

Inflammatory Cytokines That Enhance Antigen Responsiveness of Naïve CD8+ T Lymphocytes Modulate Chromatin Accessibility of Genes Impacted by Antigen Stimulation

Abstract
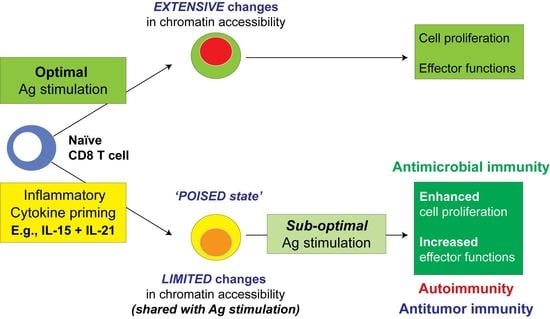
1. Introduction
2. Results
2.1. Comparison of ATACseq Peaks Modulated by Cytokine Priming and Ag Stimulation
2.2. Comparison of ATACseq Peaks Modulated by Cytokine Priming and Ag Stimulation
2.3. Chromatin Accessibility in Ag-Stimulated Cells
2.4. Chromatin Accessibility in Cytokine-Primed Cells
2.5. Transcription Factor Binding Motifs Enriched in CytP and AgS Cells
2.6. Chromatin Accessibility of Genes Implicated in CD8+ T Cell Activation
2.7. Expression of T Cell Activation Genes in CytP and AgS Cells
3. Discussion
4. Materials and Methods
4.1. Mice, Peptides and Cytokines
4.2. Cell Purification and Stimulation
4.3. ATAC Sequencing
4.4. ATACseq Data Analysis
4.5. Gene Expression and Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harty, J.T.; Tvinnereim, A.R.; White, D.W. CD8+ T cell effector mechanisms in resistance to infection. Annu. Rev. Immunol. 2000, 18, 275–308. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, S.A.; Restifo, N.P.; Yang, J.C.; Morgan, R.A.; Dudley, M.E. Adoptive cell transfer: A clinical path to effective cancer immunotherapy. Nat. Rev. 2008, 8, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, A.H. Mechanisms of costimulation. Immunol. Rev. 2009, 229, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Medzhitov, R. Regulation of adaptive immunity by the innate immune system. Science 2010, 327, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Luster, A.D. The role of chemokines in linking innate and adaptive immunity. Curr. Opin. Immunol. 2002, 14, 129–135. [Google Scholar] [CrossRef]
- Hoebe, K.; Janssen, E.; Beutler, B. The interface between innate and adaptive immunity. Nat. Immunol. 2004, 5, 971–974. [Google Scholar] [CrossRef]
- Moretta, A.; Marcenaro, E.; Parolini, S.; Ferlazzo, G.; Moretta, L. NK cells at the interface between innate and adaptive immunity. Cell Death Differ. 2008, 15, 226–233. [Google Scholar]
- Jain, A.; Pasare, C. Innate Control of Adaptive Immunity: Beyond the Three-Signal Paradigm. J. Immunol. 2017, 198, 3791–3800. [Google Scholar] [CrossRef]
- Pulendran, B.; Ahmed, R. Translating innate immunity into immunological memory: Implications for vaccine development. Cell 2006, 124, 849–863. [Google Scholar] [CrossRef]
- von Herrath, M.G.; Fujinami, R.S.; Whitton, J.L. Microorganisms and autoimmunity: Making the barren field fertile? Nat. Rev. Microbiol. 2003, 1, 151–157. [Google Scholar] [CrossRef]
- Haring, J.S.; Badovinac, V.P.; Harty, J.T. Inflaming the CD8(+) T cell response. Immunity 2006, 25, 19–29. [Google Scholar] [PubMed]
- Curtsinger, J.M.; Schmidt, C.S.; Mondino, A.; Lins, D.C.; Kedl, R.M.; Jenkins, M.K.; Mescher, M.F. Inflammatory cytokines provide a third signal for activation of naive CD4+ and CD8+ T cells. J. Immunol. 1999, 162, 3256–3262. [Google Scholar] [PubMed]
- Curtsinger, J.M.; Johnson, C.M.; Mescher, M.F. CD8 T cell clonal expansion and development of effector function require prolonged exposure to antigen, costimulation, and signal 3 cytokine. J. Immunol. 2003, 171, 5165–5171. [Google Scholar] [PubMed]
- Kolumam, G.A.; Thomas, S.; Thompson, L.J.; Sprent, J.; Murali-Krishna, K. Type I interferons act directly on CD8 T cells to allow clonal expansion and memory formation in response to viral infection. J. Exp. Med. 2005, 202, 637–650. [Google Scholar] [PubMed]
- Curtsinger, J.M.; Valenzuela, J.O.; Agarwal, P.; Lins, D.; Mescher, M.F. Type I IFNs provide a third signal to CD8 T cells to stimulate clonal expansion and differentiation. J. Immunol. 2005, 174, 4465–4469. [Google Scholar] [CrossRef]
- Morishima, N.; Owaki, T.; Asakawa, M.; Kamiya, S.; Mizuguchi, J.; Yoshimoto, T. Augmentation of effector CD8+ T cell generation with enhanced granzyme B expression by IL-27. J. Immunol. 2005, 175, 1686–1693. [Google Scholar] [CrossRef]
- Le Bon, A.; Durand, V.; Kamphuis, E.; Thompson, C.; Bulfone-Paus, S.; Rossmann, C.; Kalinke, U.; Tough, D.F. Direct stimulation of T cells by type I IFN enhances the CD8+ T cell response during cross-priming. J. Immunol. 2006, 176, 4682–4689. [Google Scholar] [CrossRef]
- Gagnon, J.; Ramanathan, S.; Leblanc, C.; Ilangumaran, S. Regulation of IL-21 signaling by suppressor of cytokine signaling-1 (SOCS1) in CD8(+) T lymphocytes. Cell Signal. 2007, 19, 806–816. [Google Scholar]
- Gagnon, J.; Ramanathan, S.; Leblanc, C.; Cloutier, A.; McDonald, P.P.; Ilangumaran, S. IL-6, in Synergy with IL-7 or IL-15, Stimulates TCR-Independent Proliferation and Functional Differentiation of CD8+ T Lymphocytes. J. Immunol. 2008, 180, 7958–7968. [Google Scholar] [CrossRef]
