
MicroRNAs as Modulators of the Immune Response in T-Cell Acute Lymphoblastic Leukemia



Abstract
:1. Introduction

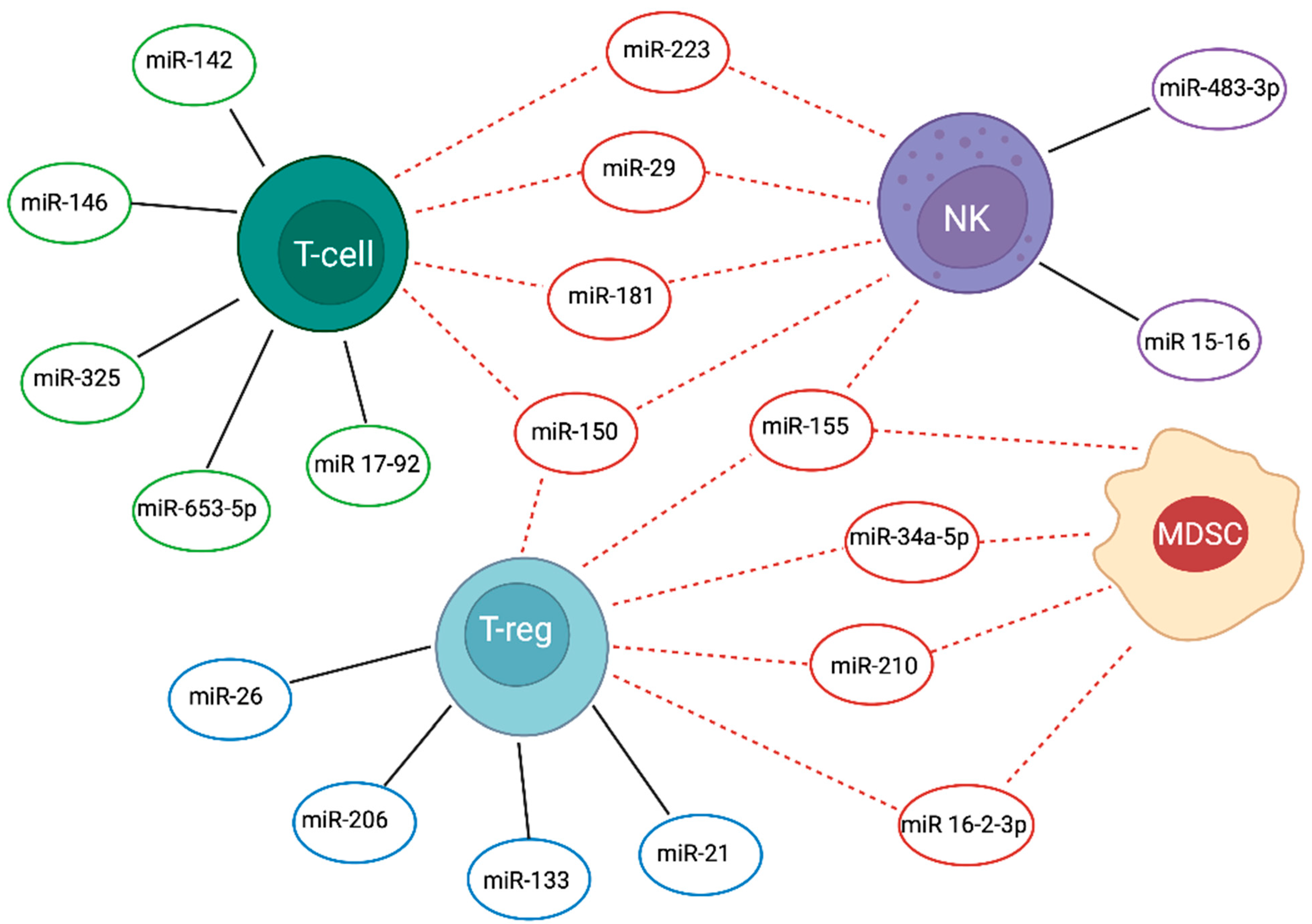{kind=link}
| T Cells | NK | T Reg | MDSC | T-ALL | Reference | |
|---|---|---|---|---|---|---|
| miR-15-16 | - | Promotes the transition to stage 4 (CD27¯CD11b+) during NK maturation | - | - | Blocks NK cell maturation and leads to accumulation of immature stage-2 and stage-3 cells | [6] |
| miR-16-2-3p | - | - | Blocks the function in the tumour microenvironment by inhibiting PD1/PDL1 signalling pathway | Blocks the function in the tumour microenvironment by inhibiting PD1/PDL1 signalling pathway | Down-regulation in ALL leads to the recruitment of both MDSCs and Tregs; inhibits functional T-cell responses in ALL | [7,8] |
| Cluster 17-92 | Increases T-cells proliferation and survival in DN–DP transition; high expression leads to autoimmunity | - | - | - | Negatively regulates E2F1 to suppress T-ALL apoptosis | [9,10,11] |
| miR-21 | - | - | Induces IL-10 and impairs Treg cell function | - | Up-regulation in T-ALL promotes survival suppressing Pdcd4 at least by stabilization of BCL-XL protein levels; immunosuppressive effects of IL-10 by Tregs leads to unchecked inflammation and promotes T-ALL progression | [12,13,14] |
| miR-26 | - | - | Promotes Tregs production and function by targeting IL-6 | - | Down-regulation in T-ALL patients leads to decreased apoptosis and induction of proliferation caused by failed repression of PIK3CD | [15,16] |
| miR-29a/b | miR-29a acts as tumour suppressor targeting DNMT3a and DNMT3b to reduce methylation in leukemic T-cells | miR-29b represses NK cells cytotoxic function and differentiation in Notch1-T-ALL | - | - | miR-29b downregulation in T-ALL causes altered epigenetic status activating methylation of DNMT3a, DNMT3b and TET1; miR-29b up-regulation via vesicular release by leukemic cells influences NK cell development; miR-29a low expression in T-ALL | [17,18,19,20] |
| miR-34a-5p | miR-34a acts as tumour suppressor, central NF-κB regulator in T-cell | - | - | Up-regulation of miR- 34a- 5p promotes the expansion of MDSCs | Down-regulation in T-ALL impacts the NF-κB signalosome increasing surface abundance of TCRA and CD3E caused by lost targeting of NF-κBIA and five PKC isozymes | [21,22,23] |
| miR-133b | - | - | Modulates Tregs differentiation by IL-17A expression | - | Reduced expression in association with low levels of IL-17 levels in T-ALL | [16,24] |
| miR-142-3p | Modulated in thymocytes differentiation | - | - | - | Over-expression in T-ALL reduces cAMP levels and interacts with PKA, to increase leukemic T-cell growth; targeting GRa it induces glucorticoid resistance | [25] |
| miR-146b-5p | High expression in thymocytes regulates the DP–SP transition | - | - | - | Tumour suppressor function delaying progression of T-ALL; Downregulated by over-expressed TAL1 in T-ALL patients | [26] |
| miR-150 | Modulates T-cell differentiation; regulates Notch3 expression | Promotes NK cell survival and maturation | Up-regulation blocks mTOR pathway and enhances Treg cell differentiation | - | Induces accumulation of hyperfunctional mature NK cells; inversely correlated to Notch3 expression | [27,28,29] |
| miR-155 | Positively regulates Th17 differentiation. Pivotal role in regulation of T-cell response during T-cell activation | Promotes the transition to stage 4 (CD27¯CD11b+) during NK cell maturation | Targets FOXP3, to modulating Tregs differentiation and function; affects IL-2 production by inhibiting SOCS1 to regulate Treg proliferation | Down-regulation promotes expansion of functional MDSCs by targeting SHIP-1 and PTEN | Blocks NK cell maturation and leads to immature stage 2 and 3 cells accumulation; Low-expression, induced by IL-10, promotes T-ALL progression | [12,30,31,32,33,34,35,36] |
| miR-181 | High expression during the DP stage and low mature T-cells | Up-regulates Notch signalling in NK cell development by targeting NLK | - | - | Is a tumour suppressor in ALL with over-expression of Smad7 and regulates TGFβ1/Smad pathway | [37,38,39] |
| miR-210 | - | - | Up-regulates FOXP3 and arginase 1, and enhances Treg cells function | Regulates MDSC function by increasing arginase activity and nitric oxide production and, in splenic MDSC, regulates arg1, CXCL12and IL16 | Up-regulation of miR-210 in ALL patients may potentiated immunosuppressive activity of tumour MDSCs | [40,41] |
| miR-223 | High expression in early thymocytes; down-regulation in DN2–DN3 stage | - | - | Down-regulation represses the differentiation and accumulation of MDSCs by targeting MEF2C | Inhibits the expression of the tumour suppressor FBXW7 and induces T-ALL cell growth in a Notch-dependent manner | [42,43,44,45] |
| miR-325 | Controls T-cell proliferation and apoptosis by binding BAG2 | - | - | - | Low-expression enhances BAG2 levels in patients with T-ALL and promotes cancer cells proliferation | [46] |
