
CD36+ Fibroblasts Secrete Protein Ligands That Growth-Suppress Triple-Negative Breast Cancer Cells While Elevating Adipogenic Markers for a Model of Cancer-Associated Fibroblast




Abstract
1. Introduction
2. Results
2.1. CD36+ Conditioned Medium Has Minimal Effect on Colony Formation of Non-Transformed MCF10A Cell Line
2.2. Recombinant Protein Ligands Induce Growth Suppression in Triple-Negative Breast Cancer Cell Lines
2.3. Recombinant SLIT3 Elevates the Expression of Its Receptor, ROBO2, Suggesting a Positive Feedback Loop for Tumor Suppression in TNBC Lines
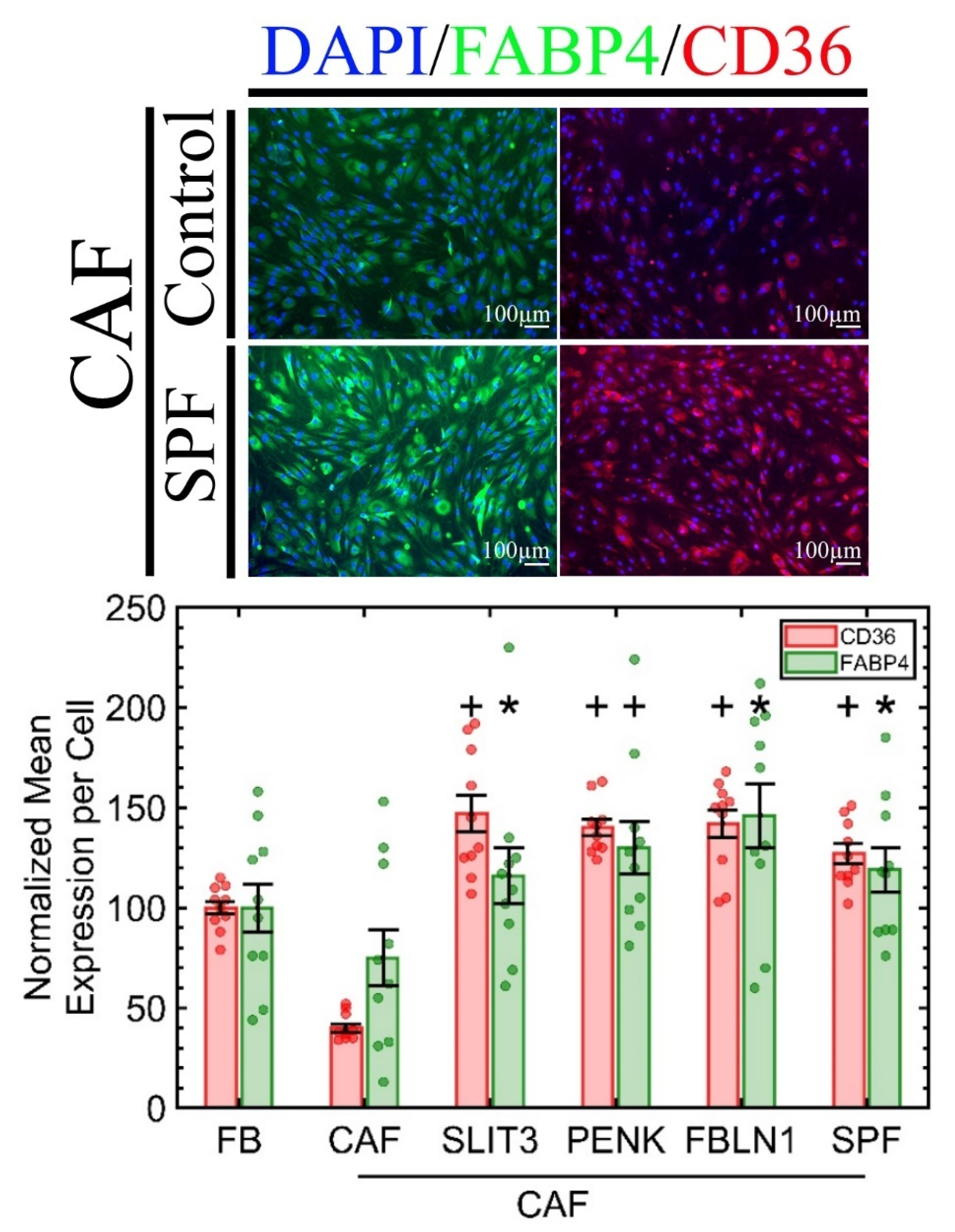
2.4. CD36 Expression in Primary CAF Is Reversible and Concomitant with the Elevation of Adipogenic Markers
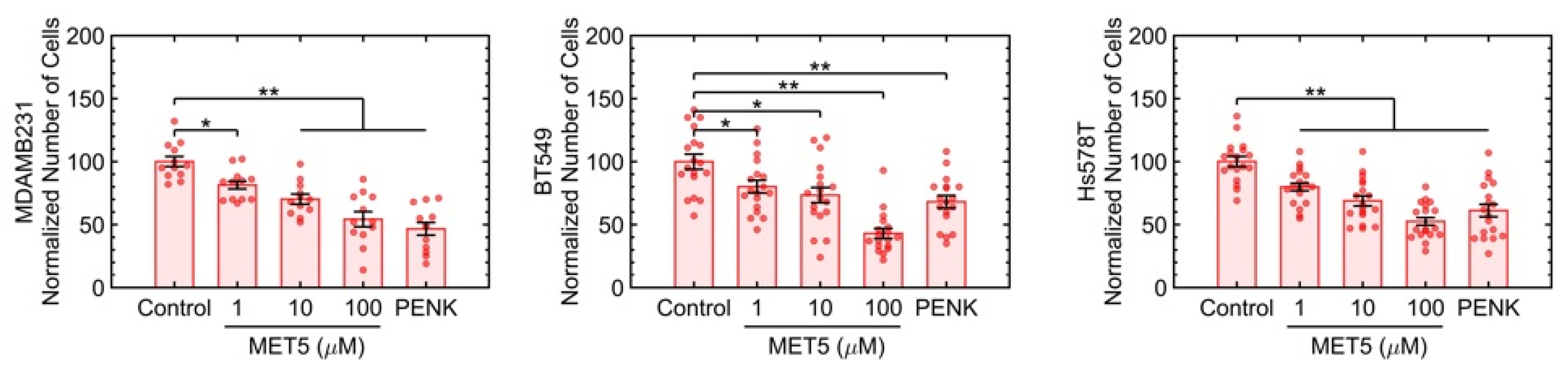
2.5. The Agonist Antibody for PENK Induces Growth Suppression in Four TNBC Lines
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cell Culture Treatment
4.3. Immunofluorescence Staining
4.4. Fluorescence Microscopy and Quantitative Analysis
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alexander, J.; Cukierman, E. Stromal dynamic reciprocity in cancer: Intricacies of fibroblastic-ECM interactions. Curr. Opin. Cell Biol. 2016, 42, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Bissell, M.J.; Hall, H.G.; Parry, G. How does the extracellular matrix direct gene expression? J. Theor. Biol. 1982, 99, 31–68. [Google Scholar] [CrossRef]
- Siemann, D. Tumor Microenvironment; Wiley-Blackwell: Hoboken, NJ, USA, 2011. [Google Scholar]
- Kalluri, R. The biology and function of fibroblasts in cancer. Nat. Rev. Cancer 2016, 16, 582–598. [Google Scholar] [CrossRef] [PubMed]
- Gascard, P.; Tlsty, T.D. Carcinoma-associated fibroblasts: Orchestrating the composition of malignancy. Genes Dev. 2016, 30, 1002–1019. [Google Scholar] [CrossRef] [PubMed]
- Peiris-Pages, M.; Smith, D.L.; Gyorffy, B.; Sotgia, F.; Lisanti, M.P. Proteomic identification of prognostic tumour biomarkers, using chemotherapy-induced cancer-associated fibroblasts. Aging 2015, 7, 816–838. [Google Scholar] [CrossRef]
- Han, C.; Liu, T.; Yin, R. Biomarkers for cancer-associated fibroblasts. Biomark. Res. 2020, 8, 64. [Google Scholar] [CrossRef]
- Bukowski, K.; Kciuk, M.; Kontek, R. Mechanisms of Multidrug Resistance in Cancer Chemotherapy. Int. J. Mol. Sci. 2020, 21, 3233. [Google Scholar] [CrossRef]