- Zeng, R.; Spolski, R.; Finkelstein, S.E.; Oh, S.; Kovanen, P.E.; Hinrichs, C.S.; Pise-Masison, C.A.; Radonovich, M.F.; Brady, J.N.; Restifo, N.P.; et al. Synergy of IL-21 and IL-15 in regulating CD8+ T cell expansion and function. J. Exp. Med. 2005, 201, 139–148. [Google Scholar] [CrossRef]
- Sawa, Y.; Arima, Y.; Ogura, H.; Kitabayashi, C.; Jiang, J.J.; Fukushima, T.; Kamimura, D.; Hirano, T.; Murakami, M. Hepatic interleukin-7 expression regulates T cell responses. Immunity 2009, 30, 447–457. [Google Scholar] [CrossRef]
- Mattei, F.; Schiavoni, G.; Belardelli, F.; Tough, D.F. IL-15 is expressed by dendritic cells in response to type I IFN, double-stranded RNA, or lipopolysaccharide and promotes dendritic cell activation. J. Immunol. 2001, 167, 1179–1187. [Google Scholar] [CrossRef] [PubMed]
- Ramanathan, S.; Dubois, S.; Chen, X.L.; Leblanc, C.; Ohashi, P.S.; Ilangumaran, S. Exposure to IL-15 and IL-21 enables autoreactive CD8 T cells to respond to weak antigens and cause disease in a mouse model of autoimmune diabetes. J. Immunol. 2011, 186, 5131–5141. [Google Scholar] [PubMed]
- Rodriguez, G.M.; D’Urbano, D.; Bobbala, D.; Chen, X.L.; Yeganeh, M.; Ramanathan, S.; Ilangumaran, S. SOCS1 Prevents Potentially Skin-Reactive Cytotoxic T Lymphocytes from Gaining the Ability to Cause Inflammatory Lesions. J. Investig. Dermatol. 2013, 133, 2013–2022. [Google Scholar] [CrossRef] [PubMed]
- Richer, M.J.; Nolz, J.C.; Harty, J.T. Pathogen-specific inflammatory milieux tune the antigen sensitivity of CD8(+) T cells by enhancing T cell receptor signaling. Immunity 2013, 38, 140–152. [Google Scholar] [CrossRef]
- Ramanathan, S.; Gagnon, J.; Dubois, S.; Forand-Boulerice, M.; Richter, M.V.; Ilangumaran, S. Cytokine Synergy in Antigen-Independent Activation and Priming of Naive CD8+ T Lymphocytes. Crit. Rev. Immunol. 2009, 29, 219–239. [Google Scholar]
- Gagnon, J.; Chen, X.L.; Forand-Boulerice, M.; Leblanc, C.; Raman, C.; Ramanathan, S.; Ilangumaran, S. Increased antigen responsiveness of naive CD8 T cells exposed to IL-7 and IL-21 is associated with decreased CD5 expression. Immunol. Cell Biol. 2010, 88, 451–460. [Google Scholar] [CrossRef]
- Bezbradica, J.S.; Medzhitov, R. Integration of cytokine and heterologous receptor signaling pathways. Nat. Immunol. 2009, 10, 333–339. [Google Scholar] [CrossRef]
- Shi, M.; Lin, T.H.; Appell, K.C.; Berg, L.J. Janus-kinase-3-dependent signals induce chromatin remodeling at the Ifng locus during T helper 1 cell differentiation. Immunity 2008, 28, 763–773. [Google Scholar] [CrossRef]
- Zhu, J.; Cote-Sierra, J.; Guo, L.; Paul, W.E. Stat5 activation plays a critical role in Th2 differentiation. Immunity 2003, 19, 739–748. [Google Scholar]
- Saeki, K.; Fukuyama, S.; Ayada, T.; Nakaya, M.; Aki, D.; Takaesu, G.; Hanada, T.; Matsumura, Y.; Kobayashi, T.; Nakagawa, R.; et al. A major lipid raft protein raftlin modulates T cell receptor signaling and enhances th17-mediated autoimmune responses. J. Immunol. 2009, 182, 5929–5937. [Google Scholar] [CrossRef] [PubMed]
- Rellahan, B.L.; Jensen, J.P.; Howcroft, T.K.; Singer, D.S.; Bonvini, E.; Weissman, A.M. Elf-1 regulates basal expression from the T cell antigen receptor zeta-chain gene promoter. J. Immunol. 1998, 160, 2794–2801. [Google Scholar] [PubMed]
- Ohnuma, K.; Uchiyama, M.; Yamochi, T.; Nishibashi, K.; Hosono, O.; Takahashi, N.; Kina, S.; Tanaka, H.; Lin, X.; Dang, N.H.; et al. Caveolin-1 triggers T-cell activation via CD26 in association with CARMA1. J. Biol. Chem. 2007, 282, 10117–10131. [Google Scholar] [CrossRef] [PubMed]
- Nayar, R.; Enos, M.; Prince, A.; Shin, H.; Hemmers, S.; Jiang, J.K.; Klein, U.; Thomas, C.J.; Berg, L.J. TCR signaling via Tec kinase ITK and interferon regulatory factor 4 (IRF4) regulates CD8+ T-cell differentiation. Proc. Natl. Acad. Sci. USA 2012, 109, E2794–E2802. [Google Scholar] [CrossRef]
- Pfeifhofer, C.; Kofler, K.; Gruber, T.; Tabrizi, N.G.; Lutz, C.; Maly, K.; Leitges, M.; Baier, G. Protein kinase C theta affects Ca2+ mobilization and NFAT cell activation in primary mouse T cells. J. Exp. Med. 2003, 197, 1525–1535. [Google Scholar] [CrossRef]
- Ruland, J.; Duncan, G.S.; Elia, A.; del Barco Barrantes, I.; Nguyen, L.; Plyte, S.; Millar, D.G.; Bouchard, D.; Wakeham, A.; Ohashi, P.S.; et al. Bcl10 is a positive regulator of antigen receptor-induced activation of NF-kappaB and neural tube closure. Cell 2001, 104, 33–42. [Google Scholar] [CrossRef]
- Liu, Z.Z.; Wang, Z.L.; Choi, T.I.; Huang, W.T.; Wang, H.T.; Han, Y.Y.; Zhu, L.Y.; Kim, H.T.; Choi, J.H.; Lee, J.S.; et al. Chd7 Is Critical for Early T-Cell Development and Thymus Organogenesis in Zebrafish. Am. J. Pathol. 2018, 188, 1043–1058. [Google Scholar] [CrossRef]
- Finco, T.S.; Justice-Healy, G.E.; Patel, S.J.; Hamilton, V.E. Regulation of the human LAT gene by the Elf-1 transcription factor. BMC Mol. Biol 2006, 7, 4. [Google Scholar] [CrossRef]
- Riera-Sans, L.; Behrens, A. Regulation of alphabeta/gammadelta T cell development by the activator protein 1 transcription factor c-Jun. J. Immunol. 2007, 178, 5690–5700. [Google Scholar] [CrossRef]
