
GDPD5 Related to Lipid Metabolism Is a Potential Prognostic Biomarker in Neuroblastoma

Abstract
1. Introduction
2. Results
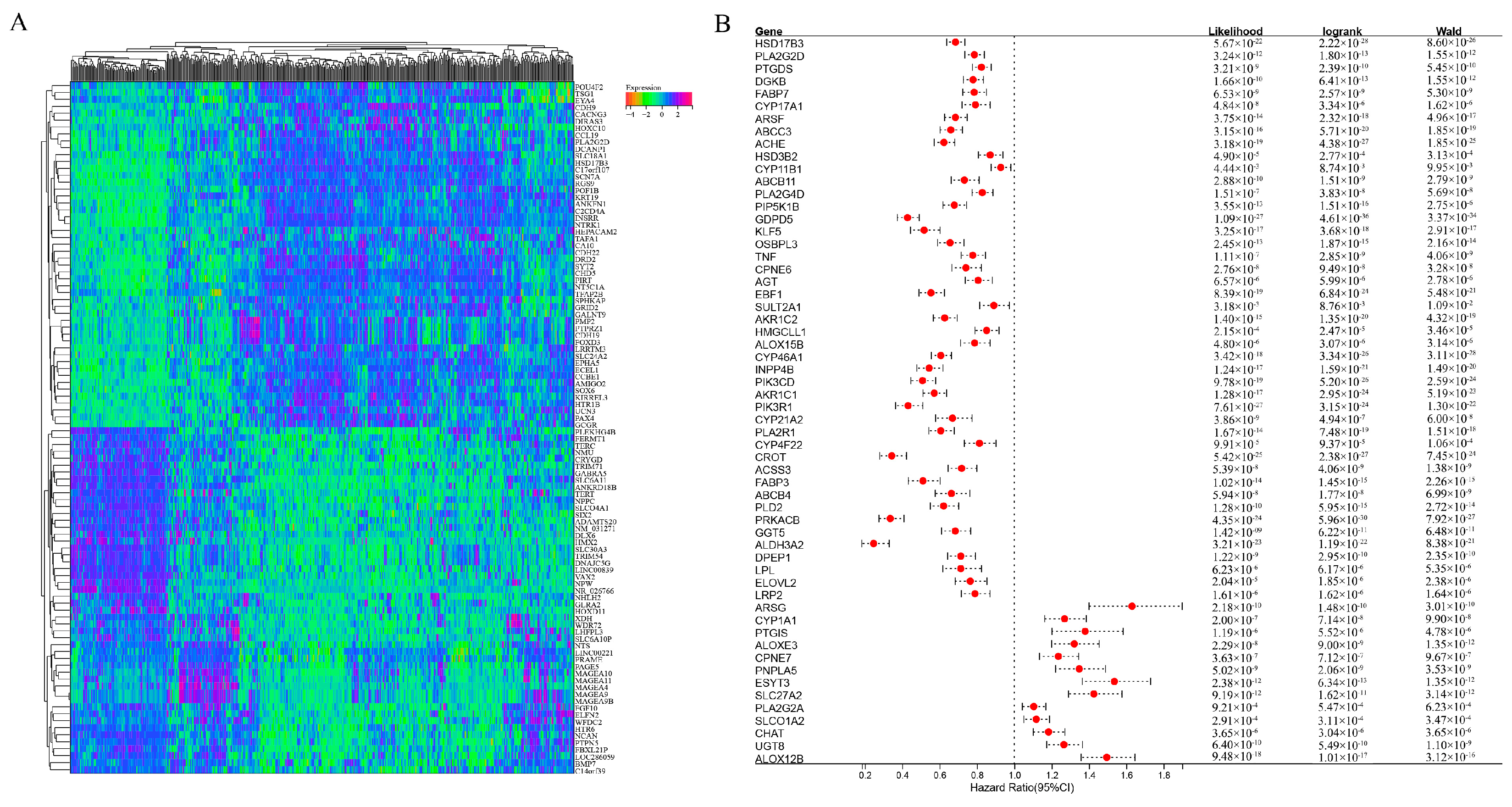
2.1. Identification of Differentially Expressed Genes Related to Lipid Metabolism between High-Risk Groups and Non-High-Risk Groups and Patient Survival in Neuroblastoma (NB)
2.2. Construction of a Prognostic Model for Lipid-Metabolism-Related Genes
2.3. Further Refinement of the Prognostic Model
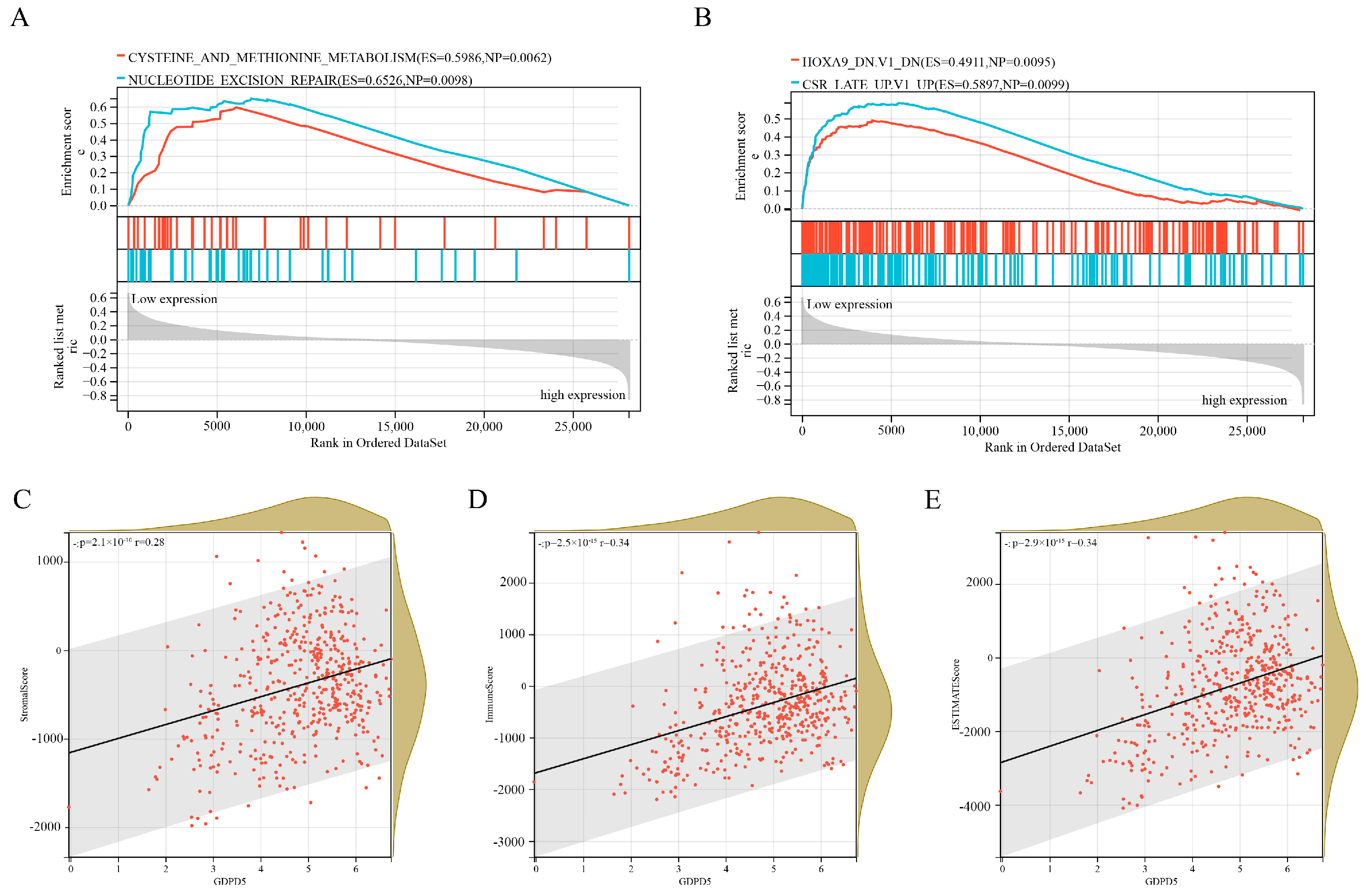
2.4. Independent Prognostic Factor GDPD5 Is Associated with Immune Infiltration
2.5. Hsa-miR-592 Is a Potential Target miRNA of GDPD5
2.6. Overexpression of GDPD5 Affects Lipid Metabolism, Migration and Proliferation of SH-SY5Y Cell Line
3. Discussion
4. Materials and Methods
4.1. Data Extraction from Online Databases
4.2. Screening of Differentially Expressed miRNAs and Lipid Metabolism Genes
4.3. Functional Enrichment Analysis and Protein–Protein Interaction Network Construction
4.4. Construction of a Prognostic Score for Lipid-metabolism-related Genes
4.5. Gene Set Enrichment Analysis (GSEA) and miRNA Prediction
4.6. Immune Infiltration and Immune Microenvironment Assessment
4.7. Plasmids Construction and Cell Culture Transfection
4.8. Western Blot Analysis
4.9. Transwell Assay
4.10. Cell Viability Assay
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| NB | Neuroblastoma |
| DEGs | Differentially Expressed Genes |
| PPI | Protein–protein interaction |
| FC | Fold Change |
| GEO | Gene Expression Omnibus |
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| DAVID | Database for Annotation, Visualization and Integrated Discovery |
| STRING | Search Tool for The Retrieval of Interaction Genes |
| OS | Overall Survival |
| DFI | Disease free interval |
| DFS | Disease free survival |
Appendix A



References
- Maris, J.; Hogarty, M.; Bagatell, R.; Cohn, S. Neuroblastoma. Lancet 2007, 369, 2106–2120. [Google Scholar] [CrossRef]
- Qiu, B.; Matthay, K.K. Advancing therapy for neuroblastoma. Nat. Rev. Clin. Oncol. 2022, 19, 515–533. [Google Scholar] [CrossRef]
