
Comparative Analysis of Whole Chloroplast Genomes of Three Common Species of Echinochloa (Gramineae) in Paddy Fields

Abstract
1. Introduction
2. Results
2.1. Differences in the Phenotype of Seeds
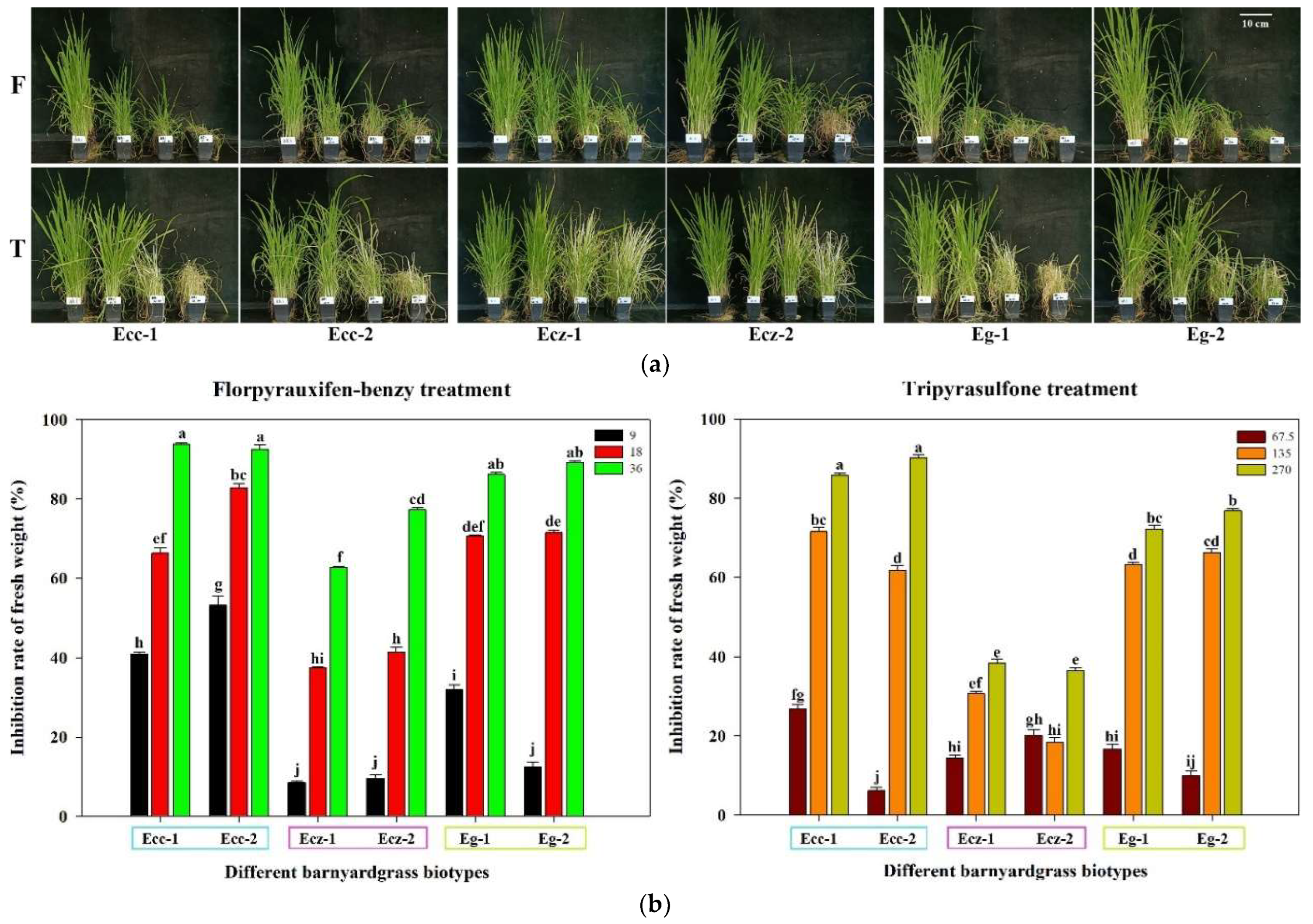
2.2. Differences in Sensitivity to New Herbicides
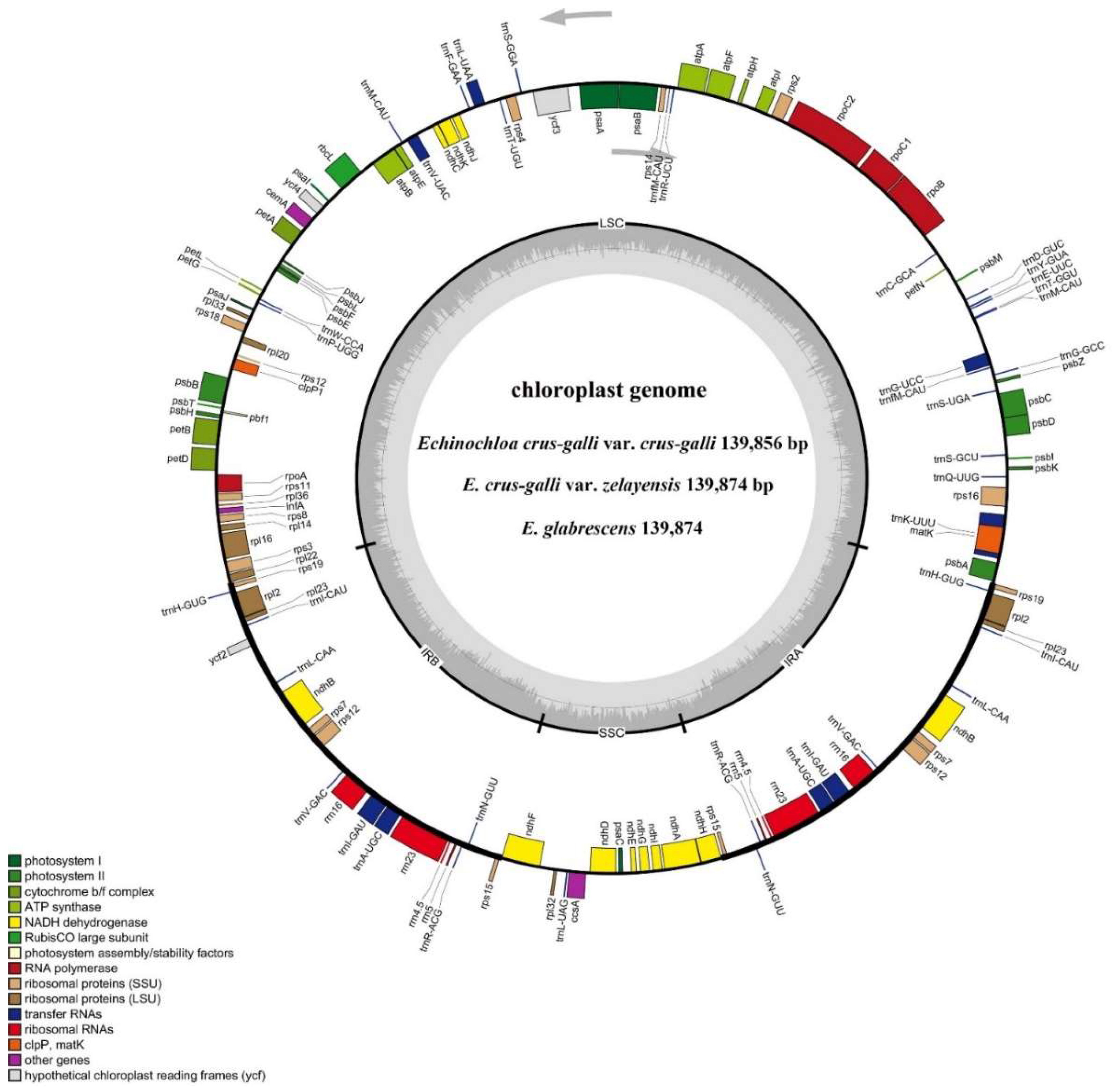
2.3. Characteristics of Chloroplast Genomes
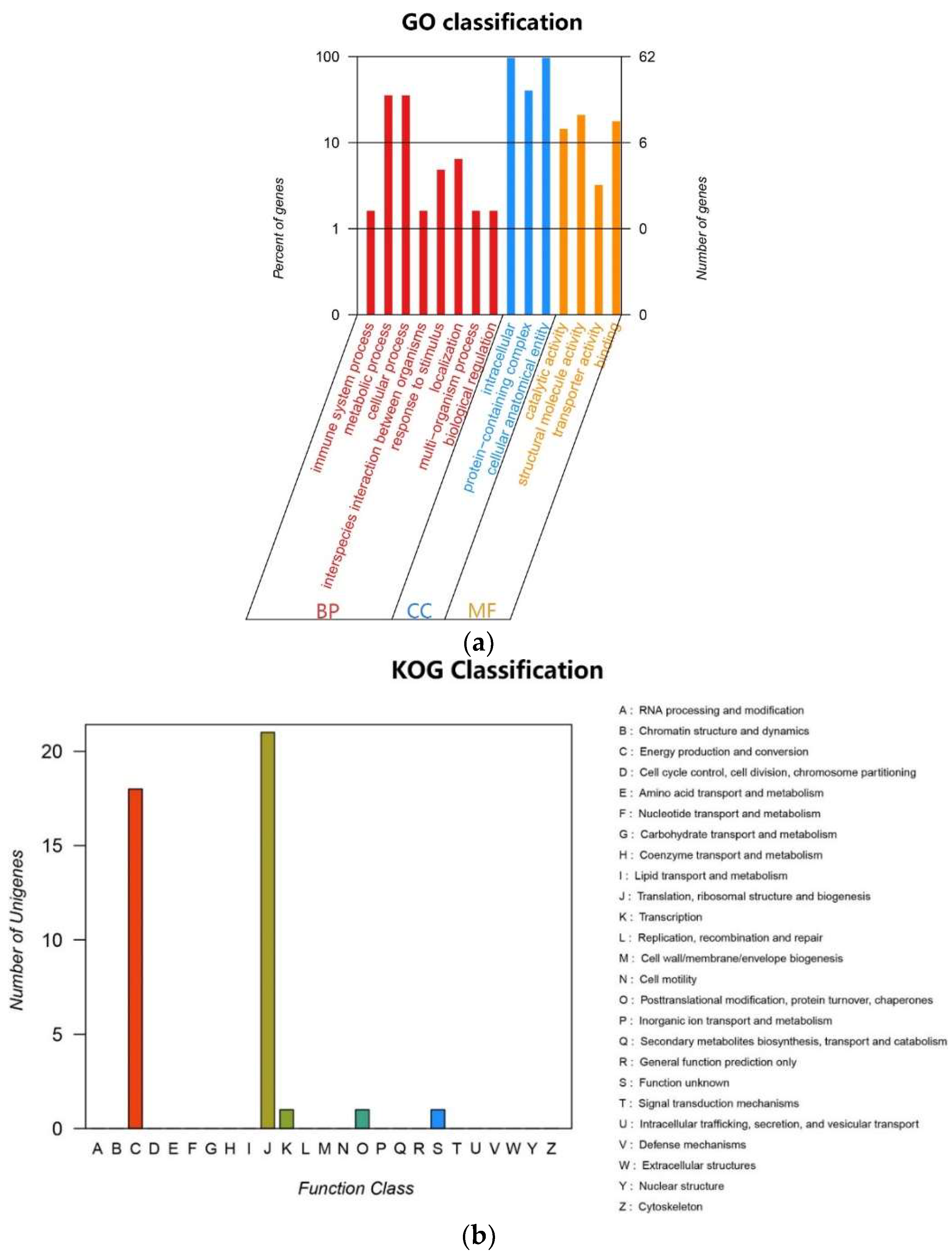
2.4. Chloroplast Genome Component
2.5. Single-Nucleotide Polymorphism Analysis
2.6. IR Expansion and Contraction
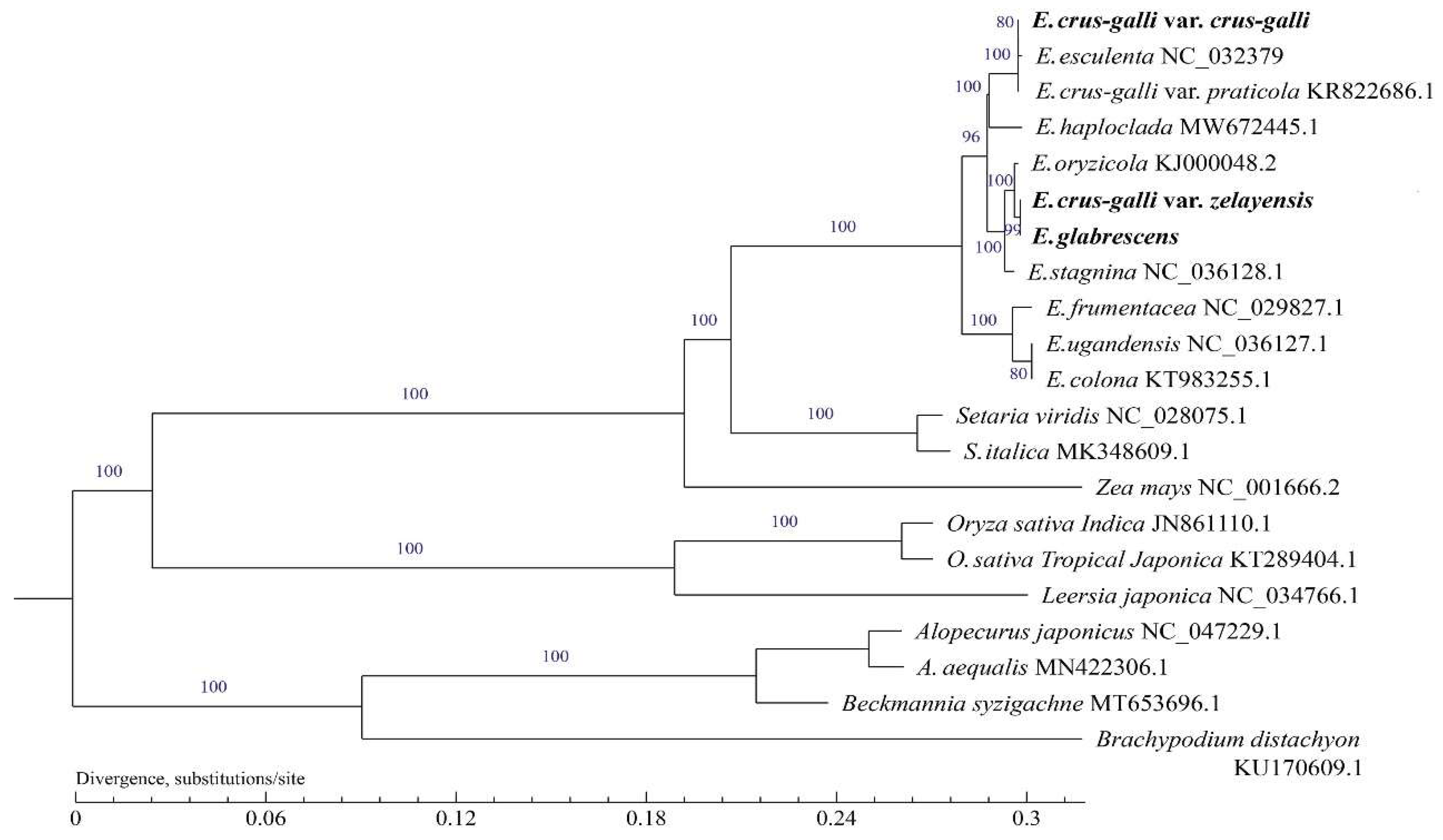
2.7. Phylogenetic Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Measurement of Awn Length and Seed Weight
4.3. Whole-Plant Bioassay to Determine Sensitivity to New Herbicides
4.4. DNA Extraction and Sequencing
4.5. DNA Sequencing and Genome Assembly
4.6. Genome Component Analysis
4.7. Gene Function Annotation and Classification Analysis
4.8. Contraction and Expansion Analysis of Inverted Repeat (IR) Regions
4.9. Phylogenetic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Michael, P. Taxonomy and distribution of Echinochloa species with special reference to their occurrence as weeds of rice. In Proceedings of the Conference on Weed Control in Rice, Los Banos, Philippines, 31 August–4 September 1981; pp. 291–306. [Google Scholar]
- Yabuno, T. Biology of Echinochloa species. In Proceedings of the Conference on Weed Control in Rice, Los Banos, Philippines, 31 August–4 September 1981; pp. 307–318. [Google Scholar]
- Xu, J.; Lv, B.; Wang, Q.; Li, J.; Dong, L. A resistance mechanism dependent upon the inhibition of ethylene biosynthesis. Pest Manag. Sci. 2013, 69, 1407–1414. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Zhang, Y.; Liu, T.; Yan, B.; Li, J.; Dong, L. Target-site and metabolic resistance mechanisms to penoxsulam in barnyardgrass (Echinochloa crus-galli (L.) P. Beauv). J. Agric. Food Chem. 2019, 67, 8085–8095. [Google Scholar] [CrossRef]
- Danquah, E.; Johnson, D.; Riches, C.; Arnold, G.; Karp, A. Genetic diversity in Echinochloa spp. collected from different geographic origins and within rice fields in Cote d’Ivoire. Weed Res. 2002, 42, 394–405. [Google Scholar] [CrossRef]
- Gibson, K.D.; Fischer, A.J.; Foin, T.C.; Hill, J.E. Implications of delayed Echinochloa spp. germination and duration of competition for integrated weed management in water-seeded rice. Weed Res. 2010, 42, 351–358. [Google Scholar] [CrossRef]
- Chauhan, B.; Johnson, D. Ecological studies on Echinochloa crus-galli and the implications for weed management in direct-seeded rice. Crop Prot. 2011, 30, 1385–1391. [Google Scholar] [CrossRef]
- Rao, A.; Johnson, D.; Sivaprasad, B.; Ladha, J.; Mortimer, A. Weed management in direct-seeded rice. Adv. Agron. 2007, 93, 153–255. [Google Scholar]