- Sallusto, F.; Kremmer, E.; Palermo, B.; Hoy, A.; Ponath, P.; Qin, S.; Forster, R.; Lipp, M.; Lanzavecchia, A. Switch in chemokine receptor expression upon TCR stimulation reveals novel homing potential for recently activated T cells. Eur. J. Immunol. 1999, 29, 2037–2045. [Google Scholar] [CrossRef]
- Schaller, M.A.; Kallal, L.E.; Lukacs, N.W. A key role for CC chemokine receptor 1 in T-cell-mediated respiratory inflammation. Am. J. Pathol. 2008, 172, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.W.; Kim, H.G.; Perry, C.J.; Kaech, S.M. CCR7 expression alters memory CD8 T-cell homeostasis by regulating occupancy in IL-7- and IL-15-dependent niches. Proc. Natl. Acad. Sci. USA 2016, 113, 8278–8283. [Google Scholar] [CrossRef]
- Acuto, O.; Michel, F. CD28-mediated co-stimulation: A quantitative support for TCR signalling. Nat. Rev. Immunol. 2003, 3, 939–951. [Google Scholar] [CrossRef]
- Zhang, Y.; Blattman, J.N.; Kennedy, N.J.; Duong, J.; Nguyen, T.; Wang, Y.; Davis, R.J.; Greenberg, P.D.; Flavell, R.A.; Dong, C. Regulation of innate and adaptive immune responses by MAP kinase phosphatase 5. Nature 2004, 430, 793–797. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Geng, J.; Shi, B.; Liu, Z.; Wang, Y.H.; Stevens, A.C.; Sprout, S.L.; Yao, M.; Wang, H.; Hu, H. Cutting Edge: Foxp1 Controls Naive CD8+ T Cell Quiescence by Simultaneously Repressing Key Pathways in Cellular Metabolism and Cell Cycle Progression. J. Immunol. 2016, 196, 3537–3541. [Google Scholar] [CrossRef]
- Ju, S.; Zhu, Y.; Liu, L.; Dai, S.; Li, C.; Chen, E.; He, Y.; Zhang, X.; Lu, B. Gadd45b and Gadd45g are important for anti-tumor immune responses. Eur. J. Immunol. 2009, 39, 3010–3018. [Google Scholar] [CrossRef] [PubMed]
- Sumida, H.; Cyster, J.G. G-Protein Coupled Receptor 18 Contributes to Establishment of the CD8 Effector T Cell Compartment. Front. Immunol. 2018, 9, 660. [Google Scholar] [CrossRef] [PubMed]
- Schluns, K.S.; Williams, K.; Ma, A.; Zheng, X.X.; Lefrancois, L. Cutting edge: Requirement for IL-15 in the generation of primary and memory antigen-specific CD8 T cells. J. Immunol. 2002, 168, 4827–4831. [Google Scholar] [CrossRef] [PubMed]
- Harker, J.A.; Wong, K.A.; Dolgoter, A.; Zuniga, E.I. Cell-Intrinsic gp130 Signaling on CD4+ T Cells Shapes Long-Lasting Antiviral Immunity. J. Immunol. 2015, 195, 1071–1081. [Google Scholar] [CrossRef] [PubMed]
- Charlton, J.J.; Chatzidakis, I.; Tsoukatou, D.; Boumpas, D.T.; Garinis, G.A.; Mamalaki, C. Programmed death-1 shapes memory phenotype CD8 T cell subsets in a cell-intrinsic manner. J. Immunol. 2013, 190, 6104–6114. [Google Scholar] [CrossRef]
- Willinger, T.; Freeman, T.; Herbert, M.; Hasegawa, H.; McMichael, A.J.; Callan, M.F. Human naive CD8 T cells down-regulate expression of the WNT pathway transcription factors lymphoid enhancer binding factor 1 and transcription factor 7 (T cell factor-1) following antigen encounter in vitro and in vivo. J. Immunol. 2006, 176, 1439–1446. [Google Scholar] [CrossRef] [PubMed]
- MaruYama, T. TGF-beta-induced IkappaB-zeta controls Foxp3 gene expression. Biochem. Biophys. Res. Commun. 2015, 464, 586–589. [Google Scholar] [CrossRef] [PubMed]
- Meisel, M.; Hermann-Kleiter, N.; Hinterleitner, R.; Gruber, T.; Wachowicz, K.; Pfeifhofer-Obermair, C.; Fresser, F.; Leitges, M.; Soldani, C.; Viola, A.; et al. The kinase PKCalpha selectively upregulates interleukin-17A during Th17 cell immune responses. Immunity 2013, 38, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Lee, Y.C.; Brown, C.; Zhang, W.; Usherwood, E.; Noelle, R.J. Dissecting the role of retinoic acid receptor isoforms in the CD8 response to infection. J. Immunol. 2014, 192, 3336–3344. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, P.; Kurtulus, S.; Wojciechowski, S.; Sholl, A.; Hoebe, K.; Morris, S.C.; Finkelman, F.D.; Grimes, H.L.; Hildeman, D.A. STAT5 is critical to maintain effector CD8+ T cell responses. J. Immunol. 2010, 185, 2116–2124. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Zhang, N. Transforming growth factor-beta signaling is constantly shaping memory T-cell population. Proc. Natl. Acad. Sci. USA 2015, 112, 11013–11017. [Google Scholar] [CrossRef]
- Nishimura, H.; Yajima, T.; Muta, H.; Podack, E.R.; Tani, K.; Yoshikai, Y. A novel role of CD30/CD30 ligand signaling in the generation of long-lived memory CD8+ T cells. J. Immunol. 2005, 175, 4627–4634. [Google Scholar] [CrossRef]
- Ritthipichai, K.; Haymaker, C.L.; Martinez, M.; Aschenbrenner, A.; Yi, X.; Zhang, M.; Kale, C.; Vence, L.M.; Roszik, J.; Hailemichael, Y.; et al. Multifaceted Role of BTLA in the Control of CD8(+) T-cell Fate after Antigen Encounter. Clin. Cancer Res. 2017, 23, 6151–6164. [Google Scholar] [CrossRef]