- Chang, H.; Lu, M.; Yang, Y.; Chou, S.; Lin, D.; Lin, K.; Hsu, W.; Jeng, Y.; Jou, S. The prognostic roles of and correlation between and protein expression in neuroblastoma. J. Clin. Pathol. 2020, 73, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Sokol, E.; Desai, A.V.; Applebaum, M.A.; Valteau-Couanet, D.; Park, J.R.; Pearson, A.D.; Schleiermacher, G.; Irwin, M.S.; Hogarty, M.; Naranjo, A.; et al. Age, Diagnostic Category, Tumor Grade, and Mitosis-Karyorrhexis Index Are Independently Prognostic in Neuroblastoma: An INRG Project. J. Clin. Oncol. 2020, 38, 1906–1918. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Lin, X.; Zhou, X.; Prochownik, E.V.; Wang, F.; Li, Y. Posttranslational control of lipogenesis in the tumor microenvironment. J. Hematol. Oncol. 2022, 15, 120. [Google Scholar] [CrossRef]
- Agostini, M.; Melino, G.; Habeb, B.; Calandria, J.M.; Bazan, N.G. Targeting lipid metabolism in cancer: Neuroblastoma. Cancer Metastasis Rev. 2022, 41, 255–260. [Google Scholar] [CrossRef]
- Yang, F.; Hu, A.; Guo, Y.; Wang, J.; Li, D.; Wang, X.; Jin, S.; Yuan, B.; Cai, S.; Zhou, Y.; et al. p113 isoform encoded by CUX1 circular RNA drives tumor progression via facilitating ZRF1/BRD4 transactivation. Mol. Cancer 2021, 20, 123. [Google Scholar] [CrossRef]
- Shulkin, B.L.; Mitchell, D.S.; Ungar, D.R.; Prakash, D.; Dole, M.G.; Castle, V.P.; Hernandez, R.J.; A Koeppe, R.; Hutchinson, R.J. Neoplasms in a pediatric population: 2-[F-18]-fluoro-2-deoxy-D-glucose PET studies. Radiology 1995, 194, 495–500. [Google Scholar] [CrossRef]
- Ruiz-Pérez, M.V.; Sainero-Alcolado, L.; Oliynyk, G.; Matuschek, I.; Balboni, N.; Ubhayasekera, S.K.A.; Snaebjornsson, M.T.; Makowski, K.; Aaltonen, K.; Bexell, D.; et al. Inhibition of fatty acid synthesis induces differentiation and reduces tumor burden in childhood neuroblastoma. iScience 2021, 24, 102128. [Google Scholar] [CrossRef]
- Rugolo, F.; Bazan, N.G.; Calandria, J.; Jun, B.; Raschellà, G.; Melino, G.; Agostini, M. The expression of ELOVL4, repressed by MYCN, defines neuroblastoma patients with good outcome. Oncogene 2021, 40, 5741–5751. [Google Scholar] [CrossRef]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef] [PubMed]
- McDermaid, A.; Monier, B.; Zhao, J.; Liu, B.; Ma, Q. Interpretation of differential gene expression results of RNA-seq data: Review and integration. Brief. Bioinform. 2019, 20, 2044–2054. [Google Scholar] [CrossRef] [PubMed]
- Gallazzini, M.; Ferraris, J.D.; Burg, M.B. GDPD5 is a glycerophosphocholine phosphodiesterase that osmotically regulates the osmoprotective organic osmolyte GPC. Proc. Natl. Acad. Sci. USA 2008, 105, 11026–11031. [Google Scholar] [CrossRef] [PubMed]
