


MicroRNA-381 Regulates Proliferation and Differentiation of Caprine Skeletal Muscle Satellite Cells by Targeting PTEN and JAG2

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Isolation, Identification and Myogenic Differentiation of Caprine SMSCs
2.2. Expression Profiles of MyoG, MyHC and miR-381 during Myogenic Differentiation
2.3. miR-381 Accelerates Myogenic Differentiation of Caprine SMSCs
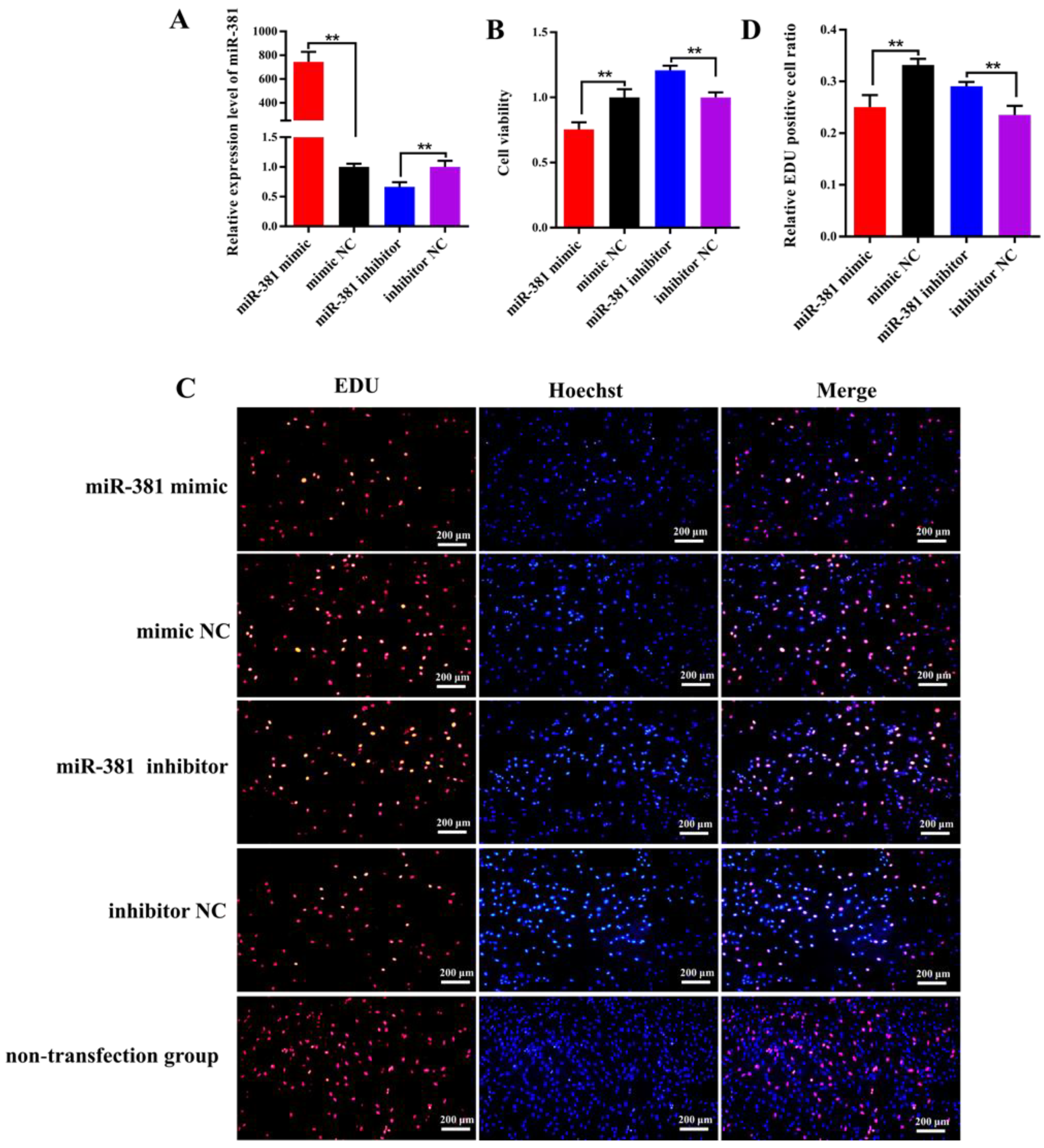
2.4. miR-381 Suppresses Viability and Proliferation of Caprine SMSCs
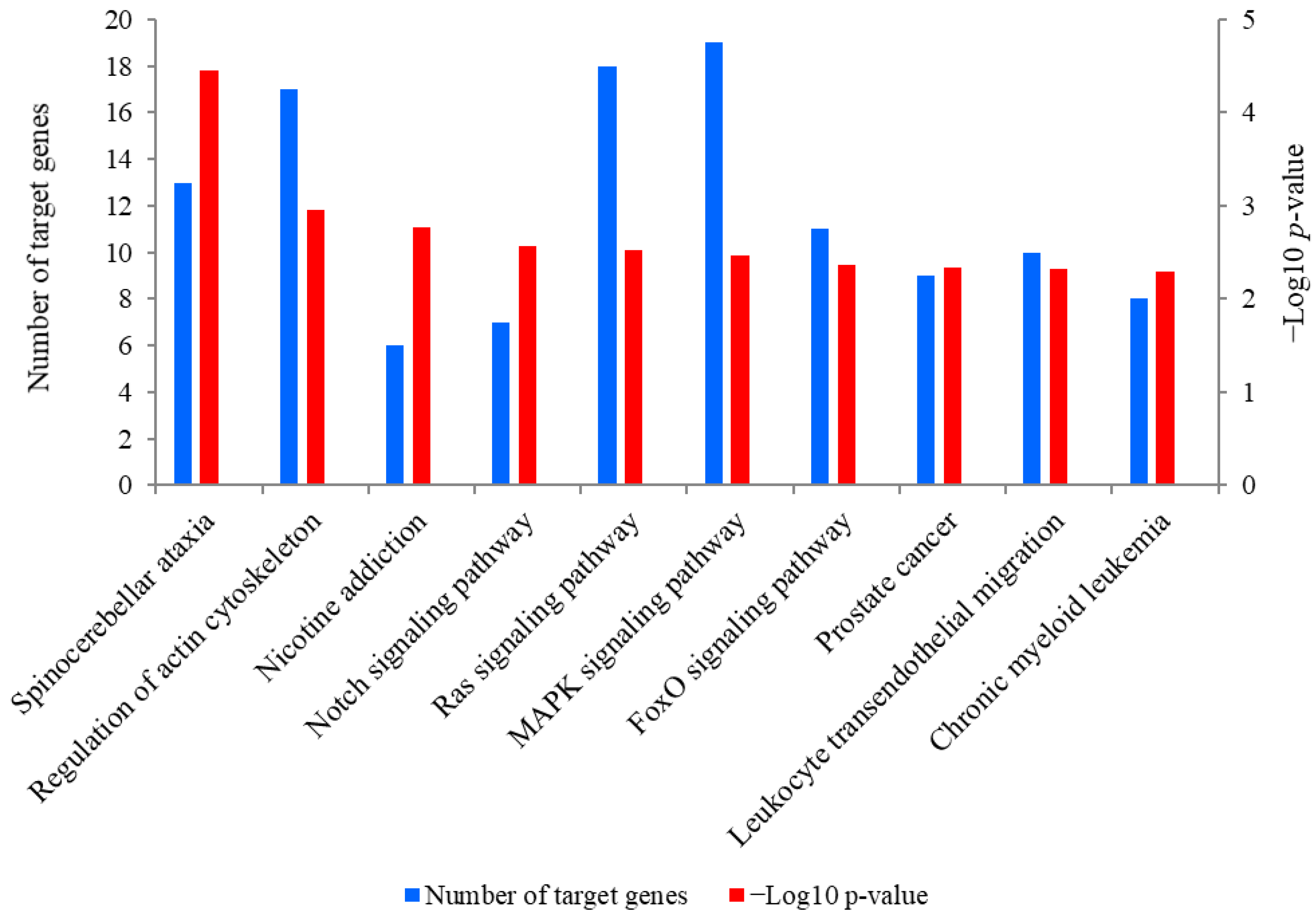
2.5. Prediction and KEGG Pathway Analysis of the Target Genes of miR-381
2.6. Validation of Target Relationships of miR-381 with the Target Genes
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Isolation, Identification and Myogenic Differentiation of Caprine SMSCs
4.3. The Effect of miR-381 on Caprine SMSC Differentiation
4.4. The Effect of miR-381 on the Viability and Proliferation of Caprine SMSCs
4.5. Prediction and Validation of the Target Genes of miR-381
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Adams, G.R. Satellite cell proliferation and skeletal muscle hypertrophy. Appl. Physiol. Nutr. Metab. 2006, 31, 782–790. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Tong, H.; Li, S.; Yan, Y. Effect of ECM2 expression on bovine skeletal muscle-derived satellite cell differentiation. Cell Biol. Int. 2018, 42, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Hou, L.; Ou, J.; Xu, G.; Jiang, F.; Hu, C.; Wang, C. MiR-199b represses porcine muscle satellite cells proliferation by targeting JAG1. Gene 2019, 691, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Ran, J.; Li, J.; Yu, C.; Cui, Z.; Amevor, F.K.; Wang, Y.; Jiang, X.; Qiu, M.; Du, H.; et al. miR-21-5p regulates the proliferation and differentiation of skeletal muscle satellite cells by targeting KLF3 in chicken. Genes 2021, 12, 814. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.; Luo, Y.; Wang, J.; Hickford, J.G.H.; Zhou, H.; Hu, J.; Liu, X.; Li, S.; Shen, J.; Ke, N.; et al. MicroRNA-432 inhibits milk fat synthesis by targeting SCD and LPL in ovine mammary epithelial cells. Food Funct. 2021, 12, 9432–9442. [Google Scholar] [CrossRef]