- Shamim, M.; Nanjappa, S.G.; Singh, A.; Plisch, E.H.; LeBlanc, S.E.; Walent, J.; Svaren, J.; Seroogy, C.; Suresh, M. Cbl-b regulates antigen-induced TCR down-regulation and IFN-gamma production by effector CD8 T cells without affecting functional avidity. J. Immunol. 2007, 179, 7233–7243. [Google Scholar] [CrossRef]
- Soares, L.R.; Tsavaler, L.; Rivas, A.; Engleman, E.G. V7 (CD101) ligation inhibits TCR/CD3-induced IL-2 production by blocking Ca2+ flux and nuclear factor of activated T cell nuclear translocation. J. Immunol. 1998, 161, 209–217. [Google Scholar]
- DeBell, K.E.; Simhadri, V.R.; Mariano, J.L.; Borrego, F. Functional requirements for inhibitory signal transmission by the immunomodulatory receptor CD300a. BMC Immunol. 2012, 13, 23. [Google Scholar] [CrossRef] [PubMed]
- Bouguermouh, S.; Van, V.Q.; Martel, J.; Gautier, P.; Rubio, M.; Sarfati, M. CD47 expression on T cell is a self-control negative regulator of type 1 immune response. J. Immunol. 2008, 180, 8073–8082. [Google Scholar] [CrossRef] [PubMed]
- Chauvin, J.M.; Pagliano, O.; Fourcade, J.; Sun, Z.; Wang, H.; Sander, C.; Kirkwood, J.M.; Chen, T.H.; Maurer, M.; Korman, A.J.; et al. TIGIT and PD-1 impair tumor antigen-specific CD8(+) T cells in melanoma patients. J. Clin. Investig. 2015, 125, 2046–2058. [Google Scholar] [CrossRef] [PubMed]
- Waugh, K.A.; Leach, S.M.; Moore, B.L.; Bruno, T.C.; Buhrman, J.D.; Slansky, J.E. Molecular Profile of Tumor-Specific CD8+ T Cell Hypofunction in a Transplantable Murine Cancer Model. J. Immunol. 2016, 197, 1477–1488. [Google Scholar] [CrossRef] [PubMed]
- Kamimura, D.; Bevan, M.J. Endoplasmic reticulum stress regulator XBP-1 contributes to effector CD8+ T cell differentiation during acute infection. J. Immunol. 2008, 181, 5433–5441. [Google Scholar] [CrossRef]
- Ma, X.; Bi, E.; Lu, Y.; Su, P.; Huang, C.; Liu, L.; Wang, Q.; Yang, M.; Kalady, M.F.; Qian, J.; et al. Cholesterol Induces CD8(+) T Cell Exhaustion in the Tumor Microenvironment. Cell Metab. 2019, 30, 143–156 e145. [Google Scholar] [CrossRef]
- Kim, H.; Kim, T.; Jeong, B.C.; Cho, I.T.; Han, D.; Takegahara, N.; Negishi-Koga, T.; Takayanagi, H.; Lee, J.H.; Sul, J.Y.; et al. Tmem64 modulates calcium signaling during RANKL-mediated osteoclast differentiation. Cell Metab. 2013, 17, 249–260. [Google Scholar] [CrossRef]
- Bando, J.K.; Gilfillan, S.; Song, C.; McDonald, K.G.; Huang, S.C.; Newberry, R.D.; Kobayashi, Y.; Allan, D.S.J.; Carlyle, J.R.; Cella, M.; et al. The Tumor Necrosis Factor Superfamily Member RANKL Suppresses Effector Cytokine Production in Group 3 Innate Lymphoid Cells. Immunity 2018, 48, 1208–1219e1204. [Google Scholar] [CrossRef]
- Reed, N.P.; Henderson, M.A.; Oltz, E.M.; Aune, T.M. Reciprocal regulation of Rag expression in thymocytes by the zinc-finger proteins, Zfp608 and Zfp609. Genes Immun. 2013, 14, 7–12. [Google Scholar] [CrossRef][Green Version]
- Voisinne, G.; Gonzalez de Peredo, A.; Roncagalli, R. CD5, an Undercover Regulator of TCR Signaling. Front. Immunol. 2018, 9, 2900. [Google Scholar] [CrossRef]
- Nolz, J.C.; Harty, J.T. IL-15 regulates memory CD8+ T cell O-glycan synthesis and affects trafficking. J. Clin. Investig. 2014, 124, 1013–1026. [Google Scholar] [CrossRef] [PubMed]
- Conley, J.M.; Gallagher, M.P.; Berg, L.J. T Cells and Gene Regulation: The Switching On and Turning Up of Genes after T Cell Receptor Stimulation in CD8 T Cells. Front. Immunol. 2016, 7, 76. [Google Scholar] [CrossRef] [PubMed]
- Mescher, M.F.; Curtsinger, J.M.; Agarwal, P.; Casey, K.A.; Gerner, M.; Hammerbeck, C.D.; Popescu, F.; Xiao, Z. Signals required for programming effector and memory development by CD8+ T cells. Immunol. Rev. 2006, 211, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Cox, M.A.; Harrington, L.E.; Zajac, A.J. Cytokines and the inception of CD8 T cell responses. Trends Immunol. 2011, 32, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Valbon, S.F.; Condotta, S.A.; Richer, M.J. Regulation of effector and memory CD8(+) T cell function by inflammatory cytokines. Cytokine 2016, 82, 16–23. [Google Scholar] [CrossRef]
- Agarwal, P.; Raghavan, A.; Nandiwada, S.L.; Curtsinger, J.M.; Bohjanen, P.R.; Mueller, D.L.; Mescher, M.F. Gene regulation and chromatin remodeling by IL-12 and type I IFN in programming for CD8 T cell effector function and memory. J. Immunol. 2009, 183, 1695–1704. [Google Scholar] [CrossRef]
- Raue, H.P.; Beadling, C.; Haun, J.; Slifka, M.K. Cytokine-mediated programmed proliferation of virus-specific CD8(+) memory T cells. Immunity 2013, 38, 131–139. [Google Scholar] [CrossRef]
- Ramanathan, S.; Dubois, S.; Gagnon, J.; Leblanc, C.; Mariathasan, S.; Ferbeyre, G.; Rottapel, R.; Ohashi, P.S.; Ilangumaran, S. Regulation of cytokine-driven functional differentiation of CD8 T cells by suppressor of cytokine signaling 1 controls autoimmunity and preserves their proliferative capacity toward foreign antigens. J. Immunol. 2010, 185, 357–366. [Google Scholar] [CrossRef]
