



Enhanced Aggression, Reduced Self-Grooming Behavior and Altered 5-HT Regulation in the Frontal Cortex in Mice Lacking Trace Amine-Associated Receptor 1 (TAAR1)

 , and
, and
Abstract
1. Introduction
2. Results
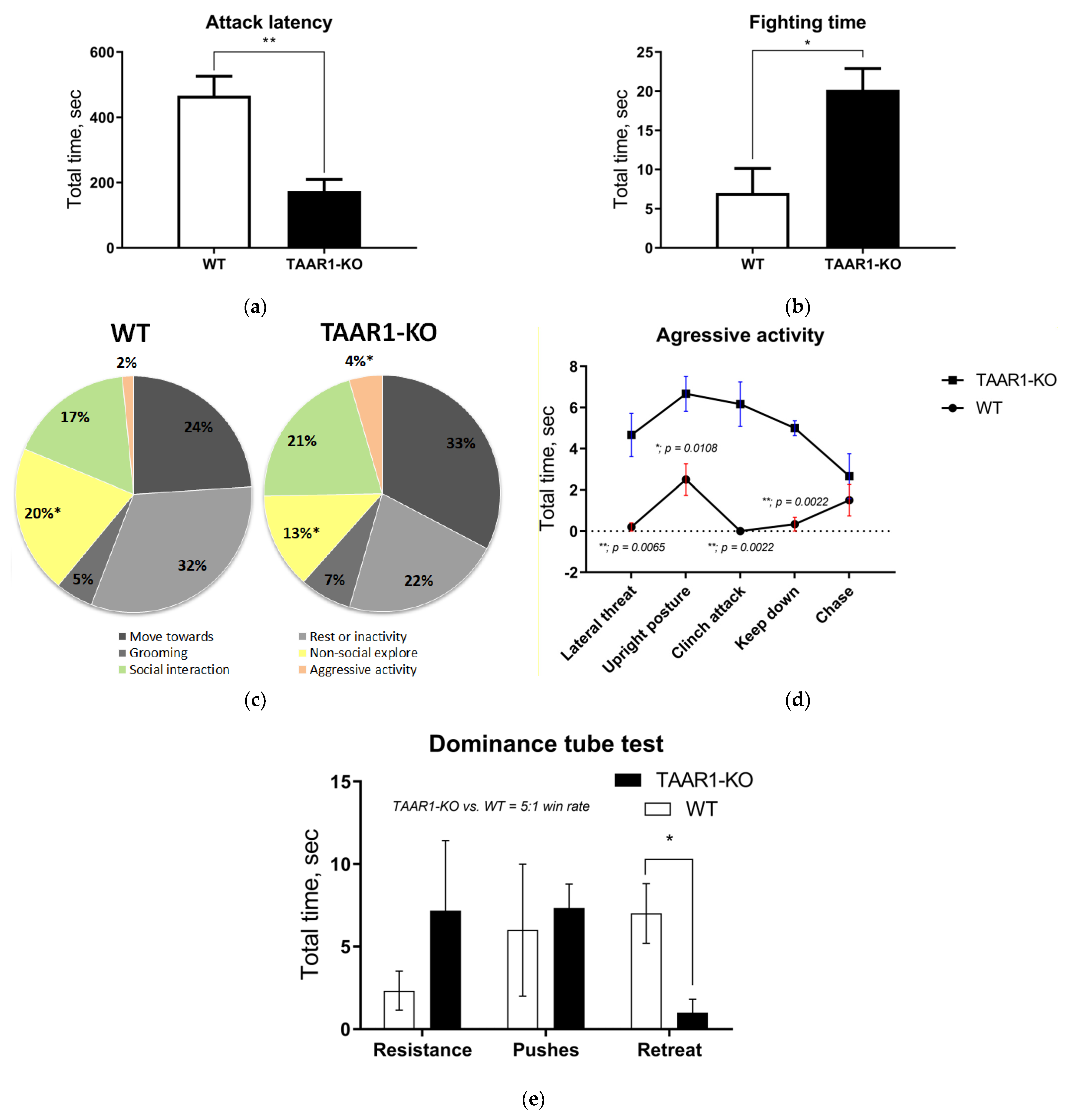
2.1. Behavioral Phenotypes
2.2. Neurochemical Analyses
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Behavioral Assays
4.3. HPLC Measurements of the Monoamines Tissue Content
4.4. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gainetdinov, R.R.; Hoener, M.C.; Berry, M.D. Trace Amines and Their Receptors. Pharmacol. Rev. 2018, 70, 549–620. [Google Scholar] [CrossRef] [PubMed]
- Boulton, A.A. Letter: Amines and theories in psychiatry. Lancet (Lond. Engl.) 1974, 2, 52–53. [Google Scholar] [CrossRef]
- Borowsky, B.; Adham, N.; Jones, K.A.; Raddatz, R.; Artymyshyn, R.; Ogozalek, K.L.; Durkin, M.M.; Lakhlani, P.P.; Bonini, J.A.; Pathirana, S.; et al. Trace amines: Identification of a family of mammalian G protein-coupled receptors. Proc. Natl. Acad. Sci. USA 2001, 98, 8966–8971. [Google Scholar] [CrossRef]
- Bunzow, J.R.; Sonders, M.S.; Arttamangkul, S.; Harrison, L.M.; Zhang, G.; Quigley, D.I.; Darland, T.; Suchland, K.L.; Pasumamula, S.; Kennedy, J.L.; et al. Amphetamine, 3,4-methylenedioxymethamphetamine, lysergic acid diethylamide, and metabolites of the catecholamine neurotransmitters are agonists of a rat trace amine receptor. Mol. Pharmacol. 2001, 60, 1181–1188. [Google Scholar] [CrossRef]
- Premont, R.T.; Gainetdinov, R.R.; Caron, M.G. Following the trace of elusive amines. Proc. Natl. Acad. Sci. USA 2001, 98, 9474. [Google Scholar] [CrossRef]
- Berry, M.D.; Gainetdinov, R.R.; Hoener, M.C.; Shahid, M. Pharmacology of human trace amine-associated receptors: Therapeutic opportunities and challenges. Pharmacol. Ther. 2017, 180, 161–180. [Google Scholar] [CrossRef]
- Grandy, D.K.; Miller, G.M.; Li, J.X. “TAARgeting Addiction” The Alamo Bears Witness to Another Revolution: An Overview of the Plenary Symposium of the 2015 Behavior, Biology and Chemistry Conference. Drug Alcohol Depend. 2016, 159, 9. [Google Scholar] [CrossRef]
- Schwartz, M.D.; Black, S.W.; Fisher, S.P.; Palmerston, J.B.; Morairty, S.R.; Hoener, M.C.; Kilduff, T.S. Trace Amine-Associated Receptor 1 Regulates Wakefulness and EEG Spectral Composition. Neuropsychopharmacology 2017, 42, 1305–1314. [Google Scholar] [CrossRef]
