

Blockage of KHSRP-NLRP3 by MCC950 Can Reverse the Effect of Manganese-Induced Neuroinflammation in N2a Cells and Rat Brain


 , , , ,
, , , ,  , , , , , and
, , , , , and
Abstract
1. Introduction
2. Results and Discussion
2.1. ManganeseCl2 Induces Toxicity in N2a Cell Lines
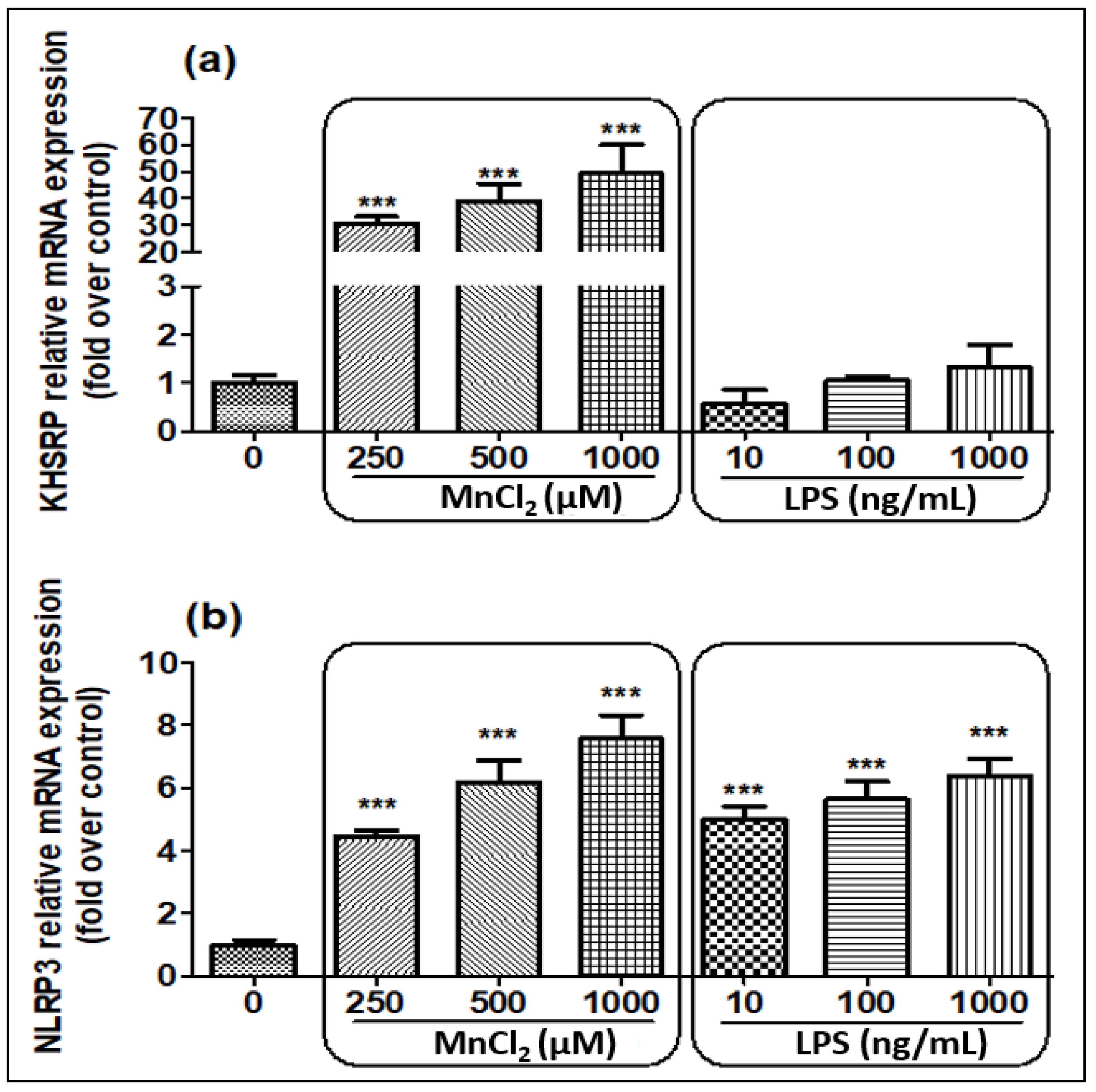
2.2. MnCl2 Treatment Induces Gene and Protein Expression of NLRP3 and KHSRP in N2a Cell Line
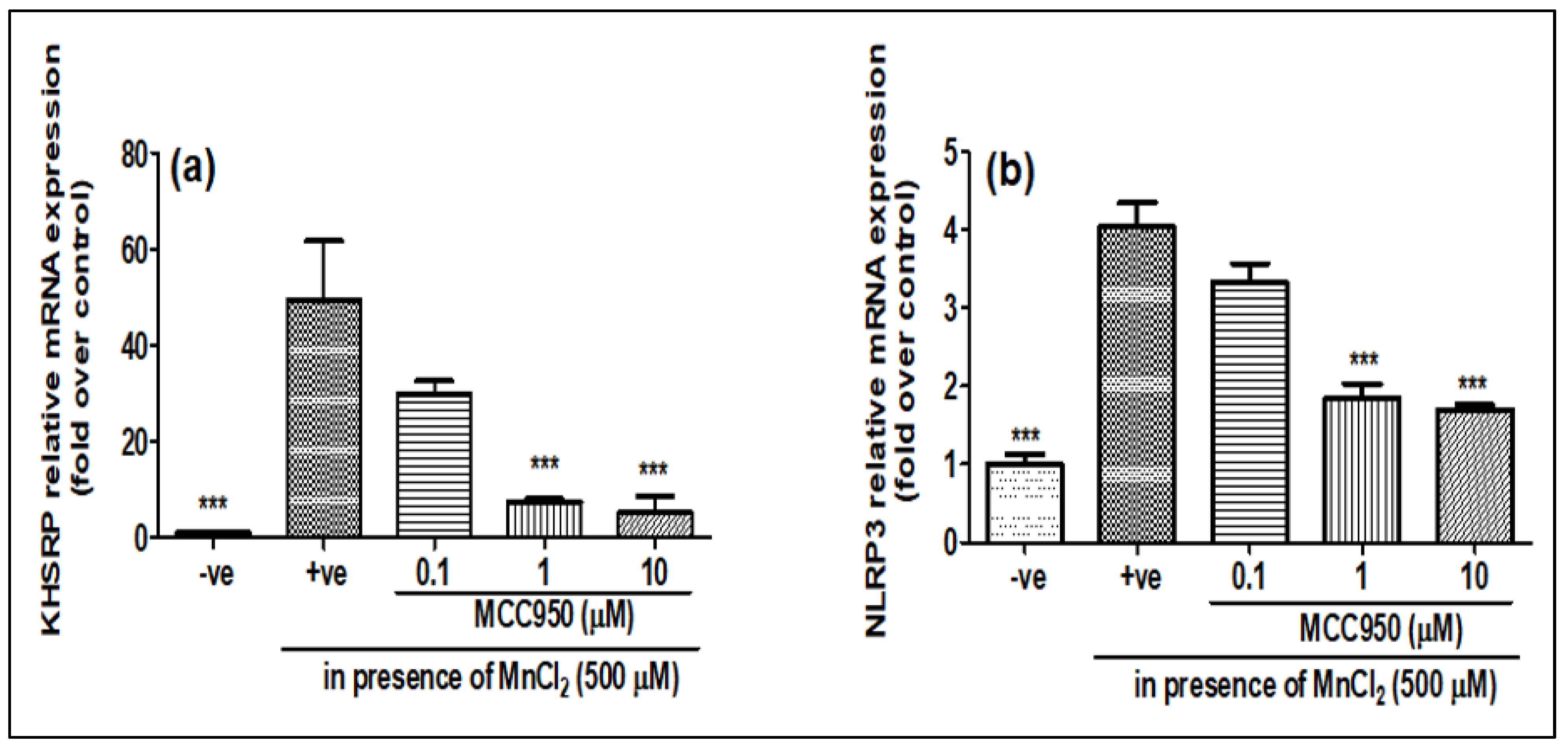
2.3. NLRP3 Inhibitor MCC950 Inhibits the Expression of KHSRP
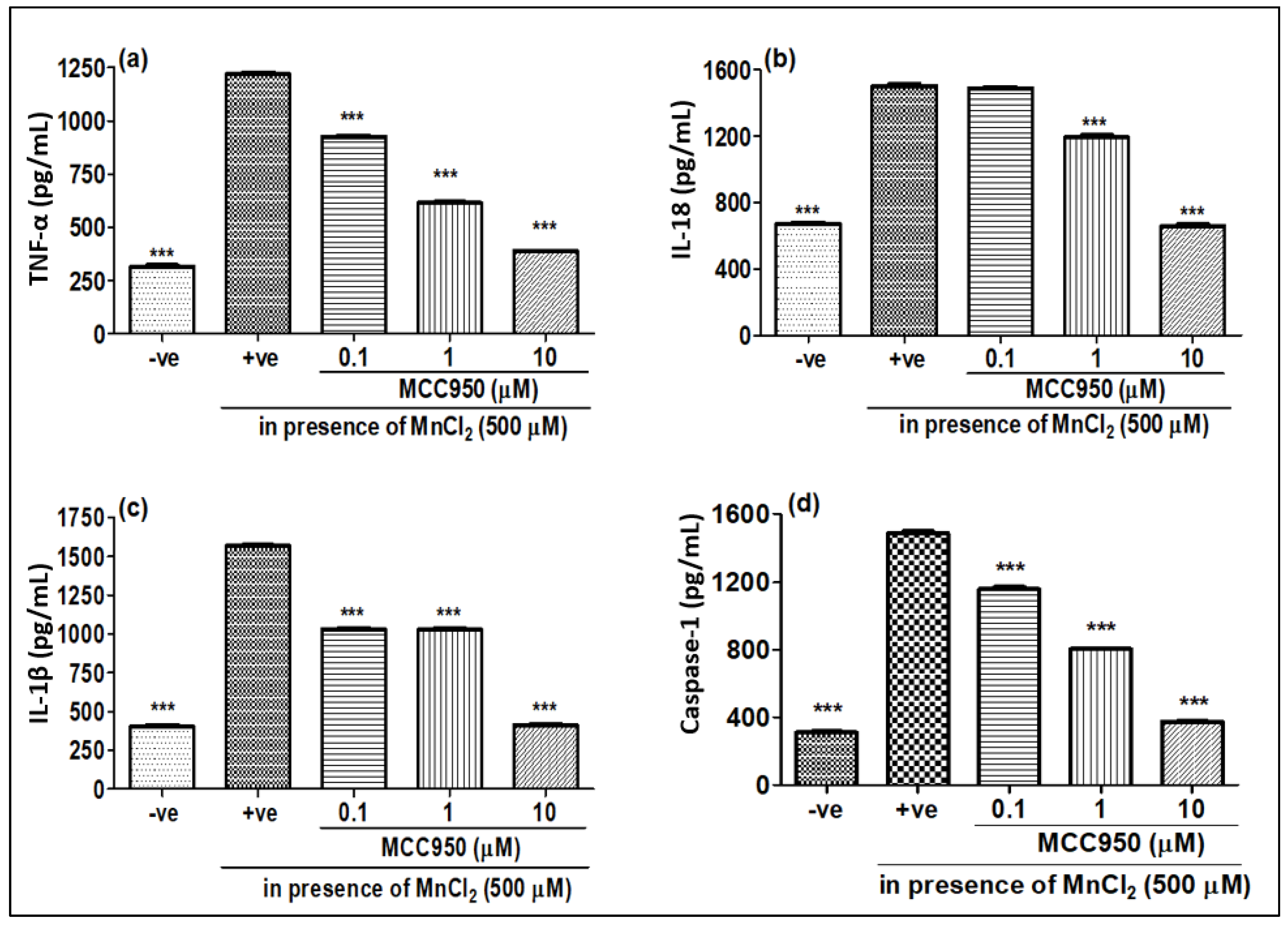
2.4. NLRP3 Inhibitor MCC950 Inhibits MnCl-2-Induced Gene Expression of Various Pro-Inflammatory Markers and Caspase-1
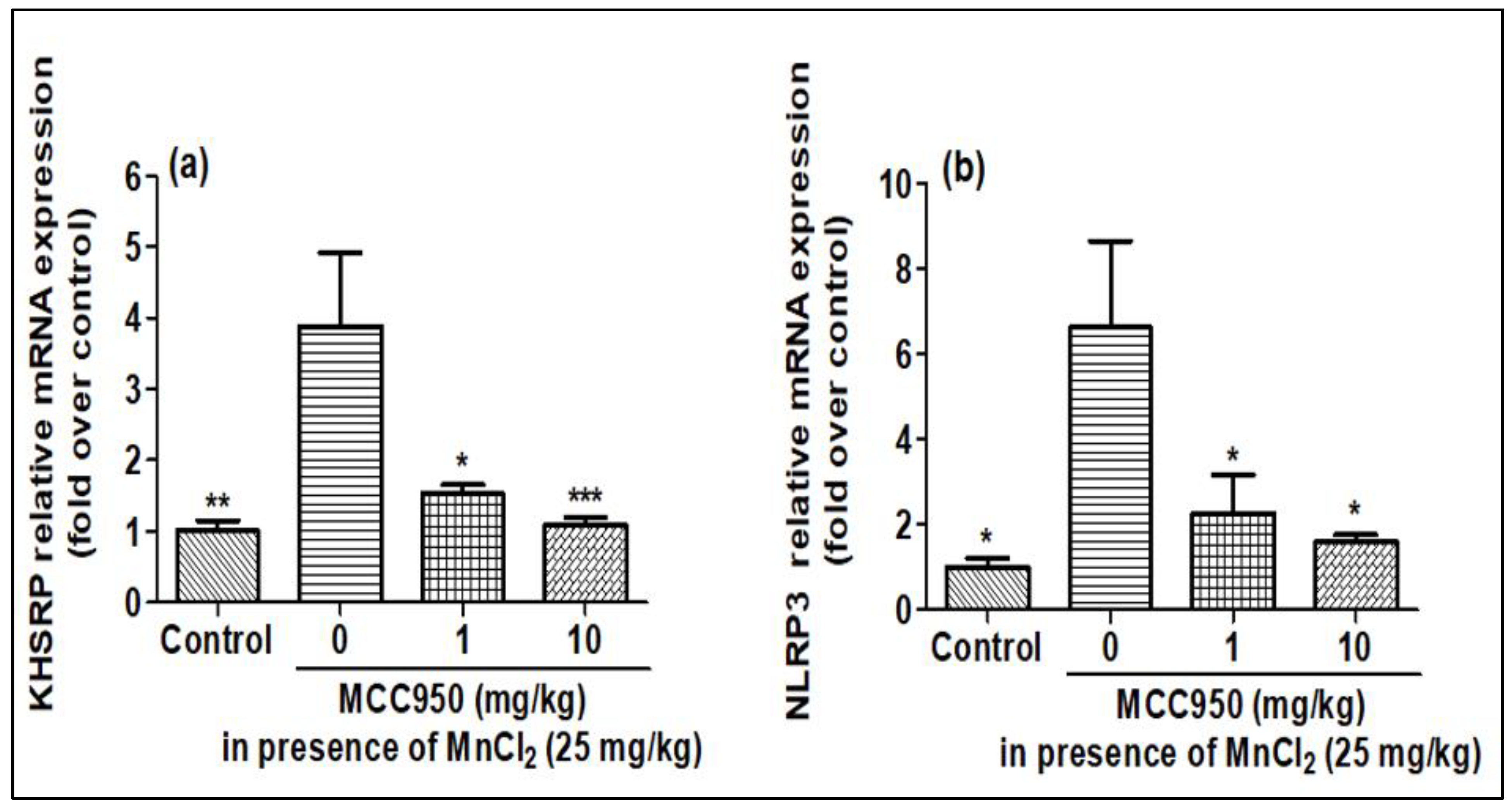
2.5. Evaluation of KHSRP and NLRP3 in Rat Brain after MnCl2 Exposure in Presence of MCC950
2.6. Effect of MnCl2 Treatment on Expression of Pro-Inflammatory Markers in Rat Brain in Presence of MCC950
3. Materials and Methods
3.1. Cell Culture
3.2. Cell Viability Assay
3.3. Cellular Experiments
3.4. Animals and Treatment
3.5. Animal Study Design and Treatment
3.6. RNA Preparation and Quantitative Real-Time PCR
3.7. Western Blotting
3.8. Measurement of TNFα, IL-1β and IL-18 Estimation
3.9. Measurement of Caspase-1
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cannon, J.R.; Greenamyre, J.T. The Role of Environmental Exposures in Neurodegeneration and Neurodegenerative Diseases. Toxicol. Sci. 2011, 124, 225–250. [Google Scholar] [CrossRef] [PubMed]
- Aschner, J.L.; Aschner, A.M. Nutritional Aspects of Manganese Homeostasis. Mol. Asp. Med. 2005, 26, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Aschner, M.; Erikson, K.M.; Dorman, D.C. Manganese Dosimetry: Species Differences and Implications for Neurotoxicity. Crit. Rev. Toxicol. 2005, 35, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Rybakowska, I.; Kaletha, K.; Anand, J.S. Manganism–Neurodegenerative Brain Disease Caused by Poisoning with Manganese. Prz. Lek. 2012, 69, 555–556. [Google Scholar]