- Jenkins, M.R.; Mintern, J.; La Gruta, N.L.; Kedzierska, K.; Doherty, P.C.; Turner, S.J. Cell cycle-related acquisition of cytotoxic mediators defines the progressive differentiation to effector status for virus-specific CD8+ T cells. J. Immunol. 2008, 181, 3818–3822. [Google Scholar] [CrossRef]
- Plumlee, C.R.; Sheridan, B.S.; Cicek, B.B.; Lefrancois, L. Environmental cues dictate the fate of individual CD8+ T cells responding to infection. Immunity 2013, 39, 347–356. [Google Scholar] [CrossRef]
- Halle, S.; Keyser, K.A.; Stahl, F.R.; Busche, A.; Marquardt, A.; Zheng, X.; Galla, M.; Heissmeyer, V.; Heller, K.; Boelter, J.; et al. In Vivo Killing Capacity of Cytotoxic T Cells Is Limited and Involves Dynamic Interactions and T Cell Cooperativity. Immunity 2016, 44, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Tubo, N.J.; Jenkins, M.K. TCR signal quantity and quality in CD4(+) T cell differentiation. Trends Immunol. 2014, 35, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Tubo, N.J.; Pagan, A.J.; Taylor, J.J.; Nelson, R.W.; Linehan, J.L.; Ertelt, J.M.; Huseby, E.S.; Way, S.S.; Jenkins, M.K. Single naive CD4+ T cells from a diverse repertoire produce different effector cell types during infection. Cell 2013, 153, 785–796. [Google Scholar] [CrossRef] [PubMed]
- van Panhuys, N.; Klauschen, F.; Germain, R.N. T-cell-receptor-dependent signal intensity dominantly controls CD4(+) T cell polarization In Vivo. Immunity 2014, 41, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Newell, E.W.; Sigal, N.; Bendall, S.C.; Nolan, G.P.; Davis, M.M. Cytometry by time-of-flight shows combinatorial cytokine expression and virus-specific cell niches within a continuum of CD8+ T cell phenotypes. Immunity 2012, 36, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Wilson, E.B.; Brooks, D.G. Inflammation makes T cells sensitive. Immunity 2013, 38, 5–7. [Google Scholar] [CrossRef] [PubMed]
- Keppler, S.J.; Theil, K.; Vucikuja, S.; Aichele, P. Effector T-cell differentiation during viral and bacterial infections: Role of direct IL-12 signals for cell fate decision of CD8(+) T cells. Eur. J. Immunol. 2009, 39, 1774–1783. [Google Scholar] [CrossRef]
- Krupica, T., Jr.; Fry, T.J.; Mackall, C.L. Autoimmunity during lymphopenia: A two-hit model. Clin. Immunol. 2006, 120, 121–128. [Google Scholar] [CrossRef]
- Fujinami, R.S.; von Herrath, M.G.; Christen, U.; Whitton, J.L. Molecular mimicry, bystander activation, or viral persistence: Infections and autoimmune disease. Clin. Microbiol. Rev. 2006, 19, 80–94. [Google Scholar] [CrossRef]
- Ellestad, K.K.; Anderson, C.C. Two Strikes and You’re Out? The Pathogenic Interplay of Coinhibitor Deficiency and Lymphopenia-Induced Proliferation. J. Immunol. 2017, 198, 2534–2541. [Google Scholar] [CrossRef]
- Horwitz, M.S.; Yanagi, Y.; Oldstone, M.B. T-cell receptors from virus-specific cytotoxic T lymphocytes recognizing a single immunodominant nine-amino-acid viral epitope show marked diversity. J. Virol. 1994, 68, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Zehn, D.; Lee, S.Y.; Bevan, M.J. Complete but curtailed T-cell response to very low-affinity antigen. Nature 2009, 458, 211–214. [Google Scholar] [CrossRef] [PubMed]
- van Gisbergen, K.P.; Klarenbeek, P.L.; Kragten, N.A.; Unger, P.P.; Nieuwenhuis, M.B.; Wensveen, F.M.; ten Brinke, A.; Tak, P.P.; Eldering, E.; Nolte, M.A.; et al. The costimulatory molecule CD27 maintains clonally diverse CD8(+) T cell responses of low antigen affinity to protect against viral variants. Immunity 2011, 35, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Martinez, R.J.; Evavold, B.D. Lower Affinity T Cells are Critical Components and Active Participants of the Immune Response. Front. Immunol. 2015, 6, 468. [Google Scholar] [CrossRef]
- Huang, J.; Zeng, X.; Sigal, N.; Lund, P.J.; Su, L.F.; Huang, H.; Chien, Y.H.; Davis, M.M. Detection, phenotyping, and quantification of antigen-specific T cells using a peptide-MHC dodecamer. Proc. Natl. Acad. Sci. USA 2016, 113, E1890–E1897. [Google Scholar] [CrossRef]
- Ozga, A.J.; Moalli, F.; Abe, J.; Swoger, J.; Sharpe, J.; Zehn, D.; Kreutzfeldt, M.; Merkler, D.; Ripoll, J.; Stein, J.V. pMHC affinity controls duration of CD8+ T cell-DC interactions and imprints timing of effector differentiation versus expansion. J. Exp. Med. 2016, 213, 2811–2829. [Google Scholar] [CrossRef]
- Schober, K.; Voit, F.; Grassmann, S.; Muller, T.R.; Eggert, J.; Jarosch, S.; Weissbrich, B.; Hoffmann, P.; Borkner, L.; Nio, E.; et al. Reverse TCR repertoire evolution toward dominant low-affinity clones during chronic CMV infection. Nat. Immunol. 2020, 21, 434–441. [Google Scholar] [CrossRef]
- Hebeisen, M.; Allard, M.; Gannon, P.O.; Schmidt, J.; Speiser, D.E.; Rufer, N. Identifying Individual T Cell Receptors of Optimal Avidity for Tumor Antigens. Front. Immunol. 2015, 6, 582. [Google Scholar] [CrossRef]