| miR-483-3p | - | Involved in the development and cytolytic function of NK by modulating IGF-1 | - | - | Higher expression level plays a positive role in T-ALL pathophysiology | [47,48] |
| miR-653-5p | Enhances apoptosis and autophagy | - | - | - | Negatively regulated by circ-PRKD promotes T-ALL cell proliferation | [49] |
2. T-Cell Acute Lymphoblastic Leukaemia
3. Overview of the Immune Mechanisms in T-ALL
4. Biogenesis and Role of miRNAs
5. miRNA and Natural Killer (NK) Cells
5.1. miRNA in NK Cell Development: An Overview
5.2. miRNAs Modulate NK Cell Function in T-ALL
6. miRNAs and T Cell
6.1. miRNAs in T Cell Development: An Overview
6.2. miRNAs Dysregulate T Cell Immune Response in T-ALL
6.3. The Modulatory Role of miRNAs in Treg Cell Function in T-ALL
7. MDSCs and Tregs
8. Conclusions and Future Perspectives
Funding
Conflicts of Interest
References
- Jia, Y.; Wei, Y. Modulators of MicroRNA Function in the Immune System. Int. J. Mol. Sci. 2020, 21, 2357. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Ansel, K.M. RNA regulation of the immune system. Immunol. Rev. 2013, 253, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Schetter, A.J.; Heegaard, N.H.; Harris, C.C. Inflammation and cancer: Interweaving microRNA, free radical, cytokine and p53 pathways. Carcinogenesis 2010, 31, 37–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tili, E.; Michaille, J.J.; Croce, C.M. MicroRNAs play a central role in molecular dysfunctions linking inflammation with cancer. Immunol. Rev. 2013, 253, 167–184. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, A.; Calin, G.A.; Fabbri, M.; Iorio, M.V.; Ferracin, M.; Shimizu, M.; Wojcik, S.E.; Aqeilan, R.I.; Zupo, S.; Dono, M.; et al. miR-15 and miR-16 induce apoptosis by targeting BCL2. Proc. Natl. Acad. Sci. USA 2005, 102, 13944–13949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salem, M.L.; Zidan, A.A.; El-Naggar, R.E.E.-D.; Saad, M.A.; El-Shanshory, M.; Bakry, U.; Zidan, M. Myeloid-derived suppressor cells and regulatory T cells share common immunoregulatory pathways-related microRNAs that are dysregulated by acute lymphoblastic leukemia and chemotherapy. Hum. Immunol. 2021, 82, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Qiu, Y.; Qiu, Z.; Qu, P. MicroRNA networks regulate the differentiation, expansion and suppression function of myeloid-derived suppressor cells in tumor microenvironment. J. Cancer 2019, 10, 4350–4356. [Google Scholar] [CrossRef] [PubMed]
- Nagel, S.; Venturini, L.; Przybylski, G.K.; Grabarczyk, P.; Schmidt, C.A.; Meyer, C.; Drexler, H.G.; Macleod, R.A.; Scherr, M. Activation of miR-17-92 by NK-like homeodomain proteins suppresses apoptosis via reduction of E2F1 in T-cell acute lymphoblastic leukemia. Leuk Lymphoma 2009, 50, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Regelin, M.; Blume, J.; Pommerencke, J.; Vakilzadeh, R.; Witzlau, K.; Lyszkiewicz, M.; Zietara, N.; Saran, N.; Schambach, A.; Krueger, A. Responsiveness of Developing T Cells to IL-7 Signals Is Sustained by miR-17 approximately 92. J. Immunol. 2015, 195, 4832–4840. [Google Scholar] [CrossRef] [Green Version]
- Tsitsiou, E.; Lindsay, M.A. microRNAs and the immune response. Curr. Opin. Pharmacol. 2009, 9, 514–520. [Google Scholar] [CrossRef] [PubMed]
- El-Maadawy, E.A.; Elshal, M.F.; Bakry, R.M.; Moussa, M.M.; El-Naby, S.; Talaat, R.M. Regulation of CD4(+)CD25(+)FOXP3(+) cells in Pediatric Acute Lymphoblastic Leukemia (ALL): Implication of cytokines and miRNAs. Mol. Immunol. 2020, 124, 1–8. [Google Scholar] [CrossRef]
- Junker, F.; Chabloz, A.; Koch, U.; Radtke, F. Dicer1 imparts essential survival cues in Notch-driven T-ALL via miR-21-mediated tumor suppressor Pdcd4 repression. Blood 2015, 126, 993–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinn, S.R.; O’Neill, L.A. The role of microRNAs in the control and mechanism of action of IL-10. Curr. Top. Microbiol. Immunol. 2014, 380, 145–155. [Google Scholar] [CrossRef]
- Yuan, T.; Yang, Y.; Chen, J.; Li, W.; Li, W.; Zhang, Q.; Mi, Y.; Goswami, R.S.; You, J.Q.; Lin, D.; et al. Regulation of PI3K signaling in T-cell acute lymphoblastic leukemia: A novel PTEN/Ikaros/miR-26b mechanism reveals a critical targetable role for PIK3CD. Leukemia 2017, 31, 2355–2364. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Tian, A.; Wang, J.; Shen, X.; Qi, G.; Tang, Y. miR26a modulates Th17/T reg balance in the EAE model of multiple sclerosis by targeting IL6. Neuromolecular Med. 2015, 17, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Huan, J.; Hornick, N.I.; Shurtleff, M.J.; Skinner, A.M.; Goloviznina, N.A.; Roberts, C.T., Jr.; Kurre, P. RNA trafficking by acute myelogenous leukemia exosomes. Cancer Res. 2013, 73, 918–929. [Google Scholar] [CrossRef] [Green Version]
