60, 3492–3505. [Google Scholar] [CrossRef] [PubMed]
- Brooks, P.J.; Zakhari, S. Acetaldehyde and the Genome: Beyond Nuclear DNA Adducts and Carcinogenesis. Environ. Mol. Mutagen. 2010, 405, 391–405. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.; Zhang, J.; Shi, S.; Gong, D.; Lu, X.; Cheang, I.; Zhang, H.; Li, X. Association of Aldehyde Exposure with Cardiovascular Disease. Ecotoxicol. Environ. Saf. 2020, 206, 111385. [Google Scholar] [CrossRef] [PubMed]
- Rumgay, H.; Shield, K.; Charvat, H.; Ferrari, P.; Sornpaisarn, B.; Obot, I.; Islami, F.; Lemmens, V.E.P.P.; Rehm, J.; Soerjomataram, I. Global Burden of Cancer in 2020 Attributable to Alcohol Consumption: A Population-Based Study. Lancet Oncol. 2021, 22, 1071–1080. [Google Scholar] [CrossRef]
- Burton, R.; Sheron, N. No Level of Alcohol Consumption Improves Health. Lancet 2018, 392, 987–988. [Google Scholar] [CrossRef]
- Fernández-Solà, J. Cardiovascular Risks and Benefits of Moderate and Heavy Alcohol Consumption. Nat. Rev. Cardiol. 2015, 12, 576–587. [Google Scholar] [CrossRef]
- Shin, M.J.; Cho, Y.; Smith, G.D. Alcohol Consumption, Aldehyde Dehydrogenase 2 Gene Polymorphisms, and Cardiovascular Health in Korea. Yonsei Med J. 2017, 58, 689–696. [Google Scholar] [CrossRef]
- Wu, S.; Zhu, W.; Thompson, P.; Hannun, Y.A. Evaluating Intrinsic and Non-Intrinsic Cancer Risk Factors. Nat. Commun. 2018, 9, 3490. [Google Scholar] [CrossRef]
- Ismahil, M.A.; Hamid, T.; Haberzettl, P.; Gu, Y.; Chandrasekar, B.; Srivastava, S.; Bhatnagar, A.; Prabhu, S.D. Chronic Oral Exposure to the Aldehyde Pollutant Acrolein Induces Dilated Cardiomyopathy. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, 2050–2060. [Google Scholar] [CrossRef]
- Grootveld, M.; Percival, B.C.; Leenders, J.; Wilson, P.B. Potential Adverse Public Health Effects Afforded by the Ingestion of Dietary Lipid Oxidation Product Toxins: Significance of Fried Food Sources. Nutrients 2020, 12, 974. [Google Scholar] [CrossRef]
- Salaspuro, M. Acetaldehyde and Gastric Cancer. J. Dig. Dis. 2011, 12, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Theruvathu, J.A.; Jaruga, P.; Nath, R.G.; Dizdaroglu, M.; Brooks, P.J. Polyamines Stimulate the Formation of Mutagenic 1,N2-Propanodeoxyguanosine Adducts from Acetaldehyde. Nucleic Acids Res. 2005, 33, 3513–3520. [Google Scholar] [CrossRef] [PubMed]
- Feron, V.J.; Til, H.P.; de Vrijer, F.; Woutersen, R.A.; Cassee, F.R.; van Bladeren, P.J. Aldehydes: Occurrence, Carcinogenic Potential, Mechanism of Action and Risk Assessment. Mutat. Res./Genet. Toxicol. 1991, 259, 363–385. [Google Scholar] [CrossRef]
- Salaspuro, M. Acetaldehyde as a Common Denominator and Cumulative Carcinogen in Digestive Tract Cancers. Scand. J. Gastroenterol. 2009, 44, 912–925. [Google Scholar] [CrossRef] [PubMed]
- Eisenbrand, G.; Baum, M.; Cartus, A.T.; Diel, P.; Engel, K.-H.; Engeli, B.; Epe, B.; Grune, T.; Guth, S.; Haller, D.; et al. Salivary Nitrate/Nitrite and Acetaldehyde in Humans: Potential Combination Effects in the Upper Gastrointestinal Tract and Possible Consequences for the in Vivo Formation of N-Nitroso Compounds—A Hypothesis. Arch. Toxicol. 2022, 96, 1905–1914. [Google Scholar] [CrossRef]