- Miller, A.M.; Bahmanof, M.; Zehn, D.; Cohen, E.E.W.; Schoenberger, S.P. Leveraging TCR Affinity in Adoptive Immunotherapy against Shared Tumor/Self-Antigens. Cancer Immunol. Res. 2019, 7, 40–49. [Google Scholar] [CrossRef]
- Segal, G.; Prato, S.; Zehn, D.; Mintern, J.D.; Villadangos, J.A. Target Density, Not Affinity or Avidity of Antigen Recognition, Determines Adoptive T Cell Therapy Outcomes in a Mouse Lymphoma Model. J. Immunol. 2016, 196, 3935–3942. [Google Scholar] [CrossRef]
- Rodriguez, G.M.; Bobbala, D.; Serrano, D.; Mayhue, M.; Champagne, A.; Saucier, C.; Steimle, V.; Kufer, T.A.; Menendez, A.; Ramanathan, S.; et al. NLRC5 elicits antitumor immunity by enhancing processing and presentation of tumor antigens to CD8(+) T lymphocytes. Oncoimmunology 2016, 5, e1151593. [Google Scholar] [CrossRef]
- He, Q.; Jiang, X.; Zhou, X.; Weng, J. Targeting cancers through TCR-peptide/MHC interactions. J. Hematol. Oncol. 2019, 12, 139. [Google Scholar] [CrossRef] [PubMed]
- Scott-Browne, J.P.; Lopez-Moyado, I.F.; Trifari, S.; Wong, V.; Chavez, L.; Rao, A.; Pereira, R.M. Dynamic Changes in Chromatin Accessibility Occur in CD8(+) T Cells Responding to Viral Infection. Immunity 2016, 45, 1327–1340. [Google Scholar] [CrossRef] [PubMed]
- Scharer, C.D.; Bally, A.P.; Gandham, B.; Boss, J.M. Cutting Edge: Chromatin Accessibility Programs CD8 T Cell Memory. J. Immunol. 2017, 198, 2238–2243. [Google Scholar] [CrossRef] [PubMed]
- Mognol, G.P.; Spreafico, R.; Wong, V.; Scott-Browne, J.P.; Togher, S.; Hoffmann, A.; Hogan, P.G.; Rao, A.; Trifari, S. Exhaustion-associated regulatory regions in CD8(+) tumor-infiltrating T cells. Proc. Natl. Acad. Sci. USA 2017, 114, E2776–E2785. [Google Scholar] [CrossRef]
- Huber, M.; Lohoff, M. IRF4 at the crossroads of effector T-cell fate decision. Eur. J. Immunol. 2014, 44, 1886–1895. [Google Scholar] [CrossRef]
- Huang, S.; Shen, Y.; Pham, D.; Jiang, L.; Wang, Z.; Kaplan, M.H.; Zhang, G.; Sun, J. IRF4 Modulates CD8(+) T Cell Sensitivity to IL-2 Family Cytokines. Immunohorizons 2017, 1, 92–100. [Google Scholar] [CrossRef]
- Best, J.A.; Blair, D.A.; Knell, J.; Yang, E.; Mayya, V.; Doedens, A.; Dustin, M.L.; Goldrath, A.W.; Immunological Genome Project, C. Transcriptional insights into the CD8(+) T cell response to infection and memory T cell formation. Nat. Immunol. 2013, 14, 404–412. [Google Scholar] [CrossRef]
- Spolski, R.; Gromer, D.; Leonard, W.J. The gamma c family of cytokines: Fine-tuning signals from IL-2 and IL-21 in the regulation of the immune response. F1000Research 2017, 6, 1872. [Google Scholar] [CrossRef]
- Shan, Q.; Zeng, Z.; Xing, S.; Li, F.; Hartwig, S.M.; Gullicksrud, J.A.; Kurup, S.P.; Van Braeckel-Budimir, N.; Su, Y.; Martin, M.D.; et al. The transcription factor Runx3 guards cytotoxic CD8(+) effector T cells against deviation towards follicular helper T cell lineage. Nat. Immunol. 2017, 18, 931–939. [Google Scholar] [CrossRef]
- Woolf, E.; Xiao, C.; Fainaru, O.; Lotem, J.; Rosen, D.; Negreanu, V.; Bernstein, Y.; Goldenberg, D.; Brenner, O.; Berke, G.; et al. Runx3 and Runx1 are required for CD8 T cell development during thymopoiesis. Proc. Natl. Acad. Sci. USA 2003, 100, 7731–7736. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Guilloty, F.; Pipkin, M.E.; Djuretic, I.M.; Levanon, D.; Lotem, J.; Lichtenheld, M.G.; Groner, Y.; Rao, A. Runx3 and T-box proteins cooperate to establish the transcriptional program of effector CTLs. J. Exp. Med. 2009, 206, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Diao, H.; Getzler, A.J.; Rogal, W.; Frederick, M.A.; Milner, J.; Yu, B.; Crotty, S.; Goldrath, A.W.; Pipkin, M.E. The Transcription Factor Runx3 Establishes Chromatin Accessibility of cis-Regulatory Landscapes that Drive Memory Cytotoxic T Lymphocyte Formation. Immunity 2018, 48, 659–674 e656. [Google Scholar] [CrossRef] [PubMed]
- Hai, T.; Hartman, M.G. The molecular biology and nomenclature of the activating transcription factor/cAMP responsive element binding family of transcription factors: Activating transcription factor proteins and homeostasis. Gene 2001, 273, 1–11. [Google Scholar] [CrossRef]
- Kurachi, M.; Barnitz, R.A.; Yosef, N.; Odorizzi, P.M.; DiIorio, M.A.; Lemieux, M.E.; Yates, K.; Godec, J.; Klatt, M.G.; Regev, A.; et al. The transcription factor BATF operates as an essential differentiation checkpoint in early effector CD8+ T cells. Nat. Immunol. 2014, 15, 373–383. [Google Scholar] [CrossRef]
- Starks, R.R.; Biswas, A.; Jain, A.; Tuteja, G. Combined analysis of dissimilar promoter accessibility and gene expression profiles identifies tissue-specific genes and actively repressed networks. Epigenetics Chromatin 2019, 12, 16. [Google Scholar] [CrossRef]
- Kiani, K.; Sanford, E.M.; Goyal, Y.; Raj, A. Changes in chromatin accessibility are not concordant with transcriptional changes for single-factor perturbations. bioRxiv 2022. [Google Scholar] [CrossRef]