- Jin, F.; Du, Z.; Tang, Y.; Wang, L.; Yang, Y. Impact of microRNA-29b on natural killer cells in T-cell acute lymphoblastic leukemia. Oncol. Lett. 2019, 18, 2394–2403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lion, E.; Willemen, Y.; Berneman, Z.N.; Van Tendeloo, V.F.; Smits, E.L. Natural killer cell immune escape in acute myeloid leukemia. Leukemia 2012, 26, 2019–2026. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, L.H.; Schiavinato, J.L.; Fraguas, M.S.; Lucena-Araujo, A.R.; Haddad, R.; Araujo, A.G.; Dalmazzo, L.F.; Rego, E.M.; Covas, D.T.; Zago, M.A.; et al. Potential roles of microRNA-29a in the molecular pathophysiology of T-cell acute lymphoblastic leukemia. Cancer Sci. 2015, 106, 1264–1277. [Google Scholar] [CrossRef]
- Hart, M.; Rheinheimer, S.; Leidinger, P.; Backes, C.; Menegatti, J.; Fehlmann, T.; Grasser, F.; Keller, A.; Meese, E. Identification of miR-34a-target interactions by a combined network based and experimental approach. Oncotarget 2016, 7, 34288–34299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hart, M.; Walch-Ruckheim, B.; Friedmann, K.S.; Rheinheimer, S.; Tanzer, T.; Glombitza, B.; Sester, M.; Lenhof, H.P.; Hoth, M.; Schwarz, E.C.; et al. miR-34a: A new player in the regulation of T cell function by modulation of NF-kappaB signaling. Cell Death Dis. 2019, 10, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pyzer, A.R.; Stroopinsky, D.; Rajabi, H.; Washington, A.; Tagde, A.; Coll, M.; Fung, J.; Bryant, M.P.; Cole, L.; Palmer, K.; et al. MUC1-mediated induction of myeloid-derived suppressor cells in patients with acute myeloid leukemia. Blood 2017, 129, 1791–1801. [Google Scholar] [CrossRef] [Green Version]
- Podshivalova, K.; Salomon, D.R. MicroRNA regulation of T-lymphocyte immunity: Modulation of molecular networks responsible for T-cell activation, differentiation, and development. Crit. Rev. Immunol. 2013, 33, 435–476. [Google Scholar] [CrossRef]
- Lv, M.; Zhang, X.; Jia, H.; Li, D.; Zhang, B.; Zhang, H.; Hong, M.; Jiang, T.; Jiang, Q.; Lu, J.; et al. An oncogenic role of miR-142-3p in human T-cell acute lymphoblastic leukemia (T-ALL) by targeting glucocorticoid receptor-alpha and cAMP/PKA pathways. Leukemia 2012, 26, 769–777. [Google Scholar] [CrossRef] [Green Version]
- Correia, N.C.; Fragoso, R.; Carvalho, T.; Enguita, F.J.; Barata, J.T. MiR-146b negatively regulates migration and delays progression of T-cell acute lymphoblastic leukemia. Sci. Rep. 2016, 6, 31894. [Google Scholar] [CrossRef]
- Ghisi, M.; Corradin, A.; Basso, K.; Frasson, C.; Serafin, V.; Mukherjee, S.; Mussolin, L.; Ruggero, K.; Bonanno, L.; Guffanti, A.; et al. Modulation of microRNA expression in human T-cell development: Targeting of NOTCH3 by miR-150. Blood 2011, 117, 7053–7062. [Google Scholar] [CrossRef]
- Saultz, J.N.; Freud, A.G.; Mundy-Bosse, B.L. MicroRNA regulation of natural killer cell development and function in leukemia. Mol. Immunol. 2019, 115, 12–20. [Google Scholar] [CrossRef]
- Warth, S.C.; Hoefig, K.P.; Hiekel, A.; Schallenberg, S.; Jovanovic, K.; Klein, L.; Kretschmer, K.; Ansel, K.M.; Heissmeyer, V. Induced miR-99a expression represses Mtor cooperatively with miR-150 to promote regulatory T-cell differentiation. EMBO J. 2015, 34, 1195–1213. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Zhang, J.; Diao, W.; Wang, D.; Wei, Y.; Zhang, C.Y.; Zen, K. MicroRNA-155 and MicroRNA-21 promote the expansion of functional myeloid-derived suppressor cells. J. Immunol. 2014, 192, 1034–1043. [Google Scholar] [CrossRef]
- Liu, C.; Li, N.; Liu, G. The Role of MicroRNAs in Regulatory T Cells. J. Immunol. Res. 2020, 2020, 3232061. [Google Scholar] [CrossRef]
- O’Connell, R.M.; Kahn, D.; Gibson, W.S.; Round, J.L.; Scholz, R.L.; Chaudhuri, A.A.; Kahn, M.E.; Rao, D.S.; Baltimore, D. MicroRNA-155 promotes autoimmune inflammation by enhancing inflammatory T cell development. Immunity 2010, 33, 607–619. [Google Scholar] [CrossRef] [Green Version]
- Thai, T.H.; Calado, D.P.; Casola, S.; Ansel, K.M.; Xiao, C.; Xue, Y.; Murphy, A.; Frendewey, D.; Valenzuela, D.; Kutok, J.L.; et al. Regulation of the germinal center response by microRNA-155. Science 2007, 316, 604–608. [Google Scholar] [CrossRef]
- Trotta, R.; Chen, L.; Ciarlariello, D.; Josyula, S.; Mao, C.; Costinean, S.; Yu, L.; Butchar, J.P.; Tridandapani, S.; Croce, C.M.; et al. miR-155 regulates IFN-gamma production in natural killer cells. Blood 2012, 119, 3478–3485. [Google Scholar] [CrossRef] [Green Version]
- Vigorito, E.; Perks, K.L.; Abreu-Goodger, C.; Bunting, S.; Xiang, Z.; Kohlhaas, S.; Das, P.P.; Miska, E.A.; Rodriguez, A.; Bradley, A.; et al. microRNA-155 regulates the generation of immunoglobulin class-switched plasma cells. Immunity 2007, 27, 847–859. [Google Scholar] [CrossRef] [Green Version]
- Yao, R.; Ma, Y.L.; Liang, W.; Li, H.H.; Ma, Z.J.; Yu, X.; Liao, Y.H. MicroRNA-155 modulates Treg and Th17 cells differentiation and Th17 cell function by targeting SOCS1. PLoS ONE 2012, 7, e46082. [Google Scholar] [CrossRef] [Green Version]