- Sabharwal, P.; Lee, C.; Park, S.; Rao, M.; Sockanathan, S. GDE2 Regulates Subtype-Specific Motor Neuron Generation through Inhibition of Notch Signaling. Neuron 2011, 71, 1058–1070. [Google Scholar] [CrossRef]
- Yanaka, N.; Nogusa, Y.; Fujioka, Y.; Yamashita, Y.; Kato, N. Involvement of membrane protein GDE2 in retinoic acid-induced neurite formation in Neuro2A cells. FEBS Lett. 2007, 581, 712–718. [Google Scholar] [CrossRef]
- Choi, B.-R.; Cave, C.; Na, C.H.; Sockanathan, S. GDE2-Dependent Activation of Canonical Wnt Signaling in Neurons Regulates Oligodendrocyte Maturation. Cell Rep. 2020, 31, 107540. [Google Scholar] [CrossRef]
- Rao, M.; Sockanathan, S. Transmembrane Protein GDE2 Induces Motor Neuron Differentiation in Vivo. Science 2005, 309, 2212–2215. [Google Scholar] [CrossRef]
- Cave, C.; Park, S.; Rodriguez, M.; Nakamura, M.; Hoke, A.; Pletnikov, M.; Sockanathan, S. GDE2 is essential for neuronal survival in the postnatal mammalian spinal cord. Mol. Neurodegener. 2017, 12, 8. [Google Scholar] [CrossRef]
- Matas-Rico, E.; van Veen, M.; Leyton-Puig, D.; van den Berg, J.; Koster, J.; Kedziora, K.M.; Molenaar, B.; Weerts, M.J.A.; Rink, I.; Medema, R.H.; et al. Glycerophosphodiesterase GDE2 Promotes Neuroblastoma Differentiation through Glypican Release and Is a Marker of Clinical Outcome. Cancer Cell 2016, 30, 548–562. [Google Scholar] [CrossRef]
- Zheng, M.; Mullikin, H.; Hester, A.; Czogalla, B.; Heidegger, H.; Vilsmaier, T.; Vattai, A.; Chelariu-Raicu, A.; Jeschke, U.; Trillsch, F.; et al. Development and Validation of a Novel 11-Gene Prognostic Model for Serous Ovarian Carcinomas Based on Lipid Metabolism Expression Profile. Int. J. Mol. Sci. 2020, 21, 9169. [Google Scholar] [CrossRef]
- Asgharzadeh, S.; Salo, J.; Ji, L.; Oberthuer, A.; Fischer, M.; Berthold, F.; Hadjidaniel, M.; Liu, C.W.; Metelitsa, L.S.; Pique-Regi, R.; et al. Clinical significance of tumor-associated inflammatory cells in metastatic neuroblastoma. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2012, 30, 3525–3532. [Google Scholar] [CrossRef] [PubMed]
- Wienke, J.; Dierselhuis, M.P.; Tytgat, G.A.; Künkele, A.; Nierkens, S.; Molenaar, J.J. The immune landscape of neuroblastoma: Challenges and opportunities for novel therapeutic strategies in pediatric oncology. Eur. J. Cancer 2020, 144, 123–150. [Google Scholar] [CrossRef] [PubMed]