- Grandi, F.C.; Modi, H.; Kampman, L.; Corces, M.R. Chromatin accessibility profiling by ATAC-seq. Nat. Protoc. 2022, 17, 1518–1552. [Google Scholar] [CrossRef]
- Shan, Q.; Zhu, S.; Chen, X.; Liu, J.; Yuan, S.; Li, X.; Peng, W.; Xue, H.H. Tcf1-CTCF cooperativity shapes genomic architecture to promote CD8(+) T cell homeostasis. Nat. Immunol. 2022, 23, 1222–1235. [Google Scholar] [CrossRef]
- Overwijk, W.W.; Restifo, N.P. B16 as a mouse model for human melanoma. Curr. Protoc. Immunol. 2001, 39. [Google Scholar] [CrossRef]
- Overwijk, W.W.; Tsung, A.; Irvine, K.R.; Parkhurst, M.R.; Goletz, T.J.; Tsung, K.; Carroll, M.W.; Liu, C.; Moss, B.; Rosenberg, S.A.; et al. gp100/pmel 17 is a murine tumor rejection antigen: Induction of “self”-reactive, tumoricidal T cells using high-affinity, altered peptide ligand. J. Exp. Med. 1998, 188, 277–286. [Google Scholar] [PubMed]
- Buenrostro, J.D.; Giresi, P.G.; Zaba, L.C.; Chang, H.Y.; Greenleaf, W.J. Transposition of native chromatin for fast and sensitive epigenomic profiling of open chromatin, DNA-binding proteins and nucleosome position. Nat. Methods 2013, 10, 1213–1218. [Google Scholar] [CrossRef] [PubMed]
- Buenrostro, J.D.; Wu, B.; Chang, H.Y.; Greenleaf, W.J. ATAC-seq: A Method for Assaying Chromatin Accessibility Genome-Wide. Curr. Protoc. Mol. Biol. 2015, 109, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Yu, N.K.; Kaang, B.K. CTCF as a multifunctional protein in genome regulation and gene expression. Exp. Mol. Med. 2015, 47, e166. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Neph, S.; Kuehn, M.S.; Reynolds, A.P.; Haugen, E.; Thurman, R.E.; Johnson, A.K.; Rynes, E.; Maurano, M.T.; Vierstra, J.; Thomas, S.; et al. BEDOPS: High-performance genomic feature operations. Bioinformatics 2012, 28, 1919–1920. [Google Scholar] [CrossRef]
- Quinlan, A.R. BEDTools: The Swiss-Army Tool for Genome Feature Analysis. Curr. Protoc. Bioinform. 2014, 47, 11–12. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Heinz, S.; Benner, C.; Spann, N.; Bertolino, E.; Lin, Y.C.; Laslo, P.; Cheng, J.X.; Murre, C.; Singh, H.; Glass, C.K. Simple combinations of lineage-determining transcription factors prime cis-regulatory elements required for macrophage and B cell identities. Mol. Cell 2010, 38, 576–589. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Term ID | Term Description | Observed Gene Count | Background Gene Count | FDR 1 |
|---|---|---|---|---|
| (A) in cytokine-primed cells | ||||
| GO:0002376 | immune system process | 63 | 1703 | 2.19 × 10−7 |
| GO:1902105 | regulation of leukocyte differentiation | 22 | 284 | 7.52 × 10−7 |
| GO:0006955 | immune response | 35 | 914 | 0.00015 |
| GO:0045582 | positive regulation of T cell differentiation | 9 | 90 | 0.00085 |
| GO:0046634 | regulation of alpha-beta T cell activation | 9 | 98 | 0.0014 |
| GO:0030098 | lymphocyte differentiation | 14 | 243 | 0.0016 |
| GO:0045619 | regulation of lymphocyte differentiation | 11 | 171 | 0.0033 |
| GO:0030217 | T cell differentiation | 10 | 154 | 0.0051 |
| GO:2000516 | positive regulation of CD4-positive, alpha-beta T cell activation | 5 | 34 | 0.0059 |
| GO:0046649 | lymphocyte activation | 16 | 378 | 0.009 |
| GO:0046632 | alpha-beta T cell differentiation | 6 | 60 | 0.0091 |
| GO:0042110 | T cell activation | 12 | 244 | 0.0118 |
| GO:0050670 | regulation of lymphocyte proliferation | 11 | 215 | 0.014 |
| GO:0070229 | negative regulation of lymphocyte apoptotic process | 4 | 39 | 0.0468 |
| (B) in both cytokine-primed and Ag-stimulated cells | ||||
| GO:0051251 | positive regulation of lymphocyte activation | 18 | 253 | 2.70 × 10−5 |
| GO:0050870 | positive regulation of T cell activation | 16 | 180 | 1.00 × 10−5 |
| GO:0051249 | regulation of lymphocyte activation | 23 | 396 | 2.13 × 10−5 |
| GO:0050863 | regulation of T cell activation | 20 | 287 | 1.05 × 10−5 |
| GO:0045621 | positive regulation of lymphocyte differentiation | 10 | 103 | 0.00045 |
| GO:0050852 | T cell receptor signaling pathway | 6 | 56 | 0.0069 |
| GO:0046635 | positive regulation of alpha-beta T cell activation | 6 | 64 | 0.0118 |
| GO:0050856 | regulation of T cell receptor signaling pathway | 5 | 39 | 0.0093 |
| GO:0042129 | regulation of T cell proliferation | 9 | 162 | 0.0207 |
| Functional Category of Genes | ATACseq Peaks Modulated in: | ||||
|---|---|---|---|---|---|
| CytP Cells | CytP & AgS Cells | Other Names | Functions in CD8+ or Other T Lymphocytes | Ref. | |
| Immune synapse | |||||
| Rftn11 | Rftn11 | Increases TCR signaling, needed for localisation of LCK to lipid rafts | [31] | ||
| Early TCR signaling | |||||
| Cd2472 | Cd2472 | CD3z | Signaling component of the TCR:CD3 complex | [32] | |
| Dpp41 | Dpp41 | Dipeptidyl peptidase IV, CD26 | Promotes T cell activation | [33] | |
| Itk2 | Itk2 | Tec family kinase | Required for efficient TCR signaling and T cell proliferation; Integrates TCR & CytR signaling | [34] | |
