

Understanding the Radiobiological Mechanisms Induced by 177Lu-DOTATATE in Comparison to External Beam Radiation Therapy
 ,
,  , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Intrinsic Radiosensitivity to 177Lu-DOTATATE and EBRT
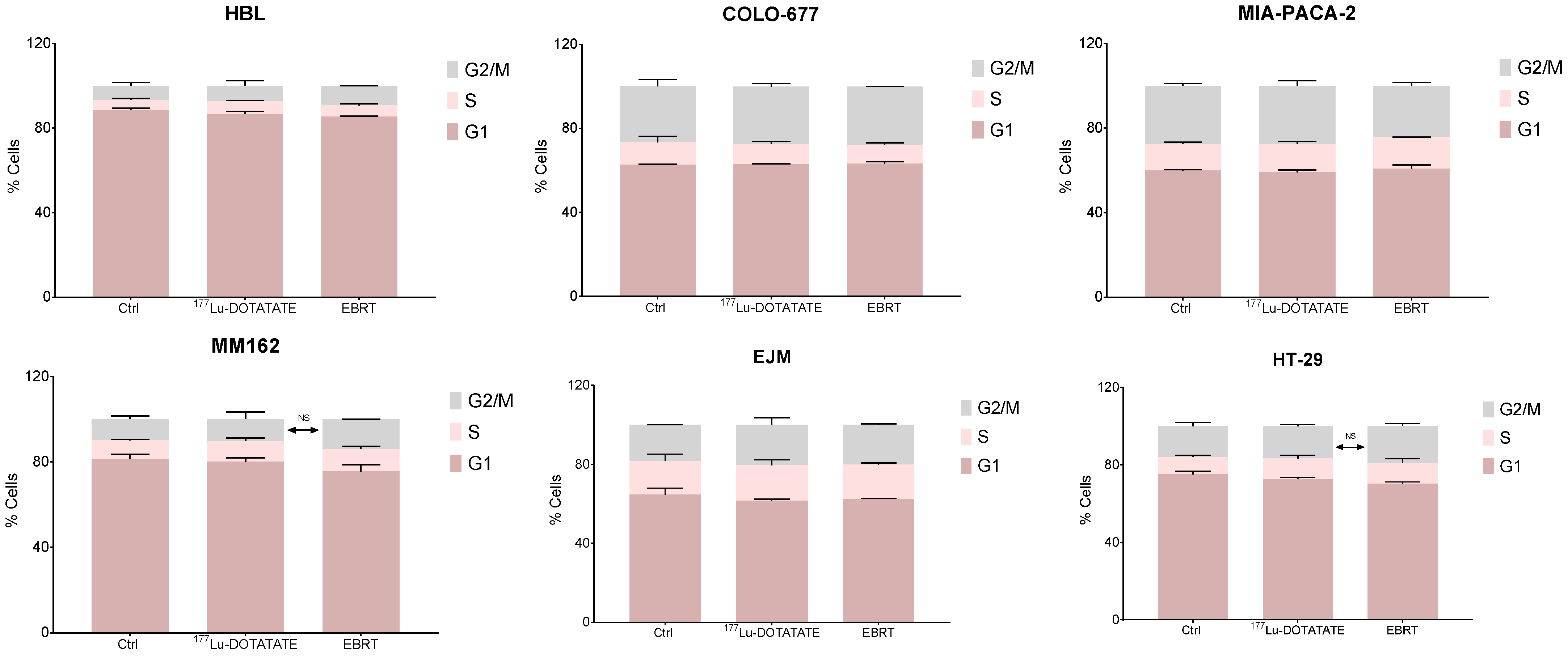
2.2. Effect of 177Lu-DOTATATE and EBRT on Cell Cycle
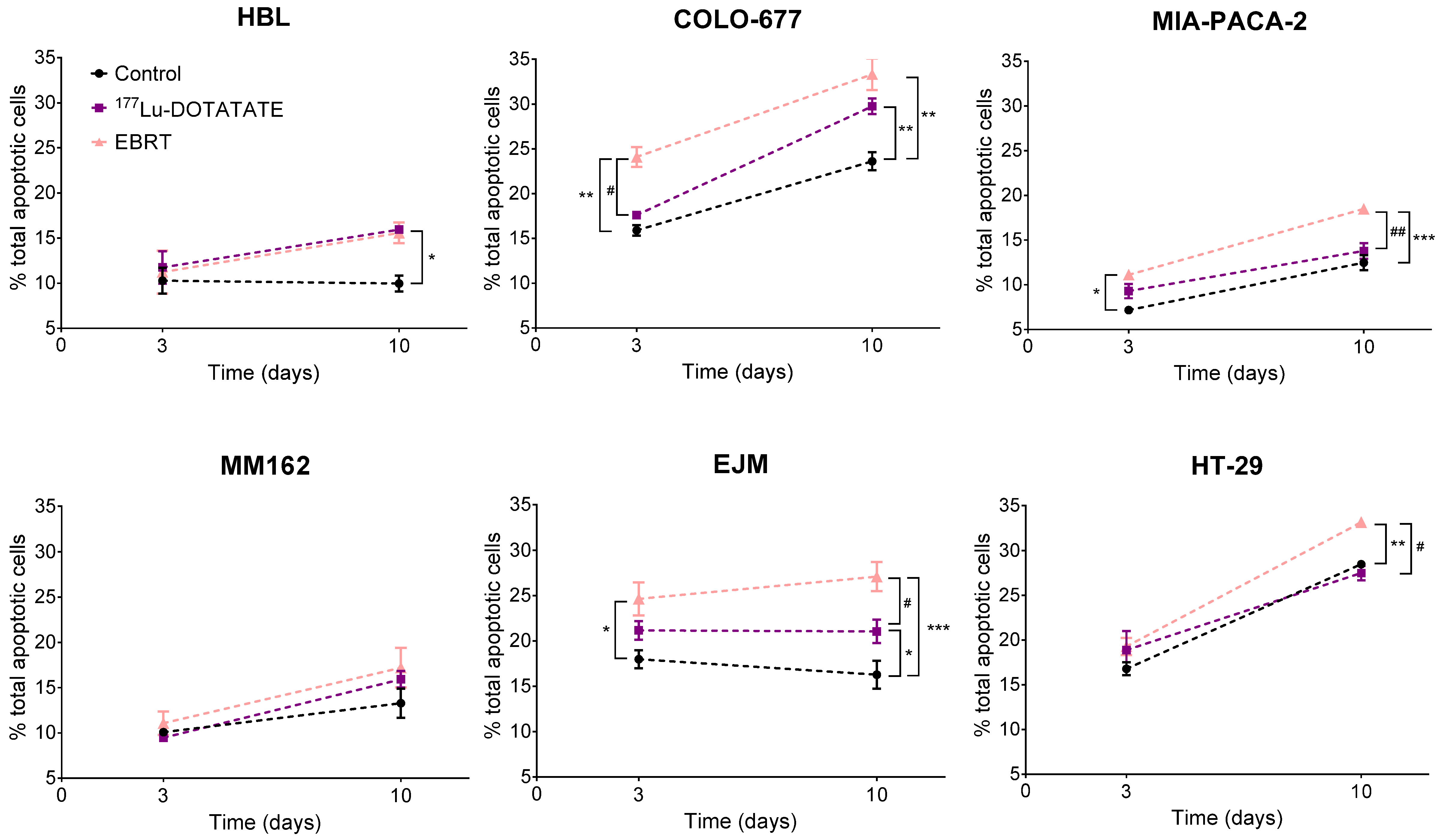
2.3. Effect of 177Lu-DOTATATE and EBRT on Apoptosis
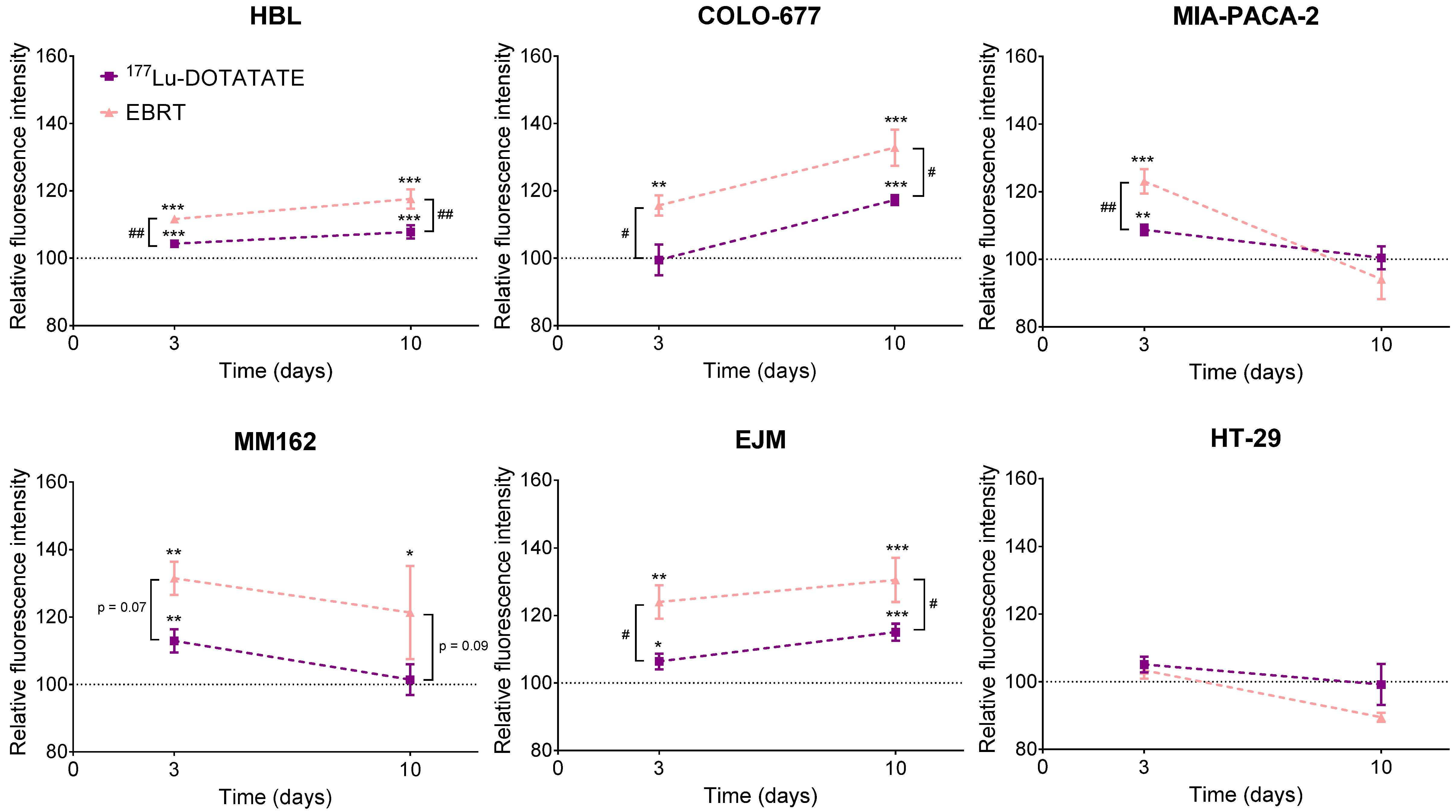
2.4. Effect of 177Lu-DOTATATE and EBRT on Autophagy
2.5. Effect of 177Lu-DOTATATE and EBRT on Reactive Oxygen Species (ROS)
2.6. Effect of 177Lu-DOTATATE and EBRT on DNA Damage
2.7. Effect of Olaparib in Combination with 177Lu-DOTATATE or EBRT on Cell Survival
2.8. Absorbed Dose Calculation of 177Lu-DOTATATE Treatment
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Cell Culture
4.2. The 177Lu-DOTATATE Production
4.3. The 177Lu-DOTATATE, EBRT and Olaparib Treatments
- 177Lu-DOTATATE: 5 MBq was added in each well, in four replicates. After 4 h of incubation at 37 °C, the medium containing 177Lu-DOTATATE was removed and replaced with fresh medium.
- EBRT: cells were irradiated at a dose of 2 Gy with a 6 MV beam from a Clinac 600 linear accelerator (Varian Medical Systems, Palo Alto, CA, USA). The collimator opening was set to 40 × 40 cm2, which gave the possibility of irradiating several plates in one batch. In order to achieve a good dose homogeneity and electronic equilibrium, a 6 mm thick polystyrene build-up was put on top of the plates. Plates were placed on a 5 cm thick polystyrene phantom for adequate backscattering conditions. The dose rate was set to 4 Gy per minute. In order to be consistent with 177Lu-DOTATATE treatment conditions, medium was replaced right after EBRT.