- Leo, D.; Targa, G.; Espinoza, S.; Villers, A.; Gainetdinov, R.R.; Ris, L. Trace Amine Associate Receptor 1 (TAAR1) as a New Target for the Treatment of Cognitive Dysfunction in Alzheimer’s Disease. Int. J. Mol. Sci. 2022, 23, 7811. [Google Scholar] [CrossRef]
- Lam, V.M.; Mielnik, C.A.; Baimel, C.; Beerepoot, P.; Espinoza, S.; Sukhanov, I.; Horsfall, W.; Gainetdinov, R.R.; Borgland, S.L.; Ramsey, A.J.; et al. Behavioral effects of a potential novel TAAR1 antagonist. Front. Pharmacol. 2018, 9, 953. [Google Scholar] [CrossRef]
- Kantrowitz, J.T. Trace Amine-Associated Receptor 1 as a Target for the Development of New Antipsychotics: Current Status of Research and Future Directions. CNS Drugs 2021, 35, 1153–1161. [Google Scholar] [CrossRef] [PubMed]
- Cho, W.K.; Shin, W.S.; An, I.; Bang, M.; Cho, D.Y.; Lee, S.H. Biological aspects of aggression and violence in Schizophrenia. Clin. Psychopharmacol. Neurosci. 2019, 17, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Volavka, J. Violence in schizophrenia and bipolar disorder. Psychiatr. Danub. 2013, 25, 24–33. [Google Scholar] [PubMed]
- Fritz, M.; Shenar, R.; Cardenas-Morales, L.; Jäger, M.; Streb, J.; Dudeck, M.; Franke, I. Aggressive and Disruptive Behavior Among Psychiatric Patients With Major Depressive Disorder, Schizophrenia, or Alcohol Dependency and the Effect of Depression and Self-Esteem on Aggression. Front. Psychiatry 2020, 11, 1381. [Google Scholar] [CrossRef]
- Kunik, M.E.; Snow, A.L.; Davila, J.A.; Steele, A.B.; Balasubramanyam, V.; Doody, R.S.; Schulz, P.E.; Kalavar, J.S.; Morgan, R.O. Causes of aggressive behavior in patients with dementia. J. Clin. Psychiatry 2010, 71, 1145–1152. [Google Scholar] [CrossRef]
- Couppis, M.H.; Kennedy, C.H. The rewarding effect of aggression is reduced by nucleus accumbens dopamine receptor antagonism in mice. Psychopharmacology 2008, 197, 449–456. [Google Scholar] [CrossRef]
- Suzuki, H.; Lucas, L.R. Neurochemical Correlates of Accumbal Dopamine D2 and Amygdaloid 5-HT1B Receptor Densities on Observational Learning of Aggression. Cogn. Affect. Behav. Neurosci. 2015, 15, 460. [Google Scholar] [CrossRef]
- Seo, D.; Patrick, C.J.; Kennealy, P.J. Role of Serotonin and Dopamine System Interactions in the Neurobiology of Impulsive Aggression and its Comorbidity with other Clinical Disorders. Aggress. Violent Behav. 2008, 13, 383. [Google Scholar] [CrossRef]
- Gutknecht, L.; Popp, S.; Waider, J.; Sommerlandt, F.M.J.; Göppner, C.; Post, A.; Reif, A.; Van Den Hove, D.; Strekalova, T.; Schmitt, A.; et al. Interaction of brain 5-HT synthesis deficiency, chronic stress and sex differentially impact emotional behavior in Tph2 knockout mice. Psychopharmacology 2015, 232, 2429. [Google Scholar] [CrossRef]
