
Short-Term Gaseous Treatments Improve Rachis Browning in Red and White Table Grapes Stored at Low Temperature: Molecular Mechanisms Underlying Its Beneficial Effect

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Material and Methods
2.1. Plant Material
2.2. Determination of Lipid Peroxidation
2.3. Analysis of Total Phenols Content by Folin–Ciocalteu Method
2.4. Antioxidant Activities Measured via 2,2-Azino-Bis-3-Ethylbenzothiazoline-6-Sulfonic Acid (ABTS) and Ferric Reducing Antioxidant Power (FRAP) Methods
2.5. Relative Gene Expression via Quantitative Real-Time RT-PCR (RT-qPCR)
2.6. Statistical Analyses
3. Results
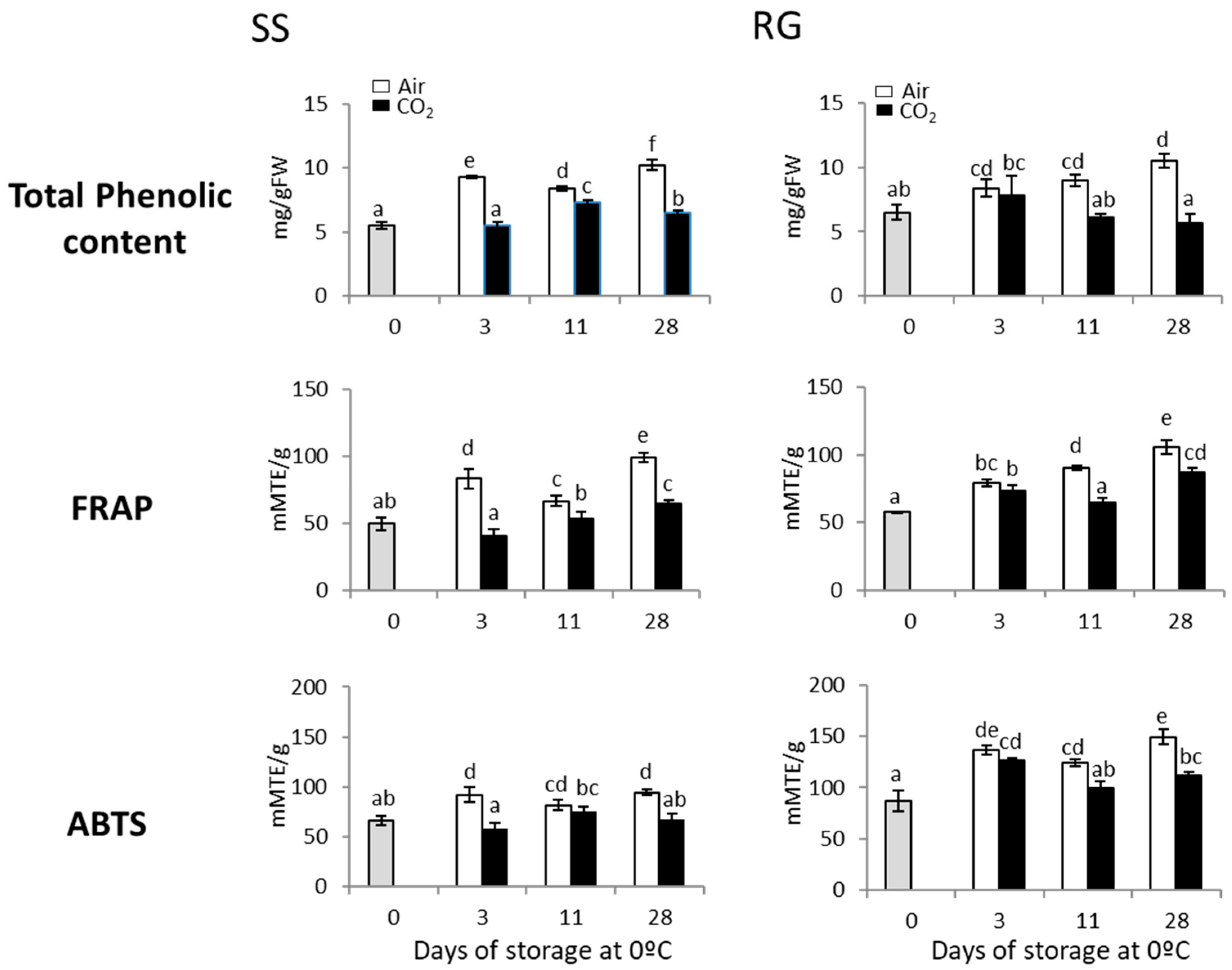
3.1. Effect of a 3-Day CO2 Treatment on Total Phenolic Content and Antioxidant Capacity in the Rachis of SS and RG Table Grapes Stored at 0 °C
3.2. Effect of a 3-Day CO2 Treatment on the Expression of VviSTSs in the Rachis of SS and RG Table Grapes Stored at 0 °C
3.3. Effect of a 3-Day CO2 Treatment on the Oxidative Stress in the Rachis of SS and RG Table Grapes Stored at 0 °C
3.4. Effect of a 3-Day CO2 Treatment on Ethylene and ABA Biosynthesis Gene Expression in the Rachis of SS and RG Table Grapes Stored at 0 °C
3.5. Effect of a 3-Day CO2 Treatment on VviERFs Expression in the Rachis of SS and RG Table Grapes Stored at 0 °C
3.6. Effect of a 3-Day CO2 Treatment on Genes Codifying Cell Wall-Degrading Enzymes in the Rachis of SS and RG Table Grapes Stored at 0 °C
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, L.; Kaplunov, T.; Zutahy, Y.; Daus, A.; Porat, R.; Lichter, A. The effects of 1-methylcyclopropane and ethylene on postharvest rachis browning in table grapes. Postharvest Biol. Technol. 2015, 107, 16–22. [Google Scholar] [CrossRef]
- Crisosto, C.H.; Smilanick, J.L.; Dokoozlian, N.K. Table grapes suffer water loss, stem browning during cooling delays. Calif. Agric. 2001, 55, 39–42. [Google Scholar] [CrossRef]
- Rosales, R.; Fernandez-Caballero, C.; Romero, I.; Escribano, M.I.; Merodio, C.; Sanchez-Ballesta, M.T. Molecular analysis of the improvement in rachis quality by high CO2 levels in table grapes stored at low temperature. Postharvest Biol. Technol. 2013, 77, 50–58. [Google Scholar] [CrossRef]
- Romanazzi, G.; Lichter, A.; Gabler, F.M.; Smilanick, J.L. Recent advances on the use of natural and safe alternatives to conventional methods to control postharvest gray mold of table grapes. Postharvest Biol. Technol. 2012, 63, 141–147. [Google Scholar] [CrossRef]
- Schanes, K.; Giljum, S.; Hertwich, E. Low carbon lifestyles: A framework to structure consumption strategies and options to reduce carbon footprints. J. Clean. Prod. 2016, 139, 1033–1043. [Google Scholar] [CrossRef]