4.4. Cell Dosimetry
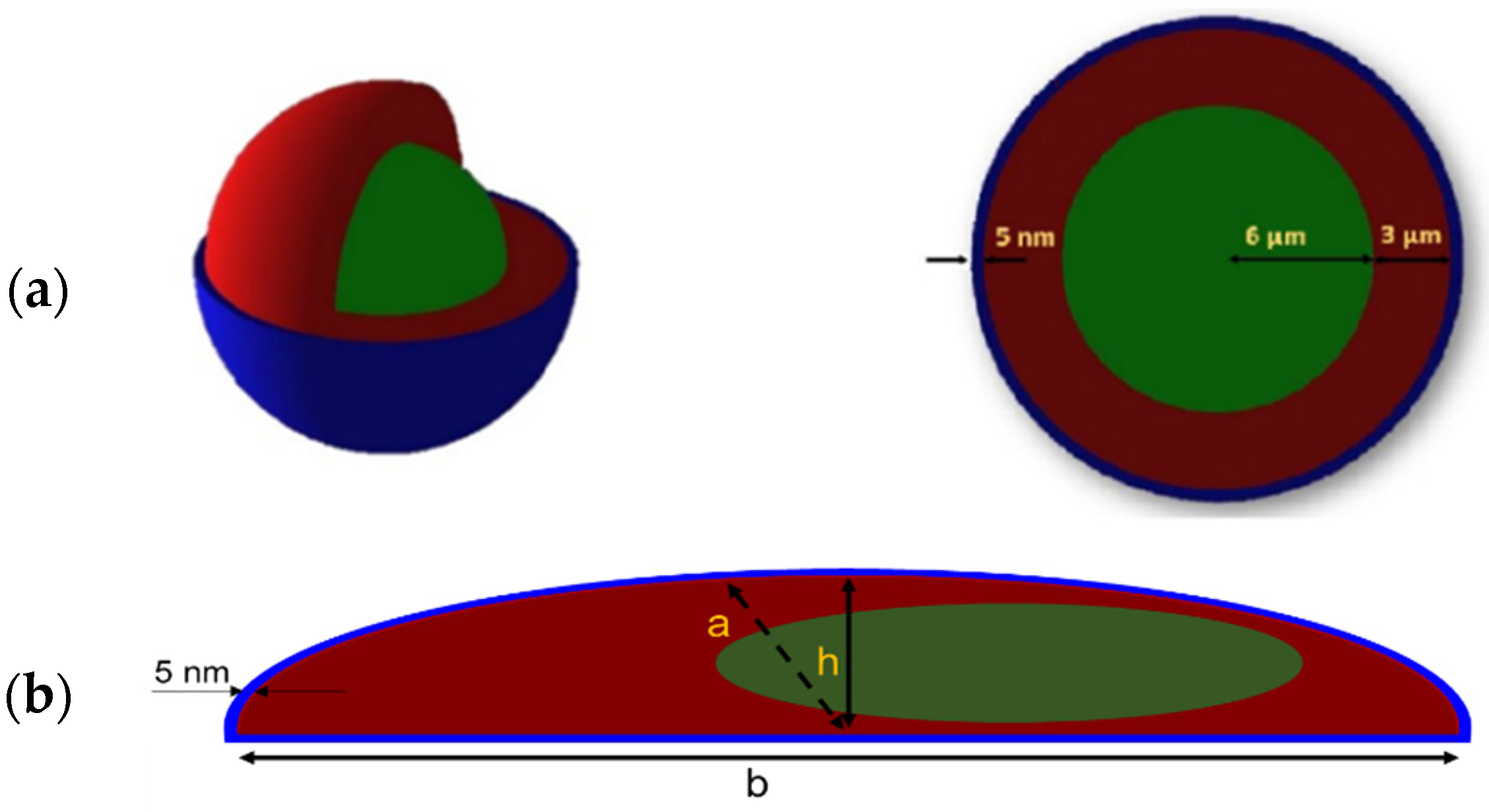
4.4.1. Geometry Set-Up
4.4.2. Radioactive Source
4.4.3. Absorbed Dose Calculation
4.5. Crystal Violet Assay
4.6. Cell Cycle Analysis
4.7. Apoptosis Analysis
4.8. Autophagy Analysis
4.9. ROS Measurement
4.10. DNA Damage Detection
4.11. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day 0 | Day 3 | Day 7 and 10 | |
|---|---|---|---|
| HBL | 80,000 | 40,000 | 1000 |
| MM162 | 50,000 | 25,000 | 2000 |
| COLO-677 | 160,000 | 80,000 | 1000 |
| EJM | 160,000 | 80,000 | 4000 |
| MIA-PACA-2 | 80,000 | 40,000 | 200 |
| HT-29 | 80,000 | 40,000 | 200 |
References
- Strosberg, J.; El-Haddad, G.; Wolin, E.; Hendifar, A.; Yao, J.; Chasen, B.; Mittra, E.; Kunz, P.L.; Kulke, M.H.; Jacene, H.; et al. Phase 3 Trial of 177Lu-Dotatate for Midgut Neuroendocrine Tumors. N. Engl. J. Med. 2017, 376, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Brabander, T.; van der Zwan, W.A.; Teunissen, J.J.M.; Kam, B.L.R.; Feelders, R.A.; de Herder, W.W.; van Eijck, C.H.J.; Franssen, G.J.H.; Krenning, E.P.; Kwekkeboom, D.J. Long-Term Efficacy, Survival, and Safety of [177Lu-DOTA0,Tyr3]octreotate in Patients with Gastroenteropancreatic and Bronchial Neuroendocrine Tumors. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2017, 23, 4617–4624. [Google Scholar] [CrossRef]
- Sistani, G.; Sutherland, D.E.K.; Mujoomdar, A.; Wiseman, D.P.; Khatami, A.; Tsvetkova, E.; Reid, R.H.; Laidley, D.T. Efficacy of 177Lu-Dotatate Induction and Maintenance Therapy of Various Types of Neuroendocrine Tumors: A Phase II Registry Study. Curr. Oncol. 2020, 28, 115–127. [Google Scholar] [CrossRef]
- Bodei, L.; Schöder, H.; Baum, R.P.; Herrmann, K.; Strosberg, J.; Caplin, M.; Öberg, K.; Modlin, I.M. Molecular profiling of neuroendocrine tumours to predict response and toxicity to peptide receptor radionuclide therapy. Lancet Oncol. 2020, 21, e431–e443. [Google Scholar] [CrossRef]
- Terry, S.Y.A.; Nonnekens, J.; Aerts, A.; Baatout, S.; de Jong, M.; Cornelissen, B.; Pouget, J.-P. Call to arms: Need for radiobiology in molecular radionuclide therapy. Eur. J. Nucl. Med. Mol. Imaging 2019, 46, 1588–1590. [Google Scholar] [CrossRef]
- Aerts, A.; Eberlein, U.; Holm, S.; Hustinx, R.; Konijnenberg, M.; Strigari, L.; van Leeuwen, F.W.B.; Glatting, G.; Lassmann, M. EANM position paper on the role of radiobiology in nuclear medicine. Eur. J. Nucl. Med. Mol. Imaging 2021, 48, 3365–3377. [Google Scholar] [CrossRef] [PubMed]
- Pouget, J.-P.; Lozza, C.; Deshayes, E.; Boudousq, V.; Navarro-Teulon, I. Introduction to radiobiology of targeted radionuclide therapy. Front. Med. 2015, 2, 12. [Google Scholar] [CrossRef] [PubMed]
- Hallqvist, A.; Svensson, J.; Hagmarker, L.; Marin, I.; Rydén, T.; Beauregard, J.-M.; Bernhardt, P. Optimizing the Schedule of PARP Inhibitors in Combination with 177Lu-DOTATATE: A Dosimetry Rationale. Biomedicines 2021, 9, 1570. [Google Scholar] [CrossRef]
