
A New Player in the Hippocampus: A Review on VGLUT3+ Neurons and Their Role in the Regulation of Hippocampal Activity and Behaviour

 ,
, 
Abstract
1. Introduction
2. Characterisation of VGLUT3
2.1. Anatomical Distribution of VGLUT3 in the Central Nervous System
2.2. Glutamate as a Secondary Neurotransmitter in VGLUT3+ Neurons
2.3. Electrophysiological Characteristics of VGLUT3
3. Implications of VGLUT3 in Physiology
4. Characteristics of the VGLUT3 KO Mice
5. The Hippocampus
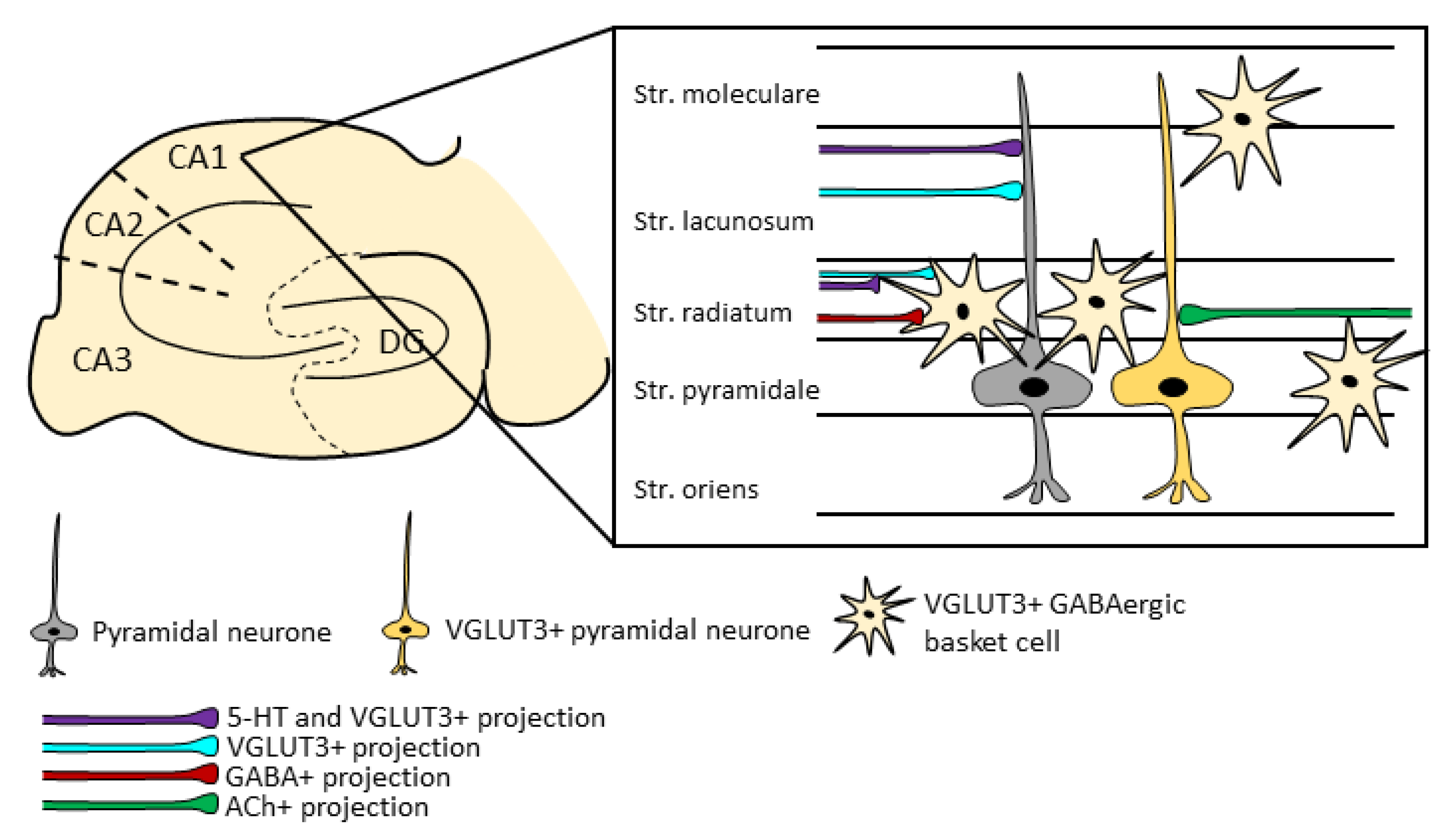
5.1. Characteristics of the VGLUT3+ Neurons in the Hippocampus
5.2. Hippocampal VGLUT3+ Projections
5.3. Role of the Hippocampal VGLUT3 Positivity
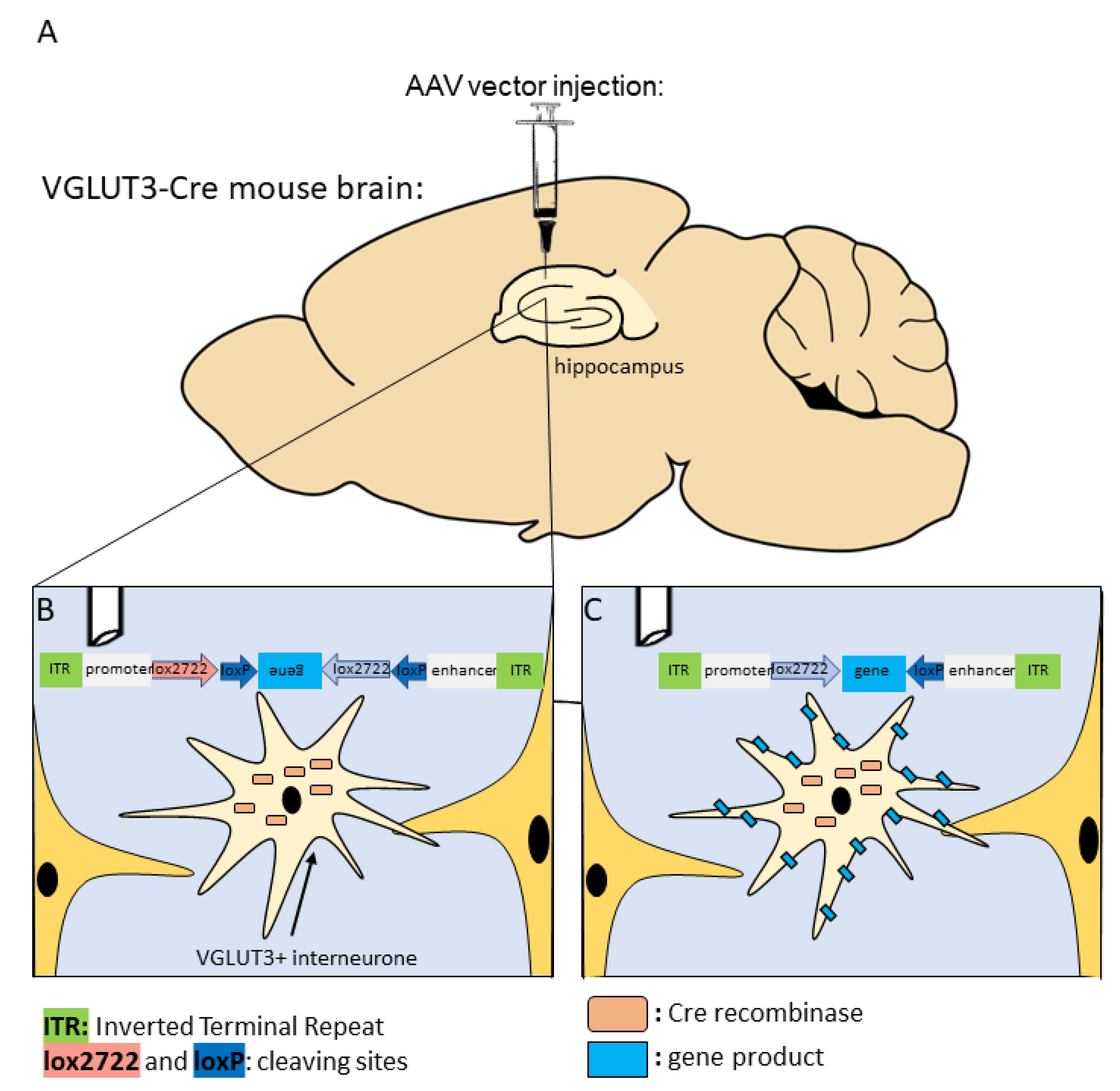
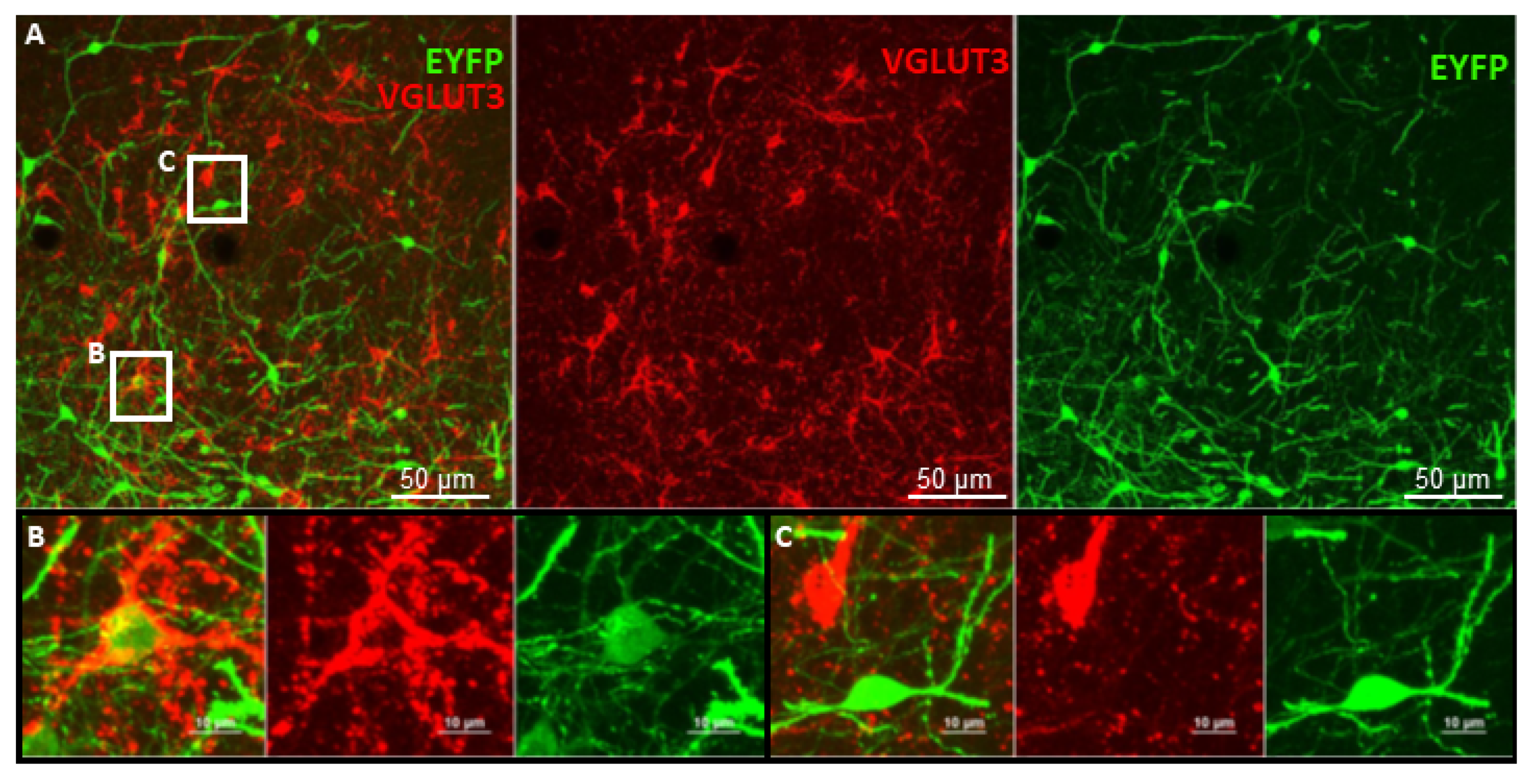
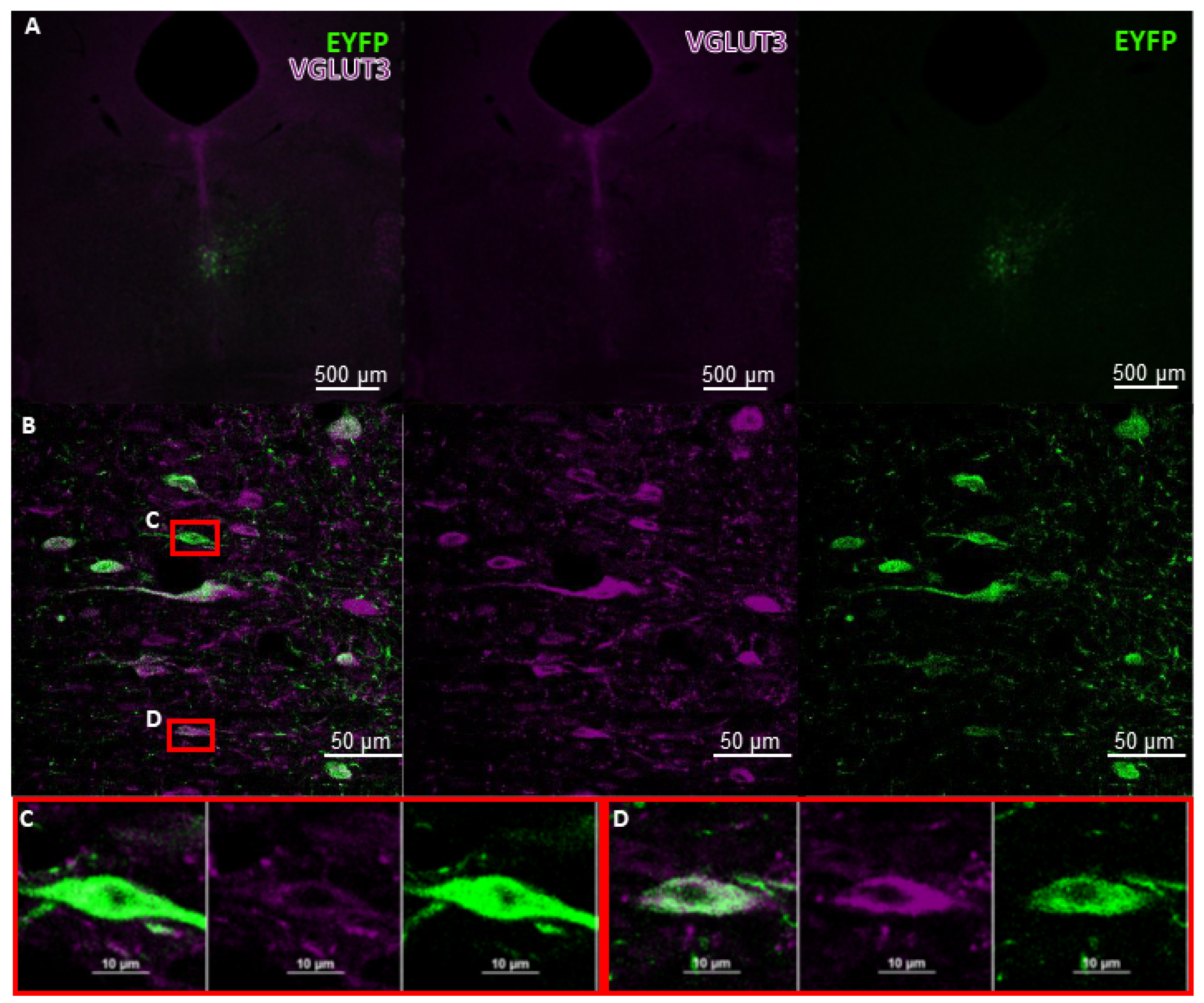
6. Future Perspectives for Selective Hippocampal VGLUT3+ Manipulations
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Burnstock, G. Do some nerve cells release more than one transmitter? Neuroscience 1976, 1, 239–248. [Google Scholar] [CrossRef]
- Takamori, S.; Rhee, J.S.; Rosenmund, C.; Jahn, R. Identification of a vesicular glutamate transporter that defines a glutamatergic phenotype in neurons. Nature 2000, 407, 189–194. [Google Scholar] [CrossRef]
- Robinson, M.B. Acute regulation of sodium-dependent glutamate transporters: A focus on constitutive and regulated trafficking. Neurotransm. Transp. 2006, 175, 251–275. [Google Scholar] [CrossRef]
- Preobraschenski, J.; Zander, J.F.; Suzuki, T.; Ahnert-Hilger, G.; Jahn, R. Vesicular glutamate transporters use flexible anion and cation binding sites for efficient accumulation of neurotransmitter. Neuron 2014, 84, 1287–1301. [Google Scholar] [CrossRef]
- Fremeau, R.T., Jr.; Burman, J.; Qureshi, T.; Tran, C.H.; Proctor, J.; Johnson, J.; Zhang, H.; Sulzer, D.; Copenhagen, D.R.; Storm-Mathisen, J.; et al. The identification of vesicular glutamate transporter 3 suggests novel modes of signaling by glutamate. Proc. Natl. Acad. Sci. USA 2002, 99, 14488–14493. [Google Scholar] [CrossRef]
- Schafer, M.K.; Varoqui, H.; Defamie, N.; Weihe, E.; Erickson, J.D. Molecular cloning and functional identification of mouse vesicular glutamate transporter 3 and its expression in subsets of novel excitatory neurons. J. Biol. Chem. 2002, 277, 50734–50748. [Google Scholar] [CrossRef] [PubMed]
- Wichmann, C.; Kuner, T. Heterogeneity of glutamatergic synapses: Cellular mechanisms and network consequences. Physiol. Rev. 2022, 102, 269–318. [Google Scholar] [CrossRef] [PubMed]
- Weston, M.C.; Nehring, R.B.; Wojcik, S.M.; Rosenmund, C. Interplay between VGLUT isoforms and endophilin A1 regulates neurotransmitter release and short-term plasticity. Neuron 2011, 69, 1147–1159. [Google Scholar] [CrossRef] [PubMed]
- Fremeau, R.T., Jr.; Voglmaier, S.; Seal, R.P.; Edwards, R.H. VGLUTs define subsets of excitatory neurons and suggest novel roles for glutamate. Trends Neurosci. 2004, 27, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Hioki, H.; Fujiyama, F.; Taki, K.; Tomioka, R.; Furuta, T.; Tamamaki, N.; Kaneko, T. Differential distribution of vesicular glutamate transporters in the rat cerebellar cortex. Neuroscience 2003, 117, 1–6. [Google Scholar] [CrossRef]
- Vigneault, E.; Poirel, O.; Riad, M.; Prud’homme, J.; Dumas, S.; Turecki, G.; Fasano, C.; Mechawar, N.; El Mestikawy, S. Distribution of vesicular glutamate transporters in the human brain. Front. Neuroanat. 2015, 9, 23. [Google Scholar] [CrossRef] [PubMed]
- Fremeau, R.T., Jr.; Troyer, M.D.; Pahner, I.; Nygaard, G.O.; Tran, C.H.; Reimer, R.J.; Bellocchio, E.E.; Fortin, D.; Storm-Mathisen, J.; Edwards, R.H. The expression of vesicular glutamate transporters defines two classes of excitatory synapse. Neuron 2001, 31, 247–260. [Google Scholar] [CrossRef]
