
Identification and Functional Characterization of Acyl-ACP Thioesterases B (GhFatBs) Responsible for Palmitic Acid Accumulation in Cotton Seeds

 ,
,
Abstract
1. Introduction
2. Results
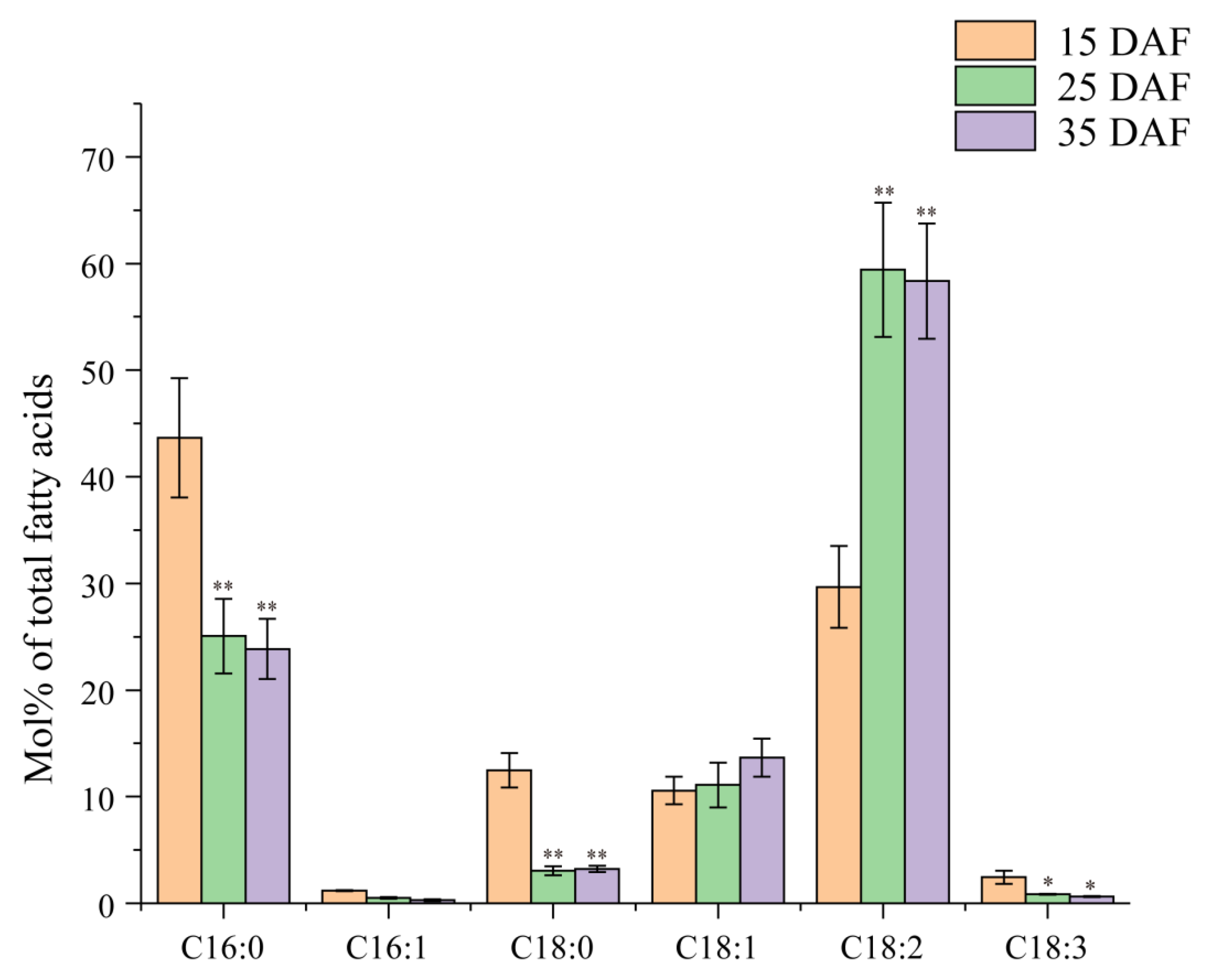
2.1. Dynamic Profiles of Fatty Acids during the Development of Cotton Seeds
2.2. Genome-Wide Identification of FatB Family Members from Four Cotton Species
2.3. Gene Structure, Conserved Domain and Phylogenetic Analysis of GhFatBs
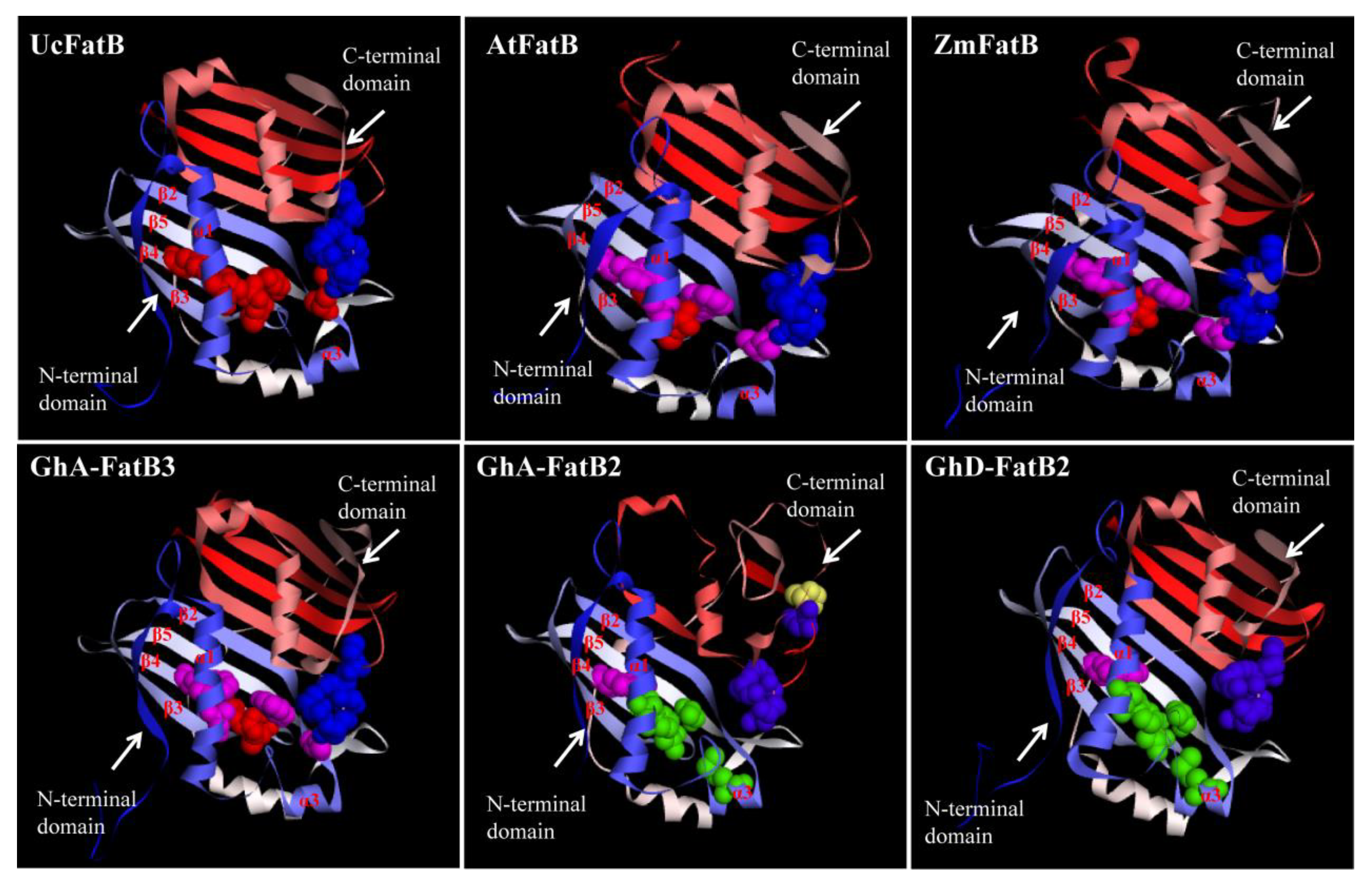
2.4. Multiple Sequence Alignments and 3D Structural Modelling of GhFatBs
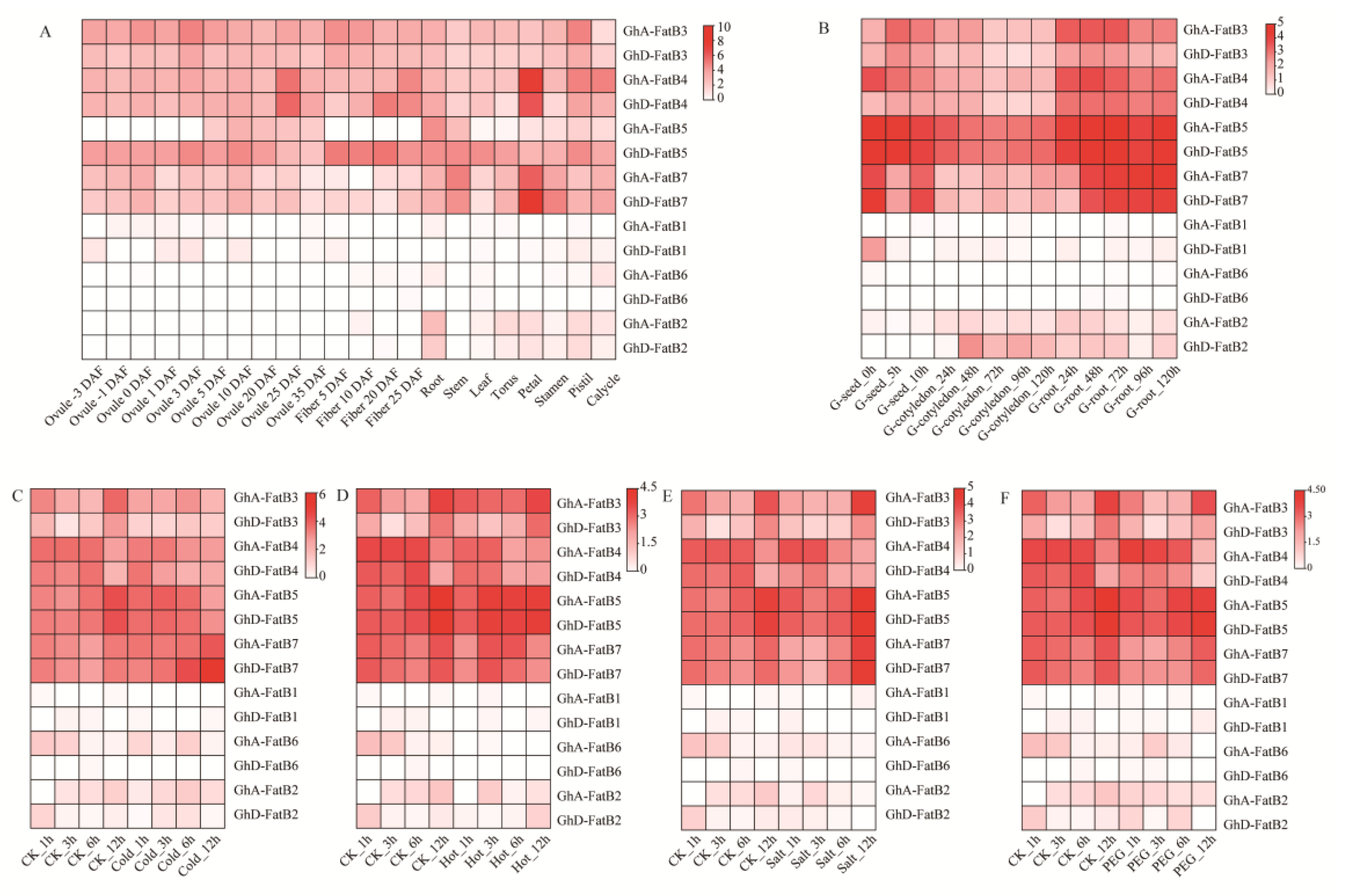
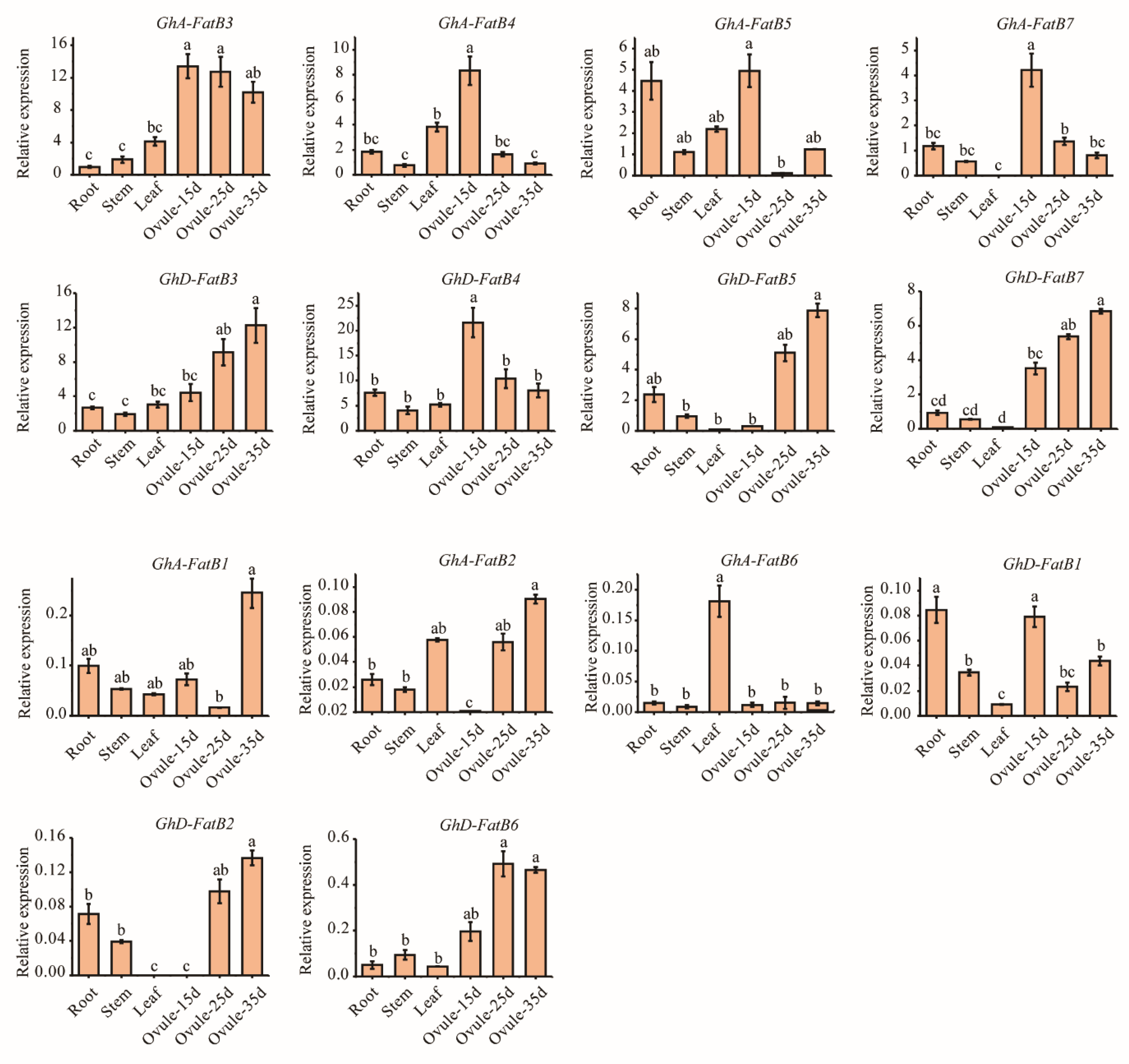
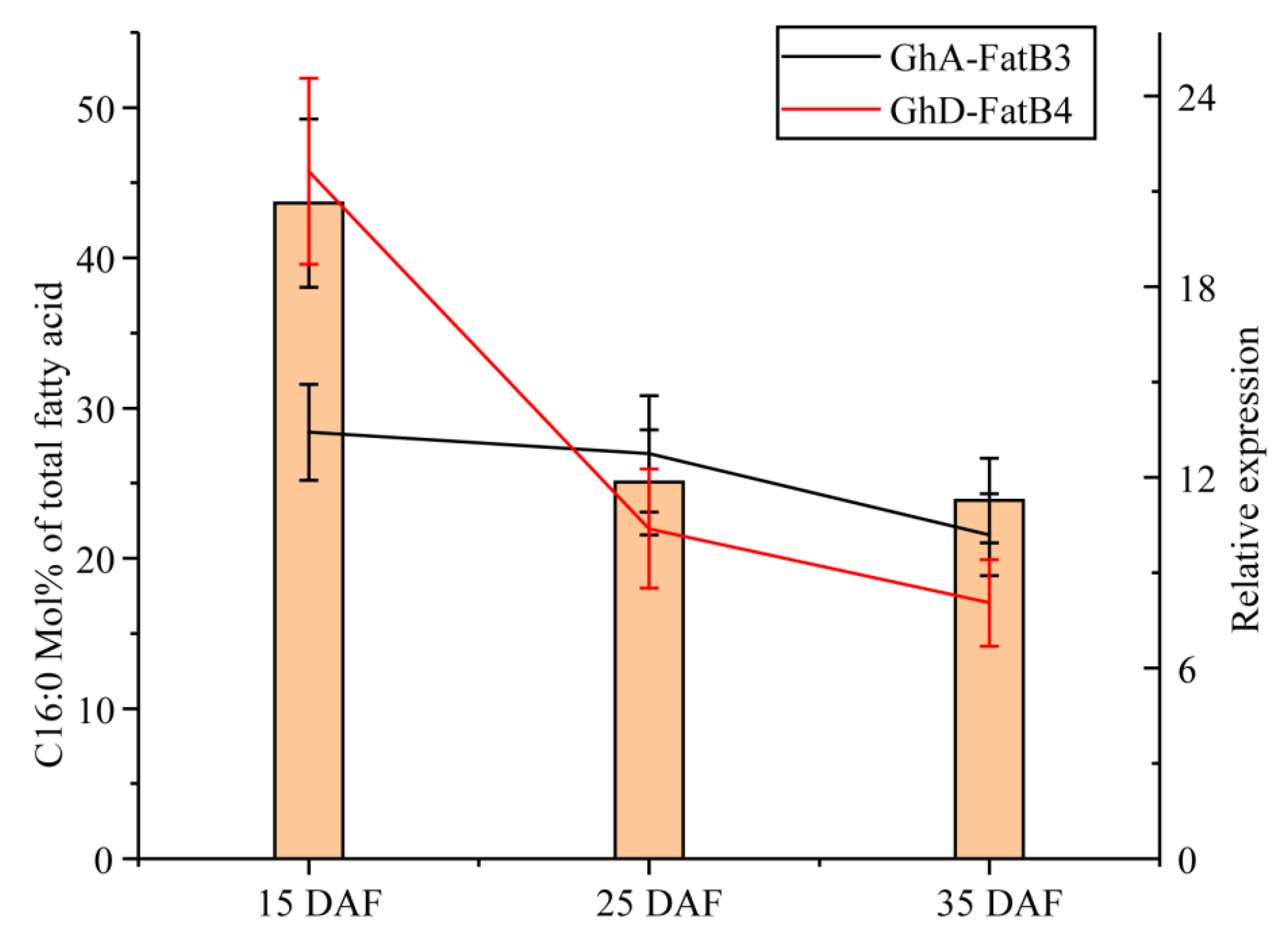
2.5. Tissue-Specific Expression Patterns of GhFatB Genes in Upland Cotton
2.6. Fatty Acid Profiles of Leaves Transiently Expressing GhFatB Genes
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Gene Isolation, Gene Structure, Functional Domain and Phylogenetic Analysis
4.3. Multiple Alignment and Three-Dimensional Model Analysis
4.4. RNA-Seq Analysis, RNA Isolation and qRT-PCR
4.5. Cloning of GhFatB Genes and Construction of Recombinant Expression Vectors
4.6. Transient Expression of GhFatB Genes in N. benthamiana
4.7. Extraction and Detection Analysis of Fatty Acid Methyl Ester
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zia, M.A.; Shah, S.H.; Shoukat, S.; Hussain, Z.; Khan, S.U.; Shafqat, N. Physicochemical features, functional characteristics, and health benefits of cottonseed oil: A review. Braz. J. Biol. 2021, 82, e243511. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Singh, S.; Green, A. High-oleic and high-stearic cottonseed oils: Nutritionally improved cooking oils developed using gene silencing. J. Am. Coll. Nutr. 2002, 21, 205S–211S. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Chen, W.; Xu, X.; Zhang, J.; Liu, Q. Engineering Trienoic Fatty Acids into Cottonseed Oil Improves Low-Temperature Seed Germination, Plant Photosynthesis and Cotton Fiber Quality. Plant Cell Physiol. 2020, 61, 1335–1347. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, Y.; Yi, H.; Cui, Y.; Hua, J. Gene network of oil accumulation reveals expression profiles in developing embryos and fatty acid composition in Upland cotton. J. Plant Physiol. 2018, 228, 101–112. [Google Scholar] [CrossRef]