- Delbart, W.; Ghanem, G.E.; Karfis, I.; Flamen, P.; Wimana, Z. Investigating intrinsic radiosensitivity biomarkers to peptide receptor radionuclide therapy with [177Lu]Lu-DOTATATE in a panel of cancer cell lines. Nucl. Med. Biol. 2021, 96–97, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Nonnekens, J.; van Kranenburg, M.; Beerens, C.E.M.T.; Suker, M.; Doukas, M.; van Eijck, C.H.J.; de Jong, M.; van Gent, D.C. Potentiation of Peptide Receptor Radionuclide Therapy by the PARP Inhibitor Olaparib. Theranostics 2016, 6, 1821–1832. [Google Scholar] [CrossRef]
- Coutard, H. Principles of X ray Therapy of Malignant Diseases. Lancet 1934, 224, 1–8. [Google Scholar] [CrossRef]
- Thariat, J.; Hannoun-Levi, J.-M.; Sun Myint, A.; Vuong, T.; Gérard, J.-P. Past, present, and future of radiotherapy for the benefit of patients. Nat. Rev. Clin. Oncol. 2013, 10, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Häggblad Sahlberg, S.; Spiegelberg, D.; Lennartsson, J.; Nygren, P.; Glimelius, B.; Stenerlöw, B. The effect of a dimeric Affibody molecule (ZEGFR:1907)2 targeting EGFR in combination with radiation in colon cancer cell lines. Int. J. Oncol. 2012, 40, 176–184. [Google Scholar] [CrossRef]
- Gholami, Y.H.; Willowson, K.P.; Forwood, N.J.; Harvie, R.; Hardcastle, N.; Bromley, R.; Ryu, H.; Yuen, S.; Howell, V.M.; Kuncic, Z.; et al. Comparison of radiobiological parameters for 90Y radionuclide therapy (RNT) and external beam radiotherapy (EBRT) in vitro. EJNMMI Phys. 2018, 5, 18. [Google Scholar] [CrossRef]
- Chavaudra, N.; Bourhis, J.; Foray, N. Quantified relationship between cellular radiosensitivity, DNA repair defects and chromatin relaxation: A study of 19 human tumour cell lines from different origin. Radiother. Oncol. 2004, 73, 373–382. [Google Scholar] [CrossRef]
- Jiang, X.; Li, X.; Li, W.; Bai, H.; Zhang, Z. PARP inhibitors in ovarian cancer: Sensitivity prediction and resistance mechanisms. J. Cell. Mol. Med. 2019, 23, 2303–2313. [Google Scholar] [CrossRef]
- Sgouros, G.; Dewaraja, Y.K.; Escorcia, F.; Graves, S.A.; Hope, T.A.; Iravani, A.; Pandit-Taskar, N.; Saboury, B.; James, S.S.; Zanzonico, P.B. Tumor Response to Radiopharmaceutical Therapies: The Knowns and the Unknowns. J. Nucl. Med. 2021, 62, 12S–22S. [Google Scholar] [CrossRef]
- O’Neill, E.; Kersemans, V.; Allen, P.D.; Terry, S.Y.A.; Torres, J.B.; Mosley, M.; Smart, S.; Lee, B.Q.; Falzone, N.; Vallis, K.A.; et al. Imaging DNA Damage Repair In Vivo after 177Lu-DOTATATE Therapy. J. Nucl. Med. 2020, 61, 743–750. [Google Scholar] [CrossRef]
- Vandersickel, V.; Slabbert, J.; Thierens, H.; Vral, A. Comparison of the colony formation and crystal violet cell proliferation assays to determine cellular radiosensitivity in a repair-deficient MCF10A cell line. Radiat. Meas. 2011, 46, 72–75. [Google Scholar] [CrossRef]
- Azzam, E.I.; Jay-Gerin, J.-P.; Pain, D. Ionizing radiation-induced metabolic oxidative stress and prolonged cell injury. Cancer Lett. 2012, 327, 48–60. [Google Scholar] [CrossRef]