- Ávila, D.S.; Puntel, R.L.; Folmer, V.; Rocha, J.B.T.; dos Santos, A.P.M.; Aschner, M. Manganese Neurotoxicity. In Handbook of Neurotoxicity; Kostrzewa, R.M., Ed.; Springer: New York, NY, USA, 2014; pp. 843–864. [Google Scholar]
- Gavin, C.E.; Gunter, K.K.; Gunter, T.E. Manganese and Calcium Transport in Mitochondria: Implications for Manganese Toxicity. Neurotoxicology 1999, 20, 445–453. [Google Scholar] [PubMed]
- Dorman, D.C.; Struve, F.M.; Marshall, M.W.; Parkinson, C.U.; James, R.A.; Wong, B.A. Tissue Manganese Concentrations in Young Male Rhesus Monkeys Following Subchronic Manganese Sulfate Inhalation. Toxicol. Sci. 2006, 92, 201–210. [Google Scholar] [CrossRef]
- Normandin, L.; Carrier, G.; Gardiner, P.F.; Kennedy, G.; Hazell, A.S.; Mergler, D.; Butterworth, R.F.; Philippe, S.; Zayed, J. Assessment of Bioaccumulation, Neuropathology, and Neurobehavior Following Subchronic (90 Days) Inhalation in Sprague-Dawley Rats Exposed to Manganese Phosphate. Toxicol. Appl. Pharmacol. 2002, 183, 135–145. [Google Scholar] [CrossRef]
- Roels, H.; Meiers, G.; Delos, M.; Ortega, I.; Lauwerys, R.; Buchet, J.P.; Lison, D. Influence of the Route of Administration and the Chemical Form (Mncl2, Mno2) on the Absorption and Cerebral Distribution of Manganese in Rats. Arch. Toxicol. 1997, 71, 223–230. [Google Scholar] [CrossRef]
- Bachiller, S.; Jimenez-Ferrer, I.; Paulus, A.; Yang, Y.; Swanberg, M.; Deierborg, T.; Boza-Serrano, A. Microglia in Neurological Diseases:A Road Map to Brain-Disease Dependent-Inflammatory Response. Front. Cell Neurosci. 2018, 12, 488. [Google Scholar] [CrossRef]
- Walsh, J.G.; Muruve, D.A.; Power, C. Inflammasomes in the Cns. Nat. Rev. Neurosci. 2014, 15, 84–97. [Google Scholar] [CrossRef]
- Zhao, F.; Cai, T.; Liu, M.; Zheng, G.; Luo, W.; Chen, J. Manganese Induces Dopaminergic Neurodegeneration Via Microglial Activation in a Rat Model of Manganism. Toxicol. Sci. 2009, 107, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Castejon, G.; Brough, D. Understanding the Mechanism of Il-1beta Secretion. Cytokine Growth Factor Rev. 2011, 22, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Schroder, K.; Tschopp, J. The Inflammasomes. Cell 2010, 140, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Pei, L.; Yao, S.; Wu, Y.; Shang, Y. Nlrp3 Inflammasome in Neurological Diseases, from Functions to Therapies. Front. Cell Neurosci. 