- Xue, J.A.; Gao, H.L.; Gao, Y.; Hao, H.T.; Liu, B.L.; Li, R.Z. Analysis of oil composition and related gene expression of three cultivated cottonseeds. J. Shanxi Agric. Univ. Nat. Sci. Ed. 2018, 38, 1–6. [Google Scholar]
- Herbert, D.; Franz, S.; Popkova, Y.; Anderegg, U.; Schiller, J.; Schwede, K.; Lorz, A.; Simon, J.C.; Saalbach, A. High-Fat Diet Exacerbates Early Psoriatic Skin Inflammation Independent of Obesity: Saturated Fatty Acids as Key Players. J. Investig. Dermatol. 2018, 138, 1999–2009. [Google Scholar] [CrossRef]
- Hong, Y.J.; Ahn, H.J.; Shin, J.; Lee, J.H.; Lee, S.K. Unsaturated fatty acids protect trophoblast cells from saturated fatty acid-induced autophagy defects. J. Reprod. Immunol. 2017, 125, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Lakhssassi, N.; Knizia, D.; Cullen, M.A.; Meksem, K. Genome-wide identification and analysis of soybean acyl-ACP thioesterase gene family reveals the role of GmFAT to improve fatty acid composition in soybean seed. Theor. Appl. Genet. 2021, 134, 3611–3623. [Google Scholar] [CrossRef]
- Rawsthorne, S. Carbon flux and fatty acid synthesis in plants. Prog. Lipid Res. 2002, 41, 182–196. [Google Scholar] [CrossRef]
- Baud, S.; Lepiniec, L. Physiological and developmental regulation of seed oil production. Prog. Lipid Res. 2010, 49, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Byers, D.M.; Gong, H. Acyl carrier protein: Structure–function relationships in a conserved multifunctional protein family. Biochem. Cell Biol. 2007, 85, 649–662. [Google Scholar] [CrossRef]
- Knutzon, D.S. Lysophosphatidic acid acyltransferase from coconut endosperm mediates the insertion of laurate at the sn-2 position of triacylglycerols in lauric rapeseed oil and can increase total laurate levels. Plant Physiol. 1999, 120, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Dormann, P. Accumulation of Palmitate in Arabidopsis Mediated by the Acyl-Acyl Carrier Protein Thioesterase FATB1. Plant Physiol. 2000, 123, 637–644. [Google Scholar] [CrossRef]
- Jones, A. Palmitoyl-acyl carrier protein (ACP) thioesterase and the evolutionary origin of plant acyl-ACP thioesterases. Plant Cell 1995, 7, 359–371. [Google Scholar] [PubMed]