- Hou, L.; Xu, J.; Li, H.; Ou, J.; Jiao, Y.; Hu, C.; Wang, C. MiR-34c represses muscle development by forming a regulatory loop with Notch1. Sci. Rep. 2017, 7, 9346. [Google Scholar] [CrossRef]
- Dai, Y.; Zhang, W.R.; Wang, Y.M.; Liu, X.F.; Li, X.; Ding, X.B.; Guo, H. MicroRNA-128 regulates the proliferation and differentiation of bovine skeletal muscle satellite cells by repressing Sp1. Mol. Cell. Biochem. 2016, 414, 37–46. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, S.Y.; Deng, S.Y.; Gao, L.; Yang, L.W.; Liu, X.N.; Shi, G.Q. MiR-27b promotes sheep skeletal muscle satellite cell proliferation by targeting myostatin gene. J. Genet. 2018, 97, 1107–1117. [Google Scholar] [CrossRef]
- Sui, M.; Zheng, Q.; Wu, H.; Zhu, L.; Ling, Y.; Wang, L.; Fang, F.; Liu, Y.; Zhang, Z.; Chu, M.; et al. The expression and regulation of miR-1 in goat skeletal muscle and satellite cell during muscle growth and development. Anim. Biotechnol. 2020, 31, 455–462. [Google Scholar] [CrossRef]
- Shen, J.; Hao, Z.; Luo, Y.; Zhen, H.; Liu, Y.; Wang, J.; Hu, J.; Liu, X.; Li, S.; Zhao, Z.; et al. Deep small RNA sequencing reveals important miRNAs related to muscle development and intramuscular fat deposition in longissimus dorsi muscle from different goat breeds. Front. Vet. Sci. 2022, 9, 911166. [Google Scholar] [CrossRef]
- Guo, J.; Zhao, W.; Zhan, S.; Li, L.; Zhong, T.; Wang, L.; Dong, Y.; Zhang, H. Identification and expression profiling of miRNAome in goat longissimus dorsi muscle from prenatal stages to a neonatal stage. PLoS ONE 2016, 11, e0165764. [Google Scholar] [CrossRef]
- Ling, Y.; Zheng, Q.; Jing, J.; Sui, M.; Zhu, L.; Li, Y.; Zhang, Y.; Liu, Y.; Fang, F.; Zhang, X. RNA-Seq reveals miRNA role shifts in seven stages of skeletal muscles in goat fetuses and kids. Front. Genet. 2020, 11, 684. [Google Scholar] [CrossRef]
- Serrano, A.L.; Baeza-Raja, B.; Perdiguero, E.; Jardí, M.; Muñoz-Cánoves, P. Interleukin-6 is an essential regulator of satellite cell-mediated skeletal muscle hypertrophy. Cell Metab. 2008, 7, 33–44. [Google Scholar] [CrossRef]
- Ling, Y.H.; Sui, M.H.; Zheng, Q.; Wang, K.Y.; Wu, H.; Li, W.Y.; Liu, Y.; Chu, M.X.; Fang, F.G.; Xu, L.N. miR-27b regulates myogenic proliferation and differentiation by targeting Pax3 in goat. Sci. Rep. 2018, 8, 3909. [Google Scholar] [CrossRef]