- Vidotto, F.; Tesio, F.; Tabacchi, M.; Ferrero, A. Herbicide sensitivity of Echinochloa spp. accessions in Italian rice fields. Crop Prot. 2007, 26, 285–293. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, G.; Jiang, Y.; Wang, Q.; Yao, Z.; Dong, L. Sensitivity of Echinochloa species to frequently used herbicides in paddy rice field. J. Nanjing Agric. Univ. 2015, 38, 804–809. (In Chinese) [Google Scholar]
- Qiao, L.; Wang, Q.; Zhang, S.; Li, Y. Review on the biology of weed of Echinochloa Beauv. Weed Sci. 2002, 3, 8–12. (In Chinese) [Google Scholar]
- Ruiz-Santaella, J.; Bastida, F.; Franco, A.; De Prado, R. Morphological and molecular characterization of different Echinochloa spp. and Oryza sativa populations. J. Agric. Food Chem. 2006, 54, 1166–1172. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Utano, A.; Yasuda, K.; Yano, A.; Soejima, A. A molecular phylogeny of wild and cultivated Echinochloa in East Asia inferred from non-coding region sequences of trnT-L-F. Weed Biol. Manag. 2005, 5, 210–218. [Google Scholar] [CrossRef]
- Nakayama, Y.; Umemoto, S.; Yamaguchi, H. Identification of polyploid groups in the genus Echinochloa by isozyme analysis. Weed Res. 1999, 44, 205–217. [Google Scholar]
- Yasuda, K.; Yano, A.; Nakayama, Y.; Yamaguchi, H. Molecular identification of Echinochloa oryzicola Vasing. and E. crus-galli (L.) Beauv. using a polymerase chain reaction-restriction fragment length polymorphism technique. Weed Biol. Manag. 2002, 2, 11–17. [Google Scholar] [CrossRef]
- Feng, J.; Zhang, T. Cytological study on Chinese species in the genus Echinochloa. J. Wuhan Bot. Res. 1993, 11, 293–299. (In Chinese) [Google Scholar]
- Qian, W.; Ge, S.; Hong, D.Y. Genetic variation within and among populations of a wild rice Oryza granulata from China detected by RAPD and ISSR markers. Theor. Appl. Genet. 2001, 102, 440–449. [Google Scholar] [CrossRef]
- Devarumath, R.; Nandy, S.; Rani, V.; Marimuthu, S.; Muraleedharan, N.; Raina, S. RAPD, ISSR and RFLP fingerprints as useful markers to evaluate genetic integrity of micropropagated plants of three diploid and triploid elite tea clones representing Camellia sinensis (China type) and C. assamica ssp. assamica (Assam-India type). Plant Cell Rep. 2002, 21, 166–173. [Google Scholar]
- Lu, Y.; Liu, D.; Guo, S.; Yu, L. Classification of Echinochloa species in Chinese paddy fields based on ISSR markers. Acta Agric. Zhejiangensis 2014, 26, 1309–1314. (In Chinese) [Google Scholar]
- Wu, D.; Shen, E.; Jiang, B.; Feng, Y.; Tang, W.; Lao, S.; Jia, L.; Lin, H.-Y.; Xie, L.; Weng, X. Genomic insights into the evolution of Echinochloa species as weed and orphan crop. Nat. Commun. 2022, 13, 689. [Google Scholar] [CrossRef]
- Altop, E.K.; Mennan, H. Genetic and morphologic diversity of Echinochloa crus-galli populations from different origins. Phytoparasitica 2011, 39, 93–102. [Google Scholar] [CrossRef]