- Herzog, E.; Gilchrist, J.; Gras, C.; Muzerelle, A.; Ravassard, P.; Giros, B.; Gaspar, P.; El Mestikawy, S. Localization of VGLUT3, the vesicular glutamate transporter type 3, in the rat brain. Neuroscience 2004, 123, 983–1002. [Google Scholar] [CrossRef] [PubMed]
- Szonyi, A.; Zicho, K.; Barth, A.M.; Gonczi, R.T.; Schlingloff, D.; Torok, B.; Sipos, E.; Major, A.; Bardoczi, Z.; Sos, K.E.; et al. Median raphe controls acquisition of negative experience in the mouse. Science 2019, 366, 6469:eaay8746. [Google Scholar] [CrossRef]
- Bai, L.; Xu, H.; Collins, J.F.; Ghishan, F.K. Molecular and functional analysis of a novel neuronal vesicular glutamate transporter. J. Biol. Chem. 2001, 276, 36764–36769. [Google Scholar] [CrossRef] [PubMed]
- Gras, C.; Herzog, E.; Bellenchi, G.C.; Bernard, V.; Ravassard, P.; Pohl, M.; Gasnier, B.; Giros, B.; El Mestikawy, S. A third vesicular glutamate transporter expressed by cholinergic and serotoninergic neurons. J. Neurosci. 2002, 22, 5442–5451. [Google Scholar] [CrossRef]
- Takamori, S.; Malherbe, P.; Broger, C.; Jahn, R. Molecular cloning and functional characterization of human vesicular glutamate transporter 3. EMBO Rep. 2002, 3, 798–803. [Google Scholar] [CrossRef] [PubMed]
- Harkany, T.; Hartig, W.; Berghuis, P.; Dobszay, M.B.; Zilberter, Y.; Edwards, R.H.; Mackie, K.; Ernfors, P. Complementary distribution of type 1 cannabinoid receptors and vesicular glutamate transporter 3 in basal forebrain suggests input-specific retrograde signalling by cholinergic neurons. Eur. J. Neurosci. 2003, 18, 1979–1992. [Google Scholar] [CrossRef] [PubMed]
- Hioki, H.; Nakamura, H.; Ma, Y.F.; Konno, M.; Hayakawa, T.; Nakamura, K.C.; Fujiyama, F.; Kaneko, T. Vesicular glutamate transporter 3-expressing nonserotonergic projection neurons constitute a subregion in the rat midbrain raphe nuclei. J. Comp. Neurol. 2010, 518, 668–686. [Google Scholar] [CrossRef]
- Hioki, H.; Fujiyama, F.; Nakamura, K.; Wu, S.X.; Matsuda, W.; Kaneko, T. Chemically specific circuit composed of vesicular glutamate transporter 3- and preprotachykinin B-producing interneurons in the rat neocortex. Cereb. Cortex. 2004, 14, 1266–1275. [Google Scholar] [CrossRef]
- Kudo, T.; Uchigashima, M.; Miyazaki, T.; Konno, K.; Yamasaki, M.; Yanagawa, Y.; Minami, M.; Watanabe, M. Three types of neurochemical projection from the bed nucleus of the stria terminalis to the ventral tegmental area in adult mice. J. Neurosci. 2012, 32, 18035–18046. [Google Scholar] [CrossRef]
- Stornetta, R.L.; Rosin, D.L.; Simmons, J.R.; McQuiston, T.J.; Vujovic, N.; Weston, M.C.; Guyenet, P.G. Coexpression of vesicular glutamate transporter-3 and gamma-aminobutyric acidergic markers in rat rostral medullary raphe and intermediolateral cell column. J. Comp. Neurol. 2005, 492, 477–494. [Google Scholar] [CrossRef]
- Somogyi, J.; Baude, A.; Omori, Y.; Shimizu, H.; El Mestikawy, S.; Fukaya, M.; Shigemoto, R.; Watanabe, M.; Somogyi, P. GABAergic basket cells expressing cholecystokinin contain vesicular glutamate transporter type 3 (VGLUT3) in their synaptic terminals in hippocampus and isocortex of the rat. Eur. J. Neurosci. 2004, 19, 552–569. [Google Scholar] [CrossRef]
- Harkany, T.; Holmgren, C.; Hartig, W.; Qureshi, T.; Chaudhry, F.A.; Storm-Mathisen, J.; Dobszay, M.B.; Berghuis, P.; Schulte, G.; Sousa, K.M.; et al. Endocannabinoid-independent retrograde signaling at inhibitory synapses in layer 2/3 of neocortex: Involvement of vesicular glutamate transporter 3. J. Neurosci. 2004, 24, 4978–4988. [Google Scholar] [CrossRef]
- Miot, S.; Voituron, N.; Sterlin, A.; Vigneault, E.; Morel, L.; Matrot, B.; Ramanantsoa, N.; Amilhon, B.; Poirel, O.; Lepicard, E.; et al. The vesicular glutamate transporter VGLUT3 contributes to protection against neonatal hypoxic stress. J. Physiol. 2012, 590, 5183–5198. [Google Scholar] [CrossRef] [PubMed]