- Salaspuro, M. Interrelationship between Alcohol, Smoking, Acetaldehyde and Cancer. Novartis Found. Symp. 2007, 285, 80–89. [Google Scholar] [CrossRef]
- Hyun, J.; Han, J.; Lee, C.; Yoon, M.; Jung, Y. Pathophysiological Aspects of Alcohol Metabolism in the Liver. Int. J. Mol. Sci. 2021, 22, 5717. [Google Scholar] [CrossRef]
- Nieminen, M.T.; Salaspuro, M. Local Acetaldehyde—An Essential Role in Alcohol-Related Upper Gastrointestinal Tract Carcinogenesis. Cancers 2018, 10, 11. [Google Scholar] [CrossRef]
- Yu, H.S.; Oyama, T.; Isse, T.; Kitagawa, K.; Pham, T.T.P.; Tanaka, M.; Kawamoto, T. Formation of Acetaldehyde-Derived DNA Adducts Due to Alcohol Exposure. Chem. Biol. Interact. 2010, 188, 367–375. [Google Scholar] [CrossRef]
- Druesne-Pecollo, N.; Tehard, B.; Mallet, Y.; Gerber, M.; Norat, T.; Hercberg, S.; Latino-Martel, P. Alcohol and Genetic Polymorphisms: Effect on Risk of Alcohol-Related Cancer. Lancet Oncol. 2009, 10, 173–180. [Google Scholar] [CrossRef]
- Cheng, X.; Blumenthal, R.M. Mammalian DNA Methyltransferases: A Structural Perspective. Structure 2008, 16, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Seitz, H.K.; Stickel, F. Molecular Mechanisms of Alcohol-Mediated Carcinogenesis. Nat. Rev. Cancer 2007, 7, 599–612. [Google Scholar] [CrossRef] [PubMed]
- Seo, W.; Gao, Y.; He, Y.; Sun, J.; Xu, H.; Feng, D.; Park, S.H.; Cho, Y.-E.; Guillot, A.; Ren, T.; et al. ALDH2 Deficiency Promotes Alcohol-Associated Liver Cancer by Activating Oncogenic Pathways via Oxidized DNA-Enriched Extracellular Vesicles. J. Hepatol. 2019, 71, 1000–1011. [Google Scholar] [CrossRef] [PubMed]
- Zakhari, S. Overview: How Is Alcohol Metabolized by the Body? Alcohol Res. Health 2006, 29, 245–254. [Google Scholar] [PubMed]
- Rumgay, H.; Murphy, N.; Ferrari, P.; Soerjomataram, I. Alcohol and Cancer: Epidemiology and Biological Mechanisms. Nutrients 2021, 13, 3173. [Google Scholar] [CrossRef] [PubMed]
- Leung, T.-M.; Nieto, N. CYP2E1 and Oxidant Stress in Alcoholic and Non-Alcoholic Fatty Liver Disease. J. Hepatol. 2013, 58, 395–398. [Google Scholar] [CrossRef]
- Stornetta, A.; Guidolin, V.; Balbo, S. Alcohol-Derived Acetaldehyde Exposure in the Oral Cavity. Cancers 2018, 10, 20. [Google Scholar] [CrossRef]
- Farah, C.S.; Jessri, M.; Currie, S.; Alnuaimi, A.; Yap, T.; McCullough, M.J. Aetiology of Oral Cavity Cancer. In Contemporary Oral Oncology; Springer: Berlin/Heidelberg, Germany, 2017; pp. 31–76. [Google Scholar]
- Salaspuro, M. Local Acetaldehyde: Its Key Role in Alcohol-Related Oropharyngeal Cancer. Visc. Med. 2020, 36, 167–174. [Google Scholar] [CrossRef]



| Order | Inclusion Criteria | Exclusion Criteria |
|---|---|---|
| 1 | Articles in English | Articles not in English |
| 2 | Beer or staling or storage effects | Reviews, short communications, thesis, and books |
| 3 | Volatile compound monitoring | No volatile compound monitoring |
| 4 | Beers analyzed by gas chromatography | Articles that evaluated other beverages or did not use gas chromatography as the analytical method for aldehyde identification |