- Gorlova, A.; Ortega, G.; Waider, J.; Bazhenova, N.; Veniaminova, E.; Proshin, A.; Kalueff, A.V.; Anthony, D.C.; Lesch, K.P.; Strekalova, T. Stress-induced aggression in heterozygous TPH2 mutant mice is associated with alterations in serotonin turnover and expression of 5-HT6 and AMPA subunit 2A receptors. J. Affect. Disord. 2020, 272, 440–451. [Google Scholar] [CrossRef]
- Dedic, N.; Dworak, H.; Zeni, C.; Rutigliano, G.; Howes, O.D. Therapeutic Potential of TAAR1 Agonists in Schizophrenia: Evidence from Preclinical Models and Clinical Studies. Int. J. Mol. Sci. 2021, 22, 13185. [Google Scholar] [CrossRef]
- Zhang, X.; Guan, W.; Yang, T.; Furlan, A.; Xiao, X.; Yu, K.; An, X.; Galbavy, W.; Ramakrishnan, C.; Deisseroth, K.; et al. Genetically identified amygdala–striatal circuits for valence-specific behaviors. Nat. Neurosci. 2021, 24, 1586–1600. [Google Scholar] [CrossRef] [PubMed]
- Kalueff, A.V.; Stewart, A.M.; Song, C.; Berridge, K.C.; Graybiel, A.M.; Fentress, J.C. Neurobiology of rodent self-grooming and its value for translational neuroscience. Nat. Rev. Neurosci. 2016, 17, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, S.; Ghisi, V.; Emanuele, M.; Leo, D.; Sukhanov, I.; Sotnikova, T.D.; Chieregatti, E.; Gainetdinov, R.R. Postsynaptic D2 dopamine receptor supersensitivity in the striatum of mice lacking TAAR1. Neuropharmacology 2015, 93, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Harmeier, A.; Obermueller, S.; Meyer, C.A.; Revel, F.G.; Buchy, D.; Chaboz, S.; Dernick, G.; Wettstein, J.G.; Iglesias, A.; Rolink, A.; et al. Trace amine-associated receptor 1 activation silences GSK3β signaling of TAAR1 and D2R heteromers. Eur. Neuropsychopharmacol. 2015, 25, 2049–2061. [Google Scholar] [CrossRef]
- Suri, D.; Teixeira, C.M.; Cagliostro, M.K.C.; Mahadevia, D.; Ansorge, M.S. Monoamine-Sensitive Developmental Periods Impacting Adult Emotional and Cognitive Behaviors. Neuropsychopharmacology 2015, 40, 88. [Google Scholar] [CrossRef]
- Mahadevia, D.; Saha, R.; Manganaro, A.; Chuhma, N.; Ziolkowski-Blake, A.; Morgan, A.A.; Dumitriu, D.; Rayport, S.; Ansorge, M.S. Dopamine promotes aggression in mice via ventral tegmental area to lateral septum projections. Nat. Commun. 2021, 12, 6796. [Google Scholar] [CrossRef]
- Peeters, D.G.A.; de Boer, S.F.; Terneusen, A.; Newman-Tancredi, A.; Varney, M.A.; Verkes, R.J.; Homberg, J.R. Enhanced aggressive phenotype of Tph2 knockout rats is associated with diminished 5-HT 1A receptor sensitivity. Neuropharmacology 2019, 153, 134–141. [Google Scholar] [CrossRef]