- Rodríguez-Rodríguez, M.; Salas, J.J.; Garcés, R.; Martínez-Force, E. Acyl-ACP thioesterases from Camelina sativa: Cloning, enzymatic characterization and implication in seed oil fatty acid composition. Phytochemistry 2014, 107, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Eccleston, V.S.; Cranmer, A.M.; Voelker, T.A.; Ohlrogge, J.B. Medium-chain fatty acid biosynthesis and utilization inBrassica napusplants expressing lauroyl-acyl carrier protein thioesterase. Planta 1996, 198, 46–53. [Google Scholar] [CrossRef]
- Eccleston, V.S.; Ohlrogge, J.B. Expression of Lauroyl-Acyl Carrier Protein Thioesterase in Brassica napus Seeds Induces Pathways for Both Fatty Acid Oxidation and Biosynthesis and Implies a Set Point for Triacylglycerol Accumulation. Plant Cell 1998, 10, 613–622. [Google Scholar]
- Dehesh, K.; Jones, A.; Knutzon, D.S.; Voelker, T.A. Production of high levels of 8:0 and 10:0 fatty acids in transgenic canola by overexpression of Ch FatB2, a thioesterase cDNA from Cuphea hookeriana. Plant J. Cell Mol. Biol. 1996, 9, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Bonaventure, G.; Bao, X.; Ohlrogge, J.; Pollard, M. metabolic responses to the reduction in palmitate caused by disruption of the fatb gene. Plant Physiol. 2019, 135, 1269–1279. [Google Scholar] [CrossRef][Green Version]
- Liu, Y.; Han, J.; Li, Z.J.; Jiang, Z.J.; Luo, L.F.; Zhang, Y.Z. Heterologous expression of Jatropha curcas fatty acyl-ACP thioesterase A (JcFATA) and B (JcFATB) affects fatty acid accumulation and promotes plant growth and development in Arabidopsis. Int. J. Mol. Sci. 2022, 23, 4209. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Sinha, S.; Das, N.; Maiti, M.K. Increasing the stearate content in seed oil of Brassica juncea by heterologous expression of MlFatB affects lipid content and germination frequency of transgenic seeds. Plant Physiol. Biochem. 2015, 96, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Luo, T.; Dou, Y.; Zhang, W.; Xiao, Y. Identification and Validation of Candidate Genes Involved in Fatty Acid Content in Oil Palm by Genome-Wide Association Analysis. Front. Plant Sci. 2019, 10, 1263. [Google Scholar] [CrossRef]
- Aznar-Moreno, J.A.; Sánchez, R.; Gidda, S.K.; Martínez-Force, E.; Salas, J.J. New Insights Into Sunflower (Helianthus annuus L.) FatA and FatB Thioesterases, Their Regulation, Structure and Distribution. Front. Plant Sci. 2018, 9, 1496. [Google Scholar] [CrossRef]