- Tatti, R.; Bhaukaurally, K.; Gschwend, O.; Seal, R.P.; Edwards, R.H.; Rodriguez, I.; Carleton, A. A population of glomerular glutamatergic neurons controls sensory information transfer in the mouse olfactory bulb. Nat. Commun. 2014, 5, 3791. [Google Scholar] [CrossRef]
- Rovira-Esteban, L.; Peterfi, Z.; Vikor, A.; Mate, Z.; Szabo, G.; Hajos, N. Morphological and physiological properties of CCK/CB1R-expressing interneurons in the basal amygdala. Brain Struct. Funct. 2017, 222, 3543–3565. [Google Scholar] [CrossRef] [PubMed]
- Ormel, L.; Stensrud, M.J.; Chaudhry, F.A.; Gundersen, V. A distinct set of synaptic-like microvesicles in atroglial cells contain VGLUT3. Glia 2012, 60, 1289–1300. [Google Scholar] [CrossRef]
- Li, D.; Herault, K.; Silm, K.; Evrard, A.; Wojcik, S.; Oheim, M.; Herzog, E.; Ropert, N. Lack of evidence for vesicular glutamate transporter expression in mouse astrocytes. J. Neurosci. 2013, 33, 4434–4455. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Matsumura, K.; Kobayashi, S.; Kaneko, T. Sympathetic premotor neurons mediating thermoregulatory functions. Neurosci. Res. 2005, 51, 1–8. [Google Scholar] [CrossRef]
- Oliveira, A.L.; Hydling, F.; Olsson, E.; Shi, T.; Edwards, R.H.; Fujiyama, F.; Kaneko, T.; Hokfelt, T.; Cullheim, S.; Meister, B. Cellular localization of three vesicular glutamate transporter mRNAs and proteins in rat spinal cord and dorsal root ganglia. Synapse 2003, 50, 117–129. [Google Scholar] [CrossRef]
- Rosin, D.L.; Chang, D.A.; Guyenet, P.G. Afferent and efferent connections of the rat retrotrapezoid nucleus. J. Comp. Neurol. 2006, 499, 64–89. [Google Scholar] [CrossRef] [PubMed]
- Zerari-Mailly, F.; Braud, A.; Davido, N.; Toure, B.; Azerad, J.; Boucher, Y. Glutamate control of pulpal blood flow in the incisor dental pulp of the rat. Eur. J. Oral Sci. 2012, 120, 402–407. [Google Scholar] [CrossRef]
- Munguba, G.C.; Camp, A.S.; Risco, M.; Tapia, M.L.; Bhattacharya, S.K.; Lee, R.K. Vesicular glutamate transporter 3 in age-dependent optic neuropathy. Mol. Vis. 2011, 17, 413–419. [Google Scholar]
- Stensrud, M.J.; Chaudhry, F.A.; Leergaard, T.B.; Bjaalie, J.G.; Gundersen, V. Vesicular glutamate transporter-3 in the rodent brain: Vesicular colocalization with vesicular gamma-aminobutyric acid transporter. J. Comp. Neurol. 2013, 521, 3042–3056. [Google Scholar] [CrossRef]
- Del Pino, I.; Brotons-Mas, J.R.; Marques-Smith, A.; Marighetto, A.; Frick, A.; Marin, O.; Rico, B. Abnormal wiring of CCK(+) basket cells disrupts spatial information coding. Nat. Neurosci. 2017, 20, 784–792. [Google Scholar] [CrossRef]
- Stensrud, M.J.; Sogn, C.J.; Gundersen, V. Immunogold characteristics of VGLUT3-positive GABAergic nerve terminals suggest corelease of glutamate. J. Comp. Neurol. 2015, 523, 2698–2713. [Google Scholar] [CrossRef] [PubMed]
- Jalabert, M.; Aston-Jones, G.; Herzog, E.; Manzoni, O.; Georges, F. Role of the bed nucleus of the stria terminalis in the control of ventral tegmental area dopamine neurons. Prog. Neuropsychopharmacol. Biol. Psychiatry 2009, 33, 1336–1346. [Google Scholar] [CrossRef]
- Omiya, Y.; Uchigashima, M.; Konno, K.; Yamasaki, M.; Miyazaki, T.; Yoshida, T.; Kusumi, I.; Watanabe, M. VGluT3-expressing CCK-positive basket cells construct invaginating synapses enriched with endocannabinoid signaling proteins in particular cortical and cortex-like amygdaloid regions of mouse brains. J. Neurosci. 2015, 35, 4215–4228. [Google Scholar] [CrossRef]
- Case, D.T.; Burton, S.D.; Gedeon, J.Y.; Williams, S.G.; Urban, N.N.; Seal, R.P. Layer- and cell type-selective co-transmission by a basal forebrain cholinergic projection to the olfactory bulb. Nat. Commun. 2017, 8, 652. [Google Scholar] [CrossRef] [PubMed]
- Nickerson Poulin, A.; Guerci, A.; El Mestikawy, S.; Semba, K. Vesicular glutamate transporter 3 immunoreactivity is present in cholinergic basal forebrain neurons projecting to the basolateral amygdala in rat. J. Comp. Neurol. 2006, 498, 690–711. [Google Scholar] [CrossRef]