- Rutigliano, G.; Accorroni, A.; Zucchi, R. The case for TAAR1 as a modulator of central nervous system function. Front. Pharmacol. 2018, 8, 987. [Google Scholar] [CrossRef]
- Takahashi, A.; Quadros, I.M.; De Almeida, R.M.M.; Miczek, K.A. Brain Serotonin Receptors and Transporters: Initiation vs. Termination of Escalated Aggression. Psychopharmacology 2011, 213, 183. [Google Scholar] [CrossRef]
- Caramaschi, D.; de Boer, S.F.; de Vries, H.; Koolhaas, J.M. Development of violence in mice through repeated victory along with changes in prefrontal cortex neurochemistry. Behav. Brain Res. 2008, 189, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Choy, O.; Raine, A.; Hamilton, R.H. Stimulation of the Prefrontal Cortex Reduces Intentions to Commit Aggression: A Randomized, Double-Blind, Placebo-Controlled, Stratified, Parallel-Group Trial. J. Neurosci. 2018, 38, 6505–6512. [Google Scholar] [CrossRef] [PubMed]
- Puig, M.V.; Gulledge, A.T. Serotonin and Prefrontal Cortex Function: Neurons, Networks, and Circuits. Mol. Neurobiol. 2011, 44, 449. [Google Scholar] [CrossRef] [PubMed]
- Leo, D.; Mus, L.; Espinoza, S.; Hoener, M.C.; Sotnikova, T.D.; Gainetdinov, R.R. Taar1-mediated modulation of presynaptic dopaminergic neurotransmission: Role of D2 dopamine autoreceptors. Neuropharmacology 2014, 81, 283–291. [Google Scholar] [CrossRef]
- Bräunig, J.; Dinter, J.; Höfig, C.S.; Paisdzior, S.; Szczepek, M.; Scheerer, P.; Rosowski, M.; Mittag, J.; Kleinau, G.; Biebermann, H. The Trace Amine-Associated Receptor 1 Agonist 3-Iodothyronamine Induces Biased Signaling at the Serotonin 1b Receptor. Front. Pharmacol. 2018, 9, 222. [Google Scholar] [CrossRef]
- Mohandass, A.; Krishnan, V.; Gribkova, E.D.; Asuthkar, S.; Baskaran, P.; Nersesyan, Y.; Hussain, Z.; Wise, L.M.; George, R.E.; Stokes, N.; et al. TRPM8 as the rapid testosterone signaling receptor: Implications in the regulation of dimorphic sexual and social behaviors. FASEB J. 2020, 34, 10887. [Google Scholar] [CrossRef]
- Zhukov, I.S.; Ptukha, M.A.; Zolotoverkhaja, E.A.; Sinitca, E.L.; Tissen, I.Y.; Karpova, I.V.; Volnova, A.B.; Gainetdinov, R.R. Evaluation of Approach to a Conspecific and Blood Biochemical Parameters in TAAR1 Knockout Mice. Brain Sci. 2022, 12, 614. [Google Scholar] [CrossRef]