- Zhang, T.Z.; Hu, Y.; Jiang, W.K.; Fang, L.; Guan, X.Y.; Chen, J.D.; Zhang, J.B.; Saski, C.A. Sequencing of allotetraploid cotton (Gossypium hirsutum L. acc. TM-1) provides a resource for fiber improvement. Nat. Biotechnol. 2015, 33, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Wang, Y.; Liu, J.; Liu, Y.; Cao, X.; Xu, S. Structural Insight into Acyl-ACP Thioesterase toward Substrate Specificity Design. ACS Chem. Biol. 2017, 12, 2830–2836. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Huang, J.; Ji, H.; Pan, L.; Hu, C.; Qiu, X.; Zhu, H.; Sui, J.; Wang, J.; Qiao, L. Identification of AhFatB genes through genome-wide analysis and knockout of AhFatB reduces the content of saturated fatty acids in peanut (Arichis hypogaea L.). Plant Sci. 2022, 319, 111247. [Google Scholar] [CrossRef] [PubMed]
- Lin, N.; Ai, T.B.; Gao, J.H.; Fan, L.H.; Wang, S.H.; Chen, F. Identification of novel acyl-ACP thioesterase gene ClFATB1 from Cinnamomum longepaniculatum. Biochem. Biokhimiia 2013, 78, 1298–1303. [Google Scholar] [CrossRef]
- Kim, H.J.; Silva, J.E.; Hieu, S.V.; Mockaitis, K.; Nam, J.W.; Cahoon, E.B. Toward production of jet fuel functionality in oilseeds: Identification of FatB acyl-acyl carrier protein thioesterases and evaluation of combinatorial expression strategies in Camelina seeds. J. Exp. Bot. 2015, 66, 4251–4265. [Google Scholar] [CrossRef]
- Davies, H.M.; Anderson, L.; Fan, C.; Hawkins, D.J. Developmental induction, purification, and further characterization of 12:0-ACP thioesterase from immature cotyledons of Umbellularia californica. Arch. Biochem. Biophys. 1991, 290, 37–45. [Google Scholar] [CrossRef]
- Li, F.; Fan, G.; Lu, C.; Xiao, G.; Zou, C.; Kohel, R.J.; Yu, S. Genome sequence of cultivated Upland cotton (Gossypium hirsutum TM-1) provides insights into genome evolution. Nat. Biotechnol. 2015, 33, 524–530. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, B.; Zheng, H.J.; Hu, Y.; Lu, G.; Yang, C.Q.; Chen, X.Y. Gossypium barbadense genome sequence provides insight into the evolution of extra-long staple fiber and specialized metabolites. Sci. Rep. 2015, 5, 14139. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-García, A.; Moreno-Pérez, A.; Muro-Pastor, A.M.; Salas, J.J.; Garcés, R.; Martínez-Force, E. Acyl-ACP thioesterases from castor (Ricinus communis L.): An enzymatic system appropriate for high rates of oil synthesis and accumulation. Phytochemistry 2010, 71, 860–869. [Google Scholar] [CrossRef] [PubMed]