| Beer Variety | Sample Treatment Applied to Assess Aldehyde Formation Effects | Storage Conditions 1 | Physicochemical Parameters | Aldehyde | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| pH | [O2] (mg·L−1) | Soluble N | 2-Methylpropanal (µg·L−1) | 3-Methylbutanal (µg·L−1) | Furfural (µg·L−1) | Reference | |||||||
| Temperature | Time | Fresh 2 | Aged 3 | Fresh 2 | Aged 3 | Fresh 2 | Aged 3 | ||||||
| Lager | Malt proteolysis | 20–40 °C | 4 d–9 m | 4.39–4.57 | 0.01–0.08 | 569–731 (mg/100 g malt d.m.) | 12.074 | 277.308 | 8.350 | 7.231 | 1.237 | 6.615 | [45] |
| Malt proteolysis | 20–40 °C | 4 d–9 m | 4.39–4.57 | 0.01–0.08 | 569–731 (mg/100 g malt d.m.) | 12.074 | 277.308 | 8.350 | 7.231 | 1.237 | 6.615 | [11] | |
| Cysteine addition | 0–30 °C | 3 m | 4.12–4.37 | Nd | Nd | ≅7 | ≅35 | ≅d | ≅d1 | ≅0 | ≅200 | [44] | |
| Oxygen exposure | 28 °C | 0 d–30 w | 4.41 | 0.0531–29.908 | Nd | Nd | Nd | 4.9 | 7.5 | 169.8 | 474.5 | [57] | |
| Different brands | 4–25 °C | 2 d–6 m | Nd | Nd | Nd | Nd | Nd | 1.40 | 5.62 | 53.50 | 210.75 | [47] | |
| Ales and lagers | Different wort and beer colors | 20–37 °C | 7 d–8 m | Nd | Nd | Nd | ≅5 | ≅70 | ≅5 | ≅150 | <1 | ≅20 | [23] |
| Ale | Nd | 4–40 °C | 4 d–24 w | 4.4–4.9 | Nd | Nd | Nd | Nd | Nd | Nd | Nd | Nd | [27] |
| Craft Durum Wheat | Yeast Strain? | 8–28 °C | 2 m–6 m | 4.44–4.76 | Nd | Nd | 11 | 4.18 | 48.49 | 92.52 | 0.17 | 0.32 | [3] |
| Blond/Amber | Different brewing? | 30 °C | 0 d–3 m | 4.30–4.38 | 0.106–0.186 | 60.5–76.1 (mg/100 mL wort) | ≅0 | ≅112 | ≅0 | ≅5 | ≅0 | ≅150 | [41] |
| Tchapalo | Different Brewer? | 4–30 °C | 4 d–6 d | Nd | Nd | Nd | Nd | Nd | Nd | Nd | Nd | Nd | [58] |
| Opaque | Different Brewer? | Room temperature | 1 d–7 d | Nd | Nd | Nd | Nd | Nd | Nd | Nd | Nd 4 | Nd | [50] |
| Nd | Nd | 4–37 °C | 7 d–6 m | Nd | Nd | Nd | >5.7 | ≅26.09 | >4.2 | ≅12 | 154.5 | 800 | [49] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreira, M.T.G.; Pereira, P.R.; Aquino, A.; Conte-Junior, C.A.; Paschoalin, V.M.F. Aldehyde Accumulation in Aged Alcoholic Beer: Addressing Acetaldehyde Impacts on Upper Aerodigestive Tract Cancer Risks. Int. J. Mol. Sci. 2022, 23, 14147. https://doi.org/10.3390/ijms232214147
Moreira MTG, Pereira PR, Aquino A, Conte-Junior CA, Paschoalin VMF. Aldehyde Accumulation in Aged Alcoholic Beer: Addressing Acetaldehyde Impacts on Upper Aerodigestive Tract Cancer Risks. International Journal of Molecular Sciences. 2022; 23(22):14147. https://doi.org/10.3390/ijms232214147
Chicago/Turabian StyleMoreira, Mariana Toledo Gonçalves, Patricia Ribeiro Pereira, Adriano Aquino, Carlos Adam Conte-Junior, and Vania Margaret Flosi Paschoalin. 2022. "Aldehyde Accumulation in Aged Alcoholic Beer: Addressing Acetaldehyde Impacts on Upper Aerodigestive Tract Cancer Risks" International Journal of Molecular Sciences 23, no. 22: 14147. https://doi.org/10.3390/ijms232214147
APA StyleMoreira, M. T. G., Pereira, P. R., Aquino, A., Conte-Junior, C. A., & Paschoalin, V. M. F. (2022). Aldehyde Accumulation in Aged Alcoholic Beer: Addressing Acetaldehyde Impacts on Upper Aerodigestive Tract Cancer Risks. International Journal of Molecular Sciences, 23(22), 14147. https://doi.org/10.3390/ijms232214147