- Revel, F.G.; Moreau, J.L.; Gainetdinov, R.R.; Bradaia, A.; Sotnikova, T.D.; Mory, R.; Durkin, S.; Zbinden, K.G.; Norcross, R.; Meyer, C.A.; et al. TAAR1 activation modulates monoaminergic neurotransmission, preventing hyperdopaminergic and hypoglutamatergic activity. Proc. Natl. Acad. Sci. USA 2011, 108, 8485–8490. [Google Scholar] [CrossRef]
- Zhukov, I.S.; Kubarskaya, L.G.; Tissen, I.Y.; Kozlova, A.A.; Dagayev, S.G.; Kashuro, V.A.; Vlasova, O.L.; Sinitca, E.L.; Karpova, I.V.; Gainetdinov, R.R. Minimal Age-Related Alterations in Behavioral and Hematological Parameters in Trace Amine-Associated Receptor 1 (TAAR1) Knockout Mice. Cell. Mol. Neurobiol. 2020, 40, 273–282. [Google Scholar] [CrossRef]
- Paumier, K.L.; Sukoff Rizzo, S.J.; Berger, Z.; Chen, Y.; Gonzales, C.; Kaftan, E.; Li, L.; Lotarski, S.; Monaghan, M.; Shen, W.; et al. Behavioral Characterization of A53T Mice Reveals Early and Late Stage Deficits Related to Parkinson’s Disease. PLoS ONE 2013, 8, e70274. [Google Scholar] [CrossRef]
- Rogel-Salazar, G.; López-Rubalcava, C. Evaluation of the anxiolytic-like effects of clomipramine in two rat strains with different anxiety vulnerability (Wistar and Wistar-Kyoto rats): Participation of 5-HT1A receptors. Behav. Pharmacol. 2011, 22, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Cromwell, H.C.; Berridge, K.C. Implementation of action sequences by a neostriatal site: A lesion mapping study of grooming syntax. J. Neurosci. 1996, 16, 3444–3458. [Google Scholar] [CrossRef] [PubMed]
- Koolhaas, J.M.; Coppens, C.M.; de Boer, S.F.; Buwalda, B.; Meerlo, P.; Timmermans, P.J.A. The Resident-intruder Paradigm: A Standardized Test for Aggression, Violence and Social Stress. J. Vis. Exp. 2013, 77, e4367. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Zhu, H.; Fan, Z.; Wang, F.; Chen, Y.; Liang, H.; Yang, Z.; Zhang, L.; Lin, L.; Zhan, Y.; et al. History of winning remodels thalamo-PFC circuit to reinforce social dominance. Science 2017, 357, 162–168. [Google Scholar] [CrossRef]
- Karpova, I.V.; Mikheev, V.V.; Marysheva, V.V.; Bychkov, E.R.; Proshin, S.N. Oxytocin-Induced Changes in Monoamine Level in Symmetric Brain Structures of Isolated Aggressive C57Bl/6 Mice. Bull. Exp. Biol. Med. 2016, 160, 605–609. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Endpoint | WT | TAAR1-KO | U | p Value |
|---|---|---|---|---|
| Incorrect transitions, % | 0.67 ± 0.02 | 0.6118 ± 0.01679 * | 50 | 0.0274 |
| Total grooming bouts, n | 118.3 ± 12.2 | 77.14 ± 10.22 * | 45.4 | 0.0145 |
| Rostral grooming bouts, n | 83.9 ± 9.1 | 57.64 ± 7.077 * | 51 | 0.0303 |