- Yang, J.; Liu, H.; Wang, K.; Li, L.; Yuan, H.; Liu, X.; Liu, Y.; Guan, W. Isolation, culture and biological characteristics of multipotent porcine skeletal muscle satellite cells. Cell Tissue Bank. 2017, 18, 513–525. [Google Scholar] [CrossRef]
- Wu, H.; Ren, Y.; Li, S.; Wang, W.; Yuan, J.; Guo, X.; Liu, D.; Cang, M. In vitro culture and induced differentiation of sheep skeletal muscle satellite cells. Cell. Biol. Int. 2012, 36, 579–587. [Google Scholar] [CrossRef]
- Asfour, H.A.; Allouh, M.Z.; Said, R.S. Myogenic regulatory factors: The orchestrators of myogenesis after 30 years of discovery. Exp. Biol. Med. 2018, 243, 118–128. [Google Scholar] [CrossRef]
- Wang, Y.; Xiao, X.; Wang, L. In vitro characterization of goat skeletal muscle satellite cells. Anim. Biotechnol. 2020, 31, 115–121. [Google Scholar] [CrossRef]
- Xu, H.; Shao, J.; Fang, J.; Yin, B.; Zhang, L.; Zhang, J.; Xia, G. miR-381 Targets KCTD15 to regulate bovine preadipocyte differentiation in vitro. Horm. Metab. Res. 2021, 53, 63–70. [Google Scholar] [CrossRef]
- Jiang, H.; Yuan, X.; Fu, Y. Down-regulation of miR-381-3p inhibits osteogenic differentiation of mouse embryonic palatal mesenchymal cells in 2, 3, 7, 8-tetrachlorodibenzo-p-dioxin-induced cleft palate of fetal mice. Chin. J. Reparative Reconstr. Surg. 2019, 33, 1174–1180. [Google Scholar]
- Wang, J.; Sun, N.; Ju, Y.; Ni, N.; Tang, Z.; Zhang, D.; Dai, X.; Chen, M.; Wang, Y.; Gu, P.; et al. miR-381-3p cooperated with Hes1 to regulate the proliferation and differentiation of retinal progenitor cells. Front. Cell Dev. Biol. 2022, 10, 853215. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.W.; Xiao, H.; Zhang, Z.X. An experimental research on different detection conditions between MTT and CCK-8. Acta Laser Biol. Sin. 2007, 16, 559–562. [Google Scholar]
- Biram, A.; Shulman, Z. Evaluation of B Cell Proliferation in vivo by EdU Incorporation Assay. Bio. Protoc. 2020, 10, e3602. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Zhu, Q.; Guo, C.; Yuan, R.; Zhang, X.; Nie, Y.; Chen, L.; Fang, Y.; Chen, K.; Zhang, J.; et al. MLL1 promotes myogenesis by epigenetically regulating Myf5. Cell Prolif. 2020, 53, e12744. [Google Scholar] [CrossRef]