- Jabbari, K.; Winkelmaier, G.; Andersen, C.; Yaswen, P.; Quilici, D.; Furuta, S.; Cheng, Q.; Parvin, B. Protein Ligands in the Secretome of CD36+ Fibroblasts Induce Growth Suppression in a Subset of Breast Cancer Cell Lines. Cancers 2021, 13, 4521. [Google Scholar] [CrossRef]
- Cheng, Q.; Jabbari, K.; Winkelmaier, G.; Andersen, C.; Yaswen, P.; Khoshdeli, M.; Parvin, B. Overexpression of CD36 in mammary fibroblasts suppresses colony growth in breast cancer cell lines. Biochem. Biophys. Res. Commun. 2020, 526, 41–47. [Google Scholar] [CrossRef]
- Alkasalias, T.; Moyano-Galceran, L.; Arsenian-Henriksson, M.; Lehti, K. Fibroblasts in the Tumor Microenvironment: Shield or Spear? Int. J. Mol. Sci. 2018, 19, 1532. [Google Scholar] [CrossRef]
- Romer, A.M.; Luhr, I.; Klein, A.; Friedl, A.; Sebens, S.; Rosel, F.; Arnold, N.; Strauss, A.; Jonat, W.; Bauer, M. Normal mammary fibroblasts induce reversion of the malignant phenotype in human primary breast cancer. Anticancer Res. 2013, 33, 1525–1536. [Google Scholar]
- DeFilippis, R.A.; Chang, H.; Dumont, N.; Rabban, J.T.; Chen, Y.Y.; Fontenay, G.V.; Berman, H.K.; Gauthier, M.L.; Zhao, J.; Hu, D.; et al. CD36 repression activates a multicellular stromal program shared by high mammographic density and tumor tissues. Cancer Discov. 2012, 2, 826–839. [Google Scholar] [CrossRef]
- Ligorio, F.; Di Cosimo, S.; Verderio, P.; Ciniselli, C.M.; Pizzamiglio, S.; Castagnoli, L.; Dugo, M.; Galbardi, B.; Salgado, R.; Loi, S.; et al. Predictive role of CD36 expression in HER2-positive breast cancer patients receiving neoadjuvant trastuzumab. J. Natl. Cancer Inst. 2022. [Google Scholar] [CrossRef]
- Neve, R.M.; Chin, K.; Fridlyand, J.; Yeh, J.; Baehner, F.L.; Fevr, T.; Clark, L.; Bayani, N.; Coppe, J.P.; Tong, F.; et al. A collection of breast cancer cell lines for the study of functionally distinct cancer subtypes. Cancer Cell 2006, 10, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Chang, H.; Giricz, O.; Lee, G.Y.; Baehner, F.L.; Gray, J.W.; Bissell, M.J.; Kenny, P.A.; Parvin, B. Molecular predictors of 3D morphogenesis by breast cancer cell lines in 3D culture. PLoS Comput. Biol. 2010, 6, e1000684. [Google Scholar] [CrossRef] [PubMed]
- Somasundaram, V.; Hemalatha, S.K.; Pal, K.; Sinha, S.; Nair, A.S.; Mukhopadhyay, D.; Srinivas, P. Selective mode of action of plumbagin through BRCA1 deficient breast cancer stem cells. BMC Cancer 2016, 16, 336. [Google Scholar] [CrossRef] [PubMed]