- Aznar-Moreno, J.A.; Venegas-Calerón, M.; Martínez-Force, E.; Garcés, R.; Salas, J.J. Acyl carrier proteins from sunflower (Helianthus annuus L.) seeds and their influence on FatA and FatB acyl-ACP thioesterase activities. Planta 2016, 244, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Ozseyhan, M.E.; Pengcheng, L.; Gunnam, N.; Zhenjing, L.; Changlu, W.; Chaofu, L. Improved fatty acid profiles in seeds of Camelina sativa by artificial microRNA mediated FATB gene suppression. Biochem. Biophys. Res. Commun. 2018, 503, 621–624. [Google Scholar] [CrossRef]
- Liu, B.; Sun, Y.; Xue, J.; Mao, X.; Jia, X.; Li, R. Stearoyl-ACP Δ(9) Desaturase 6 and 8 (GhA-SAD6 and GhD-SAD8) Are Responsible for Biosynthesis of Palmitoleic Acid Specifically in Developing Endosperm of Upland Cotton Seeds. Front Plant Sci. 2019, 10, 1–17. [Google Scholar] [CrossRef]
- Liu, F.; Zhao, Y.P.; Zhu, H.G.; Zhu, Q.H.; Sun, J. Simultaneous silencing of GhFAD2-1 and GhFATB enhances the quality of cottonseed oil with high oleic acid. J. Plant Physiol. 2017, 215, 132–139. [Google Scholar] [CrossRef]
- Anushobha, R.; Jay, S.; Kiran, K.H.; Bates, P.D. Oil-Producing Metabolons Containing DGAT1 Use Separate Substrate Pools from those Containing DGAT2 or PDAT. Plant Physiol. 2020, 184, 720–737. [Google Scholar]
- Zhao, Z.; Fan, J.; Yang, P.; Wang, Z.; Opiyo, S.O.; Mackey, D.; Xia, Y. Involvement of Arabidopsis Acyl Carrier Protein 1 in PAMP-Triggered Immunity. Mol. Plant Microbe Interact. 2022, 35, 681–693. [Google Scholar] [CrossRef]
- Jha, J.K.; Maiti, M.K.; Bhattacharjee, A.; Basu, A.; Sen, P.C.; Sen, S.K. Cloning and functional expression of an acyl-ACP thioesterase FatB type from Diploknema (Madhuca) butyracea seeds in Escherichia coli. Plant Physiol. Biochem. 2006, 44, 645–655. [Google Scholar] [CrossRef]
- Martins-Noguerol, R.; Deandrés-Gil, C.; Garcés, R.; Salas, J.J.; Martínez-Force, E.; Moreno-Pérez, A. Characterization of the acyl-ACP thioesterases from Koelreuteria paniculata reveals a new type of FatB thioesterase-ScienceDirect. Heliyon 2020, 6, e05237. [Google Scholar] [CrossRef]