- Cichocki, F.; Felices, M.; McCullar, V.; Presnell, S.R.; Al-Attar, A.; Lutz, C.T.; Miller, J.S. Cutting edge: microRNA-181 promotes human NK cell development by regulating Notch signaling. J. Immunol. 2011, 187, 6171–6175. [Google Scholar] [CrossRef] [Green Version]
- Nabhan, M.; Louka, M.L.; Khairy, E.; Tash, F.; Ali-Labib, R.; El-Habashy, S. MicroRNA-181a and its target Smad 7 as potential biomarkers for tracking child acute lymphoblastic leukemia. Gene 2017, 628, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Wallaert, A.; Van Loocke, W.; Hernandez, L.; Taghon, T.; Speleman, F.; Van Vlierberghe, P. Comprehensive miRNA expression profiling in human T-cell acute lymphoblastic leukemia by small RNA-sequencing. Sci. Rep. 2017, 7, 7901. [Google Scholar] [CrossRef] [Green Version]
- Mei, Y.; Gao, C.; Wang, K.; Cui, L.; Li, W.; Zhao, X.; Liu, F.; Wu, M.; Deng, G.; Ding, W.; et al. Effect of microRNA-210 on prognosis and response to chemotherapeutic drugs in pediatric acute lymphoblastic leukemia. Cancer Sci. 2014, 105, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Noman, M.Z.; Janji, B.; Hu, S.; Wu, J.C.; Martelli, F.; Bronte, V.; Chouaib, S. Tumor-Promoting Effects of Myeloid-Derived Suppressor Cells Are Potentiated by Hypoxia-Induced Expression of miR-210. Cancer Res. 2015, 75, 3771–3787. [Google Scholar] [CrossRef] [Green Version]
- Begley, C.G.; Aplan, P.D.; Davey, M.P.; Nakahara, K.; Tchorz, K.; Kurtzberg, J.; Hershfield, M.S.; Haynes, B.F.; Cohen, D.I.; Waldmann, T.A.; et al. Chromosomal translocation in a human leukemic stem-cell line disrupts the T-cell antigen receptor delta-chain diversity region and results in a previously unreported fusion transcript. Proc. Natl. Acad. Sci. USA 1989, 86, 2031–2035. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Palermo, R.; Talora, C.; Campese, A.F.; Checquolo, S.; Bellavia, D.; Tottone, L.; Testa, G.; Miele, E.; Indraccolo, S.; et al. Notch and NF-kB signaling pathways regulate miR-223/FBXW7 axis in T-cell acute lymphoblastic leukemia. Leukemia 2014, 28, 2324–2335. [Google Scholar] [CrossRef]
- Shu, Y.; Wang, Y.; Lv, W.Q.; Peng, D.Y.; Li, J.; Zhang, H.; Jiang, G.J.; Yang, B.J.; Liu, S.; Zhang, J.; et al. ARRB1-Promoted NOTCH1 Degradation Is Suppressed by OncomiR miR-223 in T-cell Acute Lymphoblastic Leukemia. Cancer Res. 2020, 80, 988–998. [Google Scholar] [CrossRef] [PubMed]
- Ye, F. MicroRNA expression and activity in T-cell acute lymphoblastic leukemia. Oncotarget 2018, 9, 5445–5458. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, F.; Zhang, S.; Xu, X. MicroRNA-325 inhibits the proliferation and induces the apoptosis of T cell acute lymphoblastic leukemia cells in a BAG2-dependent manner. Exp. Ther. Med. 2021, 21, 631. [Google Scholar] [CrossRef]
- Li, W.Y.; Chen, X.M.; Xiong, W.; Guo, D.M.; Lu, L.; Li, H.Y. Detection of microvesicle miRNA expression in ALL subtypes and analysis of their functional roles. J. Huazhong Univ. Sci. Technol. Med. Sci. 2014, 34, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Ni, F.; Guo, C.; Sun, R.; Fu, B.; Yang, Y.; Wu, L.; Ren, S.; Tian, Z.; Wei, H. MicroRNA transcriptomes of distinct human NK cell populations identify miR-362-5p as an essential regulator of NK cell function. Sci. Rep. 2015, 5, 9993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mavrakis, K.J.; Van Der Meulen, J.; Wolfe, A.L.; Liu, X.; Mets, E.; Taghon, T.; Khan, A.A.; Setty, M.; Rondou, P.; Vandenberghe, P.; et al. A cooperative microRNA-tumor suppressor gene network in acute T-cell lymphoblastic leukemia (T-ALL). Nat. Genet. 2011, 43, 673–678. [Google Scholar] [CrossRef]
- Chiaretti, S.; Vitale, A.; Cazzaniga, G.; Orlando, S.M.; Silvestri, D.; Fazi, P.; Valsecchi, M.G.; Elia, L.; Testi, A.M.; Mancini, F.; et al. Clinico-biological features of 5202 patients with acute lymphoblastic leukemia enrolled in the Italian AIEOP and GIMEMA protocols and stratified in age cohorts. Haematologica 2013, 98, 1702–1710. [Google Scholar] [CrossRef]
- Guruharsha, K.G.; Kankel, M.W.; Artavanis-Tsakonas, S. The Notch signalling system: Recent insights into the complexity of a conserved pathway. Nat. Rev. Genet. 2012, 13, 654–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giaimo, B.D.; Borggrefe, T. Introduction to Molecular Mechanisms in Notch Signal Transduction and Disease Pathogenesis. Adv. Exp. Med. Biol. 2018, 1066, 3–30. [Google Scholar] [CrossRef] [PubMed]
- Zema, S.; Pelullo, M.; Nardozza, F.; Felli, M.P.; Screpanti, I.; Bellavia, D. A Dynamic Role of Mastermind-Like 1: A Journey Through the Main (Path)ways Between Development and Cancer. Front. Cell Dev. Biol. 2020, 8, 613557. [Google Scholar] [CrossRef] [PubMed]
- Tsaouli, G.; Barbarulo, A.; Vacca, A.; Screpanti, I.; Felli, M.P. Molecular Mechanisms of Notch Signaling in Lymphoid Cell Lineages Development: NF-kappaB and Beyond. Adv. Exp. Med. Biol. 2020, 1227, 145–164. [Google Scholar] [CrossRef] [PubMed]