- Yenyuwadee, S.; Aliazis, K.; Wang, Q.; Christofides, A.; Shah, R.; Patsoukis, N.; Boussiotis, V.A. Immune cellular components and signaling pathways in the tumor microenvironment. Semin. Cancer Biol. 2022, 86, 187–201. [Google Scholar] [CrossRef] [PubMed]
- Fabian, M.R.; Sonenberg, N.; Filipowicz, W. Regulation of mRNA Translation and Stability by microRNAs. Annu. Rev. Biochem. 2010, 79, 351–379. [Google Scholar] [CrossRef]
- Campos Cogo, S.; Gradowski Farias da Costa do Nascimento, T.; de Almeida Brehm Pinhatti, F.; de França Junior, N.; Santos Rodrigues, B.; Cavalli, L.R.; Elifio-Esposito, S. An overview of neuroblastoma cell lineage phenotypes and in vitro models. Exp. Biol. Med. 2020, 245, 1637–1647. [Google Scholar] [CrossRef]
- Batchuluun, B.; Pinkosky, S.L.; Steinberg, G.R. Lipogenesis inhibitors: Therapeutic opportunities and challenges. Nat. Rev. Drug Discov. 2022, 21, 283–305. [Google Scholar] [CrossRef]
- Wang, X.; Song, H.; Liang, J.; Jia, Y.; Zhang, Y. Abnormal expression of HADH, an enzyme of fatty acid oxidation, affects tumor development and prognosis (Review). Mol. Med. Rep. 2022, 26, 355. [Google Scholar] [CrossRef]
- Park, S.; Baek, I.J.; Ryu, J.H.; Chun, C.H.; Jin, E.J. PPARalpha-ACOT12 axis is responsible for maintaining cartilage homeostasis through modulating de novo lipogenesis. Nat. Commun. 2022, 13, 3. [Google Scholar] [CrossRef]
- Bolanos, J.P.; Almeida, A.; Moncada, S. Glycolysis: A bioenergetic or a survival pathway? Trends Biochem. Sci. 2010, 35, 145–149. [Google Scholar] [CrossRef]
- Tarnopolsky, M.A. Myopathies Related to Glycogen Metabolism Disorders. Neurotherapeutics 2018, 15, 915–927. [Google Scholar] [CrossRef]
- Qin, J.; Chai, G.; Brewer, J.M.; Lovelace, L.L.; Lebioda, L. Structures of asymmetric complexes of human neuron specific enolase with resolved substrate and product and an analogous complex with two inhibitors indicate subunit interaction and inhibitor cooperativity. J. Inorg. Biochem. 2012, 111, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Tan, V.P.; Miyamoto, S. HK2/hexokinase-II integrates glycolysis and autophagy to confer cellular protection. Autophagy 2015, 11, 963–964. [Google Scholar] [CrossRef]