- Katoh, Y.; Katoh, M. Comparative genomics on SLIT1, SLIT2, and SLIT3 orthologs. Oncol. Rep. 2005, 14, 1351–1355. [Google Scholar] [CrossRef]
- Condac, E.; Strachan, H.; Gutierrez-Sanchez, G.; Brainard, B.; Giese, C.; Heiss, C.; Johnson, D.; Azadi, P.; Bergmann, C.; Orlando, R.; et al. The C-terminal fragment of axon guidance molecule Slit3 binds heparin and neutralizes heparin’s anticoagulant activity. Glycobiology 2012, 22, 1183–1192. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kidd, T.; Brose, K.; Mitchell, K.J.; Fetter, R.D.; Tessier-Lavigne, M.; Goodman, C.S.; Tear, G. Roundabout controls axon crossing of the CNS midline and defines a novel subfamily of evolutionarily conserved guidance receptors. Cell 1998, 92, 205–215. [Google Scholar] [CrossRef]
- Nguyen Ba-Charvet, K.T.; Brose, K.; Ma, L.; Wang, K.H.; Marillat, V.; Sotelo, C.; Tessier-Lavigne, M.; Chedotal, A. Diversity and specificity of actions of Slit2 proteolytic fragments in axon guidance. J. Neurosci. 2001, 21, 4281–4289. [Google Scholar] [CrossRef]
- Tong, M.; Jun, T.; Nie, Y.; Hao, J.; Fan, D. The Role of the Slit/Robo Signaling Pathway. J. Cancer 2019, 10, 2694–2705. [Google Scholar] [CrossRef]
- Chance, R.K.; Bashaw, G.J. Slit-Dependent Endocytic Trafficking of the Robo Receptor Is Required for Son of Sevenless Recruitment and Midline Axon Repulsion. PLoS Genet. 2015, 11, e1005402. [Google Scholar] [CrossRef]
- Huang, Z.; Wen, P.; Kong, R.; Cheng, H.; Zhang, B.; Quan, C.; Bian, Z.; Chen, M.; Zhang, Z.; Chen, X.; et al. USP33 mediates Slit-Robo signaling in inhibiting colorectal cancer cell migration. Int. J. Cancer 2015, 136, 1792–1802. [Google Scholar] [CrossRef]
- Achari, A.E.; Jain, S.K. Adiponectin, a Therapeutic Target for Obesity, Diabetes, and Endothelial Dysfunction. Int. J. Mol. Sci. 2017, 18, 1321. [Google Scholar] [CrossRef]
- Gao, H.; Volat, F.; Sandhow, L.; Galitzky, J.; Nguyen, T.; Esteve, D.; Astrom, G.; Mejhert, N.; Ledoux, S.; Thalamas, C.; et al. CD36 Is a Marker of Human Adipocyte Progenitors with Pronounced Adipogenic and Triglyceride Accumulation Potential. Stem Cells 2017, 35, 1799–1814. [Google Scholar] [CrossRef]
- Krusinova, E.; Pelikanova, T. Fatty acid binding proteins in adipose tissue: A promising link between metabolic syndrome and atherosclerosis? Diabetes Res. Clin. Pract. 2008, 82, S127–S134. [Google Scholar] [CrossRef]