- Sanchez-Ballesta, M.T.; Jiménez, J.B.; Romero, I.; Orea, J.M.; Maldonado, R.; Ureña, Á.G.; Escribano, M.I.; Merodio, C. Effect of high CO2 pretreatment on quality, fungal decay and molecular regulation of stilbene phytoalexin biosynthesis in stored table grapes. Postharvest Biol. Technol. 2006, 42, 209–216. [Google Scholar] [CrossRef]
- Vazquez-Hernandez, M.; Romero, I.; Escribano, M.I.; Merodio, C.; Sanchez-Ballesta, M.T. Deciphering the role of CBF/DREB transcription factors and dehydrins in maintaining the quality of table grapes cv. Autumn Royal treated with high CO2 levels and stored at 0 °C. Front. Plant Sci. 2017, 8, 1591. [Google Scholar] [CrossRef]
- Romero, I.; Domínguez, I.; Doménech-Carbó, A.; Gavara, R.; Escribano, M.I.; Merodio, C.; Sanchez-Ballesta, M.T. Effect of high levels of CO2 on the electrochemical behavior and the enzymatic and non-enzymatic antioxidant systems in black and white table grapes stored at 0 °C. J. Sci. Food Agric. 2019, 99, 6859–6867. [Google Scholar] [CrossRef]
- Sanchez-Ballesta, M.T.; Alvarez, I.; Escribano, M.I.; Merodio, C.; Romero, I. Effect of high CO2 levels and low temperature on stilbene biosynthesis pathway gene expression and stilbenes production in white, red and black table grape cultivars during postharvest storage. Plant Physiol. Biochem. 2020, 151, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Suehiro, Y.; Mochida, K.; Tsuma, M.; Yasuda, Y.; Itamura, H.; Esumi, T. Effects of abscisic acid/ethephon treatments on berry development and maturation in the yellow-green skinned ‘Shine Muscat’ grape. Hortic. J. 2019, 88, 189–201. [Google Scholar] [CrossRef]
- Cantín, C.M.; Fidelibus, M.W.; Crisosto, C.H. Application of abscisic acid (ABA) at veraison advanced red color development and maintained postharvest quality of “Crimson Seedless” grapes. Postharvest Biol. Technol. 2007, 46, 237–241. [Google Scholar] [CrossRef]
- Ohme-Takagi, M.; Shinshi, H. Ethylene-inducible DNA binding proteins that interact with an ethylene-responsive element. Plant Cell 1995, 7, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Romero, I.; Vazquez-Hernandez, M.; Escribano, M.I.; Merodio, C.; Sanchez-Ballesta, M.T. Expression profiles and DNA-binding affinity of five ERF genes in bunches of Vitis vinifera cv. Cardinal treated with high levels of CO2 at low temperature. Front. Plant Sci. 2016, 7, 1748. [Google Scholar] [CrossRef] [PubMed]
- Balic, I.; Moreno, A.; Sanhueza, D.; Huerta, C.; Orellana, A.; Defilippi, B.G.; Campos-Vargas, R. Molecular and physiological study of postharvest rachis browning of table grape cv Red Globe. Postharvest Biol. Technol. 2012, 72, 47–56. [Google Scholar] [CrossRef]