- Gleimer, M.; von Boehmer, H. Thymocyte selection: Chemokine signaling is not only about the destination. Curr. Biol. 2010, 20, R316–R318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felli, M.P.; Maroder, M.; Mitsiadis, T.A.; Campese, A.F.; Bellavia, D.; Vacca, A.; Mann, R.S.; Frati, L.; Lendahl, U.; Gulino, A.; et al. Expression pattern of notch1, 2 and 3 and Jagged1 and 2 in lymphoid and stromal thymus components: Distinct ligand-receptor interactions in intrathymic T cell development. Int. Immunol. 1999, 11, 1017–1025. [Google Scholar] [CrossRef] [Green Version]
- Yashiro-Ohtani, Y.; Ohtani, T.; Pear, W.S. Notch regulation of early thymocyte development. Semin. Immunol. 2010, 22, 261–269. [Google Scholar] [CrossRef]
- Kannan, S.; Aitken, M.J.L.; Herbrich, S.M.; Golfman, L.S.; Hall, M.G.; Mak, D.H.; Burks, J.K.; Song, G.; Konopleva, M.; Mullighan, C.G.; et al. Antileukemia Effects of Notch-Mediated Inhibition of Oncogenic PLK1 in B-Cell Acute Lymphoblastic Leukemia. Mol. Cancer Ther. 2019, 18, 1615–1627. [Google Scholar] [CrossRef]
- Girardi, T.; Vicente, C.; Cools, J.; De Keersmaecker, K. The genetics and molecular biology of T-ALL. Blood 2017, 129, 1113–1123. [Google Scholar] [CrossRef]
- Bellavia, D.; Campese, A.F.; Alesse, E.; Vacca, A.; Felli, M.P.; Balestri, A.; Stoppacciaro, A.; Tiveron, C.; Tatangelo, L.; Giovarelli, M.; et al. Constitutive activation of NF-kappaB and T-cell leukemia/lymphoma in Notch3 transgenic mice. EMBO J. 2000, 19, 3337–3348. [Google Scholar] [CrossRef] [Green Version]
- Palomero, T.; Ferrando, A. Therapeutic targeting of NOTCH1 signaling in T-cell acute lymphoblastic leukemia. Clin. Lymphoma Myeloma 2009, 9, S205–S210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsaouli, G.; Ferretti, E.; Bellavia, D.; Vacca, A.; Felli, M.P. Notch/CXCR4 Partnership in Acute Lymphoblastic Leukemia Progression. J. Immunol. Res. 2019, 2019, 5601396. [Google Scholar] [CrossRef] [PubMed]
- Belver, L.; Ferrando, A. The genetics and mechanisms of T cell acute lymphoblastic leukaemia. Nat. Rev. Cancer 2016, 16, 494–507. [Google Scholar] [CrossRef] [PubMed]
- Bernasconi-Elias, P.; Hu, T.; Jenkins, D.; Firestone, B.; Gans, S.; Kurth, E.; Capodieci, P.; Deplazes-Lauber, J.; Petropoulos, K.; Thiel, P.; et al. Characterization of activating mutations of NOTCH3 in T-cell acute lymphoblastic leukemia and anti-leukemic activity of NOTCH3 inhibitory antibodies. Oncogene 2016, 35, 6077–6086. [Google Scholar] [CrossRef] [Green Version]
- Zhdanovskaya, N.; Firrincieli, M.; Lazzari, S.; Pace, E.; Rossi, P.S.; Felli, M.P.; Talora, C.; Screpanti, I.; Palermo, R. Targeting Notch to Maximize Chemotherapeutic Benefits: Rationale, Advanced Strategies, and Future Perspectives. Cancers 2021, 13, 5106. [Google Scholar] [CrossRef] [PubMed]
- Ferrandino, F.; Grazioli, P.; Bellavia, D.; Campese, A.F.; Screpanti, I.; Felli, M.P. Notch and NF-kappaB: Coach and Players of Regulatory T-Cell Response in Cancer. Front. Immunol. 2018, 9, 2165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastorczak, A.; Domka, K.; Fidyt, K.; Poprzeczko, M.; Firczuk, M. Mechanisms of Immune Evasion in Acute Lymphoblastic Leukemia. Cancers 2021, 13, 1536. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Galat, V.; Galat, Y.; Lee, Y.K.A.; Wainwright, D.; Wu, J. NK cell-based cancer immunotherapy: From basic biology to clinical development. J. Hematol. Oncol. 2021, 14, 7. [Google Scholar] [CrossRef]
- Moretta, L.; Montaldo, E.; Vacca, P.; Del Zotto, G.; Moretta, F.; Merli, P.; Locatelli, F.; Mingari, M.C. Human natural killer cells: Origin, receptors, function, and clinical applications. Int. Arch. Allergy Immunol. 2014, 164, 253–264. [Google Scholar] [CrossRef]
- Valenzuela-Vazquez, L.; Nunez-Enriquez, J.C.; Sanchez-Herrera, J.; Jimenez-Hernandez, E.; Martin-Trejo, J.A.; Espinoza-Hernandez, L.E.; Medina-Sanson, A.; Flores-Villegas, L.V.; Penaloza-Gonzalez, J.G.; Refugio Torres-Nava, J.; et al. Functional characterization of NK cells in Mexican pediatric patients with acute lymphoblastic leukemia: Report from the Mexican Interinstitutional Group for the Identification of the Causes of Childhood Leukemia. PLoS ONE 2020, 15, e0227314. [Google Scholar] [CrossRef]
- Duault, C.; Kumar, A.; Khani, A.T.; Lee, S.J.; Yang, L.; Huang, M.; Hurtz, C.; Manning, B.; Ghoda, L.; McDonald, T.; et al. Activated natural killer cells predict poor clinical prognosis in high-risk B- and T-cell acute lymphoblastic leukemia. Blood 2021, 138, 1465–1480. [Google Scholar] [CrossRef]