- Kobashigawa, S.; Kashino, G.; Suzuki, K.; Yamashita, S.; Mori, H. Ionizing Radiation-Induced Cell Death Is Partly Caused by Increase of Mitochondrial Reactive Oxygen Species in Normal Human Fibroblast Cells. Radiat. Res. 2015, 183, 455–464. [Google Scholar] [CrossRef]
- Katerji, M.; Filippova, M.; Duerksen-Hughes, P. Approaches and Methods to Measure Oxidative Stress in Clinical Samples: Research Applications in the Cancer Field. Oxidative Med. Cell. Longev. 2019, 2019, 1279250. [Google Scholar] [CrossRef]
- Penninckx, S.; Pariset, E.; Cekanaviciute, E.; Costes, S.V. Quantification of radiation-induced DNA double strand break repair foci to evaluate and predict biological responses to ionizing radiation. NAR Cancer 2021, 3, zcab046. [Google Scholar] [CrossRef]
- Kataoka, Y.; Bindokas, V.P.; Duggan, R.C.; Murley, J.S.; Grdina, D.J. Flow Cytometric Analysis of Phosphorylated Histone H2AX Following Exposure to Ionizing Radiation in Human Microvascular Endothelial Cells. J. Radiat. Res. 2006, 47, 245–257. [Google Scholar] [CrossRef]
- Qin, C.; Ji, Z.; Zhai, E.; Xu, K.; Zhang, Y.; Li, Q.; Jing, H.; Wang, X.; Song, X. PARP inhibitor olaparib enhances the efficacy of radiotherapy on XRCC2-deficient colorectal cancer cells. Cell Death Dis. 2022, 13, 505. [Google Scholar] [CrossRef]
- Senra, J.M.; Telfer, B.A.; Cherry, K.E.; McCrudden, C.M.; Hirst, D.G.; O’Connor, M.J.; Wedge, S.R.; Stratford, I.J. Inhibition of PARP-1 by Olaparib (AZD2281) Increases the Radiosensitivity of a Lung Tumor Xenograft. Mol. Cancer 2011, 10, 1949–1958. [Google Scholar] [CrossRef]
- Cullinane, C.; Waldeck, K.; Kirby, L.; Rogers, B.E.; Eu, P.; Tothill, R.W.; Hicks, R.J. Enhancing the anti-tumour activity of 177 Lu-DOTA-octreotate radionuclide therapy in somatostatin receptor-2 expressing tumour models by targeting PARP. Sci. Rep. 2020, 10, 10196. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Qiu, F.; Stolniceanu, C.R.; Yu, F.; Zang, S.; Xiang, Y.; Huang, Y.; Matovic, M.; Stefanescu, C.; Tang, Q.; et al. Combined use of 177Lu-DOTATATE peptide receptor radionuclide therapy and fluzoparib for treatment of well-differentiated neuroendocrine tumors: A preclinical study. J. Neuroendocrinol. 2022, 34, e13109. [Google Scholar] [CrossRef]
- Powell, C.; Mikropoulos, C.; Kaye, S.B.; Nutting, C.M.; Bhide, S.A.; Newbold, K.; Harrington, K.J. Pre-clinical and clinical evaluation of PARP inhibitors as tumour-specific radiosensitisers. Cancer Treat. Rev. 2010, 36, 566–575. [Google Scholar] [CrossRef]
- Verhagen, C.V.M.; de Haan, R.; Hageman, F.; Oostendorp, T.P.D.; Carli, A.L.E.; O’Connor, M.J.; Jonkers, J.; Verheij, M.; van den Brekel, M.W.; Vens, C. Extent of radiosensitization by the PARP inhibitor olaparib depends on its dose, the radiation dose and the integrity of the homologous recombination pathway of tumor cells. Radiother. Oncol. 2015, 116, 358–365. [Google Scholar] [CrossRef]
