
Group I Metabotropic Glutamate Receptors and Interacting Partners: An Update



{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Group I mGlu Receptors
3. MAPKs
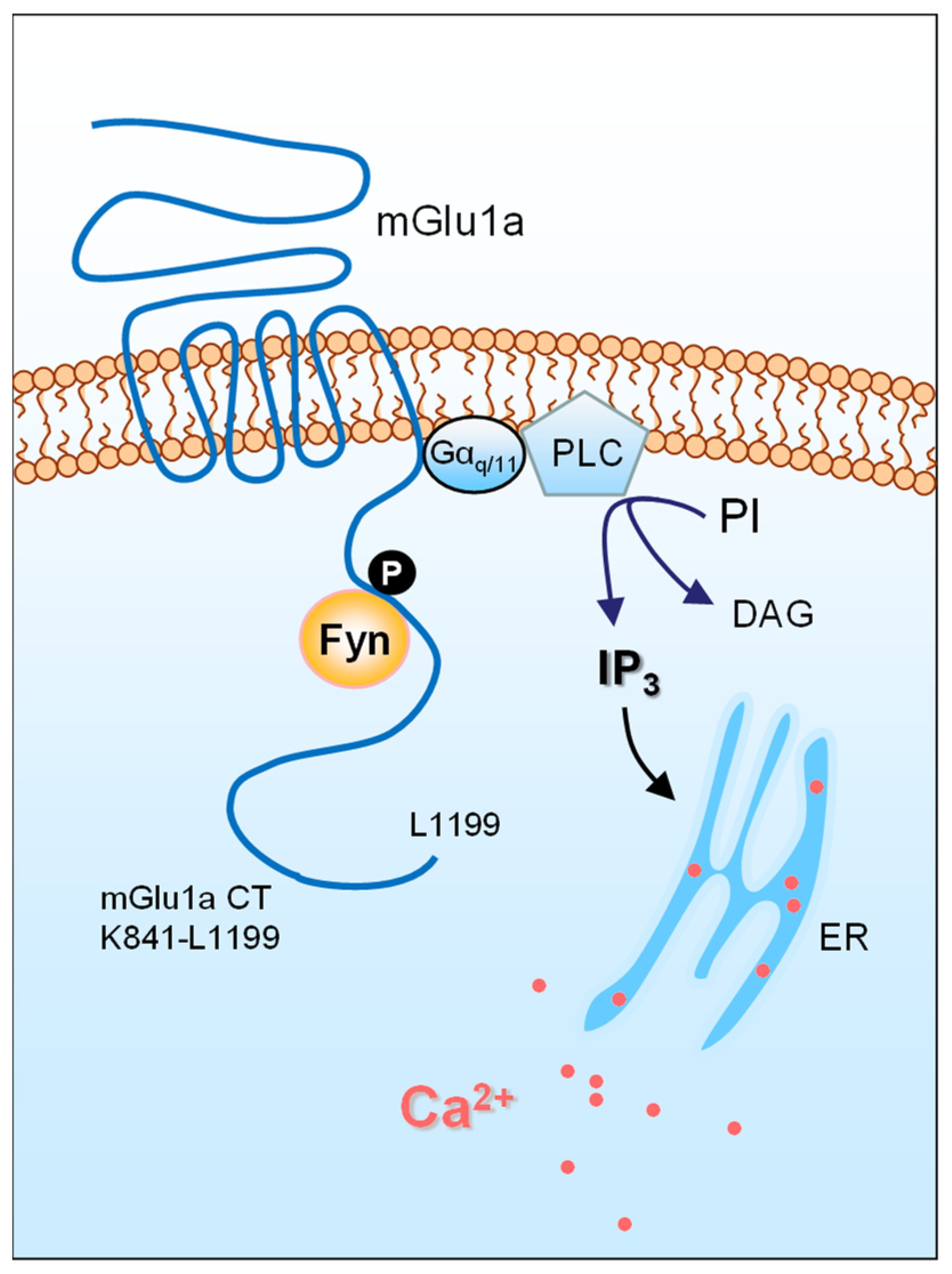
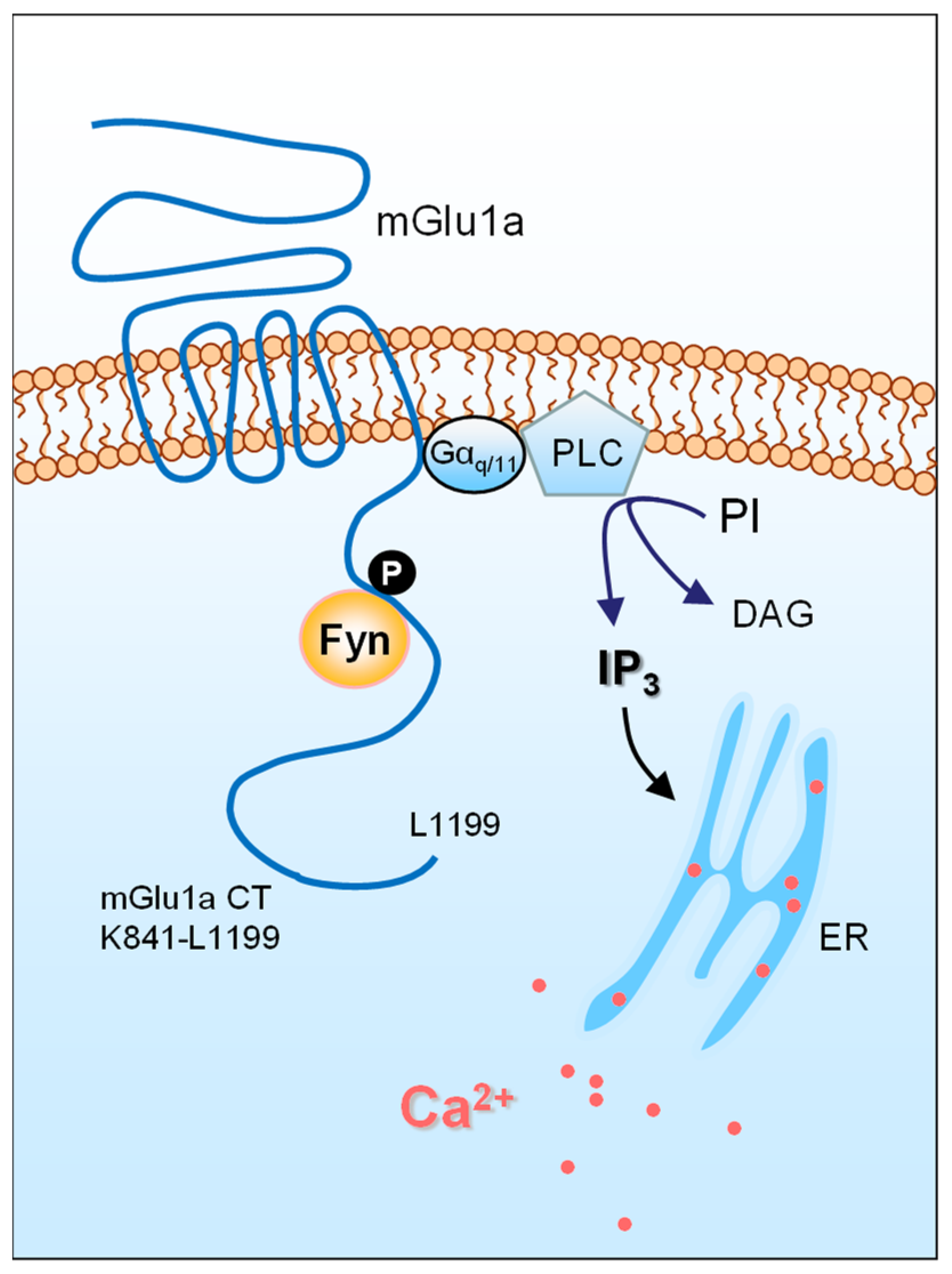
4. Fyn
5. CaMKII and PKC
6. Scaffolding Proteins
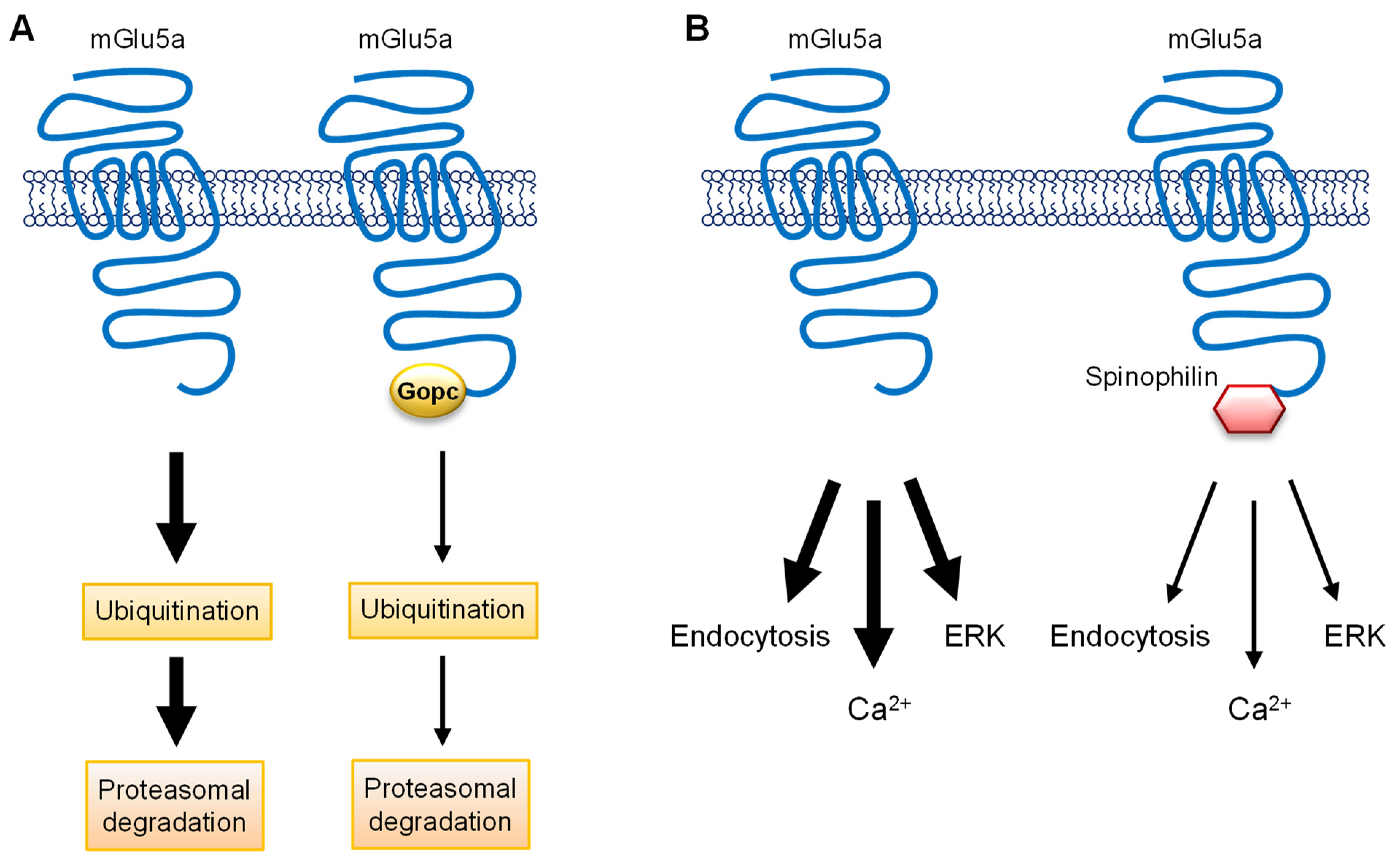
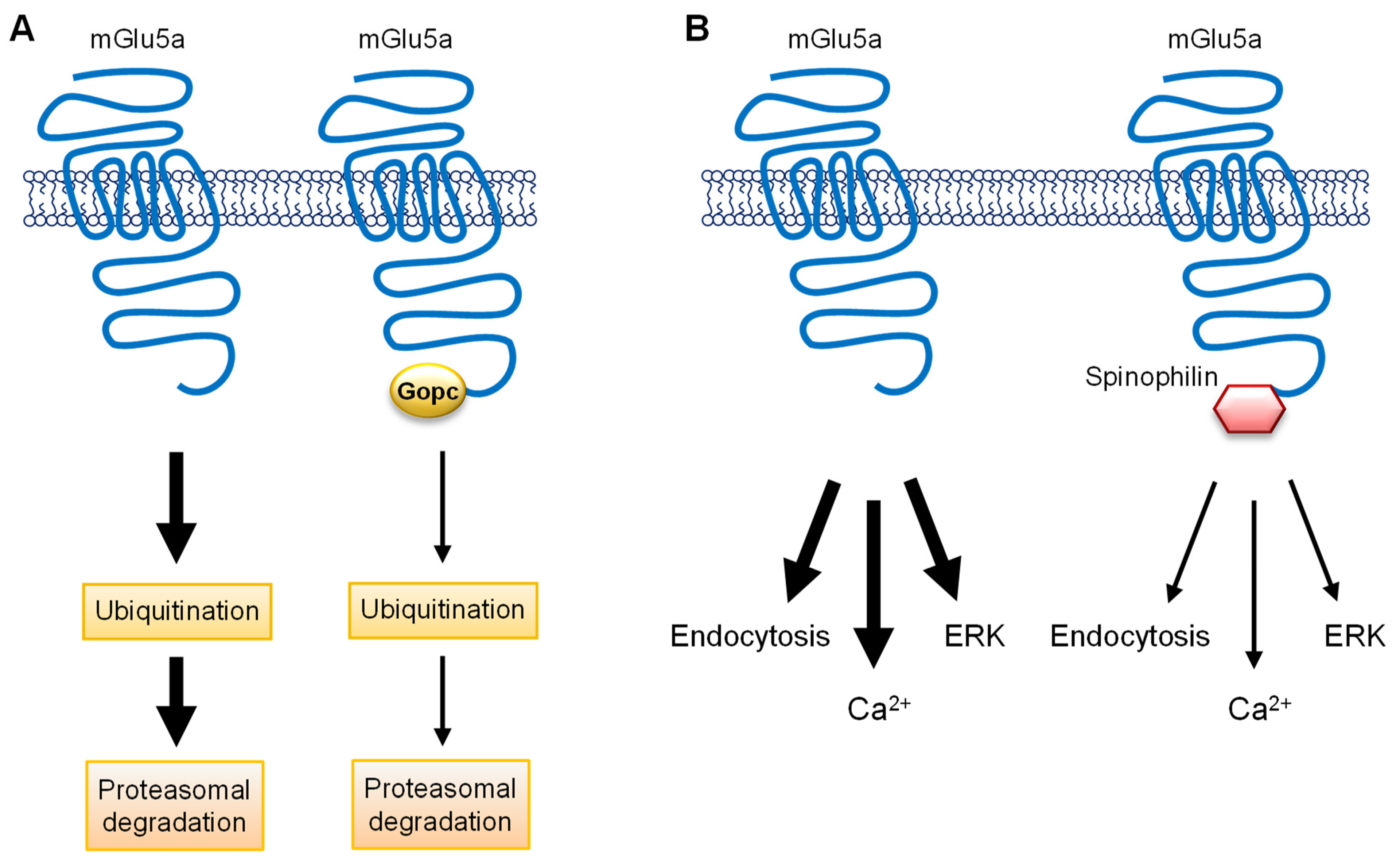
7. Proteins Involved in mGlu1/5 Trafficking
8. Other Interacting Partners
9. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AMPA | α-amino-3-hydroxy-5-methylisoxazole-4-propionic acid |
| BDNF | Brain-derived neurotrophic factor |
| BNST | bed nucleus of the stria terminalis |
| CaMKII | Ca2+/calmodulin-dependent protein kinase II |
| CT | C-terminus |
| DAG | diacylglycerol |
| ERK | extracellular signal-regulated kinase |
| Gopc | Golgi-associated PDZ and coiled-coil motif-containing protein |
| GPCR | G protein-coupled receptor |
| iGlu | ionotropic glutamate |
| IL2 | second intracellular loop |
| INM | inner nuclear membrane |
| IP3 | inositol-1,4,5-triphosphate |
| JNK | c-Jun N-terminal kinase |
| LTD | long-term depression |
| MAPK | mitogen-activated protein kinase |
| mGlu | metabotropic glutamate |
| NAc | nucleus accumbens |
| NMDA | N-methyl-d-aspartate |
| nRTK | non-receptor tyrosine kinase |
| PDZ | [PSD95/Drosophila disc large tumor suppressor (Dlg1)/zonula occludens-1 (zo-1)] |
| PKC | protein kinase C |
| PrPC | cellular prion protein |
| PrPSC | scrapie prion protein |
| PSD | postsynaptic density |
| SAPK | stress-activated protein kinase |
| SFK | Src family kinase |
| SNAP23 | synaptosome-associated protein 23 |
| S-SCAM | synaptic scaffolding molecule |
| SNARE | soluble N-ethylmaleimide-sensitive-factor attachment protein receptor |
| SNX | sorting nexin |
References
- Traynelis, S.F.; Wollmuth, L.P.; McBain, C.J.; Menniti, F.S.; Vance, K.M.; Ogden, K.K.; Hansen, K.B.; Yuan, H.; Myers, S.J.; Dingledine, R. Glutamate receptor ion channels: Structure, regulation, and function. Pharmacol. Rev. 2010, 62, 405–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregory, K.J.; Gouodet, C. International union of basic and clinical pharmacology. CXI. Pharmacology, signaling, and physiology of metabotropic glutamate receptors. Pharmacol. Rev. 2021, 73, 521–569. [Google Scholar] [CrossRef] [PubMed]
- Enz, R. Structure of metabotropic glutamate receptor C-terminal domains in contact with interacting proteins. Front. Mol. Neurosci. 2012, 5, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagni, L. Diversity of metabotropic glutamate receptor-interacting proteins and pathophysiological functions. Adv. Exp. Med. Biol. 2012, 970, 63–79. [Google Scholar] [PubMed]
- Suh, Y.H.; Chang, K.; Roche, K.W. Metabotropic glutamate receptor trafficking. Mol. Cell. Neurosci. 2018, 91, 10–24. [Google Scholar] [CrossRef]
- Bhattacharyya, S. Inside story of group I metabotropic glutamate receptors (mGluRs). Int. J. Biochem. Cell. Biol. 2016, 77, 205–212. [Google Scholar] [CrossRef]
- Ferraguti, F.; Crepaldi, L.; Nicoletti, F. Metabotropic glutamate 1 receptor: Current concepts and perspectives. Pharmacol. Rev. 2008, 60, 536–581. [Google Scholar] [CrossRef]