| Caudal grooming bouts, n | 34.4 ± 3.8 | 19.5 ± 3.781 * | 44.5 | 0.0127 |
| Interrupted grooming bouts, % | 0.40 ± 0.04 | 0.3854 ± 0.06415 | 70 | 0.2057 |
| Paws grooming bouts, n | 37.57 ± 4.715 | 25.36 ± 2.946 | 57 | 0.0603 |
| Nose grooming bouts, n | 31.14 ± 2.721 | 27.21 ± 3.551 | 75 | 0.3004 |
| Head grooming bouts, n | 15.14 ± 3.572 | 5.071 ± 1.777 * | 43 | 0.0099 |
| Body grooming bouts, n | 24.86 ± 2.957 | 14.17 ± 3.153 * | 48.5 | 0.0217 |
| Tail grooming bouts, n | 9.571 + 1.797 | 4.786 + 1.1 * | 52 | 0.0333 |
| Paws to nose transitions, n | 16.5 ± 1.504 | 16.64 ± 2.269 | 95.5 | 0.9185 |
| Nose to head transitions, n | 2.214 ± 0.5565 | 1 ± 0.4804 * | 56.5 | 0.0478 |
| Head to body transitions, n | 1.929 ± 0.5494 | 1.214 ± 0.6305 | 58.5 | 0.0595 |
| Body to tail transitions, n | 5.357 ± 1.137 | 3.357 ± 0.7956 | 70.5 | 0.211 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhukov, I.S.; Karpova, I.V.; Krotova, N.A.; Tissen, I.Y.; Demin, K.A.; Shabanov, P.D.; Budygin, E.A.; Kalueff, A.V.; Gainetdinov, R.R. Enhanced Aggression, Reduced Self-Grooming Behavior and Altered 5-HT Regulation in the Frontal Cortex in Mice Lacking Trace Amine-Associated Receptor 1 (TAAR1). Int. J. Mol. Sci. 2022, 23, 14066. https://doi.org/10.3390/ijms232214066
Zhukov IS, Karpova IV, Krotova NA, Tissen IY, Demin KA, Shabanov PD, Budygin EA, Kalueff AV, Gainetdinov RR. Enhanced Aggression, Reduced Self-Grooming Behavior and Altered 5-HT Regulation in the Frontal Cortex in Mice Lacking Trace Amine-Associated Receptor 1 (TAAR1). International Journal of Molecular Sciences. 2022; 23(22):14066. https://doi.org/10.3390/ijms232214066
Chicago/Turabian StyleZhukov, Ilya S., Inessa V. Karpova, Nataliya A. Krotova, Ilya Y. Tissen, Konstantin A. Demin, Petr D. Shabanov, Evgeny A. Budygin, Allan V. Kalueff, and Raul R. Gainetdinov. 2022. "Enhanced Aggression, Reduced Self-Grooming Behavior and Altered 5-HT Regulation in the Frontal Cortex in Mice Lacking Trace Amine-Associated Receptor 1 (TAAR1)" International Journal of Molecular Sciences 23, no. 22: 14066. https://doi.org/10.3390/ijms232214066
APA StyleZhukov, I. S., Karpova, I. V., Krotova, N. A., Tissen, I. Y., Demin, K. A., Shabanov, P. D., Budygin, E. A., Kalueff, A. V., & Gainetdinov, R. R. (2022). Enhanced Aggression, Reduced Self-Grooming Behavior and Altered 5-HT Regulation in the Frontal Cortex in Mice Lacking Trace Amine-Associated Receptor 1 (TAAR1). International Journal of Molecular Sciences, 23(22), 14066. https://doi.org/10.3390/ijms232214066