- Rose, M.; Burgess, J.T.; O’Byrne, K.; Richard, D.J.; Bolderson, E. PARP Inhibitors: Clinical Relevance, Mechanisms of Action and Tumor Resistance. Front. Cell Dev. Biol. 2020, 8, 564601. [Google Scholar] [CrossRef]
- Ruigrok, E.A.M.; Verkaik, N.S.; de Blois, E.; de Ridder, C.; Stuurman, D.; Roobol, S.J.; Van Gent, D.C.; de Jong, M.; Van Weerden, W.M.; Nonnekens, J. Preclinical Assessment of the Combination of PSMA-Targeting Radionuclide Therapy with PARP Inhibitors for Prostate Cancer Treatment. Int. J. Mol. Sci. 2022, 23, 8037. [Google Scholar] [CrossRef]
- Pouget, J.-P.; Santoro, L.; Piron, B.; Paillas, S.; Ladjohounlou, R.; Pichard, A.; Poty, S.; Deshayes, E.; Constanzo, J.; Bardiès, M. From the target cell theory to a more integrated view of radiobiology in Targeted radionuclide therapy: The Montpellier group’s experience. Nucl. Med. Biol. 2022, 104–105, 53–64. [Google Scholar] [CrossRef]
- Tamborino, G.; De Saint-Hubert, M.; Struelens, L.; Seoane, D.C.; Ruigrok, E.A.M.; Aerts, A.; van Cappellen, W.A.; de Jong, M.; Konijnenberg, M.W.; Nonnekens, J. Cellular dosimetry of [177Lu]Lu-DOTA-[Tyr3]octreotate radionuclide therapy: The impact of modeling assumptions on the correlation with in vitro cytotoxicity. EJNMMI Phys. 2020, 7, 8. [Google Scholar] [CrossRef] [PubMed]
- Spetz, J.; Rudqvist, N.; Langen, B.; Parris, T.Z.; Dalmo, J.; Schüler, E.; Wängberg, B.; Nilsson, O.; Helou, K.; Forssell-Aronsson, E. Time-dependent transcriptional response of GOT1 human small intestine neuroendocrine tumor after 177Lu[Lu]-octreotate therapy. Nucl. Med. Biol. 2018, 60, 11–18. [Google Scholar] [CrossRef]
- Eves, P.; Layton, C.; Hedley, S.; Dawson, R.A.; Wagner, M.; Morandini, R.; Ghanem, G.; Neil, S.M. Characterization of an in vitro model of human melanoma invasion based on reconstructed human skin. Br. J. Dermatol. 2000, 142, 210–222. [Google Scholar] [CrossRef]
- Gembarska, A.; Luciani, F.; Fedele, C.; Russell, E.A.; Dewaele, M.; Villar, S.; Zwolinska, A.; Haupt, S.; de Lange, J.; Yip, D.; et al. MDM4 is a key therapeutic target in cutaneous melanoma. Nat. Med. 2012, 18, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Rambow, F.; Rogiers, A.; Marin-Bejar, O.; Aibar, S.; Femel, J.; Dewaele, M.; Karras, P.; Brown, D.; Chang, Y.H.; Debiec-Rychter, M.; et al. Toward Minimal Residual Disease-Directed Therapy in Melanoma. Cell 2018, 174, 843–855.e19. [Google Scholar] [CrossRef]
- Hill, R.P. The changing paradigm of tumour response to irradiation. Br. J. Radiol. 2017, 90, 20160474. [Google Scholar] [CrossRef]
- Marin, G.; Vanderlinden, B.; Karfis, I.; Guiot, T.; Wimana, Z.; Reynaert, N.; Vandenberghe, S.; Flamen, P. A dosimetry procedure for organs-at-risk in 177Lu peptide receptor radionuclide therapy of patients with neuroendocrine tumours. Phys. Med. 2018, 56, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Kyriakou, I.; Emfietzoglou, D.; Ivanchenko, V.; Bordage, M.C.; Guatelli, S.; Lazarakis, P.; Tran, H.N.; Incerti, S. Microdosimetry of electrons in liquid water using the low-energy models of Geant4. J. Appl. Phys. 2017, 122, 024303. [Google Scholar] [CrossRef]