- Masu, M.; Tanabe, Y.; Tsuchida, K.; Shigemoto, R.; Nakanishi, S. Sequence and expression of a metabotropic glutamate receptor. Nature 1991, 349, 760–765. [Google Scholar] [CrossRef]
- Nicoletti, F.; Bockaert, J.; Collingridge, G.L.; Conn, P.J.; Ferraguti, F.; Schoepp, D.D.; Wroblewski, J.T.; Pin, J.P. Metabotropic glutamate receptors: From the workbench to the bedside. Neuropharmacology 2011, 60, 1017–1041. [Google Scholar] [CrossRef] [Green Version]
- Abe, T.; Sugihara, H.; Nawa, H.; Shigemoto, R.; Mizuno, N.; Nakanishi, S. Molecular characterization of a novel metabotropic glutamate receptor mGluR5 coupled to inositol phosphate/Ca2+ signal transduction. J. Biol. Chem. 1992, 267, 13361–13368. [Google Scholar] [CrossRef]
- Niswender, C.M.; Conn, P.J. Metabotropic glutamate receptors: Physiology, pharmacology, and disease. Annu. Rev. Pharmcol. Toxicol. 2010, 50, 295–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spampinato, S.F.; Copani, A.; Nicoletti, F.; Sortino, M.A.; Caraci, F. Metabotropic glutamate receptors in glial cells: A new potential target for neuroprotection? Front. Mol. Neurosci. 2018, 11, 414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catania, M.V.; D’Antoni, S.; Bonaccorso, C.M.; Aronica, E.; Bear, M.F.; Nicoletti, F. Group I metabotropic glutamate receptors: A role in neurodevelopmental diseases? Mol. Neurobiol. 2007, 35, 298–307. [Google Scholar] [CrossRef]
- Nicoletti, F.; Battaglia, G.; Storto, M.; Ngomba, R.T.; Iacovelli, L.; Arcella, A.; Gradini, R.; Sale, P.; Rampello, L.; De Vita, T.; et al. Metabotropic glutamate receptors: Beyond the regulation of synaptic transmission. Psychoneuroendocrinology 2007, 32 (Suppl. 1), S40–S45. [Google Scholar] [CrossRef]
- Yue, J.; Lopez, J.M. Understanding MAPK signaling pathways in apoptosis. Int. J. Mol. Sci. 2020, 21, 2346. [Google Scholar] [CrossRef] [Green Version]
- Songyang, Z.; Lu, K.P.; Kwon, Y.T.; Tsai, L.H.; Filhol, O.; Cochet, C.; Brickey, D.A.; Soderling, T.R.; Bartleson, C.; Graves, D.J.; et al. A structure basis for substrate specificities of protein Ser/Thr kinases: Primary sequence preference of casein kinases I and II, NIMA, phosphorylase kinase, calmodulin-dependent kinase II, CDK5, and Erk1. Mol. Cell. Biol. 1996, 16, 6486–6493. [Google Scholar] [CrossRef] [Green Version]
- Mao, L.M.; Wang, J.Q. Synaptically localized mitogen-activated protein kinases: Local substrates and regulation. Mol. Neurobiol. 2016, 53, 6309–6315. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.H.; Mao, L.M.; Choe, E.S.; Wang, J.Q. Synaptic ERK2 phosphorylates and regulates metabotropic glutamate receptor 1 in vitro and in neurons. Mol. Neurobiol. 2017, 54, 7156–7170. [Google Scholar] [CrossRef]
- Martin, L.J.; Blackstone, C.D.; Huganir, R.L.; Price, D.L. Cellular location of a metabotropic glutamate receptor in rat brain. Neuron 1992, 9, 259–270. [Google Scholar] [CrossRef]
- Shigemoto, R.; Nomura, S.; Ohishi, H.; Sugihara, H.; Nakanishi, S.; Mizuno, N. Immunohistochemical localization of a metabotropic glutamate receptor, mGluR5, in the rat brain. Neurosci. Lett. 1993, 163, 53–57. [Google Scholar] [CrossRef]