- Gupta, A.; Mahajan, V.; Kumar, M.; Gupta, H. Biodiversity in the barnyard millet (Echinochloa frumentacea Link, Poaceae) germplasm in India. Genet. Resour. Crop Evol. 2009, 56, 883–889. [Google Scholar] [CrossRef]
- Howe, C.J.; Barbrook, A.C.; Koumandou, V.L.; Nisbet, R.E.R.; Symington, H.A.; Wightman, T.F. Evolution of the chloroplast genome. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2003, 358, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Plunkett, G.M.; Downie, S.R. Expansion and contraction of the chloroplast inverted repeat in Apiaceae subfamily Apioideae. Syst. Bot. 2000, 25, 648–667. [Google Scholar] [CrossRef]
- Wicke, S.; Schneeweiss, G.M.; Depamphilis, C.W.; Müller, K.F.; Quandt, D. The evolution of the plastid chromosome in land plants: Gene content, gene order, gene function. Plant Mol. Biol. 2011, 76, 273–297. [Google Scholar] [CrossRef]
- Cosner, M.E.; Raubeson, L.A.; Jansen, R.K. Chloroplast DNA rearrangements in Campanulaceae: Phylogenetic utility of highly rearranged genomes. BMC Evol. Biol. 2004, 4, 27. [Google Scholar] [CrossRef] [PubMed]
- Hong, Z.; Wu, Z.; Zhao, K.; Yang, Z.; Zhang, N.; Guo, J.; Tembrock, L.R.; Xu, D. Comparative analyses of five complete chloroplast genomes from the genus Pterocarpus (Fabacaeae). Int. J. Mol. Sci. 2020, 21, 3758. [Google Scholar] [CrossRef]
- Lu, R.-S.; Li, P.; Qiu, Y.-X. The complete chloroplast genomes of three Cardiocrinum (Liliaceae) species: Comparative genomic and phylogenetic analyses. Front. Plant Sci. 2017, 7, 2054. [Google Scholar] [CrossRef]
- Niu, Y.-T.; Jabbour, F.; Barrett, R.L.; Ye, J.-F.; Zhang, Z.-Z.; Lu, K.-Q.; Lu, L.-M.; Chen, Z.-D. Combining complete chloroplast genome sequences with target loci data and morphology to resolve species limits in Triplostegia (Caprifoliaceae). Mol. Phylogenetics Evol. 2018, 129, 15–26. [Google Scholar] [CrossRef]
- Pinard, D.; Myburg, A.A.; Mizrachi, E. The plastid and mitochondrial genomes of Eucalyptus grandis. BMC Genom. 2019, 20, 132. [Google Scholar] [CrossRef]
- Wu, F.-H.; Chan, M.-T.; Liao, D.-C.; Hsu, C.-T.; Lee, Y.-W.; Daniell, H.; Duvall, M.R.; Lin, C.-S. Complete chloroplast genome of Oncidium Gower Ramsey and evaluation of molecular markers for identification and breeding in Oncidiinae. BMC Plant Biol. 2010, 10, 68. [Google Scholar] [CrossRef]
- Li, P.; Lu, R.-S.; Xu, W.-Q.; Ohi-Toma, T.; Cai, M.-Q.; Qiu, Y.-X.; Cameron, K.M.; Fu, C.-X. Comparative genomics and phylogenomics of East Asian tulips (Amana, Liliaceae). Front. Plant Sci. 2017, 8, 451. [Google Scholar] [CrossRef]
- Bi, Y.; Zhang, M.-F.; Xue, J.; Dong, R.; Du, Y.-P.; Zhang, X.-H. Chloroplast genomic resources for phylogeny and DNA barcoding: A case study on Fritillaria. Sci. Rep. 2018, 8, 1184. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Liu, H.; Hu, J.; Liang, Y.; Liang, J.; Wuyun, T.; Tan, X. Five complete chloroplast genome sequences from Diospyros: Genome organization and comparative analysis. PLoS ONE 2016, 11, e0159566. [Google Scholar] [CrossRef] [PubMed]