- Asai, M.; Cortés-Giraldo, M.A.; Giménez-Alventosa, V.; Giménez Gómez, V.; Salvat, F. The PENELOPE Physics Models and Transport Mechanics. Implementation into Geant4. Front. Phys. 2021, 9, 660. [Google Scholar] [CrossRef]
- Scott, J.A. Photon, Electron, Proton and Neutron Interaction Data for Body Tissues: ICRU Report 46. International Commission on Radiation Units and Measurements, Bethesda, 1992, $40.00. J. Nucl. Med. 1993, 34, 171. [Google Scholar]
- Livechart—Table of Nuclides—Nuclear Structure and Decay Data. Available online: https://www-nds.iaea.org/relnsd/vcharthtml/VChartHTML.html (accessed on 1 June 2022).
- Penninckx, S.; Pariset, E.; Acuna, A.U.; Lucas, S.; Costes, S.V. Considering Cell Proliferation to Optimize Detection of Radiation-Induced 53BP1 Positive Foci in 15 Mouse Strains Ex Vivo. Radiat. Res. 2020, 195, 47–59. [Google Scholar] [CrossRef]









| CDI | AF (%) | |||
|---|---|---|---|---|
| 177Lu-DOTATATE | EBRT | 177Lu-DOTATATE | EBRT | |
| HBL | 0.84 | 0.72 | 20 | 34 |
| MM162 | 0.95 | 0.80 | 12 | 26 |
| COLO-677 | NA | NA | 0 | 5 |
| EJM | NA | NA | 7 | 5 |
| MIA-PACA-2 | NA | 0.58 | 0 | 43 |
| HT-29 | NA | 0.63 | 4 | 34 |
| % of Branching Ratios Used for the Simulations [44] | Energies (keV) | |
|---|---|---|
| Gamma | 17.3 | 71, 113, 136, 208, 250 and 321 |
| X-rays | 8.5 | 9, 55, 56 and 64 |
| β particles | 100.0 | 0 to 497 (Emax) |
| Auger electrons | 22.1 | 8, 48, 103 and 111 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delbart, W.; Karabet, J.; Marin, G.; Penninckx, S.; Derrien, J.; Ghanem, G.E.; Flamen, P.; Wimana, Z. Understanding the Radiobiological Mechanisms Induced by 177Lu-DOTATATE in Comparison to External Beam Radiation Therapy. Int. J. Mol. Sci. 2022, 23, 12369. https://doi.org/10.3390/ijms232012369
Delbart W, Karabet J, Marin G, Penninckx S, Derrien J, Ghanem GE, Flamen P, Wimana Z. Understanding the Radiobiological Mechanisms Induced by 177Lu-DOTATATE in Comparison to External Beam Radiation Therapy. International Journal of Molecular Sciences. 2022; 23(20):12369. https://doi.org/10.3390/ijms232012369
Chicago/Turabian StyleDelbart, Wendy, Jirair Karabet, Gwennaëlle Marin, Sébastien Penninckx, Jonathan Derrien, Ghanem E. Ghanem, Patrick Flamen, and Zéna Wimana. 2022. "Understanding the Radiobiological Mechanisms Induced by 177Lu-DOTATATE in Comparison to External Beam Radiation Therapy" International Journal of Molecular Sciences 23, no. 20: 12369. https://doi.org/10.3390/ijms232012369
APA StyleDelbart, W., Karabet, J., Marin, G., Penninckx, S., Derrien, J., Ghanem, G. E., Flamen, P., & Wimana, Z. (2022). Understanding the Radiobiological Mechanisms Induced by 177Lu-DOTATATE in Comparison to External Beam Radiation Therapy. International Journal of Molecular Sciences, 23(20), 12369. https://doi.org/10.3390/ijms232012369