- Hu, J.H.; Yang, L.; Kammermeier, P.J.; Moore, C.G.; Brakeman, P.R.; Tu, J.; Yu, S.; Petralia, R.S.; Li, Z.; Zhang, P.W.; et al. Preso1 dynamically regulates group I metabotropic glutamate receptors. Nat. Neurosci. 2012, 15, 836–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, D.Z.; Mao, L.M.; Wang, J.Q. The role of extracellular signal-regulated kinases (ERK) in the regulation of mGlu5 receptors in neurons. J. Mol. Neurosci. 2018, 66, 629–638. [Google Scholar] [CrossRef]
- Campbell, R.R.; Domingo, R.D.; Williams, A.R.; Wroten, M.G.; McGregor, H.A.; Waltermire, R.S.; Greentree, D.I.; Goulding, S.P.; Thompson, A.B.; Lee, K.M.; et al. Increased alcohol-drinking induced by manipulations of mGlu5 phosphorylation within the bed nucleus of the stria terminalis. J. Neurosci. 2019, 39, 2745–2761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.M.; Hu, J.H.; Milshteyn, A.; Zhang, P.W.; Moore, C.G.; Park, S.; Datko, M.C.; Domingo, R.D.; Reyes, C.M.; Wang, X.J.J.; et al. A prolyl-isomerase mediates dopamine-dependent plasticity and cocaine motor sensitization. Cell 2013, 154, 637–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fultz, E.K.; Quadir, S.G.; Martin, D.; Flaherty, D.M.; Worley, P.F.; Kippin, T.E.; Szumlinski, K.K. ERK-directed phosphorylation of mGlu5 gates methamphetamine reward and reinforcement in mouse. Int. J. Mol. Sci. 2021, 22, 1473. [Google Scholar] [CrossRef]
- Yang, J.H.; Sohn, S.; Kim, S.; Kim, J.; Oh, J.H.; Ryu, I.S.; Go, B.S.; Choe, E.S. Repeated nicotine exposure increases the intracellular interaction between ERK-mGluR5 in the nucleus accumbens more in adult than adolescent rats. Addict. Biol. 2021, 26, e12913. [Google Scholar] [CrossRef]
- Mao, L.M.; Geosling, R.; Penman, B.; Wang, J.Q. Local substrates of non-receptor tyrosine kinases at synaptic sites in neurons. Sheng Li Xue Bao 2017, 69, 657–665. [Google Scholar]
- Ohnishi, H.; Murata, Y.; Okazawa, H.; Matozaki, T. Src family kinases: Modulators of neurotransmitter receptor function and behavior. Trends Neurosci. 2011, 34, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Rajani, V.; Sengar, A.S.; Salter, M.W. Src and Fyn regulation of NMDA receptors in health and disease. Neuropharmacology 2021, 193, 108615. [Google Scholar] [CrossRef]
- Jin, D.Z.; Mao, L.M.; Wang, J.Q. An essential role of Fyn in the modulation of metabotropic glutamate receptor 1 in neurons. eNeuro 2017, 4, ENEURO.0096-17.2017. [Google Scholar] [CrossRef] [Green Version]
- Orlando, L.R.; Dunah, A.W.; Standaert, D.G.; Young, A.B. Tyrosine phosphorylation of the metabotropic glutamate receptor mGluR5 in striatal neurons. Neuropharmacology 2002, 43, 161–173. [Google Scholar] [CrossRef]
- Giuffrida, R.; Musumeci, S.; D’Antoni, S.D.; Bonaccorso, C.M.; Giuffrida-Stella, A.M.; Oostra, B.A.; Catania, M.V. A reduced number of metabotropic glutamate receptor subtype 5 receptors are associated with constitutive homer proteins in a mouse model of fragile X syndrome. J. Neurosci. 2005, 25, 8908–8916. [Google Scholar] [CrossRef] [Green Version]