- Romero, I.; Domínguez, I.; Morales-Diaz, N.; Escribano, M.I.; Merodio, C.; Sanchez-Ballesta, M.T. Regulation of flavonoid biosynthesis pathway by a single or dual short-term CO2 treatment in black table grapes stored at low temperature. Plant Physiol. Biochem. 2020, 156, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Dong, C.; Wei, J.; Guo, L.; Meng, Y.; Wu, B.; Chen, J. A transcriptional study of the effects of nitric oxide on rachis browning in table grapes cv. Thompson Seedless. Postharvest Biol. Technol. 2021, 175, 111471. [Google Scholar] [CrossRef]
- Tomás-Barberán, F.A.; Espín, J.C. Phenolic compounds and related enzymes as determinants of quality in fruits and vegetables. J. Sci. Food Agric. 2001, 81, 853–876. [Google Scholar] [CrossRef]
- Langcake, P.; Pryce, R.J. Oxidative dimerisation of 4-hydroxystilbenes in vitro: Production of a grapevine phytoalexin mimic. J. Chem. Soc. Chem. Commun. 1977, 7, 208–210. [Google Scholar] [CrossRef]
- Cantos, E.; Espín, J.C.; Tomás-Barberán, F.A. Postharvest stilbene-enrichment of red and white table grape varieties using UV-C irradiation pulses. J. Agric. Food Chem. 2002, 50, 6322–6329. [Google Scholar] [CrossRef]
- Cayuela, J.A.; Vázquez, A.; Pérez, A.G.; García, J.M. Control of table grapes postharvest decay by ozone treatment and resveratrol induction. Food Sci. Technol. Int. 2009, 15, 495–502. [Google Scholar] [CrossRef]
- Suehiro, Y.; Mochida, K.; Itamura, H.; Esumi, T. Skin browning and expression of PPO, STS, and CHS genes in the grape berries of “Shine Muscat”. J. Jpn. Soc. Hortic. Sci. 2014, 83, 122–132. [Google Scholar] [CrossRef]
- Singleton, V.; Rossi, J. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3-new capabilities and interfaces. Nucleic Acids Res. 2012, 40, 115. [Google Scholar] [CrossRef] [PubMed]
- Ciaffi, M.; Paolacci, A.R.; Paolocci, M.; Alicandri, E.; Bigini, V.; Badiani, M.; Muganu, M. Transcriptional regulation of stilbene synthases in grapevine germplasm differentially susceptible to downy mildew. BMC Plant Biol. 2019, 19, 404. [Google Scholar] [CrossRef] [PubMed]
- Carvajal-Millán, E.; Carvallo, T.; Orozco, J.A.; Martínez, M.A.; Tapia, I.; Guerrero, V.M.; Rascón-Chu, A.; Llamas, J.; Gardea, A.A. Polyphenol oxidase activity, color changes, and dehydration in table grape rachis during development and storage as affected by N-(2-chloro-4-pyridyl)-N-phenylurea. J. Agric. Food Chem. 2001, 49, 946–951. [Google Scholar] [CrossRef]
- Campos-Vargas, R.; Zamora, P.; Contreras, R.; Köhler, H.; Zúñiga, G.E.; Pérez-Donoso, A.; Defilippi, B.G. Cold storage effects on oxidative stress of cv. Red Globe table grape rachise. Cienc. Investig. Agrar. 2012, 39, 91–104. [Google Scholar] [CrossRef]