- Ma, J.; Sun, S.; Whelan, J.; Shou, H.X. CRISPR/Cas9-Mediated Knockout of GmFATB1 Significantly Reduced the Amount of Saturated Fatty Acids in Soybean Seeds. Int. J. Mol. Sci. 2021, 22, 3877. [Google Scholar] [CrossRef] [PubMed]
- Iskandarov, U.; Silva, J.E.; Kim, H.J.; Andersson, M.; Cahoon, R.E.; Mockaitis, K.; Cahoon, E.B. A Specialized Diacylglycerol Acyltransferase Contributes to the Extreme Medium-Chain Fatty Acid Content of Cuphea Seed Oil. Plant Physiol. 2017, 174, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, J.; Garcia-Ruiz, H.; Bates, P.D.; Mirkov, T.E.; Wang, X. A stearoyl-acyl carrier protein desaturase, NbSACPD-C, is critical for ovule development in Nicotiana benthamiana. Plant J. 2014, 80, C502. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Babar, M.D.; Parthasarathy, S.; Gibson, R.; Parliament, K.; Flook, J.; Patterson, T.; Friedemann, P.; Kumpatla, S.; Thompson, S. A truncated FatB resulting from a single nucleotide insertion is responsible for reducing saturated fatty acids in maize seed oil. Theor. Appl. Genet. 2014, 127, 1537–1547. [Google Scholar] [CrossRef] [PubMed]
- Lindqvist, Y.; Huang, W.J.; Schneider, G.J. Crystal structure of delta(9) stearoyl-acyl carrier protein desaturase from castor seed and its relationship to other di-iron proteins. Embo J. 1996, 15, 4081–4092. [Google Scholar] [CrossRef]
- Shang, X.; Cheng, C.; Ding, J.; Guo, W. Identification of candidate genes from the SAD gene family in cotton for determination of cottonseed oil composition. Mol. Genet. Genom. 2017, 292, 1–14. [Google Scholar] [CrossRef]
- Jiang, Y.; Guo, W.; Zhu, H.; Ruan, Y.L.; Zhang, T. Overexpression of GhSusA1 increases plant biomass and improves cotton fiber yield and quality. Plant Biotechnol. J. 2012, 10, 301–312. [Google Scholar] [CrossRef]
- Liu, B.L.; Sun, Y.; Hang, W.; Wang, X.D.; Xue, J.A.; Ma, R.Y.; Jia, X.Y.; Li, R.Z. Characterization of a Novel Acyl-ACP Desaturase Gene Responsible for Palmitoleic Acid Accumulation in a Diatom Phaeodactylum tricornutum. Front. Microbiol. 2020, 11, 584589. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID. | Gene Name | Chromosome Location | Protein Length (aa) | Relative Molecular Weight (kD) | Theoretical pI | NCBI Accession | Hydrophobic Region (aa) |