- Liu, L.L.; Beziat, V.; Oei, V.Y.S.; Pfefferle, A.; Schaffer, M.; Lehmann, S.; Hellstrom-Lindberg, E.; Soderhall, S.; Heyman, M.; Grander, D.; et al. Ex Vivo Expanded Adaptive NK Cells Effectively Kill Primary Acute Lymphoblastic Leukemia Cells. Cancer Immunol. Res. 2017, 5, 654–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Liu, S.; Zhang, B.; Qiao, L.; Zhang, Y.; Zhang, Y. T Cell Dysfunction and Exhaustion in Cancer. Front. Cell Dev. Biol. 2020, 8, 17. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.H.; Hwang, H.J.; Yoo, J.W.; Kim, H.; Choi, E.S.; Hwang, S.H.; Cho, Y.U.; Jang, S.; Park, C.J.; Im, H.J.; et al. Expression of Immune Checkpoint Receptors on T-Cells and Their Ligands on Leukemia Blasts in Childhood Acute Leukemia. Anticancer. Res. 2019, 39, 5531–5539. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Xu, J.; Liu, Q.; Li, J.; Xi, Y. Expression and significance of CD47, PD1 and PDL1 in T-cell acute lymphoblastic lymphoma/leukemia. Pathol. Res. Pract. 2019, 215, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Anand, P.; Guillaumet-Adkins, A.; Dimitrova, V.; Yun, H.; Drier, Y.; Sotudeh, N.; Rogers, A.; Ouseph, M.M.; Nair, M.; Potdar, S.; et al. Single-cell RNA-seq reveals developmental plasticity with coexisting oncogenic states and immune evasion programs in ETP-ALL. Blood 2021, 137, 2463–2480. [Google Scholar] [CrossRef] [PubMed]
- Bayon-Calderon, F.; Toribio, M.L.; Gonzalez-Garcia, S. Facts and Challenges in Immunotherapy for T-Cell Acute Lymphoblastic Leukemia. Int. J. Mol. Sci. 2020, 21, 7685. [Google Scholar] [CrossRef] [PubMed]
- Hornick, N.I.; Doron, B.; Abdelhamed, S.; Huan, J.; Harrington, C.A.; Shen, R.; Cambronne, X.A.; Verghese, S.C.; Kurre, P. AML suppresses hematopoiesis by releasing exosomes that contain microRNAs targeting c-MYB. Sci. Signal 2016, 9, ra88. [Google Scholar] [CrossRef] [Green Version]
- Yeh, Y.Y.; Ozer, H.G.; Lehman, A.M.; Maddocks, K.; Yu, L.; Johnson, A.J.; Byrd, J.C. Characterization of CLL exosomes reveals a distinct microRNA signature and enhanced secretion by activation of BCR signaling. Blood 2015, 125, 3297–3305. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, T.; Kono, K.; Mizukami, Y.; Kawaguchi, Y.; Mimura, K.; Watanabe, M.; Izawa, S.; Fujii, H. Distribution of Th17 cells and FoxP3(+) regulatory T cells in tumor-infiltrating lymphocytes, tumor-draining lymph nodes and peripheral blood lymphocytes in patients with gastric cancer. Cancer Sci. 2010, 101, 1947–1954. [Google Scholar] [CrossRef] [PubMed]
- Grazioli, P.; Orlando, A.; Giordano, N.; Noce, C.; Peruzzi, G.; Scafetta, G.; Screpanti, I.; Campese, A.F. NF-kappaB1 Regulates Immune Environment and Outcome of Notch-Dependent T-Cell Acute Lymphoblastic Leukemia. Front. Immunol. 2020, 11, 541. [Google Scholar] [CrossRef] [PubMed]
- Manlove, L.S.; Berquam-Vrieze, K.E.; Pauken, K.E.; Williams, R.T.; Jenkins, M.K.; Farrar, M.A. Adaptive Immunity to Leukemia Is Inhibited by Cross-Reactive Induced Regulatory T Cells. J. Immunol. 2015, 195, 4028–4037. [Google Scholar] [CrossRef] [Green Version]
- Niedzwiecki, M.; Budzilo, O.; Adamkiewicz-Drozynska, E.; Pawlik-Gwozdecka, D.; Zielinski, M.; Maciejka-Kemblowska, L.; Szczepanski, T.; Trzonkowski, P. CD4(+)CD25(high)CD127(low/-)FoxP3 (+) Regulatory T-Cell Population in Acute Leukemias: A Review of the Literature. J. Immunol. Res. 2019, 2019, 2816498. [Google Scholar] [CrossRef] [Green Version]
- Hossain, F.; Majumder, S.; Ucar, D.A.; Rodriguez, P.C.; Golde, T.E.; Minter, L.M.; Osborne, B.A.; Miele, L. Notch Signaling in Myeloid Cells as a Regulator of Tumor Immune Responses. Front. Immunol. 2018, 9, 1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Lopez, S.; Monsalve, E.M.; de Avila, R.M.J.; Gonzalez-Gomez, J.; de Leon, H.N.; Ruiz-Marcos, F.; Baladron, V.; Nueda, M.L.; Garcia-Leon, M.J.; Screpanti, I.; et al. NOTCH3 signaling is essential for NF-kappaB activation in TLR-activated macrophages. Sci. Rep. 2020, 10, 14839. [Google Scholar] [CrossRef] [PubMed]
- Du, T.; Zamore, P.D. MicroPrimer: The biogenesis and function of microRNA. Development 2005, 132, 4645–4652. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Lee, Y.; Yeom, K.H.; Kim, Y.K.; Jin, H.; Kim, V.N. The Drosha-DGCR8 complex in primary microRNA processing. Genes Dev. 2004, 18, 3016–3027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, R.; Qin, Y.; Macara, I.G.; Cullen, B.R. Exportin-5 mediates the nuclear export of pre-microRNAs and short hairpin RNAs. Genes Dev. 2003, 17, 3011–3016. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Segal, M.; Slack, F.J. Challenges identifying efficacious miRNA therapeutics for cancer. Expert Opin. Drug Discov. 2020, 15, 987–992. [Google Scholar] [CrossRef] [PubMed]
- Freud, A.G.; Caligiuri, M.A. Human natural killer cell development. Immunol. Rev. 2006, 214, 56–72. [Google Scholar] [CrossRef] [PubMed]
- Freud, A.G.; Keller, K.A.; Scoville, S.D.; Mundy-Bosse, B.L.; Cheng, S.; Youssef, Y.; Hughes, T.; Zhang, X.; Mo, X.; Porcu, P.; et al. NKp80 Defines a Critical Step during Human Natural Killer Cell Development. Cell Rep. 2016, 16, 379–391. [Google Scholar] [CrossRef] [Green Version]