- Maréchal, A.; Brisson, N. Recombination and the maintenance of plant organelle genome stability. New Phytol. 2010, 186, 299–317. [Google Scholar] [CrossRef]
- Duan, H.; Guo, J.; Xuan, L.; Wang, Z.; Li, M.; Yin, Y.; Yang, Y. Comparative chloroplast genomics of the genus Taxodium. BMC Genom. 2020, 21, 114. [Google Scholar] [CrossRef] [PubMed]
- Kugita, M.; Kaneko, A.; Yamamoto, Y.; Takeya, Y.; Matsumoto, T.; Yoshinaga, K. The complete nucleotide sequence of the hornwort (Anthoceros formosae) chloroplast genome: Insight into the earliest land plants. Nucleic Acids Res. 2003, 31, 716–721. [Google Scholar] [CrossRef]
- Henry, R.J. Plant Diversity and Evolution: Genotypic and Phenotypic Variation in Higher Plants; Cabi Publishing: Wallingford, UK, 2005. [Google Scholar]
- Yamane, K.; Yasui, Y.; Ohnishi, O. Intraspecific cpDNA variations of diploid and tetraploid perennial buckwheat, Fagopyrum cymosum (Polygonaceae). Am. J. Bot. 2003, 90, 339–346. [Google Scholar] [CrossRef]
- Li, L.; Hu, Y.; He, M.; Zhang, B.; Wu, W.; Cai, P.; Huo, D.; Hong, Y. Comparative chloroplast genomes: Insights into the evolution of the chloroplast genome of Camellia sinensis and the phylogeny of Camellia. BMC genomics 2021, 22, 138. [Google Scholar] [CrossRef]
- Liu, J.; Fang, J.; He, Z.; Li, J.; Dong, L. Target site–based resistance to penoxsulam in late watergrass (Echinochloa phyllopogon) from China. Weed Sci. 2019, 67, 1–9. [Google Scholar] [CrossRef]
- Qiong, P.; Heping, H.; Xia, Y.; Lianyang, B.; Qin, Y.; Powles, S.B. Quinclorac resistance in Echinochloa crus-galli from China. Rice Sci. 2019, 26, 300–308. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, Z.; Gu, T.; Dong, M.; Peng, Q.; Bai, L.; Li, Y. Quantitative proteomics reveals ecological fitness cost of multi-herbicide resistant barnyardgrass (Echinochloa crus-galli L.). J. Proteom. 2017, 150, 160–169. [Google Scholar] [CrossRef]
- Shinozaki, K.; Ohme, M.; Tanaka, M.; Wakasugi, T.; Hayashida, N.; Matsubayashi, T.; Zaita, N.; Chunwongse, J.; Obokata, J.; Yamaguchi-Shinozaki, K. The complete nucleotide sequence of the tobacco chloroplast genome: Its gene organization and expression. EMBO J. 1986, 5, 2043–2049. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, T.; Xu, L.; Zhan, Z.; Bao, G.; Zhang, T.; Ding, Z.; Sun, N.; Sun, S.; Xie, M. Comparative analysis of whole chloroplast genomes of Ligusticum sinense and L. jeholense, Umbelliferae. Res. Sq. 2020. [Google Scholar] [CrossRef]
- Fan, R.; Ma, W.; Liu, S.; Huang, Q. Integrated analysis of three newly sequenced fern chloroplast genomes: Genome structure and comparative analysis. Ecol. Evol. 2021, 11, 4550–4563. [Google Scholar] [CrossRef] [PubMed]
- NCBI. 2022. Available online: https://www.ncbi.nlm.nih.gov/nuccore/?term=Echinochloa+chloroplast+complete+genome (accessed on 18 October 2022).