- Liu, E.C.; Niu, L.F.; Yi, Y.; Wang, L.M.; Ai, Y.W.; Zhao, Y.; Wang, H.X.; Min, T. Expression analysis of ERFs during storage under modified atmosphere packaging (High-concentration of CO2) of fresh-cut lotus root. HortScience 2020, 55, 216–223. [Google Scholar] [CrossRef]
- Holderbaum, D.F.; Kon, T.; Kudo, T.; Guerra, M.P. Enzymatic browning, polyphenol oxidase activity, and polyphenols in four apple cultivars: Dynamics during fruit development. HortScience 2010, 45, 1150–1154. [Google Scholar] [CrossRef]
- Tanaka, E.; Okumura, S.; Takamiya, R.; Hosaka, H.; Shimamura, Y.; Murata, M. Cinnamaldehyde inhibits enzymatic browning of cut lettuce by repressing the induction of phenylalanine ammonia-lyase without promotion of microbial growth. J. Agric. Food Chem. 2011, 59, 6705–6709. [Google Scholar] [CrossRef]
- Kaur, S.; Arora, N.K.; Gill, K.B.S.; Sharma, S.; Gill, M.I.S. Hexanal formulation reduces rachis browning and postharvest losses in table grapes cv. ‘Flame Seedless’. Sci. Hortic. 2019, 248, 265–273. [Google Scholar] [CrossRef]
- Adrian, M.; Rajaei, H.; Jeandet, P.; Veneau, J.; Bessis, R. Resveratrol oxidation in Botrytis cinerea conidia. Phytopathology 1998, 88, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Aziz, A.; Verhagen, B.; Magnin-Robert, M.; Couderchet, M.; Clément, C.; Jeandet, P.; Trotel-Aziz, P. Effectiveness of beneficial bacteria to promote systemic resistance of grapevine to gray mold as related to phytoalexin production in vineyards. Plant Soil 2016, 405, 141–153. [Google Scholar] [CrossRef]
- Xu, D.; Deng, Y.; Han, T.; Jiang, L.; Xi, P.; Wang, Q.; Jiang, Z.; Gao, L. In vitro and in vivo effectiveness of phenolic compounds for the control of postharvest gray mold of table grapes. Postharvest Biol. Technol. 2018, 139, 106–114. [Google Scholar] [CrossRef]
- Ruenroengklin, N.; Yang, B.; Lin, H.; Chen, F.; Jiang, Y. Degradation of anthocyanin from litchi fruit pericarp by H2O2 and hydroxyl radical. Food Chem. 2009, 116, 995–998. [Google Scholar] [CrossRef]
- Lin, Y.; Lin, Y.; Lin, M.; Fan, Z.; Lin, H. Influence of hydrogen peroxide on the ROS metabolism and its relationship to pulp breakdown of fresh longan during storage. Food Chem. X 2021, 12, 100159. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Lin, H.; Fan, Z.; Wang, H.; Lin, M.; Chen, Y.; Hung, Y.C.; Lin, Y. Inhibitory effect of propyl gallate on pulp breakdown of longan fruit and its relationship with ROS metabolism. Postharvest Biol. Technol. 2020, 168, 111272. [Google Scholar] [CrossRef]
- Tao, D.; Wang, J.; Zhang, L.; Jiang, Y.; Lv, M. 1-Methylcyclopropene alleviates peel browning of ‘Nanguo’ pears by regulating energy, antioxidant and lipid metabolisms after long term refrigeration. Sci. Hortic. 2019, 247, 254–263. [Google Scholar] [CrossRef]