- Freud, A.G.; Mundy-Bosse, B.L.; Yu, J.; Caligiuri, M.A. The Broad Spectrum of Human Natural Killer Cell Diversity. Immunity 2017, 47, 820–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caligiuri, M.A. Human natural killer cells. Blood 2008, 112, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Cimmino, A.; Fabbri, M.; Ferracin, M.; Wojcik, S.E.; Shimizu, M.; Taccioli, C.; Zanesi, N.; Garzon, R.; Aqeilan, R.I.; et al. MiR-15a and miR-16-1 cluster functions in human leukemia. Proc. Natl. Acad. Sci. USA 2008, 105, 5166–5171. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, R.P.; Leong, J.W.; Schneider, S.E.; Ireland, A.R.; Berrien-Elliott, M.M.; Singh, A.; Schappe, T.; Jewell, B.A.; Sexl, V.; Fehniger, T.A. MicroRNA-15/16 Antagonizes Myb To Control NK Cell Maturation. J. Immunol. 2015, 195, 2806–2817. [Google Scholar] [CrossRef] [Green Version]
- Bezman, N.A.; Chakraborty, T.; Bender, T.; Lanier, L.L. miR-150 regulates the development of NK and iNKT cells. J. Exp. Med. 2011, 208, 2717–2731. [Google Scholar] [CrossRef] [Green Version]
- Haraguchi, K.; Suzuki, T.; Koyama, N.; Kumano, K.; Nakahara, F.; Matsumoto, A.; Yokoyama, Y.; Sakata-Yanagimoto, M.; Masuda, S.; Takahashi, T.; et al. Notch activation induces the generation of functional NK cells from human cord blood CD34-positive cells devoid of IL-15. J. Immunol. 2009, 182, 6168–6178. [Google Scholar] [CrossRef]
- Yang, C.; Shen, C.; Feng, T.; Li, H. Noncoding RNA in NK cells. J. Leukoc. Biol. 2019, 105, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Starczynowski, D.T.; Morin, R.; McPherson, A.; Lam, J.; Chari, R.; Wegrzyn, J.; Kuchenbauer, F.; Hirst, M.; Tohyama, K.; Humphries, R.K.; et al. Genome-wide identification of human microRNAs located in leukemia-associated genomic alterations. Blood 2011, 117, 595–607. [Google Scholar] [CrossRef] [Green Version]
- Romanski, A.; Bug, G.; Becker, S.; Kampfmann, M.; Seifried, E.; Hoelzer, D.; Ottmann, O.G.; Tonn, T. Mechanisms of resistance to natural killer cell-mediated cytotoxicity in acute lymphoblastic leukemia. Exp. Hematol. 2005, 33, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Leong, J.W.; Sullivan, R.P.; Fehniger, T.A. microRNA management of NK-cell developmental and functional programs. Eur. J. Immunol. 2014, 44, 2862–2868. [Google Scholar] [CrossRef] [Green Version]
- Garzon, R.; Heaphy, C.E.; Havelange, V.; Fabbri, M.; Volinia, S.; Tsao, T.; Zanesi, N.; Kornblau, S.M.; Marcucci, G.; Calin, G.A.; et al. MicroRNA 29b functions in acute myeloid leukemia. Blood 2009, 114, 5331–5341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, S.M.; Xing, C.Y.; Chen, C.Q.; Lin, S.S.; Dong, P.H.; Yu, F.J. miR-15a and miR-16-1 inhibit the proliferation of leukemic cells by down-regulating WT1 protein level. J. Exp. Clin. Cancer Res. 2011, 30, 110. [Google Scholar] [CrossRef] [Green Version]
- Rouce, R.H.; Shaim, H.; Sekine, T.; Weber, G.; Ballard, B.; Ku, S.; Barese, C.; Murali, V.; Wu, M.F.; Liu, H.; et al. The TGF-beta/SMAD pathway is an important mechanism for NK cell immune evasion in childhood B-acute lymphoblastic leukemia. Leukemia 2016, 30, 800–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, S.; Dong, J.; Lu, L.F. Cell-intrinsic and -extrinsic roles of miRNAs in regulating T cell immunity. Immunol. Rev. 2021, 304, 126–140. [Google Scholar] [CrossRef]
- Tapeh, B.E.G.; Mosayyebi, B.; Samei, M.; Basmenj, B.H.; Mohammadi, A.; Alivand, M.R.; Hassanpour, P.; Solali, S. microRNAs involved in T-cell development, selection, activation, and hemostasis. J. Cell Physiol. 2020, 235, 8461–8471. [Google Scholar] [CrossRef]
- Hu, L.; Mao, L.; Liu, S.; Zhao, J.; Chen, C.; Guo, M.; He, Z.; Yang, J.; Xu, W.; Xu, L. Functional Role of MicroRNAs in Thymocyte Development. Int. Arch. Allergy Immunol. 2019, 178, 315–322. [Google Scholar] [CrossRef]
- Zhou, B.; Wang, S.; Mayr, C.; Bartel, D.P.; Lodish, H.F. MiR-150, a microRNA expressed in mature B and T cells, blocks early B cell development when expressed prematurely. Proc. Natl. Acad. Sci. USA 2007, 104, 7080–7085. [Google Scholar] [CrossRef] [Green Version]
- Ferrandino, F.; Bernardini, G.; Tsaouli, G.; Grazioli, P.; Campese, A.F.; Noce, C.; Ciuffetta, A.; Vacca, A.; Besharat, Z.M.; Bellavia, D.; et al. Intrathymic Notch3 and CXCR4 combinatorial interplay facilitates T-cell leukemia propagation. Oncogene 2018, 37, 6285–6298. [Google Scholar] [CrossRef]
- Moloudizargari, M.; Hekmatirad, S.; Mofarahe, Z.S.; Asghari, M.H. Exosomal microRNA panels as biomarkers for hematological malignancies. Curr. Probl. Cancer 2021, 45, 100726. [Google Scholar] [CrossRef] [PubMed]
- Mansour, M.R.; Sanda, T.; Lawton, L.N.; Li, X.; Kreslavsky, T.; Novina, C.D.; Brand, M.; Gutierrez, A.; Kelliher, M.A.; Jamieson, C.H.; et al. The TAL1 complex targets the FBXW7 tumor suppressor by activating miR-223 in human T cell acute lymphoblastic leukemia. J. Exp. Med. 2013, 210, 1545–1557. [Google Scholar] [CrossRef] [PubMed]
- Fragoso, R.; Mao, T.; Wang, S.; Schaffert, S.; Gong, X.; Yue, S.; Luong, R.; Min, H.; Yashiro-Ohtani, Y.; Davis, M.; et al. Modulating the strength and threshold of NOTCH oncogenic signals by mir-181a-1/b-1. PLoS Genet. 2012, 8, e1002855. [Google Scholar] [CrossRef]