2017, 11, 63. [Google Scholar] [CrossRef]
- Li, X.; Lin, W.J.; Chen, C.Y.; Si, Y.; Zhang, X.; Lu, L.; Suswam, E.; Zheng, L.; King, P.H. Ksrp: A Checkpoint for Inflammatory Cytokine Production in Astrocytes. Glia 2012, 60, 1773–1784. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Zhao, J.; Yang, L.; Nie, X.; Han, J.; Ma, X.; Wan, C.; Jiang, J. Khsrp Participates in Manganese-Induced Neurotoxicity in Rat Striatum and Pc12 Cells. J. Mol. Neurosci. 2015, 55, 454–465. [Google Scholar] [CrossRef]
- Fechir, M.; Linker, K.; Pautz, A.; Hubrich, T.; Forstermann, U.; Rodriguez-Pascual, F.; Kleinert, H. Tristetraprolin Regulates the Expression of the Human Inducible Nitric-Oxide Synthase Gene. Mol. Pharmacol. 2005, 67, 2148–2161. [Google Scholar] [CrossRef]
- Jayasooriya, R.G.; Lee, K.T.; Lee, H.J.; Choi, Y.H.; Jeong, J.W.; Kim, G.Y. Anti-Inflammatory Effects of Beta-Hydroxyisovalerylshikonin in Bv2 Microglia Are Mediated through Suppression of the Pi3k/Akt/Nf-Kb Pathway and Activation of the Nrf2/Ho-1 Pathway. Food Chem. Toxicol. 2014, 65, 82–89. [Google Scholar] [CrossRef]
- Jeong, Y.H.; Kim, Y.; Song, H.; Chung, Y.S.; Park, S.B.; Kim, H.S. Anti-Inflammatory Effects of Alpha-Galactosylceramide Analogs in Activated Microglia: Involvement of the P38 Mapk Signaling Pathway. PLoS ONE 2014, 9, e87030. [Google Scholar] [CrossRef]
- Linker, K.; Pautz, A.; Fechir, M.; Hubrich, T.; Greeve, J.; Kleinert, H. Involvement of Ksrp in the Post-Transcriptional Regulation of Human Inos Expression-Complex Interplay of Ksrp with Ttp and Hur. Nucleic Acids Res. 2005, 33, 4813–4827. [Google Scholar] [CrossRef]
- Bikkavilli, R.K.; Malbon, C.C. Dishevelled-Ksrp Complex Regulates Wnt Signaling through Post-Transcriptional Stabilization of Beta-Catenin Mrna. J. Cell Sci. 2010, 123, 1352–1362. [Google Scholar] [CrossRef] [PubMed]