- Fanning, J.; Hossler, C.A.; Kesterson, J.P.; Donahue, R.N.; McLaughlin, P.J.; Zagon, I.S. Expression of the opioid growth factor-opioid growth factor receptor axis in human ovarian cancer. Gynecol. Oncol. 2012, 124, 319–324. [Google Scholar] [CrossRef]
- Zagon, I.S.; Donahue, R.; McLaughlin, P.J. Targeting the opioid growth factor: Opioid growth factor receptor axis for treatment of human ovarian cancer. Exp. Biol. Med. 2013, 238, 579–587. [Google Scholar] [CrossRef]
- Cheng, F.; McLaughlin, P.J.; Verderame, M.F.; Zagon, I.S. The OGF-OGFr axis utilizes the p21 pathway to restrict progression of human pancreatic cancer. Mol. Cancer 2008, 7, 5. [Google Scholar] [CrossRef]
- Zagon, I.S.; Porterfield, N.K.; McLaughlin, P.J. Opioid growth factor—Opioid growth factor receptor axis inhibits proliferation of triple negative breast cancer. Exp. Biol. Med. 2013, 238, 589–599. [Google Scholar] [CrossRef]
- Zagon, I.S.; Verderame, M.F.; Hankins, J.; McLaughlin, P.J. Overexpression of the opioid growth factor receptor potentiates growth inhibition in human pancreatic cancer cells. Int. J. Oncol. 2007, 30, 775–783. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gara, R.K.; Kumari, S.; Ganju, A.; Yallapu, M.M.; Jaggi, M.; Chauhan, S.C. Slit/Robo pathway: A promising therapeutic target for cancer. Drug Discov. Today 2015, 20, 156–164. [Google Scholar] [CrossRef]
- Jiang, Z.; Liang, G.; Xiao, Y.; Qin, T.; Chen, X.; Wu, E.; Ma, Q.; Wang, Z. Targeting the SLIT/ROBO pathway in tumor progression: Molecular mechanisms and therapeutic perspectives. Ther. Adv. Med. Oncol. 2019, 11, 1758835919855238. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.; Fernandis, A.Z.; Rao, Y.; Ganju, R.K. Slit protein-mediated inhibition of CXCR4-induced chemotactic and chemoinvasive signaling pathways in breast cancer cells. J. Biol. Chem. 2004, 279, 9115–9124. [Google Scholar] [CrossRef] [PubMed]
- Pupa, S.M.; Giuffre, S.; Castiglioni, F.; Bertola, L.; Cantu, M.; Bongarzone, I.; Baldassari, P.; Mortarini, R.; Argraves, W.S.; Anichini, A.; et al. Regulation of breast cancer response to chemotherapy by fibulin-1. Cancer Res. 2007, 67, 4271–4277. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.V.; Rodriguez-Manzaneque, J.C.; Thai, S.N.M.; Twal, W.O.; Luque, A.; Lyons, K.M.; Argraves, W.S.; Iruela-Arispe, M.L. Fibulin-1 acts as a cofactor for the matrix metalloprotease ADAMTS-1. J. Biol. Chem. 2005, 280, 34796–34804. [Google Scholar] [CrossRef]