- Wu, M.; Li, Q.; Hu, Z.; Li, X.; Chen, S. The complete Amomum kravanh chloroplast genome sequence and phylogenetic analysis of the commelinids. Molecules 2017, 22, 1875. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Cui, Y.; Chen, X.; Li, Y.; Xu, Z.; Duan, B.; Li, Y.; Song, J.; Yao, H. Complete chloroplast genomes of Papaver rhoeas and Papaver orientale: Molecular structures, comparative analysis, and phylogenetic analysis. Molecules 2018, 23, 437. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Liu, J.; Luo, L.; Wei, X.; Zhang, J.; Qi, Y.; Zhang, B.; Liu, H.; Xiao, P. Complete chloroplast genome sequences of Schisandra chinensis: Genome structure, comparative analysis, and phylogenetic relationship of basal angiosperms. Sci. China Life Sci. 2017, 60, 1286–1290. [Google Scholar] [CrossRef]
- Doorduin, L.; Gravendeel, B.; Lammers, Y.; Ariyurek, Y.; Chin-A-Woeng, T.; Vrieling, K. The complete chloroplast genome of 17 individuals of pest species Jacobaea vulgaris: SNPs, microsatellites and barcoding markers for population and phylogenetic studies. DNA Res. 2011, 18, 93–105. [Google Scholar] [CrossRef]
- He, S.; Wang, Y.; Volis, S.; Li, D.; Yi, T. Genetic diversity and population structure: Implications for conservation of wild soybean (Glycine soja Sieb. et Zucc) based on nuclear and chloroplast microsatellite variation. Int. J. Mol. Sci. 2012, 13, 12608–12628. [Google Scholar] [CrossRef]
- Yang, A.H.; Zhang, J.J.; Yao, X.H.; Huang, H.W. Chloroplast microsatellite markers in Liriodendron tulipifera (Magnoliaceae) and cross-species amplification in L. chinense. Am. J. Bot. 2011, 98, e123–e126. [Google Scholar] [CrossRef]
- Xue, J.; Wang, S.; Zhou, S.L. Polymorphic chloroplast microsatellite loci in Nelumbo (Nelumbonaceae). Am. J. Bot. 2012, 99, e240–e244. [Google Scholar] [CrossRef]
- Han, Y.; Gao, Y.; Zhai, X.; Zhou, H.; Ding, Q.; Ma, L. Assembly and comparative analysis of chloroplast genome of wheat K-CMS line and maintainer line. Res. Sq. 2020. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. Chloroplast evolution: Secondary symbiogenesis and multiple losses. Curr. Biol. 2002, 12, R62–R64. [Google Scholar] [CrossRef]
- Landegren, U.; Nilsson, M.; Kwok, P.-Y. Reading bits of genetic information: Methods for single-nucleotide polymorphism analysis. Genome Res. 1998, 8, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Germano, J.; Klein, A.S. Species-specific nuclear and chloroplast single nucleotide polymorphisms to distinguish Picea glauca, P. mariana and P. rubens. Theor. Appl. Genet. 1999, 99, 37–49. [Google Scholar] [CrossRef]
- Jeffares, D.C.; Tomiczek, B.; Sojo, V.; Reis, M.D. A beginners guide to estimating the non-synonymous to synonymous rate ratio of all protein-coding genes in a genome. In Parasite Genomics Protocols; Springer: Berlin/Heidelberg, Germany, 2015; pp. 65–90. [Google Scholar]