- Inestrosa, N.C.; Arenas, E. Emerging Roles of Wnts in the Adult Nervous System. Nat. Rev. Neurosci. 2010, 11, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Lim, J.Y.; Lee, J.N.; Choe, S.K.; Kim, Y.I.; Song, S.R.; Cho, M.; So, H.S.; Park, R. Activation of Beta-Catenin by Inhibitors of Glycogen Synthase Kinase-3 Ameliorates Cisplatin-Induced Cytotoxicity and Pro-Inflammatory Cytokine Expression in Hei-Oc1 Cells. Toxicology 2014, 320, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.G.; Wang, R.; Khan, M.; Mahesh, V.; Brann, D.W. Role of Dickkopf-1, an Antagonist of the Wnt/Beta-Catenin Signaling Pathway, in Estrogen-Induced Neuroprotection and Attenuation of Tau Phosphorylation. J. Neurosci. 2008, 28, 8430–8441. [Google Scholar] [CrossRef]
- Peng, D.J.; Zhang, Y.W.; Li, Z.C.; Li, S.J.; Cai, M.; Qin, W.X.; Ou, S.Y.; Huang, X.W.; Yuan, Z.X.; Jiang, Y.M. Preventive Impacts of Pas-Na on the Slow Growth and Activated Inflammatory Responses in Mn-Exposed Rats. J. Trace Elem. Med. Biol. 2019, 54, 134–141. [Google Scholar] [CrossRef]
- Briata, P.; Bordo, D.; Puppo, M.; Gorlero, F.; Rossi, M.; Perrone-Bizzozero, N.; Gherzi, R. Diverse Roles of the Nucleic Acid-Binding Protein Khsrp in Cell Differentiation and Disease. Wiley Interdiscip. Rev. RNA 2016, 7, 227–240. [Google Scholar] [CrossRef]
- Bollmann, F.; Art, J.; Henke, J.; Schrick, K.; Besche, V.; Bros, M.; Li, H.; Siuda, D.; Handler, N.; Bauer, F.; et al. Resveratrol Post-Transcriptionally Regulates Pro-Inflammatory Gene Expression Via Regulation of Ksrp Rna Binding Activity. Nucleic Acids Res. 2014, 42, 12555–12569. [Google Scholar] [CrossRef]
- Santamaria, A.B.; Sulsky, S.I. Risk Assessment of an Essential Element: Manganese. J. Toxicol. Environ. Health A 2010, 73, 128–155. [Google Scholar] [CrossRef]
- Swanson, K.V.; Deng, M.; Ting, J.P. The Nlrp3 Inflammasome: Molecular Activation and Regulation to Therapeutics. Nat. Rev. Immunol. 2019, 19, 477–489. [Google Scholar] [CrossRef]