- Mohamedi, Y.; Fontanil, T.; Cobo, T.; Vega, J.A.; Cobo, J.; Garcia-Suarez, O.; Cal, S.; Obaya, A.J. The molecular interaction of ADAMTS-1 and fibulin-1 and its potential contribution to breast cancer biology. J. Cancer Metastasis Treat. 2019, 5, 37. [Google Scholar] [CrossRef]
- Tang, D.F.; Lin, T.L.; Wang, Y.Y.; Cao, H. High expression of proenkephalin is associated with favorable outcomes in patients with gastrointestinal stromal tumors. Cancer Manag. Res. 2019, 11, 6681–6690. [Google Scholar] [CrossRef]
- Zhang, H.P.; Yu, Z.L.; Wu, B.B.; Sun, F.R. PENK inhibits osteosarcoma cell migration by activating the PI3K/Akt signaling pathway. J. Orthop. Surg. Res. 2020, 15, 162. [Google Scholar] [CrossRef]
- Chen, Y.J.; Liao, W.X.; Huang, S.Z.; Yu, Y.F.; Wen, J.Y.; Chen, J.; Lin, D.G.; Wu, X.Y.; Jiang, N.; Li, X. Prognostic and immunological role of CD36: A pan-cancer analysis. J. Cancer 2021, 12, 4762–4773. [Google Scholar] [CrossRef]
- Geutskens, S.B.; Hordijk, P.L.; van Hennik, P.B. The chemorepellent Slit3 promotes monocyte migration. J. Immunol. 2010, 185, 7691–7698. [Google Scholar] [CrossRef] [PubMed]
- Pupa, S.M.; Argraves, S.W.; Forti, S.; Casalini, P.; Berno, V.; Agresti, R.; Aiello, P.; Invernizzi, A.; Baldassari, P.; Otwal, W.; et al. Immunological and pathobiological roles of fibulin-1 in breast cancer. Oncogene 2004, 23, 2153–2160. [Google Scholar] [CrossRef]
- Machelska, H.; Celik, M.O. Opioid Receptors in Immune and Glial Cells-Implications for Pain Control. Front. Immunol. 2020, 11, 300. [Google Scholar] [CrossRef] [PubMed]
- Ovadia, H.; Magenheim, Y.; Behar, O.; Rosen, H. Molecular characterization of immune derived proenkephalin mRNA and the involvement of the adrenergic system in its expression in rat lymphoid cells. J. Neuroimmunol. 1996, 68, 77–83. [Google Scholar] [CrossRef]
- Plein, L.M.; Rittner, H.L. Opioids and the immune system—Friend or foe. Br. J. Pharmacol. 2018, 175, 2717–2725. [Google Scholar] [CrossRef]
- Lee, G.Y.; Kenny, P.A.; Lee, E.H.; Bissell, M.J. Three-dimensional culture models of normal and malignant breast epithelial cells. Nat. Methods 2007, 4, 359–365. [Google Scholar] [CrossRef]
- Winkelmaier, G.; Parvin, B. An enhanced loss function simplifies the deep learning model for characterizing the 3D organoid models. Bioinformatics 2021, 37, 3084–3085. [Google Scholar] [CrossRef]