- Mao, L.M.; Wang, J.Q. Dopamine D2 receptors are involved in the regulation of Fyn and metabotropic glutamate receptor 5 phosphorylation in the rat striatum in vivo. J. Neurosci. Res. 2016, 94, 329–338. [Google Scholar] [CrossRef] [Green Version]
- Mao, L.M.; Wang, J.Q. Linkage of non-receptor tyrosine kinase Fyn to mGlu5 receptors in striatal neurons in a depression model. Neuroscience 2020, 433, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Mundell, S.J.; Matharu, A.L.; Pula, G.; Holman, D.; Roberts, P.J.; Kelly, E. Metabotropic glutamate receptor 1 internalization induced by muscarinic acetylcholine receptor activation: Differential dependency of internalization of splice variants on nonvisual arrestins. Mol. Pharmacol. 2002, 61, 1114–1123. [Google Scholar] [CrossRef] [Green Version]
- Mundell, S.J.; Pula, G.; McIIhinney, R.A.J.; Roberts, P.J.; Kelly, E. Desensitization and internalization of metabotropic glutamate receptor 1a following activation of heterologous Gq/11-coupled receptors. Biochemistry 2004, 43, 7541–7551. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.Z.; Guo, M.L.; Xue, B.; Fibuch, E.E.; Choe, E.S.; Mao, L.M.; Wang, J.Q. Phosphorylation and feedback regulation of metabotropic glutamate receptor 1 by calcium/calmodulin-dependent protein kinase II. J. Neurosci. 2013, 33, 3402–3412. [Google Scholar] [CrossRef] [Green Version]
- Jin, D.Z.; Guo, M.L.; Xue, B.; Mao, L.M.; Wang, J.Q. Differential regulation of CaMKIIα interactions with mGluR5 and NMDA receptors by Ca2+ in neurons. J. Neurochem. 2013, 127, 620–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marks, C.; Chonesy, B.C.; Wang, X.; Stephenson, J.R.; Niswender, C.M.; Colbran, R.J. Activated CaMKIIα binds to the mGlu5 metabotropic glutamate receptor and modulates calcium mobilization. Mol. Pharmacol. 2018, 94, 1352–1362. [Google Scholar] [CrossRef] [Green Version]
- Raka, F.; Di Sebastiano, A.R.; Kulhawy, S.C.; Ribeiro, F.M.; Godin, C.M.; Caetano, F.A.; Angers, S.; Ferguson, S.S.G. Ca2+/calmodulin-dependent protein kinase II interacts with group I metabotropic glutamate and facilitates receptor endocytosis and ERK1/2 signaling: Role of β-amyloid. Mol. Brain 2015, 8, 21. [Google Scholar] [CrossRef] [Green Version]
- Mao, L.M.; Liu, X.Y.; Zhang, G.C.; Chu, X.P.; Fibuch, E.E.; Wang, L.S.; Liu, Z.; Wang, J.Q. Phosphorylation of group I metabotropic glutamate receptors (mGluR1/5) in vitro and in vivo. Neuropharmacology 2008, 55, 403–408. [Google Scholar] [CrossRef] [Green Version]
- Schwendt, M.; Olive, M.F. Protein kinase Cε activity regulates mGluR5 surface expression in the rat nucleus accumbens. J. Neurosci. Res. 2017, 95, 1079–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vergouts, M.; Doyen, P.J.; Peeters, M.; Opsomer, R.; Hermans, E. Constitutive downregulation protein kinase C epsilon in hSOD1G93A astrocytes influences mGluR5 signaling and the regulation of glutamate uptake. Glia 2018, 66, 749–761. [Google Scholar] [CrossRef] [PubMed]
- Bockaert, J.; Perroy, J.; Ango, F. The complex formed by group I metabotropic glutamate receptor (mGluR) and Homer1a plays a central role in metaplasticity and homeostatic synaptic scaling. J. Neurosci. 2021, 41, 5567–5578. [Google Scholar] [CrossRef] [PubMed]