- Liu, X.; Xiao, K.; Zhang, A.; Zhu, W.; Zhang, H.; Tan, F.; Huang, Q.; Wu, X.; Zha, D. Metabolomic Analysis, Combined with Enzymatic and Transcriptome Assays, to Reveal the Browning Resistance Mechanism of Fresh-Cut Eggplant. Foods 2022, 11, 1174. [Google Scholar] [CrossRef]
- Omar, A.S.M.; Aboryia, M.S. Effect of calcium nanoparticles coupled with oxalic acid in minimizing rachis browning incidence and its relation to phenolic compounds in “Superior Seedless” grapes during shelf life. J. Plant Prod. 2022, 13, 63–73. [Google Scholar] [CrossRef]
- Lo’ay, A.A.; Taha, N.A.; EL-Khateeb, Y.A. Storability of ‘Thompson Seedless’ grapes: Using biopolymer coating chitosan and polyvinyl alcohol blending with salicylic acid and antioxidant enzymes activities during cold storage. Sci. Hortic. 2019, 249, 314–321. [Google Scholar] [CrossRef]
- Mathooko, F.M. Regulation of ethylene biosynthesis in higher plants by carbon dioxide. Postharvest Biol. Technol. 1996, 7, 1–26. [Google Scholar] [CrossRef]
- Becatti, E.; Chkaiban, L.; Tonutti, P.; Forcato, C.; Bonghi, C.; Ranier, A.M. Short-term postharvest carbon dioxide treatments induce selective molecular and metabolic changes in grape berries. J. Agric. Food Chem. 2010, 58, 8012–8020. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Zheng, X.; Zhai, D.; Song, W.; Tan, B.; Li, J.; Feng, J. Expression patterns of ACS and ACO gene families and ethylene production in rachis and berry of grapes. HortScience 2017, 52, 413–422. [Google Scholar] [CrossRef]
- Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AP2/ERF family transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta-Gene Regul. Mech. 2012, 1819, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Abiri, R.; Shaharuddin, N.A.; Maziah, M.; Yusof, Z.N.B.; Atabaki, N.; Sahebi, M.; Valdiani, A.; Kalhori, N.; Azizi, P.; Hanafi, M.M. Role of ethylene and the APETALA 2/ethylene response factor superfamily in rice under various abiotic and biotic stress conditions. Environ. Exp. Bot. 2017, 134, 33–44. [Google Scholar] [CrossRef]
- Kuang, J.F.; Chen, J.Y.; Luo, M.; Wu, K.Q.; Sun, W.; Jiang, Y.M.; Lu, W.J. Histone deacetylase HD2 interacts with ERF1 and is involved in longan fruit senescence. J. Exp. Bot. 2012, 63, 441–454. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.L.; Yin, X.R.; Chen, K.S. Roles of APETALA2/ethylene-response factors in regulation of fruit quality. CRC. Crit. Rev. Plant Sci. 2016, 35, 120–130. [Google Scholar] [CrossRef]