- McDonald, M.J.; Wang, W.-C.; Huang, H.-D.; Leu, J.-Y. Clusters of nucleotide substitutions and insertion/deletion mutations are associated with repeat sequences. PLoS Biol. 2011, 9, e1000622. [Google Scholar] [CrossRef]
- Wang, W.; Messing, J. High-throughput sequencing of three Lemnoideae (duckweeds) chloroplast genomes from total DNA. PLoS ONE 2011, 6, e24670. [Google Scholar] [CrossRef]
- Raubeson, L.A.; Peery, R.; Chumley, T.W.; Dziubek, C.; Fourcade, H.M.; Boore, J.L.; Jansen, R.K. Comparative chloroplast genomics: Analyses including new sequences from the angiosperms Nuphar advena and Ranunculus macranthus. BMC Genom. 2007, 8, 174. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, X.; Liu, G.; Yin, Y.; Chen, K.; Yun, Q.; Zhao, D.; Al-Mssallem, I.S.; Yu, J. The complete chloroplast genome sequence of date palm (Phoenix dactylifera L.). PLoS ONE 2010, 5, e12762. [Google Scholar] [CrossRef]
- Daniell, H.; Lin, C.-S.; Yu, M.; Chang, W.-J. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering. Genome Biol. 2016, 17, 134. [Google Scholar] [CrossRef]
- Logacheva, M.D.; Krinitsina, A.A.; Belenikin, M.S.; Khafizov, K.; Konorov, E.A.; Kuptsov, S.V.; Speranskaya, A.S. Comparative analysis of inverted repeats of polypod fern (Polypodiales) plastomes reveals two hypervariable regions. BMC Plant Biol. 2017, 17, 61–73. [Google Scholar] [CrossRef][Green Version]
- Kim, J.; Hwang, I.; Cho, K. Effect of storage conditions on the dormancy release and the induction of secondary dormancy in weed seeds. Korean J. Weed Sci. 1996, 16, 200–209. [Google Scholar]
- Li, H.-T.; Yi, T.-S.; Gao, L.-M.; Ma, P.-F.; Zhang, T.; Yang, J.-B.; Gitzendanner, M.A.; Fritsch, P.W.; Cai, J.; Luo, Y. Origin of angiosperms and the puzzle of the Jurassic gap. Nat. Plants 2019, 5, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, Y.; Henry, R.J.; Rossetto, M.; Wang, Y.; Chen, S. Plant DNA barcoding: From gene to genome. Biol. Rev. 2015, 90, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, Y.; Song, M.; Guan, Y.; Ma, X. Species identification of Dracaena using the complete chloroplast genome as a super-barcode. Front. Pharmacol. 2019, 10, 1441. [Google Scholar] [CrossRef]
- Yang, J.-B.; Yang, S.-X.; Li, H.-T.; Yang, J.; Li, D.-Z. Comparative chloroplast genomes of Camellia species. PLoS ONE 2013, 8, e73053. [Google Scholar] [CrossRef]
- Zhang, Y.; Song, M.-F.; Sun, H.-F.; Tang, D.-Y.; Xu, A.-S.; Zhang, Z.-L. Complete chloroplast genome analysis of two important medicinal Alpinia species: Alpinia galanga and Alpinia kwangsiensis. Front. Plant Sci. 2021, 12, 2908. [Google Scholar] [CrossRef]
- Gao, Y.; Li, J.; Pan, X.; Liu, D.; Napier, R.; Dong, L. Quinclorac resistance induced by the suppression of the expression of 1-aminocyclopropane-1-carboxylic acid (ACC) synthase and ACC oxidase genes in Echinochloa crus-galli var. zelayensis. Pestic. Biochem. Physiol. 2018, 146, 25–32. [Google Scholar] [CrossRef]