- Brakeman, P.R.; Lanahan, A.A.; O’Brien, R.; Roche, K.; Barnes, C.A.; Huganir, R.L.; Worley, P.F. Homer: A protein that selectively binds metabotropic glutamate receptors. Nature 1997, 386, 284–288. [Google Scholar] [CrossRef] [PubMed]
- Tu, J.C.; Xiao, B.; Yuan, J.P.; Lanahan, A.A.; Leoffert, K.; Li, M.; Linden, D.J.; Worley, P.F. Homer binds a novel proline-rich motif and links and links group 1 metabotropic glutamate receptors with IP3 receptors. Neuron 1998, 21, 717–726. [Google Scholar] [CrossRef] [Green Version]
- Buscemi, L.; Ginet, V.; Lopatar, J.; Montana, V.; Pucci, L.; Spagnuolo, P.; Zehnder, T.; Grubisic, V.; Truttman, A.; Sala, C.; et al. Homer1 scaffold proteins govern Ca2+ dynamics in normal and reactive astrocytes. Cereb. Cortex 2017, 27, 2365–2384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chokshi, V.; Gao, M.; Grier, B.D.; Owens, A.; Wang, H.; Worley, P.F.; Lee, H.K. Input-specific metaplasticity in the visual cortex requires Homer1a-mediated mGluR5 signaling. Neuron 2019, 104, 736–748. [Google Scholar] [CrossRef]
- Diering, G.H.; Nirujogi, R.S.; Roth, R.H.; Worley, P.F.; Pandey, A.; Huganir, R.L. Homer1a drives homeostatic scaling-down of excitatory synapses during sleep. Science 2017, 355, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Chokshi, V.; Druciak, B.; Worley, P.F.; Lee, H.K. Homer1a is required for establishment of contralateral bias and maintenance of ocular dominance in mouse visual cortex. J. Neurosci. 2019, 39, 3897–3905. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.; Huang, M.; Zhang, Y.; Wang, W.; Ma, X.; Shi, H.; Worley, P.F.; Kim, D.K.; Fedorovich, S.V.; Jiang, W.; et al. Disabling phosphorylation at the Homer ligand of the metabotropic glutamate receptor 5 alleviates complete Freund’s adjuvant-induced inflammatory pain. Neuropharmacology 2020, 170, 108046. [Google Scholar] [CrossRef]
- Guo, W.; Ceolin, L.; Collins, K.A.; Perroy, J.; Huber, K.M. Elevated CaMKIIα and hyperphosphorylation of Homer mediate circuit dysfunction of in a Fragile X syndrome mouse model. Cell Rep. 2015, 13, 2297–2311. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Molinaro, G.; Collins, K.; Hays, S.A.; Paylor, R.; Worley, P.F.; Szumlinski, K.K.; Huber, K.M. Selective disruption of metabotropic glutamate receptor 5-Homer interactions mimics phenotypes of Fragile X Syndrome in mice. J. Neurosci. 2016, 36, 2131–2147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.H.; Worley, P.F.; Kammermeier, P.J. Dynamic regulation of Homer binding to group I metabotropic glutamate receptors by Preso1 and converging kinase cascades. J. Pharmacol. Exp. Ther. 2017, 361, 122–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitano, J.; Kimura, K.; Yamazaki, Y.; Soda, T.; Shigemoto, R.; Nakajima, Y.; Nakanishi, S. Tamalin, a PDZ domain-containing, links a protein complex formation of group 1 metabotropic glutamate receptors the guanine nucleotide exchange factor cytohesins. J. Neurosci. 2002, 22, 1280–1289. [Google Scholar] [CrossRef] [Green Version]