- Sarkar, S.; Rokad, D.; Malovic, E.; Luo, J.; Harischandra, D.S.; Jin, H.; Anantharam, V.; Huang, X.; Lewis, M.; Kanthasamy, A.; et al. Manganese Activates Nlrp3 Inflammasome Signaling and Propagates Exosomal Release of Asc in Microglial Cells. Sci. Signal. 2019, 12, 63. [Google Scholar] [CrossRef]
- Sriram, K.; Lin, G.X.; Jefferson, A.M.; Roberts, J.R.; Chapman, R.S.; Chen, B.T.; Soukup, J.M.; Ghio, A.J.; Antonini, J.M. Dopaminergic Neurotoxicity Following Pulmonary Exposure to Manganese-Containing Welding Fumes. Arch. Toxicol. 2010, 84, 521–540. [Google Scholar] [CrossRef] [PubMed]
- Barhoumi, R.; Faske, J.; Liu, X.; Tjalkens, R.B. Manganese Potentiates Lipopolysaccharide-Induced Expression of Nos2 in C6 Glioma Cells through Mitochondrial-Dependent Activation of Nuclear Factor Kappab. Brain Res. Mol. Brain Res. 2004, 122, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.Y.; Liu, L.Z. Manganese Potentiates Nitric Oxide Production by Microglia. Brain Res. Mol. Brain Res. 1999, 68, 22–28. [Google Scholar] [CrossRef]
- Filipov, N.M.; Seegal, R.F.; Lawrence, D.A. Manganese Potentiates in Vitro Production of Proinflammatory Cytokines and Nitric Oxide by Microglia through a Nuclear Factor Kappa B-Dependent Mechanism. Toxicol. Sci. 2005, 84, 139–148. [Google Scholar] [CrossRef]
- Liao, S.L.; Ou, Y.C.; Chen, S.Y.; Chiang, A.N.; Chen, C.J. Induction of Cyclooxygenase-2 Expression by Manganese in Cultured Astrocytes. Neurochem. Int. 2007, 50, 905–915. [Google Scholar] [CrossRef]
- Chen, C.J.; Ou, Y.C.; Lin, S.Y.; Liao, S.L.; Chen, S.Y.; Chen, J.H. Manganese Modulates Pro-Inflammatory Gene Expression in Activated Glia. Neurochem. Int. 2006, 49, 62–71. [Google Scholar] [CrossRef]
- Zhang, P.; Lokuta, K.M.; Turner, D.E.; Liu, B. Synergistic Dopaminergic Neurotoxicity of Manganese and Lipopolysaccharide: Differential Involvement of Microglia and Astroglia. J. Neurochem. 2010, 112, 434–443. [Google Scholar] [CrossRef]