- Gras, C.; Amilhon, B.; Lepicard, E.M.; Poirel, O.; Vinatier, J.; Herbin, M.; Dumas, S.; Tzavara, E.T.; Wade, M.R.; Nomikos, G.G.; et al. The vesicular glutamate transporter VGLUT3 synergizes striatal acetylcholine tone. Nat. Neurosci. 2008, 11, 292–300. [Google Scholar] [CrossRef]
- Higley, M.J.; Gittis, A.H.; Oldenburg, I.A.; Balthasar, N.; Seal, R.P.; Edwards, R.H.; Lowell, B.B.; Kreitzer, A.C.; Sabatini, B.L. Cholinergic interneurons mediate fast VGluT3-dependent glutamatergic transmission in the striatum. PLoS ONE 2011, 6, e19155. [Google Scholar] [CrossRef]
- Nelson, A.B.; Bussert, T.G.; Kreitzer, A.C.; Seal, R.P. Striatal cholinergic neurotransmission requires VGLUT3. J. Neurosci. 2014, 34, 8772–8777. [Google Scholar] [CrossRef] [PubMed]
- Shutoh, F.; Ina, A.; Yoshida, S.; Konno, J.; Hisano, S. Two distinct subtypes of serotonergic fibers classified by co-expression with vesicular glutamate transporter 3 in rat forebrain. Neurosci. Lett. 2008, 432, 132–136. [Google Scholar] [CrossRef] [PubMed]
- Voisin, A.N.; Mnie-Filali, O.; Giguere, N.; Fortin, G.M.; Vigneault, E.; El Mestikawy, S.; Descarries, L.; Trudeau, L.E. Axonal segregation and role of the vesicular glutamate transporter VGLUT3 in serotonin neurons. Front. Neuroanat. 2016, 10, 39. [Google Scholar] [CrossRef] [PubMed]
- Calizo, L.H.; Akanwa, A.; Ma, X.; Pan, Y.Z.; Lemos, J.C.; Craige, C.; Heemstra, L.A.; Beck, S.G. Raphe serotonin neurons are not homogenous: Electrophysiological, morphological and neurochemical evidence. Neuropharmacology 2011, 61, 524–543. [Google Scholar] [CrossRef]
- Ren, J.; Friedmann, D.; Xiong, J.; Liu, C.D.; Ferguson, B.R.; Weerakkody, T.; DeLoach, K.E.; Ran, C.; Pun, A.; Sun, Y.; et al. Anatomically defined and functionally distinct dorsal raphe serotonin sub-systems. Cell 2018, 175, 472–487 e420. [Google Scholar] [CrossRef]
- Gagnon, D.; Parent, M. Distribution of VGLUT3 in highly collateralized axons from the rat dorsal raphe nucleus as revealed by single-neuron reconstructions. PLoS ONE 2014, 9, e87709. [Google Scholar] [CrossRef]
- Commons, K.G. Locally collateralizing glutamate neurons in the dorsal raphe nucleus responsive to substance P contain vesicular glutamate transporter 3 (VGLUT3). J. Chem. Neuroanat. 2009, 38, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Le Maitre, E.; Fabre, V.; Bernard, J.F.; David Xu, Z.Q.; Hokfelt, T. Chemical neuroanatomy of the dorsal raphe nucleus and adjacent structures of the mouse brain. J. Comp. Neurol. 2010, 518, 3464–3494. [Google Scholar] [CrossRef]
- Liu, Z.; Zhou, J.; Li, Y.; Hu, F.; Lu, Y.; Ma, M.; Feng, Q.; Zhang, J.E.; Wang, D.; Zeng, J.; et al. Dorsal raphe neurons signal reward through 5-HT and glutamate. Neuron 2014, 81, 1360–1374. [Google Scholar] [CrossRef]
- Zou, W.J.; Song, Y.L.; Wu, M.Y.; Chen, X.T.; You, Q.L.; Yang, Q.; Luo, Z.Y.; Huang, L.; Kong, Y.; Feng, J.; et al. A discrete serotonergic circuit regulates vulnerability to social stress. Nat. Commun. 2020, 11, 4218. [Google Scholar] [CrossRef]
- Wang, H.L.; Zhang, S.; Qi, J.; Wang, H.; Cachope, R.; Mejias-Aponte, C.A.; Gomez, J.A.; Mateo-Semidey, G.E.; Beaudoin, G.M.J.; Paladini, C.A.; et al. Dorsal raphe dual serotonin-glutamate neurons drive reward by establishing excitatory synapses on VTA mesoaccumbens dopamine neurons. Cell Rep. 2019, 26, 1128–1142.e7. [Google Scholar] [CrossRef]
- Sengupta, A.; Holmes, A. A Discrete dorsal raphe to basal amygdala 5-HT circuit calibrates aversive memory. Neuron 2019, 103, 489–505.e7. [Google Scholar] [CrossRef]
- Sengupta, A.; Bocchio, M.; Bannerman, D.M.; Sharp, T.; Capogna, M. Control of Amygdala Circuits by 5-HT Neurons via 5-HT and Glutamate Cotransmission. J. Neurosci. 2017, 37, 1785–1796. [Google Scholar] [CrossRef]