- Pandey, S.; Ramsakha, N.; Sharma, R.; Gulia, R.; Ojha, P.; Lu, W.; Bhattacharyya, S. The post-synaptic scaffolding protein tamalin regulates ligand-mediated trafficking of metabotropic glutamate receptors. J. Biol. Chem. 2020, 295, 8575–8588. [Google Scholar] [CrossRef]
- Neyman, S.; Braunewell, K.H.; O’Connell, K.E.; Dev, K.K.; Manahan-Vaughan, D. Inhibition of the interaction between group I metabotropic glutamate receptors and PDZ-domain proteins prevents hippocampal long-term depression, but not long-term potentiation. Front. Synaptic Neurosci. 2019, 11, 13. [Google Scholar] [CrossRef]
- Zhang, J.; Cheng, S.; Xiong, Y.; Ma, Y.; Luo, D.; Jeromin, A.; Zhang, H.; He, J. A novel association of mGluR1a with the PDZ scaffold protein CAL modulates receptor activity. FEBS Lett. 2008, 582, 4117–4124. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.; Zhang, J.; Zhu, P.; Ma, Y.; Xiong, Y.; Sun, L.; Xu, J.; Zhang, H.; He, J. The PDZ domain protein CAL interacts with mGluR5a and modulates receptor expression. J. Neurochem. 2010, 112, 588–598. [Google Scholar] [CrossRef] [PubMed]
- Klussendorf, M.; Song, I.; Schau, L.; Morellini, F.; Dityatev, A.; Koliwer, J.; Kreienkamp, H.J. The Golgi-associated PDZ domain protein Gopc/PIST is required for synaptic targeting of mGluR5. Mol. Neurobiol. 2021, 58, 5618–5634. [Google Scholar] [CrossRef]
- Luo, W.Y.; Xing, S.Q.; Zhu, P.; Zhang, C.G.; Yang, H.M.; Van Halm-Lutterodt, N.; Gu, L.; Zhang, H. PDZ scaffold protein CAL couples with metabotropic glutamate receptor 5 to protect against cell apoptosis and is a potential target in the treatment of Parkinson’s disease. Neurotherapeutics 2019, 16, 761–783. [Google Scholar] [CrossRef] [PubMed]
- Sarrouilhe, D.; Di Tommaso, A.; Metaye, T.; Ladeveze, V. Spinophilin: From partners to functions. Biochimie 2006, 88, 1099–1113. [Google Scholar] [CrossRef] [PubMed]
- Dunn, H.A.; Ferguson, S.S.G. PDZ protein regulation of G protein-coupled receptor trafficking and signaling pathways. Mol. Pharmacol. 2015, 88, 624–639. [Google Scholar] [CrossRef] [Green Version]
- Di Sebastiano, A.R.; Fahim, S.; Dunn, H.A.; Walther, C.; Ribeiro, F.M.; Cregan, S.P.; Angers, S.; Schmid, S.; Ferguson, S.S.G. Role of spinophilin in group I metabotropic glutamate receptor endocytosis, signaling, and synaptic plasticity. J. Biol. Chem. 2016, 291, 17602–17615. [Google Scholar] [CrossRef] [Green Version]
- Raynaud, F.; Homburger, V.; Seveno, M.; Vigy, O.; Moutin, E.; Fagni, L.; Perroy, J. SNAP23-Kif5 complex controls mGlu1 receptor trafficking. J. Mol. Cell. Biol. 2018, 10, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Ravichandran, V.; Chawla, A.; Roche, P.A. Identification of a novel syntaxin- and synaptobrevin/VAMP-binding protein, SNAP-23, expressed in non-neuronal tissues. J. Biol. Chem. 1996, 271, 13300–13303. [Google Scholar] [CrossRef] [Green Version]
- Uemura, T.; Shepherd, S.; Ackerman, L.; Jan, L.Y.; Jan, Y.N. Numb, a gene required in determination of cell fate during sensory organ formation in Drosophila embryos. Cell 1989, 58, 349–360. [Google Scholar] [CrossRef]