- Cheng, Q.; Khoshdeli, M.; Ferguson, B.S.; Jabbari, K.; Zang, C.; Parvin, B. YY1 is a Cis-regulator in the organoid models of high mammographic density. Bioinformatics 2020, 36, 1663–1667. [Google Scholar] [CrossRef]
- Khoshdeli, M.; Winkelmaier, G.; Parvin, B. Deep Fusion of Contextual and Object-based Representations for Delineation of Multiple Nuclear Phenotypes. Bioinformatics 2019, 35, 4860–4861. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Wen, Q.; Parvin, B. Coupled Segmentation of Nuclear and Membrane-bound Macromolecules through Voting and Multiphase Level Set. Pattern Recognit. 2015, 48, 882–893. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | CD36 | ROBO2 | FABP4 |
|---|---|---|---|
| Permeabilization (Triton X-100) | 0.5% | 0.5% | 0.5% |
| Blocking Solution (BSA) | 1% | 1% | 1% |
| Primary Antibody | Novus Biological NB 400-144 | Novus Biological NBP1-81399 | R&D Systems AF3150 |
| 1:250 | 1:250 | 1:250 | |
| Secondary Antibody | Abcam Ab175471 | Abcam Ab150077 | Novus Biological NB710-58353 |
| 1:250 | 1:250 | 1:250 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jabbari, K.; Cheng, Q.; Winkelmaier, G.; Furuta, S.; Parvin, B. CD36+ Fibroblasts Secrete Protein Ligands That Growth-Suppress Triple-Negative Breast Cancer Cells While Elevating Adipogenic Markers for a Model of Cancer-Associated Fibroblast. Int. J. Mol. Sci. 2022, 23, 12744. https://doi.org/10.3390/ijms232112744
Jabbari K, Cheng Q, Winkelmaier G, Furuta S, Parvin B. CD36+ Fibroblasts Secrete Protein Ligands That Growth-Suppress Triple-Negative Breast Cancer Cells While Elevating Adipogenic Markers for a Model of Cancer-Associated Fibroblast. International Journal of Molecular Sciences. 2022; 23(21):12744. https://doi.org/10.3390/ijms232112744
Chicago/Turabian StyleJabbari, Kosar, Qingsu Cheng, Garrett Winkelmaier, Saori Furuta, and Bahram Parvin. 2022. "CD36+ Fibroblasts Secrete Protein Ligands That Growth-Suppress Triple-Negative Breast Cancer Cells While Elevating Adipogenic Markers for a Model of Cancer-Associated Fibroblast" International Journal of Molecular Sciences 23, no. 21: 12744. https://doi.org/10.3390/ijms232112744
APA StyleJabbari, K., Cheng, Q., Winkelmaier, G., Furuta, S., & Parvin, B. (2022). CD36+ Fibroblasts Secrete Protein Ligands That Growth-Suppress Triple-Negative Breast Cancer Cells While Elevating Adipogenic Markers for a Model of Cancer-Associated Fibroblast. International Journal of Molecular Sciences, 23(21), 12744. https://doi.org/10.3390/ijms232112744