- Martin-Ibanez, R.; Jenstad, M.; Berghuis, P.; Edwards, R.H.; Hioki, H.; Kaneko, T.; Mulder, J.; Canals, J.M.; Ernfors, P.; Chaudhry, F.A.; et al. Vesicular glutamate transporter 3 (VGLUT3) identifies spatially segregated excitatory terminals in the rat substantia nigra. Eur. J. Neurosci. 2006, 23, 1063–1070. [Google Scholar] [CrossRef]
- Sos, K.E.; Mayer, M.I.; Cserep, C.; Takacs, F.S.; Szonyi, A.; Freund, T.F.; Nyiri, G. Cellular architecture and transmitter phenotypes of neurons of the mouse median raphe region. Brain Struct. Funct. 2017, 222, 287–299. [Google Scholar] [CrossRef]
- Belmer, A.; Beecher, K.; Jacques, A.; Patkar, O.L.; Sicherre, F.; Bartlett, S.E. Axonal Non-segregation of the vesicular glutamate transporter VGLUT3 within serotonergic projections in the mouse forebrain. Front. Cell Neurosci. 2019, 13, 193. [Google Scholar] [CrossRef]
- Gaspar, P.; Lillesaar, C. Probing the diversity of serotonin neurons. Philos. Trans. R Soc. Lond B Biol. Sci. 2012, 367, 2382–2394. [Google Scholar] [CrossRef]
- Senft, R.A.; Freret, M.E.; Sturrock, N.; Dymecki, S.M. Neurochemically and hodologically distinct ascending VGLUT3 versus serotonin subsystems comprise the r2-Pet1 median raphe. J. Neurosci. 2021, 41, 2581–2600. [Google Scholar] [CrossRef]
- Amilhon, B.; Lepicard, E.; Renoir, T.; Mongeau, R.; Popa, D.; Poirel, O.; Miot, S.; Gras, C.; Gardier, A.M.; Gallego, J.; et al. VGLUT3 (vesicular glutamate transporter type 3) contribution to the regulation of serotonergic transmission and anxiety. J. Neurosci. 2010, 30, 2198–2210. [Google Scholar] [CrossRef]
- El Mestikawy, S.; Wallen-Mackenzie, A.; Fortin, G.M.; Descarries, L.; Trudeau, L.E. From glutamate co-release to vesicular synergy: Vesicular glutamate transporters. Nat. Rev. Neurosci. 2011, 12, 204–216. [Google Scholar] [CrossRef]
- Fasano, C.; Rocchetti, J.; Pietrajtis, K.; Zander, J.F.; Manseau, F.; Sakae, D.Y.; Marcus-Sells, M.; Ramet, L.; Morel, L.J.; Carrel, D.; et al. Regulation of the hippocampal network by VGLUT3-positive CCK- GABAergic basket cells. Front. Cell Neurosci. 2017, 11, 140. [Google Scholar] [CrossRef]
- Pelkey, K.A.; Calvigioni, D.; Fang, C.; Vargish, G.; Ekins, T.; Auville, K.; Wester, J.C.; Lai, M.; Mackenzie-Gray Scott, C.; Yuan, X.; et al. Paradoxical network excitation by glutamate release from VGluT3(+) GABAergic interneurons. Elife 2020, 9, e51996. [Google Scholar] [CrossRef]
- Zimmermann, J.; Herman, M.A.; Rosenmund, C. Co-release of glutamate and GABA from single vesicles in GABAergic neurons exogenously expressing VGLUT3. Front. Synaptic Neurosci. 2015, 7, 16. [Google Scholar] [CrossRef]
- Cropper, E.C.; Jing, J.; Vilim, F.S.; Weiss, K.R. Peptide cotransmitters as dynamic, intrinsic modulators of network activity. Front. Neural Circ. 2018, 12, 78. [Google Scholar] [CrossRef]
- Nusbaum, M.P.; Blitz, D.M.; Marder, E. Functional consequences of neuropeptide and small-molecule co-transmission. Nat. Rev. Neurosci. 2017, 18, 389–403. [Google Scholar] [CrossRef]
- Larsson, M.; Broman, J. Synaptic Organization of VGLUT3 Expressing low-threshold mechanosensitive c fiber terminals in the rodent spinal cord. eNeuro 2019, 6, ENEURO.0007-19.2019. [Google Scholar] [CrossRef]
- Crepel, F.; Galante, M.; Habbas, S.; McLean, H.; Daniel, H. Role of the vesicular transporter VGLUT3 in retrograde release of glutamate by cerebellar Purkinje cells. J. Neurophysiol. 2011, 105, 1023–1032. [Google Scholar] [CrossRef]
- Fuzik, J.; Zeisel, A.; Mate, Z.; Calvigioni, D.; Yanagawa, Y.; Szabo, G.; Linnarsson, S.; Harkany, T. Integration of electrophysiological recordings with single-cell RNA-seq data identifies neuronal subtypes. Nat. Biotechnol. 2016, 34, 175–183. [Google Scholar] [CrossRef]
- Kohus, Z.; Kali, S.; Rovira-Esteban, L.; Schlingloff, D.; Papp, O.; Freund, T.F.; Hajos, N.; Gulyas, A.I. Properties and dynamics of inhibitory synaptic communication within the CA3 microcircuits of pyramidal cells and interneurons expressing parvalbumin or cholecystokinin. J. Physiol. 2016, 594, 3745–3774. [Google Scholar] [CrossRef]