- Zhao, X.; Yin, L.; Wu, Y.; Han, M.; Zhuang, Y.; Cong, Y.; Liu, Y.; Chen, G.; Jiang, J. Manganese Induces Neuroinflammation Via Nf-Κb/Ros Nlrp3 Pathway in Rat Brain Striatum and Hapi Cells. Mol. Cell. Toxicol. 2019, 15, 173–183. [Google Scholar] [CrossRef]
- Min, H.; Turck, C.W.; Nikolic, J.M.; Black, D.L. A New Regulatory Protein, Ksrp, Mediates Exon Inclusion through an Intronic Splicing Enhancer. Genes Dev. 1997, 11, 1023–1036. [Google Scholar] [CrossRef]
- Bae, J.H.; Jang, B.C.; Suh, S.I.; Ha, E.; Baik, H.H.; Kim, S.S.; Lee, M.Y.; Shin, D.H. Manganese Induces Inducible Nitric Oxide Synthase (Inos) Expression Via Activation of Both Map Kinase and Pi3k/Akt Pathways in Bv2 Microglial Cells. Neurosci. Lett. 2006, 398, 151–154. [Google Scholar] [CrossRef]
- Crittenden, P.L.; Filipov, N.M. Manganese-Induced Potentiation of in Vitro Proinflammatory Cytokine Production by Activated Microglial Cells Is Associated with Persistent Activation of P38 Mapk. Toxicol. In Vitro 2008, 22, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.A.; Sullivan, K.A.; Carbone, D.L.; Hanneman, W.H.; Tjalkens, R.B. Manganese Potentiates Nuclear Factor-Kappab-Dependent Expression of Nitric Oxide Synthase 2 in Astrocytes by Activating Soluble Guanylate Cyclase and Extracellular Responsive Kinase Signaling Pathways. J. Neurosci. Res. 2008, 86, 2028–2038. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Hatter, A.; Liu, B. Manganese Chloride Stimulates Rat Microglia to Release Hydrogen Peroxide. Toxicol. Lett. 2007, 173, 88–100. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Gherzi, R.; Ong, S.E.; Chan, E.L.; Raijmakers, R.; Pruijn, G.J.; Stoecklin, G.; Moroni, C.; Mann, M.; Karin, M. Au Binding Proteins Recruit the Exosome to Degrade Are-Containing Mrnas. Cell 2001, 107, 451–464. [Google Scholar] [CrossRef]