- Gao, Y.; Pan, X.; Sun, X.; Li, J.; Dong, L. Is the protection of photosynthesis related to the mechanism of quinclorac resistance in Echinochloa crus-galli var. zelayensis? Gene 2019, 683, 133–148. [Google Scholar] [CrossRef]
- Wortley, A.H.; Rudall, P.J.; Harris, D.J.; Scotland, R.W. How much data are needed to resolve a difficult phylogeny? Case study in Lamiales. Syst. Biol. 2005, 54, 697–709. [Google Scholar] [CrossRef]
- Petersen, G.; Aagesen, L.; Seberg, O.; Larsen, I.H. When is enough, enough in phylogenetics? A case in point from Hordeum (Poaceae). Cladistics 2011, 27, 428–446. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Tembrock, L.R.; Ge, S. Are differences in genomic data sets due to true biological variants or errors in genome assembly: An example from two chloroplast genomes. PLoS ONE 2015, 10, e0118019. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Echinochloa spp. | Length of Awn (cm) | Dry Weight Per 1000 Seeds (g) |
|---|---|---|
| E. crusgalli var. crusgalli | 1.63 ± 0.05 a | 2.806 ± 0.011 b |
| E. crusgalli var. zelayensis | 0.00 ± 0.00 b | 2.139 ± 0.016 c |
| E. glabrescens | 0.00 ± 0.00 b | 3.006 ± 0.032 a |
| Genome Features | E. crus-galli var. crus-galli | E. crus-galli var. zelayensis | E. glabrescens |
|---|---|---|---|
| Genome size (bp) | 139,856 | 139,874 | 139,874 |
| LSC length (bp) | 81,843 | 81,890 | 81,890 |
| SSC length (bp) | 12,517 | 12,514 | 12,514 |
| IR length (bp) | 22,748 | 22,735 | 22,735 |
| Intergenic region length (bp) | 79,775 | 79,751 | 79,751 |
| Overall GC content (%) | 38.63 | 38.63 | 38.63 |
| GC content of LSC (%) | 36.47 | 36.46 | 36.46 |
| GC content of SSC (%) | 33.21 | 33.22 | 33.22 |
| GC content of IRs (%) | 44.01 | 44.03 | 44.02 |
| Total genes | 132 | 132 | 132 |
| Number of protein-coding genes | 84 | 84 | 84 |
| Echinochloa spp. | Type | ncRNA Number | Total Length (bp) | Average Length (bp) | Length/ Genome (%) |
|---|---|---|---|---|---|
| E. crus-galli var. crus-galli | tRNA | 40 | 2972 * | 74 | 2.13 |
| rrn23 | 2 | 5778 | 2889 | 4.13 | |
| rrn4.5 | 2 | 190 | 95 | 0.14 | |
| rrn16 | 2 | 2982 | 1491 | 2.13 | |
| rrn5 | 2 | 242 | 121 | 0.17 | |
| E. crus-galli var. zelayensis | tRNA | 40 | 2976 | 74 | 2.13 |
| rrn23 | 2 | 5579 | 2789 | 3.99 | |
| rrn4.5 | 2 | 190 | 95 | 0.14 | |
| rrn16 | 2 | 2982 | 1491 | 2.13 | |
| rrn5 | 2 | 242 | 121 | 0.17 | |
| E. glabrescens | tRNA | 40 | 2976 | 74 | 2.13 |
| rrn23 | 2 | 5380 | 2690 | 3.85 | |
| rrn4.5 | 2 | 190 | 95 | 0.14 | |
| rrn16 | 2 | 2982 | 1491 | 2.13 | |
| rrn5 | 2 | 242 | 121 | 0.17 |
| Category | Groups | Genes |
|---|---|---|
| Photosynthesis | Subunits_of_photosystem_I | psaA, psaB, psaC, psaI, and psaJ |