- Drobna, M.; Szarzynska-Zawadzka, B.; Dawidowska, M. T-cell acute lymphoblastic leukemia from miRNA perspective: Basic concepts, experimental approaches, and potential biomarkers. Blood Rev. 2018, 32, 457–472. [Google Scholar] [CrossRef]
- Lorsbach, R.B.; Moore, J.; Mathew, S.; Raimondi, S.C.; Mukatira, S.T.; Downing, J.R. TET1, a member of a novel protein family, is fused to MLL in acute myeloid leukemia containing the t(10;11)(q22;q23). Leukemia 2003, 17, 637–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebarowska, K.; Mroczek, A.; Kowalczyk, J.R.; Lejman, M. MicroRNA as a Prognostic and Diagnostic Marker in T-Cell Acute Lymphoblastic Leukemia. Int. J. Mol. Sci. 2021, 22, 5317. [Google Scholar] [CrossRef] [PubMed]
- Fuziwara, C.S.; Kimura, E.T. Insights into Regulation of the miR-17-92 Cluster of miRNAs in Cancer. Front. Med. 2015, 2, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, Z.; Fang, Z.G.; Wu, J.Y.; Liu, J.J. The depletion of Circ-PRKDC enhances autophagy and apoptosis in T-cell acute lymphoblastic leukemia via microRNA-653-5p/Reelin mediation of the PI3K/AKT/mTOR signaling pathway. Kaohsiung J. Med. Sci. 2021, 37, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Correia, N.C.; Melao, A.; Povoa, V.; Sarmento, L.; de Cedron, G.M.; Malumbres, M.; Enguita, F.J.; Barata, J.T. MicroRNAs regulate TAL1 expression in T-cell acute lymphoblastic leukemia. Oncotarget 2016, 7, 8268–8281. [Google Scholar] [CrossRef] [Green Version]
- Hori, S.; Nomura, T.; Sakaguchi, S. Control of regulatory T cell development by the transcription factor Foxp3. Science 2003, 299, 1057–1061. [Google Scholar] [CrossRef] [Green Version]
- Cobb, B.S.; Hertweck, A.; Smith, J.; O’Connor, E.; Graf, D.; Cook, T.; Smale, S.T.; Sakaguchi, S.; Livesey, F.J.; Fisher, A.G.; et al. A role for Dicer in immune regulation. J. Exp. Med. 2006, 203, 2519–2527. [Google Scholar] [CrossRef]
- Haas, J.D.; Nistala, K.; Petermann, F.; Saran, N.; Chennupati, V.; Schmitz, S.; Korn, T.; Wedderburn, L.R.; Forster, R.; Krueger, A.; et al. Expression of miRNAs miR-133b and miR-206 in the Il17a/f locus is co-regulated with IL-17 production in alphabeta and gammadelta T cells. PLoS ONE 2011, 6, e20171. [Google Scholar] [CrossRef] [PubMed]
- Grazioli, P.; Felli, M.P.; Screpanti, I.; Campese, A.F. The mazy case of Notch and immunoregulatory cells. J. Leukoc. Biol. 2017, 102, 361–368. [Google Scholar] [CrossRef] [Green Version]
- Lv, M.; Wang, K.; Huang, X.J. Myeloid-derived suppressor cells in hematological malignancies: Friends or foes. J. Hematol. Oncol. 2019, 12, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baghbani, E.; Noorolyai, S.; Duijf, P.H.G.; Silvestris, N.; Kolahian, S.; Hashemzadeh, S.; Kojabad, B.A.; FallahVazirabad, A.; Baradaran, B. The impact of microRNAs on myeloid-derived suppressor cells in cancer. Hum. Immunol. 2021, 82, 668–678. [Google Scholar] [CrossRef]
- Hassan, N.M.; Refaat, L.A.; Ismail, G.N.; Abdellateif, M.; Fadel, S.A.; AbdelAziz, R.S. Diagnostic, prognostic and predictive values of miR-100 and miR-210 in pediatric acute lymphoblastic Leukemia. Hematology 2020, 25, 405–413. [Google Scholar] [CrossRef]
- Nair, R.A.; Verma, V.K.; Beevi, S.S.; Rawoof, A.; Alexander, L.E.; Prasad, E.R.; Kumari, P.K.; Kumar, P.; Kumar, D.L. MicroRNA Signatures in Blood or Bone Marrow Distinguish Subtypes of Pediatric Acute Lymphoblastic Leukemia. Transl. Oncol. 2020, 13, 100800. [Google Scholar] [CrossRef]
- Lv, M.; Zhu, S.; Peng, H.; Cheng, Z.; Zhang, G.; Wang, Z. B-cell acute lymphoblastic leukemia-related microRNAs: Uncovering their diverse and special roles. Am. J. Cancer Res. 2021, 11, 1104–1120. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Del Gaizo, M.; Sergio, I.; Lazzari, S.; Cialfi, S.; Pelullo, M.; Screpanti, I.; Felli, M.P. MicroRNAs as Modulators of the Immune Response in T-Cell Acute Lymphoblastic Leukemia. Int. J. Mol. Sci. 2022, 23, 829. https://doi.org/10.3390/ijms23020829
Del Gaizo M, Sergio I, Lazzari S, Cialfi S, Pelullo M, Screpanti I, Felli MP. MicroRNAs as Modulators of the Immune Response in T-Cell Acute Lymphoblastic Leukemia. International Journal of Molecular Sciences. 2022; 23(2):829. https://doi.org/10.3390/ijms23020829
Chicago/Turabian StyleDel Gaizo, Martina, Ilaria Sergio, Sara Lazzari, Samantha Cialfi, Maria Pelullo, Isabella Screpanti, and Maria Pia Felli. 2022. "MicroRNAs as Modulators of the Immune Response in T-Cell Acute Lymphoblastic Leukemia" International Journal of Molecular Sciences 23, no. 2: 829. https://doi.org/10.3390/ijms23020829
APA StyleDel Gaizo, M., Sergio, I., Lazzari, S., Cialfi, S., Pelullo, M., Screpanti, I., & Felli, M. P. (2022). MicroRNAs as Modulators of the Immune Response in T-Cell Acute Lymphoblastic Leukemia. International Journal of Molecular Sciences, 23(2), 829. https://doi.org/10.3390/ijms23020829