- Grimes, W.N.; Seal, R.P.; Oesch, N.; Edwards, R.H.; Diamond, J.S. Genetic targeting and physiological features of VGLUT3+ amacrine cells. Vis. Neurosci. 2011, 28, 381–392. [Google Scholar] [CrossRef]
- Peng, Z.; Wang, G.P.; Zeng, R.; Guo, J.Y.; Chen, C.F.; Gong, S.S. Temporospatial expression and cellular localization of VGLUT3 in the rat cochlea. Brain Res. 2013, 1537, 100–110. [Google Scholar] [CrossRef]
- Obholzer, N.; Wolfson, S.; Trapani, J.G.; Mo, W.; Nechiporuk, A.; Busch-Nentwich, E.; Seiler, C.; Sidi, S.; Sollner, C.; Duncan, R.N.; et al. Vesicular glutamate transporter 3 is required for synaptic transmission in zebrafish hair cells. J. Neurosci. 2008, 28, 2110–2118. [Google Scholar] [CrossRef]
- Ruel, J.; Emery, S.; Nouvian, R.; Bersot, T.; Amilhon, B.; Van Rybroek, J.M.; Rebillard, G.; Lenoir, M.; Eybalin, M.; Delprat, B.; et al. Impairment of SLC17A8 encoding vesicular glutamate transporter-3, VGLUT3, underlies nonsyndromic deafness DFNA25 and inner hair cell dysfunction in null mice. Am. J. Hum. Genet. 2008, 83, 278–292. [Google Scholar] [CrossRef]
- Noh, J.; Seal, R.P.; Garver, J.A.; Edwards, R.H.; Kandler, K. Glutamate co-release at GABA/glycinergic synapses is crucial for the refinement of an inhibitory map. Nat. Neurosci. 2010, 13, 232–238. [Google Scholar] [CrossRef]
- Joshi, Y.; Petit, C.P.; Miot, S.; Guillet, M.; Sendin, G.; Bourien, J.; Wang, J.; Pujol, R.; El Mestikawy, S.; Puel, J.L.; et al. VGLUT3-p.A211V variant fuses stereocilia bundles and elongates synaptic ribbons. J. Physiol. 2021, 599, 5397–5416. [Google Scholar] [CrossRef]
- Seal, R.P.; Akil, O.; Yi, E.; Weber, C.M.; Grant, L.; Yoo, J.; Clause, A.; Kandler, K.; Noebels, J.L.; Glowatzki, E.; et al. Sensorineural deafness and seizures in mice lacking vesicular glutamate transporter 3. Neuron 2008, 57, 263–275. [Google Scholar] [CrossRef]
- Akil, O.; Seal, R.P.; Burke, K.; Wang, C.; Alemi, A.; During, M.; Edwards, R.H.; Lustig, L.R. Restoration of hearing in the VGLUT3 knockout mouse using virally mediated gene therapy. Neuron 2012, 75, 283–293. [Google Scholar] [CrossRef]
- Kim, K.X.; Payne, S.; Yang-Hood, A.; Li, S.Z.; Davis, B.; Carlquist, J.; Babak, V.-G.; Gantz, J.A.; Kallogjeri, D.; Fitzpatrick, J.A.J.; et al. Vesicular glutamatergic transmission in noise-induced loss and repair of cochlear ribbon synapses. J. Neurosci. 2019, 39, 4434–4447. [Google Scholar] [CrossRef]
- Honsek, S.D.; Seal, R.P.; Sandkuhler, J. Presynaptic inhibition of optogenetically identified VGluT3+ sensory fibres by opioids and baclofen. Pain 2015, 156, 243–251. [Google Scholar] [CrossRef]
- Lou, S.; Duan, B.; Vong, L.; Lowell, B.B.; Ma, Q. Runx1 controls terminal morphology and mechanosensitivity of VGLUT3-expressing C-mechanoreceptors. J. Neurosci. 2013, 33, 870–882. [Google Scholar] [CrossRef]
- Balazsfi, D.; Fodor, A.; Torok, B.; Ferenczi, S.; Kovacs, K.J.; Haller, J.; Zelena, D. Enhanced innate fear and altered stress axis regulation in VGluT3 knockout mice. Stress 2018, 21, 151–161. [Google Scholar] [CrossRef]
- Horvath, H.R.; Fazekas, C.L.; Balazsfi, D.; Jain, S.K.; Haller, J.; Zelena, D. Contribution of vesicular glutamate transporters to stress response and related psychopathologies: Studies in VGluT3 knockout mice. Cell Mol. Neurobiol. 2018, 38, 37–52. [Google Scholar] [CrossRef]
- Olivan, A.M.; Perez-Rodriguez, R.; Roncero, C.; Arce, C.; Gonzalez, M.P.; Oset-Gasque, M.J. Plasma membrane and vesicular glutamate transporter expression in chromaffin cells of bovine adrenal medulla. J. Neurosci. Res. 2011, 89, 44–57. [Google Scholar] [CrossRef]