- Naito, M.; Mori, M.; Inagawa, M.; Miyata, K.; Hashimoto, N.; Tanaka, S.; Asahara, H. Dnmt3a regulates proliferation of muscle satellite cells via p57Kip2. PLoS Genet. 2016, 12, e1006167. [Google Scholar] [CrossRef]
- Fang, Z.; Zhong, M.; Wang, Y.; Yuan, X.; Guo, H.; Yao, Y.; Feng, M.; Chen, J.; Xiong, J.; Xiang, X. miR-381 and miR-489 suppress cell proliferation and invasion by targeting CUL4B via the Wnt/β-catenin pathway in gastric cancer. Int. J. Oncol. 2019, 54, 733–743. [Google Scholar] [CrossRef]
- Zhang, P.F.; Pei, X.; Li, K.S.; Jin, L.N.; Wang, F.; Wu, J.; Zhang, X.M. Circular RNA circFGFR1 promotes progression and anti-PD-1 resistance by sponging miR-381-3p in non-small cell lung cancer cells. Mol. Cancer 2019, 18, 179. [Google Scholar] [CrossRef]
- Xue, Y.; Xu, W.; Zhao, W.; Wang, W.; Zhang, D.; Wu, P. miR-381 inhibited breast cancer cells proliferation, epithelial-to-mesenchymal transition and metastasis by targeting CXCR4. Biomed. Pharmacother. 2017, 86, 426–433. [Google Scholar] [CrossRef]
- Zhu, X.S.; Zhou, H.Y.; Yang, F.; Zhang, H.S.; Ma, K.Z. miR-381-3p inhibits high glucose-induced vascular smooth muscle cell proliferation and migration by targeting HMGB1. J. Gene Med. 2021, 23, e3274. [Google Scholar] [CrossRef]
- Bai, S.Y.; Li, M.L.; Ren, Y.; Su, X.M. HDAC8-inhibitor PCI-34051-induced exosomes inhibit human bronchial smooth muscle cell proliferation via miR-381-3p mediated TGFB3. Pulm. Pharmacol. Ther. 2021, 71, 102096. [Google Scholar] [CrossRef]
- Fujimaki, S.; Seko, D.; Kitajima, Y.; Yoshioka, K.; Tsuchiya, Y.; Masuda, S.; Ono, Y. Notch1 and Notch2 coordinately regulate stem cell function in the quiescent and activated states of muscle satellite cells. Stem Cells 2018, 36, 278–285. [Google Scholar] [CrossRef]
- Wen, Y.; Bi, P.; Liu, W.; Asakura, A.; Keller, C.; Kuang, S. Constitutive Notch activation upregulates Pax7 and promotes the self-renewal of skeletal muscle satellite cells. Mol. Cell Biol. 2012, 32, 2300–2311. [Google Scholar] [CrossRef]
- Mourikis, P.; Sambasivan, R.; Castel, D.; Rocheteau, P.; Bizzarro, V.; Tajbakhsh, S. A critical requirement for notch signaling in maintenance of the quiescent skeletal muscle stem cell state. Stem Cells 2012, 30, 243–252. [Google Scholar] [CrossRef]