|---|---|---|---|---|---|---|---|
| Gh_A03G1318 | GhA-FatB1 | A03:91082126..91083883 | 392 | 44.71 | 9.27 | XP_012479545.1 | 1~13 |
| Gh_A05G1069 | GhA-FatB2 | A05:10796095..10800280 | 420 | 48.21 | 8.65 | XP_012440572.1 | 14~18, 20~22 |
| Gh_A06G0514 | GhA-FatB3 | A06:10526649..10529358 | 418 | 46.17 | 7.65 | XP_012451088.1 | 1~14 |
| Gh_A07G0026 | GhA-FatB4 | A07:285980..287966 | 420 | 46.47 | 7.68 | XP_012448973.1 | 1~14 |
| Gh_A08G2137 | GhA-FatB5 | A08:102491658..102494334 | 413 | 45.65 | 6.42 | XP_012477171.1 | 1~14 |
| Gh_A12G1306 | GhA-FatB6 | A12:69335538..69337285 | 397 | 45.1 | 8.11 | XP_012438233.1 | 1~14 |
| Gh_A13G1750 | GhA-FatB7 | A13:76380863..76383241 | 413 | 46.01 | 8.53 | XP_012461322.1 | 1~13 |
| Gh_D02G1759 | GhD-FatB1 | D02:59968397..59970154 | 392 | 44.56 | 9.26 | XP_012479545.1 | 1~14 |
| Gh_D05G1218 | GhD-FatB2 | D05:10460475..10462206 | 392 | 45.34 | 8.78 | XP_012440572.1 | 14~22 |
| Gh_D06G0571 | GhD-FatB3 | D06:8969914..8972587 | 418 | 46.17 | 8.5 | XP_012451088.1 | 1~14 |
| Gh_D07G0034 | GhD-FatB4 | D07:343874..345855 | 420 | 46.46 | 7.07 | XP_012448973.1 | 1~14 |
| Gh_D08G2504 | GhD-FatB5 | D08:64827154..64829829 | 413 | 45.66 | 5.76 | XP_012477171.1 | 1~14 |
| Gh_D12G1429 | GhD-FatB6 | D12:44061277..44062989 | 392 | 44.72 | 8.94 | XP_012438233.1 | 1~14 |
| Gh_D13G2098 | GhD-FatB7 | D13:56456813..56459196 | 412 | 45.75 | 7.66 | XM_016864037.1 | 1~13 |
| Protein Name | C-Terminal Catalytic Center | N-Terminal Acyl Binding Pocket | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| UcFatB | D281 | N283 | H285 | E319 | C320 | T137 | S219 | M163 | M197 | R199 | F201 |
| AtFatB | D313 | N315 | H317 | E351 | C352 | A169 | W251 | I195 | M229 | R231 | W233 |
| ZmFatB | D324 | N326 | H328 | E362 | C363 | A174 | W256 | I200 | M234 | R236 | W238 |
| GhA-FatB3 | D315 | N317 | H319 | E353 | C354 | A171 | W253 | I197 | M231 | R233 | W235 |
| GhD-FatB3 | D315 | N317 | H319 | E353 | C354 | A171 | W253 | I197 | M231 | R233 | W235 |
| GhA-FatB4 | D315 | N317 | H319 | E353 | C354 | A171 | W253 | I197 | M231 | R233 | W235 |
| GhD-FatB4 | D315 | N317 | H319 | E353 | C354 | A171 | W253 | I197 | M231 | R233 | W235 |