- Balazsfi, D.; Farkas, L.; Csikota, P.; Fodor, A.; Zsebok, S.; Haller, J.; Zelena, D. Sex-dependent role of vesicular glutamate transporter 3 in stress-regulation and related anxiety phenotype during the early postnatal period. Stress 2016, 19, 434–438. [Google Scholar] [CrossRef]
- Prakash, N.; Stark, C.J.; Keisler, M.N.; Luo, L.; Der-Avakian, A.; Dulcis, D. Serotonergic plasticity in the dorsal raphe nucleus characterizes susceptibility and resilience to anhedonia. J. Neurosci. 2020, 40, 569–584. [Google Scholar] [CrossRef] [PubMed]
- Gammelsaeter, R.; Coppola, T.; Marcaggi, P.; Storm-Mathisen, J.; Chaudhry, F.A.; Attwell, D.; Regazzi, R.; Gundersen, V. A role for glutamate transporters in the regulation of insulin secretion. PLoS ONE 2011, 6, e22960. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.L.; Moazzami, A.R.; Longhurst, J.C. Stimulation of cardiac sympathetic afferents activates glutamatergic neurons in the parabrachial nucleus: Relation to neurons containing nNOS. Brain Res. 2005, 1053, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, E.J.; Muntner, P.; Alonso, A.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Das, S.R.; et al. Heart disease and stroke statistics-2019 update: A report from the american heart association. Circulation 2019, 139, e56–e528. [Google Scholar] [CrossRef]
- Krzyzanowska, W.; Pomierny, B.; Budziszewska, B.; Filip, M.; Pera, J. N-Acetylcysteine and ceftriaxone as preconditioning strategies in focal brain ischemia: Influence on glutamate transporters expression. Neurotox. Res. 2016, 29, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Castillo, J.; Alvarez-Sabin, J.; Davalos, A.; Diez-Tejedor, E.; Lizasoain, I.; Martinez-Vila, E.; Vivancos, J.; Zarranz, J.J. Consensus review. Pharmacological neuroprotection in cerebral ischemia: Is it still a therapeutic option? Neurologia 2003, 18, 368–384. [Google Scholar] [PubMed]
- Krzyzanowska, W.; Pomierny, B.; Bystrowska, B.; Pomierny-Chamiolo, L.; Filip, M.; Budziszewska, B.; Pera, J. Ceftriaxone- and N-acetylcysteine-induced brain tolerance to ischemia: Influence on glutamate levels in focal cerebral ischemia. PLoS ONE 2017, 12, e0186243. [Google Scholar] [CrossRef]
- Sanchez-Mendoza, E.; Burguete, M.C.; Castello-Ruiz, M.; Gonzalez, M.P.; Roncero, C.; Salom, J.B.; Arce, C.; Canadas, S.; Torregrosa, G.; Alborch, E.; et al. Transient focal cerebral ischemia significantly alters not only EAATs but also VGLUTs expression in rats: Relevance of changes in reactive astroglia. J. Neurochem. 2010, 113, 1343–1355. [Google Scholar] [CrossRef] [PubMed]
- Callaerts-Vegh, Z.; Moechars, D.; Van Acker, N.; Daneels, G.; Goris, I.; Leo, S.; Naert, A.; Meert, T.; Balschun, D.; D’Hooge, R. Haploinsufficiency of VGluT1 but not VGluT2 impairs extinction of spatial preference and response suppression. Behav. Brain Res. 2013, 245, 13–21. [Google Scholar] [CrossRef]
- Wallen-Mackenzie, A.; Gezelius, H.; Thoby-Brisson, M.; Nygard, A.; Enjin, A.; Fujiyama, F.; Fortin, G.; Kullander, K. Vesicular glutamate transporter 2 is required for central respiratory rhythm generation but not for locomotor central pattern generation. J. Neurosci. 2006, 26, 12294–12307. [Google Scholar] [CrossRef]
- Wallen-Mackenzie, A.; Wootz, H.; Englund, H. Genetic inactivation of the vesicular glutamate transporter 2 (VGLUT2) in the mouse: What have we learnt about functional glutamatergic neurotransmission? Ups J. Med. Sci. 2010, 115, 11–20. [Google Scholar] [CrossRef]
- Moechars, D.; Weston, M.C.; Leo, S.; Callaerts-Vegh, Z.; Goris, I.; Daneels, G.; Buist, A.; Cik, M.; van der Spek, P.; Kass, S.; et al. Vesicular glutamate transporter VGLUT2 expression levels control quantal size and neuropathic pain. J. Neurosci. 2006, 26, 12055–12066. [Google Scholar] [CrossRef]
- Fremeau, R.T., Jr.; Kam, K.; Qureshi, T.; Johnson, J.; Copenhagen, D.R.; Storm-Mathisen, J.; Chaudhry, F.A.; Nicoll, R.A.; Edwards, R.H. Vesicular glutamate transporters 1 and 2 target to functionally distinct synaptic release sites. Science 2004, 304, 1815–1819. [Google Scholar] [CrossRef]