- Pouysségur, J.; Marchiq Parks, S.; Durivault, J.; Ždralević, M.; Vucetic, M. ‘Warburg effect’ controls tumor growth, bacterial, viral infections and immunity—Genetic deconstruction and therapeutic perspectives. Semin. Cancer Biol. 2022, 86, 334–346. [Google Scholar] [CrossRef] [PubMed]
- Irwin, M.; Naranjo, A.; Zhang, F.; Cohn, S.; London, W.; Gastier-Foster, J.; Ramirez, N.C.; Pfau, R.; Reshmi, S.; Wagner, E.; et al. Revised Neuroblastoma Risk Classification System: A Report from the Children’s Oncology Group. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2021, 39, 3229–3241. [Google Scholar] [CrossRef]
- Pinto, N.; Applebaum, M.; Volchenboum, S.; Matthay, K.; London, W.; Ambros, P.; Nakagawara, A.; Berthold, F.; Schleiermacher, G.; Park, J.R.; et al. Advances in Risk Classification and Treatment Strategies for Neuroblastoma. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2015, 33, 3008–3017. [Google Scholar] [CrossRef] [PubMed]
- Shao, F.; Wang, Z.; Wang, S. MYCNIdentification of -Related Gene as a Potential Biomarker for Neuroblastoma Prognostic Model by Integrated Analysis and Quantitative Real-Time PCR. DNA Cell Biol. 2021, 40, 332–347. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Li, X.; Tian, X.; Zhao, Q. Identification of a Five-Gene Signature Derived from MYCN Amplification and Establishment of a Nomogram for Predicting the Prognosis of Neuroblastoma. Front. Mol. Biosci. 2021, 8, 1185. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Zhao, Q.; Xu, Y.; Zhu, L. A signature of 29 immune-related genes pairs to predict prognosis in patients with neuroblastoma. Int. Immunopharmacol. 2020, 88, 106994. [Google Scholar] [CrossRef] [PubMed]
- Sha, Y.; Han, L.; Sun, B.; Zhao, Q. Identification of a Glycosyltransferase Signature for Predicting Prognosis and Immune Microenvironment in Neuroblastoma. Front. Cell Dev. Biol. 2022, 9, 769580. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y. Adipocyte and lipid metabolism in cancer drug resistance. J. Clin. Investig. 2019, 129, 3006–3017. [Google Scholar] [CrossRef]
- Broadfield, L.A.; Pane, A.A.; Talebi, A.; Swinnen, J.V.; Fendt, S.-M. Lipid metabolism in cancer: New perspectives and emerging mechanisms. Dev. Cell 2021, 56, 1363–1393. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Cheng, C.; Tan, Z.; Li, N.; Tang, M.; Yang, L.; Cao, Y. Emerging roles of lipid metabolism in cancer metastasis. Mol. Cancer 2017, 16, 76. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Liu, Y.; Wen, S.; Xu, C.; Gu, L. In silico development and clinical validation of novel 8 gene signature based on lipid metabolism related genes in colon adenocarcinoma. Pharmacol. Res. 2021, 169, 105644. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Zeng, Q.; Fan, T.; Lei, Y.; Wang, F.; Zheng, S.; Wang, X.; Zeng, H.; Tan, F.; Sun, N.; et al. Clinical Significance and Immunometabolism Landscapes of a Novel Recurrence-Associated Lipid Metabolism Signature in Early-Stage Lung Adenocarcinoma: A Comprehensive Analysis. Front. Immunol. 2022, 13, 783495. [Google Scholar] [CrossRef] [PubMed]
- Attiyeh, E.F.; London, W.B.; Mossé, Y.P.; Wang, Q.; Winter, C.; Khazi, D.; McGrady, P.W.; Seeger, R.C.; Look, A.T.; Shimada, H.; et al. Chromosome 1p and 11q Deletions and Outcome in Neuroblastoma. N. Engl. J. Med. 2005, 353, 2243–2253. [Google Scholar] [CrossRef]
- Jiang, M.; Stanke, J.; Lahti, J.M. The Connections between Neural Crest Development and Neuroblastoma. Curr. Top. Dev. Biol. 2011, 94, 77–127. [Google Scholar] [CrossRef]
- Zhang, W.; Yu, Y.; Hertwig, F.; Thierry-Mieg, J.; Zhang, W.; Thierry-Mieg, D.; Wang, J.; Furlanello, C.; Devanarayan, V.; Cheng, J.; et al. Comparison of RNA-seq and microarray-based models for clinical endpoint prediction. Genome Biol. 2015, 16, 133. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Belinda, P.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef]
- Lu, T.-P.; Lee, C.-Y.; Tsai, M.-H.; Chiu, Y.-C.; Hsiao, C.K.; Lai, L.-C.; Chuang, E.Y. miRSystem: An Integrated System for Characterizing Enriched Functions and Pathways of MicroRNA Targets. PLoS ONE 2012, 7, e42390. [Google Scholar] [CrossRef]
- Newman, A.M.; Liu, C.L.; Green, M.R.; Gentles, A.J.; Feng, W.; Xu, Y.; Hoang, C.D.; Diehn, M.; Alizadeh, A.A. Robust enumeration of cell subsets from tissue expression profiles. Nat. Methods 2015, 12, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Yoshihara, K.; Shahmoradgoli, M.; Martínez, E.; Vegesna, R.; Kim, H.; Torres-Garcia, W.; Trevino, V.; Shen, H.; Laird, P.W.; Levine, D.A.; et al. Inferring tumour purity and stromal and immune cell admixture from expression data. Nat. Commun. 2013, 4, 2612. [Google Scholar] [CrossRef] [PubMed]