| Subunits_of_photosystem_II | pbsN, psbA, psbB, psbC, psbD, psbE, psbF, psbH, psbI, psbJ, psbK, psbL, psbM, psbT, and psbZ | |
| Subunits_of_NADH_dehydrogenase | ndhA, ndhB, ndhB, ndhC, ndhD, ndhE, ndhF, ndhG, ndhH, ndhI, ndhJ, and ndhK | |
| Subunits_of_cytochrome_b/f_complex | petA, petB, petD, petG, petL, and petN | |
| Subunits_of_ATP_synthase | atpA, atpB, atpE, atpF, atpH, and atpI | |
| Large_subunit_of_Rubisco | rbcL | |
| Self-replication | Large_subunits_of_ribosome | rpl14, rpl16, rpl2, rpl2, rpl20, rpl22, rpl23, rpl23, rpl32, rpl33, and rpl36 |
| Small_subunits_of_ribosome | rps11, rps12, rps12, rps14, rps15, rps15, rps16, rps18, rps19, rps19, rps2, rps3, rps4, rps7, rps7, and rps8 | |
| DNA-dependent_RNA_polymerase | rpoA, rpoB, rpoC1, and rpoC2 | |
| Ribosomal_RNAs | 8 rRNA | |
| Transfer_RNAs | 40 tRNAs | |
| Other genes | Maturase | matK |
| Protease | clpP1 | |
| Envelope_membrane_protein | cemA | |
| Acetyl-CoA_carboxylase | ||
| C-type_cytochrome_synthesis_gene | ccsA | |
| Translation_initiation_factor | infA | |
| protochlorophillide_reductase_subunit | ||
| Genes of unknown function | Proteins_of_unknown_function | ycf2, ycf3, and ycf4 |
| Types of Repeats | E. crus-galli var. crus-galli | E. crus-galli var. zelayensis | E. glabrescens | |
|---|---|---|---|---|
| SSR Region Distribution | Genome | 136 | 139 | 139 |
| Coding | 43 | 42 | 42 | |
| IRa | 5 | 5 | 5 | |
| IRb | 5 | 5 | 5 | |
| LSC | 110 | 113 | 113 | |
| SSC | 16 | 16 | 16 | |
| LR Hamming Distance | 0 | 8 | 9 | 9 |
| 1 | 0 | 2 | 2 | |
| 2 | 21 | 21 | 21 | |
| 3 | 33 | 36 | 36 | |
| Barnyard Grass Species | Start | Stop | Synonymous | Non-Synonymous | CDS | Intergenic | Total_SNP |
|---|---|---|---|---|---|---|---|
| E. crus-galli var. crus-galli | 0 | 1 | 76 | 21 | 98 | 223 | 321 |
| E. glabrescens | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Shen, G.; Yuan, G.; Tian, Z. Comparative Analysis of Whole Chloroplast Genomes of Three Common Species of Echinochloa (Gramineae) in Paddy Fields. Int. J. Mol. Sci. 2022, 23, 13864. https://doi.org/10.3390/ijms232213864
Gao Y, Shen G, Yuan G, Tian Z. Comparative Analysis of Whole Chloroplast Genomes of Three Common Species of Echinochloa (Gramineae) in Paddy Fields. International Journal of Molecular Sciences. 2022; 23(22):13864. https://doi.org/10.3390/ijms232213864
Chicago/Turabian StyleGao, Yuan, Guohui Shen, Guohui Yuan, and Zhihui Tian. 2022. "Comparative Analysis of Whole Chloroplast Genomes of Three Common Species of Echinochloa (Gramineae) in Paddy Fields" International Journal of Molecular Sciences 23, no. 22: 13864. https://doi.org/10.3390/ijms232213864
APA StyleGao, Y., Shen, G., Yuan, G., & Tian, Z. (2022). Comparative Analysis of Whole Chloroplast Genomes of Three Common Species of Echinochloa (Gramineae) in Paddy Fields. International Journal of Molecular Sciences, 23(22), 13864. https://doi.org/10.3390/ijms232213864