- Zhou, L.; Yang, D.; Wang, D.J.; Xie, Y.J.; Zhou, J.H.; Zhou, L.; Huang, H.; Han, S.; Shao, C.Y.; Li, H.S.; et al. Numb deficiency in cerebellar Purkinjie cells impairs synaptic expression of metabotropic glutamate receptor and motor coordination. Proc. Natl. Acad. Sci. USA 2015, 112, 15474–15479. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Cai, X.Y.; Gao, W.; Shen, Y. Numb-p72, but not Numb-p65, contributes to the trafficking of group I metabotropic glutamate receptors. Neuroreport 2018, 29, 902–906. [Google Scholar] [CrossRef]
- Wang, N.; Wang, D.D.; Shen, Y. Numb deficiency causes impaired trafficking of mGlu5 in neurons and autistic-like behaviors. Neurosci. Lett. 2019, 707, 134291. [Google Scholar] [CrossRef]
- Wang, N.; Wang, D.D.; Hou, X.; Li, X.; Shen, Y. Different roles of Numb-p72 and Numb-p65 on the trafficking of metabotropic glutamate receptor 5. Mol. Biol. Rep. 2021, 48, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Vieira, N.; Rito, T.; Correia-Neves, M.; Sousa, N. Sorting out sorting nexins functions in the nervous system in health and disease. Mol. Neurobiol. 2021, 58, 4070–4106. [Google Scholar] [CrossRef] [PubMed]
- Heydorn, A.; Sondergaard, B.P.; Ersboll, B.; Holst, B.; Nielsen, F.C.; Haft, C.R.; Wistler, J.; Schwartz, T.W. A library of 7TM receptor C-terminal tails. Interactions with the proposed post-endocytic sorting proteins ERM-binding phosphoprotein 50 (EBP50), N-ethylmaleimide-sensitive factor (NSF), sorting nexin 1 (SNX1), and G protein-coupled receptor-associated sorting protein (GASP). J. Biol. Chem. 2004, 279, 54291–54303. [Google Scholar]
- Pandey, S.; Mahato, P.K.; Bhattacharyya, S. Metabotropic glutamate receptor 1 recycles to the cell surface in protein phosphatase 2A-dependent manner in non-neuronal and neuronal cell lines. J. Neurochem. 2014, 131, 602–614. [Google Scholar] [CrossRef]
- Sharma, R.; Gulia, R.; Bhattacharyya, S. A critical role for sorting Nextin 1 in the trafficking of metabotropic glutamate receptors. J. Neurosci. 2018, 38, 8605–8620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodman, O.B., Jr.; Krupnick, J.G.; Santini, F.; Gurevich, V.V.; Penn, R.B.; Gagnon, A.W.; Keen, J.H.; Benovic, J.L. Beta-arrestin acts as a clathrin adaptor in endocytosis of the beta2-adrenergic receptor. Nature 1996, 383, 447–450. [Google Scholar] [CrossRef]
- Van Gastel, J.; Hendrickx, J.O.; Leysen, H.; Santos-Otte, P.; Luttrell, L.M.; Martin, B.; Maudsley, S. β-Arrestin based receptor signaling paradigms: Potential therapeutic targets for complex age-related disorders. Front. Pharmacol. 2018, 9, 1369. [Google Scholar] [CrossRef]
- Eng, A.G.; Kelver, D.A.; Hedrick, T.P.; Swanson, G.T. Transduction of group I mGluR-mediated synaptic plasticity by β-arrestin2 signalling. Nat. Commun. 2016, 7, 13571. [Google Scholar] [CrossRef] [Green Version]
- Stoppel, L.J.; Auerbach, B.D.; Senter, R.K.; Preza, A.R.; Lefkowitz, R.J.; Bear, M.F. β-arrestin2 couples metabotropic glutamate receptor 5 to neuronal protein synthesis and is a potential target to treat fragile X. Cell Rep. 2017, 18, 2807–2814. [Google Scholar] [CrossRef]