- Racle, J.; De Jonge, K.; Baumgaertner, P.; Speiser, D.E.; Gfeller, D. Simultaneous enumeration of cancer and immune cell types from bulk tumor gene expression data. eLife 2017, 6, e26476. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Features | Number (%) |
|---|---|
| Sex | |
| Female | 211 (42.4%) |
| Male | 287 (57.6%) |
| Age | |
| ≤18 months | 300 (60.2%) |
| >18 months | 198 (39.8%) |
| MYCN amplification | |
| No | 401 (80.5%) |
| Unknown | 5 (1.0%) |
| Yes | 92 (18.5%) |
| INSS stage | |
| St1 | 121 (24.3%) |
| St2 | 78 (15.7%) |
| St3 | 63 (12.7%) |
| St4 | 183 (36.7%) |
| St4S | 53 (10.6%) |
| Clinical risk | |
| High | 176 (35.3%) |
| Low | 322 (64.7%) |
| Class label | |
| Favorable | 181 (36.3%) |
| Unfavorable | 91 (18.3%) |
| Unknown | 226 (45.4%) |
| Progression | |
| No | 315 (63.3%) |
| Yes | 183 (36.7%) |
| Death from disease | |
| No | 393 (78.9%) |
| Yes | 105 (21.1%) |
| Low Risk Score | High Risk Score | Accuracy | |
|---|---|---|---|
| COG (High) | 26 | 150 | 85.23% |
| COG (Low and medium) | 301 | 21 | 93.48% |
| Total | 327 | 171 | 498 (100%) |
| Low Risk Score | High Risk Score | Accuracy | |
|---|---|---|---|
| COG (High) | 8 | 168 | 95.45% |
| COG (Low and medium) | 241 | 81 | 74.84% |
| Total | 327 | 171 | 498 (100%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, T.; Peng, J.; Li, Q.; Zhang, Y.; Huang, Y.; Xu, L.; Yang, G.; Tan, D.; Zhang, Q.; Tan, Y. GDPD5 Related to Lipid Metabolism Is a Potential Prognostic Biomarker in Neuroblastoma. Int. J. Mol. Sci. 2022, 23, 13740. https://doi.org/10.3390/ijms232213740
Luo T, Peng J, Li Q, Zhang Y, Huang Y, Xu L, Yang G, Tan D, Zhang Q, Tan Y. GDPD5 Related to Lipid Metabolism Is a Potential Prognostic Biomarker in Neuroblastoma. International Journal of Molecular Sciences. 2022; 23(22):13740. https://doi.org/10.3390/ijms232213740
Chicago/Turabian StyleLuo, Tengling, Junwei Peng, Qijun Li, Yao Zhang, Yun Huang, Lei Xu, Genling Yang, Dongmei Tan, Qian Zhang, and Yi Tan. 2022. "GDPD5 Related to Lipid Metabolism Is a Potential Prognostic Biomarker in Neuroblastoma" International Journal of Molecular Sciences 23, no. 22: 13740. https://doi.org/10.3390/ijms232213740
APA StyleLuo, T., Peng, J., Li, Q., Zhang, Y., Huang, Y., Xu, L., Yang, G., Tan, D., Zhang, Q., & Tan, Y. (2022). GDPD5 Related to Lipid Metabolism Is a Potential Prognostic Biomarker in Neuroblastoma. International Journal of Molecular Sciences, 23(22), 13740. https://doi.org/10.3390/ijms232213740