| GhA-FatB5 | D308 | N310 | H312 | E346 | C347 | A164 | W246 | I190 | M224 | R226 | W228 |
| GhD-FatB5 | D308 | N310 | H312 | E346 | C347 | A164 | W246 | I190 | M224 | R226 | W228 |
| GhA-FatB7 | D314 | N316 | H318 | E352 | C353 | A170 | W251 | I196 | M229 | R231 | W233 |
| GhD-FatB7 | D313 | N315 | H317 | E351 | C352 | A169 | W251 | I195 | M229 | R231 | W233 |
| GhA-FatB1 | D267 | N269 | H271 | E305 | C306 | A124 | W204 | I150 | M182 | R184 | W186 |
| GhD-FatB1 | D267 | N269 | H271 | E305 | C306 | A124 | W204 | I150 | M182 | R184 | W186 |
| GhA-FatB6 | D276 | N278 | H280 | E314 | C315 | A133 | W215 | I159 | M193 | R195 | W197 |
| GhD-FatB6 | D276 | N278 | H280 | E314 | C315 | A133 | W215 | I159 | M193 | R195 | W197 |
| GhA-FatB2 | D315 | N317 | H319 | E360 | P361 | S149 | A232 | V170 | V210 | F212 | W214 |
| GhD-FatB2 | D293 | N295 | H297 | E331 | C332 | S149 | A232 | V170 | V210 | F212 | W214 |
| Second structure | Random coil | α1 helix | β5 sheets | β2 sheets | β4 sheets | β4 sheets | β4 sheets | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, B.; Sun, Y.; Wang, X.; Xue, J.; Wang, J.; Jia, X.; Li, R. Identification and Functional Characterization of Acyl-ACP Thioesterases B (GhFatBs) Responsible for Palmitic Acid Accumulation in Cotton Seeds. Int. J. Mol. Sci. 2022, 23, 12805. https://doi.org/10.3390/ijms232112805
Liu B, Sun Y, Wang X, Xue J, Wang J, Jia X, Li R. Identification and Functional Characterization of Acyl-ACP Thioesterases B (GhFatBs) Responsible for Palmitic Acid Accumulation in Cotton Seeds. International Journal of Molecular Sciences. 2022; 23(21):12805. https://doi.org/10.3390/ijms232112805
Chicago/Turabian StyleLiu, Baoling, Yan Sun, Xiaodan Wang, Jinai Xue, Jiping Wang, Xiaoyun Jia, and Runzhi Li. 2022. "Identification and Functional Characterization of Acyl-ACP Thioesterases B (GhFatBs) Responsible for Palmitic Acid Accumulation in Cotton Seeds" International Journal of Molecular Sciences 23, no. 21: 12805. https://doi.org/10.3390/ijms232112805
APA StyleLiu, B., Sun, Y., Wang, X., Xue, J., Wang, J., Jia, X., & Li, R. (2022). Identification and Functional Characterization of Acyl-ACP Thioesterases B (GhFatBs) Responsible for Palmitic Acid Accumulation in Cotton Seeds. International Journal of Molecular Sciences, 23(21), 12805. https://doi.org/10.3390/ijms232112805