- Schuster-Gossler, K.; Cordes, R.; Gossler, A. Premature myogenic differentiation and depletion of progenitor cells cause severe muscle hypotrophy in Delta1 mutants. Proc. Natl. Acad. Sci. USA 2007, 104, 537–542. [Google Scholar] [CrossRef]
- Esteves de Lima, J.; Bonnin, M.A.; Birchmeier, C.; Duprez, D. Muscle contraction is required to maintain the pool of muscle progenitors via YAP and NOTCH during fetal myogenesis. eLife 2016, 5, e15593. [Google Scholar] [CrossRef]
- Yue, F.; Bi, P.; Wang, C.; Shan, T.; Nie, Y.; Ratliff, T.L.; Gavin, T.P.; Kuang, S. Pten is necessary for the quiescence and maintenance of adult muscle stem cells. Nat. Commun. 2017, 8, 14328. [Google Scholar] [CrossRef]
- Chen, J.F.; Mandel, E.M.; Thomson, J.M.; Wu, Q.; Callis, T.E.; Hammond, S.M.; Conlon, F.L.; Wang, D.Z. The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation. Nat. Genet. 2006, 38, 228–233. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, J.; Wang, J.; Zhen, H.; Liu, Y.; Li, L.; Luo, Y.; Hu, J.; Liu, X.; Li, S.; Hao, Z.; et al. MicroRNA-381 Regulates Proliferation and Differentiation of Caprine Skeletal Muscle Satellite Cells by Targeting PTEN and JAG2. Int. J. Mol. Sci. 2022, 23, 13587. https://doi.org/10.3390/ijms232113587
Shen J, Wang J, Zhen H, Liu Y, Li L, Luo Y, Hu J, Liu X, Li S, Hao Z, et al. MicroRNA-381 Regulates Proliferation and Differentiation of Caprine Skeletal Muscle Satellite Cells by Targeting PTEN and JAG2. International Journal of Molecular Sciences. 2022; 23(21):13587. https://doi.org/10.3390/ijms232113587
Chicago/Turabian StyleShen, Jiyuan, Jiqing Wang, Huimin Zhen, Yan Liu, Lu Li, Yuzhu Luo, Jiang Hu, Xiu Liu, Shaobin Li, Zhiyun Hao, and et al. 2022. "MicroRNA-381 Regulates Proliferation and Differentiation of Caprine Skeletal Muscle Satellite Cells by Targeting PTEN and JAG2" International Journal of Molecular Sciences 23, no. 21: 13587. https://doi.org/10.3390/ijms232113587
APA StyleShen, J., Wang, J., Zhen, H., Liu, Y., Li, L., Luo, Y., Hu, J., Liu, X., Li, S., Hao, Z., Li, M., & Zhao, Z. (2022). MicroRNA-381 Regulates Proliferation and Differentiation of Caprine Skeletal Muscle Satellite Cells by Targeting PTEN and JAG2. International Journal of Molecular Sciences, 23(21), 13587. https://doi.org/10.3390/ijms232113587