- Suswam, E.A.; Nabors, L.B.; Huang, Y.; Yang, X.; King, P.H. Il-1beta Induces Stabilization of Il-8 Mrna in Malignant Breast Cancer Cells Via the 3′ Untranslated Region: Involvement of Divergent Rna-Binding Factors Hur, Ksrp and Tiar. Int. J. Cancer 2005, 113, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Mou, S.Q.; Li, W.J.; Zhang, N.; Zhou, Z.Y.; Ding, W.; Bian, Z.Y.; Liao, H.H. Resveratrol Inhibits Ischemia-Induced Myocardial Senescence Signals and Nlrp3 Inflammasome Activation. Oxid. Med. Cell. Longev. 2020, 2020, 2647807. [Google Scholar] [CrossRef]
- LePage, K.T.; Dickey, R.W.; Gerwick, W.H.; Jester, E.L.; Murray, T.F. On the Use of Neuro-2a Neuroblastoma Cells Versus Intact Neurons in Primary Culture for Neurotoxicity Studies. Crit. Rev. Neurobiol. 2005, 17, 27–50. [Google Scholar] [CrossRef]
- Mokgobu, M.I.; Cholo, M.C.; Anderson, R.; Steel, H.C.; Motheo, M.P.; Hlatshwayo, T.N.; Tintinger, G.R.; Theron, A.J. Oxidative Induction of Pro-Inflammatory Cytokine Formation by Human Monocyte-Derived Macrophages Following Exposure to Manganese in Vitro. J. Immunotoxicol. 2015, 12, 98–103. [Google Scholar] [CrossRef]
- Coll, R.C.; Hill, J.R.; Day, C.J.; Zamoshnikova, A.; Boucher, D.; Massey, N.L.; Chitty, J.L.; Fraser, J.A.; Jennings, M.P.; Robertson, A.A.B.; et al. Mcc950 Directly Targets the Nlrp3 Atp-Hydrolysis Motif for Inflammasome Inhibition. Nat. Chem. Biol. 2019, 15, 556–559. [Google Scholar] [CrossRef]
- Coll, R.C.; Robertson, A.A.; Chae, J.J.; Higgins, S.C.; Munoz-Planillo, R.; Inserra, M.C.; Vetter, I.; Dungan, L.S.; Monks, B.G.; Stutz, A.; et al. A Small-Molecule Inhibitor of the Nlrp3 Inflammasome for the Treatment of Inflammatory Diseases. Nat. Med. 2015, 21, 248–255. [Google Scholar] [CrossRef]
- Nolan, T.; Hands, R.E.; Bustin, S.A. Quantification of Mrna Using Real-Time Rt-Pcr. Nat. Protoc. 2006, 1, 1559–1582. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, T.; Yang, P.C. Western Blot: Technique, Theory, and Trouble Shooting. N. Am. J. Med. Sci. 2012, 4, 429–434. [Google Scholar] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence |
|---|---|
| KHSRP Forward: 5′–3′ | TCCATCCTGCCTTAGTGGGT |
| KHSRP Reverse: 5′–3′ | TAAGCCTCTGCACCCATCG |
| GAPDH Forward: 5′–3′ | CATCACTGCCACCCAGAAGACTG |
| GAPDH Reverse: 5′–3′ | ATGCCAGTGAGCTTCCCGTTCAG |