- Yin, X.; Allan, A.C.; Xu, Q.; Burdon, J.; Dejnoprat, S.; Chen, K.; Ferguson, I.B. Differential expression of kiwifruit ERF genes in response to postharvest abiotic stress. Postharvest Biol. Technol. 2012, 66, 1–7. [Google Scholar] [CrossRef]
- Min, T.; Fang, F.; Ge, H.; Shi, Y.N.; Luo, Z.R.; Yao, Y.C.; Grierson, D.; Yin, X.R.; Chen, K.S. Two novel anoxia-induced ethylene response factors that interact with promoters of deastringency-related genes from persimmon. PLoS ONE 2014, 9, e97043. [Google Scholar] [CrossRef]
- Min, T.; Liu, E.C.; Xie, J.; Yi, Y.; Wang, L.M.; Ai, Y.W.; Wang, H.-X. Effects of vacuum packaging on enzymatic browning and ethylene response factor (ERF) gene expression of fresh-cut lotus root. HortScience 2019, 54, 331–336. [Google Scholar] [CrossRef]
- Brummell, D.A. Cell wall disassembly in ripening fruit. Funct. Plant Biol. 2006, 33, 103–119. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wu, Z.; Zhu, Z.; Xu, L.; Wu, B.; Li, J. Botrytis cinerea mediated cell wall degradation accelerates spike stalk browning in Munage grape. J. Food Biochem. 2022, 46, 14271. [Google Scholar] [CrossRef] [PubMed]
- Lo’ay, A.A.; Taha, N.A. Evaluation rachis browning phenomena of ‘Superior Seedless’ vines grafted on different rootstocks during shelf life. Sci. Hortic. 2020, 261, 109040. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romero, I.; Rosales, R.; Escribano, M.I.; Merodio, C.; Sanchez-Ballesta, M.T. Short-Term Gaseous Treatments Improve Rachis Browning in Red and White Table Grapes Stored at Low Temperature: Molecular Mechanisms Underlying Its Beneficial Effect. Int. J. Mol. Sci. 2022, 23, 13304. https://doi.org/10.3390/ijms232113304
Romero I, Rosales R, Escribano MI, Merodio C, Sanchez-Ballesta MT. Short-Term Gaseous Treatments Improve Rachis Browning in Red and White Table Grapes Stored at Low Temperature: Molecular Mechanisms Underlying Its Beneficial Effect. International Journal of Molecular Sciences. 2022; 23(21):13304. https://doi.org/10.3390/ijms232113304
Chicago/Turabian StyleRomero, Irene, Raquel Rosales, M. Isabel Escribano, Carmen Merodio, and M. Teresa Sanchez-Ballesta. 2022. "Short-Term Gaseous Treatments Improve Rachis Browning in Red and White Table Grapes Stored at Low Temperature: Molecular Mechanisms Underlying Its Beneficial Effect" International Journal of Molecular Sciences 23, no. 21: 13304. https://doi.org/10.3390/ijms232113304
APA StyleRomero, I., Rosales, R., Escribano, M. I., Merodio, C., & Sanchez-Ballesta, M. T. (2022). Short-Term Gaseous Treatments Improve Rachis Browning in Red and White Table Grapes Stored at Low Temperature: Molecular Mechanisms Underlying Its Beneficial Effect. International Journal of Molecular Sciences, 23(21), 13304. https://doi.org/10.3390/ijms232113304