| Prkcq1 | Prkcq1 | PKC theta | Critical signal strength regulator for the activation of NF-kB, NF-AT, AP-1 transcription factors | [35] | |
| Signal transduction to the nucleus | |||||
| Bcl102 | Bcl102 | B-cell lymphoma/ leukemia 10 | Essential for TCR-induced NF-kB activation | [36] | |
| Chd71 | Chd71 | Chromatin remodeler | T cell development in Zebrafish | [37] | |
| Elf12 | Elf12 | Regulates transcription of TCR ζ chain, LAT | [32,38] | ||
| Jun1 | AP-1 family transcription factor, activated by TCR signaling | [39] | |||
| Cytoskeletal remodeling & cell migration | |||||
| Ccr11 | Promotes migration of activated T cells | [40,41] | |||
| Ccr71 | Ccr71 | Controls CD8 T cell homeostasis by directing memory cells to IL-7-dependent niches | [42] | ||
| Co-stimulation | |||||
| Cd282 | Cd282 | T cell co-stimulatory receptor | [43] | ||
| Functional differentiation | |||||
| Dusp102 | Dusp102 | Mkp5 | Facilitates proliferation & cytokine production | [44] | |
| Foxp12 | Foxp12 | Maintains quiescence in naive T cells | [45] | ||
| Gadd45g1 | Promotes effector functions in CD8 T cells | [46] | |||
| Gpr182 | Gpr182 | Controls effector-memory CD8 T cells | [47] | ||
| Il15ra | Il15ra2 | Ligand-specificity subunit of IL-15R; controls memory CD8 T cell homeostasis | [48] | ||
| Il6st2 | Il6st2 | gp130, IL-6 signal transducer | Needed for efficient generation of memory CD8 T cells | [49] | |
| H2-Aa2 | MHC-II antigen | Over-expressed in PD-1 KO CD8 T-CM phenotype cells | [50] | ||
| Lef12 | Lef12 | Lymphoid enhancer binding factor 1; Possess HDAC activity | suppresses CD4 lineage genes in CD8 T cells; highly expressed in quiescent cells; downregulated by TCR and IL-15 signaling | [51] | |
| Nfkbiz1 | Nfkbiz1 | IkBz | promotes Th17 differentiation | [52] | |
| Prkca2 | Prkca2 | PKC alpha | Promotes Th17 resposne by upregulating IL-17A expression | [53] | |
| Rara2 | Rara2 | Essential for CD4 T cell activation and CD8 T cell survival | [54] | ||
| Stat5b2 | Stat5b2 | Critical to maintain effector T cell responses | [55] | ||
| Tgfbr22 | Tgfbr22 | Regulates IL-15-mediated CD8 T cell homeostasis | [56] | ||
| Tnfsf81 | Tnfsf81 | TNF Superfamily Member 8; CD153; CD30L | Induces prolifertaion of T cells | [57] | |
| Negative regulation | |||||
| Btla1 | B and T-lymphocyte attenuator; CD272 | Negative regulation of T cell activation | [58] | ||
| Cblb2 | Cblb2 | Casitas B Lymphoma Proto-Oncogene B | Negative regulation of TCR signaling pathways | [59] | |
| Cd1012 | Negative regulation of T cell activation | [60] | |||
| Cd300a1 | Negative regulator of T cell activation | [61] | |||
| Cd472 | Cd472 | Promotes/inhibits T cell activation | [62] | ||
| Tigit2 | CD115T | Suppresses CD8 T cell activation | [63] | ||
| Associated with Exhaustion | |||||
| Trib12 | Tribbles Pseudokinase 1 | Expressed in hypofunctional CD8 T cells in tumors and during chronic infection | [64] | ||
| Xbp12 | X-box-binding protein-1 | Effector T cell differentiation; Induces CD8 T cell exhaustion accompanied by increased expression of inhibitory receptors | [65,66] | ||
| Unknown functions in mature T cells | |||||
| Tmem642 | Tmem642 | Positively regulates TNFSF11-mediated NF-AT-dependent Ca2+ signaling in osteoclasts; not yet implicated in Lymphocyte activation | [67] | ||
| Tnfsf112 | TNF Superfamily Member 11; RANKL; CD254 | Suppresses effector cytokine production in group 3 innate lymphoid cells | [68] | ||
| Zfp6091 | Zfp6091 | Represses Rag gene expression during T cell development | [69] | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quenum, A.J.I.; Santharam, M.A.; Ramanathan, S.; Ilangumaran, S. Inflammatory Cytokines That Enhance Antigen Responsiveness of Naïve CD8+ T Lymphocytes Modulate Chromatin Accessibility of Genes Impacted by Antigen Stimulation. Int. J. Mol. Sci. 2022, 23, 14122. https://doi.org/10.3390/ijms232214122
Quenum AJI, Santharam MA, Ramanathan S, Ilangumaran S. Inflammatory Cytokines That Enhance Antigen Responsiveness of Naïve CD8+ T Lymphocytes Modulate Chromatin Accessibility of Genes Impacted by Antigen Stimulation. International Journal of Molecular Sciences. 2022; 23(22):14122. https://doi.org/10.3390/ijms232214122
Chicago/Turabian StyleQuenum, Akouavi Julite Irmine, Madanraj Appiya Santharam, Sheela Ramanathan, and Subburaj Ilangumaran. 2022. "Inflammatory Cytokines That Enhance Antigen Responsiveness of Naïve CD8+ T Lymphocytes Modulate Chromatin Accessibility of Genes Impacted by Antigen Stimulation" International Journal of Molecular Sciences 23, no. 22: 14122. https://doi.org/10.3390/ijms232214122
APA StyleQuenum, A. J. I., Santharam, M. A., Ramanathan, S., & Ilangumaran, S. (2022). Inflammatory Cytokines That Enhance Antigen Responsiveness of Naïve CD8+ T Lymphocytes Modulate Chromatin Accessibility of Genes Impacted by Antigen Stimulation. International Journal of Molecular Sciences, 23(22), 14122. https://doi.org/10.3390/ijms232214122