| NLRP3 Forward: 5′–3′ | TCACAACTCGCCCAAGGAGGAA |
| NLRP3 Reverse: 5′–3′ | AAGAGACCACGGCAGAAGCTAG |
| TNFα Forward: 5′–3′ | GGTGCCTATGTCTCAGCCTCTT |
| TNFα Reverse: 5′–3′ | GCCATAGAACTGATGAGAGGGAG |
| IL-18 Forward: 5′–3′ | GACAGCCTGTGTTCGAGGATATG |
| IL-18 Reverse: 5′–3′ | TGTTCTTACAGGAGAGGGTAGAC |
| IL-1β Forward: 5′–3′ | TGGACCTTCCAGGATGAGGACA |
| IL-1β Reverse: 5′–3′ | GTTCATCTCGGAGCCTGTAGTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, S.; Shaikh, I.A.; More, S.S.; Mahnashi, M.H.; Almohaimeed, H.M.; El-Sherbiny, M.; Ghoneim, M.M.; Umar, A.; Soni, H.K.; Agrawal, H.; et al. Blockage of KHSRP-NLRP3 by MCC950 Can Reverse the Effect of Manganese-Induced Neuroinflammation in N2a Cells and Rat Brain. Int. J. Mol. Sci. 2022, 23, 13224. https://doi.org/10.3390/ijms232113224
Singh S, Shaikh IA, More SS, Mahnashi MH, Almohaimeed HM, El-Sherbiny M, Ghoneim MM, Umar A, Soni HK, Agrawal H, et al. Blockage of KHSRP-NLRP3 by MCC950 Can Reverse the Effect of Manganese-Induced Neuroinflammation in N2a Cells and Rat Brain. International Journal of Molecular Sciences. 2022; 23(21):13224. https://doi.org/10.3390/ijms232113224
Chicago/Turabian StyleSingh, Sharad, Ibrahim Ahmed Shaikh, Sunil S. More, Mater H. Mahnashi, Hailah M. Almohaimeed, Mohamed El-Sherbiny, Mohammed M. Ghoneim, Ahmad Umar, Harshit Kumar Soni, Himanshu Agrawal, and et al. 2022. "Blockage of KHSRP-NLRP3 by MCC950 Can Reverse the Effect of Manganese-Induced Neuroinflammation in N2a Cells and Rat Brain" International Journal of Molecular Sciences 23, no. 21: 13224. https://doi.org/10.3390/ijms232113224
APA StyleSingh, S., Shaikh, I. A., More, S. S., Mahnashi, M. H., Almohaimeed, H. M., El-Sherbiny, M., Ghoneim, M. M., Umar, A., Soni, H. K., Agrawal, H., Mannasaheb, B. A., Khan, A. A., Muddapur, U. M., & Iqubal, S. M. S. (2022). Blockage of KHSRP-NLRP3 by MCC950 Can Reverse the Effect of Manganese-Induced Neuroinflammation in N2a Cells and Rat Brain. International Journal of Molecular Sciences, 23(21), 13224. https://doi.org/10.3390/ijms232113224