- Takada, L.T.; Geschwind, M.D. Prion diseases. Semin. Neurol. 2013, 33, 348–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linden, R. The biological function of the prion protein: A cell surface scaffold of signaling modules. Front. Mol. Neurosci. 2017, 10, 77. [Google Scholar] [CrossRef] [Green Version]
- Matsubara, T.; Satoh, K.; Homma, T.; Nakagaki, T.; Yamaguchi, N.; Atarashi, R.; Sudo, Y.; Uezono, Y.; Ishibashi, D.; Nishida, N. Prion protein interacts with the metabotropic glutamate receptor 1 and regulates the organization of Ca2+ signaling. Biochem. Biophys. Res. Commun. 2020, 525, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Beraldo, F.H.; Arantes, C.P.; Santos, T.G.; Machado, C.F.; Roffe, M.; Hajj, G.N.; Lee, K.S.; Magalhaes, A.C.; Caetano, F.A.; Mancini, G.L.; et al. Metabotropic glutamate receptors transduce signals from neurite outgrowth after binding of the prion protein to laminin γ1 chain. FASEB J. 2011, 25, 265–279. [Google Scholar] [CrossRef]
- Um, J.W.; Kaufman, A.C.; Kostylev, M.; Heiss, J.K.; Stagi, M.; Takahashi, H.; Kerrisk, M.E.; Vortmeyer, A.; Wisniewski, T.; Koleske, A.J.; et al. Metabotropic glutamate receptor 5 is a co-receptor for Alzheimer Aβ oligomer bound to cellular prion protein. Neuron 2013, 79, 887–902. [Google Scholar] [CrossRef] [Green Version]
- Goniotaki, D.; Lakkaraju, A.K.K.; Shrivastava, A.N.; Bakirci, P.; Sorce, S.; Senatore, A.; Marpakwar, R.; Hornemann, S.; Gasparini, F.; Triller, A.; et al. Inhibition of group-I metabotropic glutamate receptors protects against prion toxicity. PLoS Pathog. 2017, 13, e1006733. [Google Scholar] [CrossRef] [PubMed]
- Jong, Y.J.I.; Harmon, S.K.; O’Malley, K.L. GPCR signalling from within the cell. Br. J. Pharmacol. 2018, 175, 4026–4035. [Google Scholar] [CrossRef] [Green Version]
- Jong, Y.J.I.; Sergin, I.; Purgert, C.A.; O’Malley, K.L. Location-dependent signaling of the group I metabotropic glutamate receptor mGlu5. Mol. Pharmacol. 2014, 86, 774–785. [Google Scholar] [CrossRef] [Green Version]
- Sergin, I.; Jong, Y.J.; Harmon, S.K.; Kumar, V.; O’Malley, K.L. Sequences within the C terminus of the metabotropic glutamate receptor 5 (mGluR5) are responsible for inner nuclear membrane localization. J. Biol. Chem. 2017, 292, 3637–3655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, L.-M.; Bodepudi, A.; Chu, X.-P.; Wang, J.Q. Group I Metabotropic Glutamate Receptors and Interacting Partners: An Update. Int. J. Mol. Sci. 2022, 23, 840. https://doi.org/10.3390/ijms23020840
Mao L-M, Bodepudi A, Chu X-P, Wang JQ. Group I Metabotropic Glutamate Receptors and Interacting Partners: An Update. International Journal of Molecular Sciences. 2022; 23(2):840. https://doi.org/10.3390/ijms23020840
Chicago/Turabian StyleMao, Li-Min, Alaya Bodepudi, Xiang-Ping Chu, and John Q. Wang. 2022. "Group I Metabotropic Glutamate Receptors and Interacting Partners: An Update" International Journal of Molecular Sciences 23, no. 2: 840. https://doi.org/10.3390/ijms23020840
APA StyleMao, L.-M., Bodepudi, A., Chu, X.-P., & Wang, J. Q. (2022). Group I Metabotropic Glutamate Receptors and Interacting Partners: An Update. International Journal of Molecular Sciences, 23(2), 840. https://doi.org/10.3390/ijms23020840