- Gezelius, H.; Wallen-Mackenzie, A.; Enjin, A.; Lagerstrom, M.; Kullander, K. Role of glutamate in locomotor rhythm generating neuronal circuitry. J. Physiol.-Paris 2006, 100, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Divito, C.B.; Steece-Collier, K.; Case, D.T.; Williams, S.P.; Stancati, J.A.; Zhi, L.; Rubio, M.E.; Sortwell, C.E.; Collier, T.J.; Sulzer, D.; et al. Loss of VGLUT3 Produces circadian-dependent hyperdopaminergia and ameliorates motor dysfunction and l-dopa-mediated dyskinesias in a model of parkinson’s disease. J. Neurosci. 2015, 35, 14983–14999. [Google Scholar] [CrossRef]
- Sakae, D.Y.; Ramet, L.; Henrion, A.; Poirel, O.; Jamain, S.; El Mestikawy, S.; Daumas, S. Differential expression of VGLUT3 in laboratory mouse strains: Impact on drug-induced hyperlocomotion and anxiety-related behaviors. Genes Brain Behav. 2019, 18, e12528. [Google Scholar] [CrossRef] [PubMed]
- Sakae, D.Y.; Marti, F.; Lecca, S.; Vorspan, F.; Martin-Garcia, E.; Morel, L.J.; Henrion, A.; Gutierrez-Cuesta, J.; Besnard, A.; Heck, N.; et al. The absence of VGLUT3 predisposes to cocaine abuse by increasing dopamine and glutamate signaling in the nucleus accumbens. Mol. Psychiatry 2015, 20, 1448–1459. [Google Scholar] [CrossRef]
- Gangarossa, G.; Guzman, M.; Prado, V.F.; Prado, M.A.; Daumas, S.; El Mestikawy, S.; Valjent, E. Role of the atypical vesicular glutamate transporter VGLUT3 in l-DOPA-induced dyskinesia. Neurobiol. Dis. 2016, 87, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Mansouri-Guilani, N.; Bernard, V.; Vigneault, E.; Vialou, V.; Daumas, S.; El Mestikawy, S.; Gangarossa, G. VGLUT3 gates psychomotor effects induced by amphetamine. J. Neurochem. 2019, 148, 779–795. [Google Scholar] [CrossRef]
- Fontaine, H.M.; Silva, P.R.; Neiswanger, C.; Tran, R.; Abraham, A.D.; Land, B.B.; Neumaier, J.F.; Chavkin, C. Stress decreases serotonin tone in the nucleus accumbens in male mice to promote aversion and potentiate cocaine preference via decreased stimulation of 5-HT1B receptors. Neuropsychopharmacology 2021. [Google Scholar] [CrossRef] [PubMed]
- Ramet, L.; Zimmermann, J.; Bersot, T.; Poirel, O.; De Gois, S.; Silm, K.; Sakae, D.Y.; Mansouri-Guilani, N.; Bourque, M.J.; Trudeau, L.E.; et al. Characterization of a human point mutation of VGLUT3 (p.A211V) in the rodent brain suggests a nonuniform distribution of the transporter in synaptic vesicles. J. Neurosci. 2017, 37, 4181–4199. [Google Scholar] [CrossRef]
- Fazekas, C.L.; Balazsfi, D.; Horvath, H.R.; Balogh, Z.; Aliczki, M.; Puhova, A.; Balagova, L.; Chmelova, M.; Jezova, D.; Haller, J.; et al. Consequences of VGluT3 deficiency on learning and memory in mice. Physiol. Behav. 2019, 212, 112688. [Google Scholar] [CrossRef]
- Cheng, X.R.; Yang, Y.; Zhou, W.X.; Zhang, Y.X. Expression of VGLUTs contributes to degeneration and acquisition of learning and memory. Neurobiol. Learn. Mem. 2011, 95, 361–375. [Google Scholar] [CrossRef]
- Cossart, R.; Khazipov, R. How development sculpts hippocampal circuits and function. Physiol. Rev. 2022, 102, 343–378. [Google Scholar] [CrossRef] [PubMed]
- Wirth, S.; Soumier, A.; Eliava, M.; Derdikman, D.; Wagner, S.; Grinevich, V.; Sirigu, A. Territorial blueprint in the hippocampal system. Trends Cogn. Sci. 2021, 25, 831–842. [Google Scholar] [CrossRef]
- Sachser, R.M.; Haubrich, J.; Lunardi, P.S.; de Oliveira Alvares, L. Forgetting of what was once learned: Exploring the role of postsynaptic ionotropic glutamate receptors on memory formation, maintenance, and decay. Neuropharmacology 2017, 112, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Manahan-Vaughan, D. Role of metabotropic glutamate receptors in persistent forms of hippocampal plasticity and learning. Neuropharmacology 2013, 66, 65–81. [Google Scholar] [CrossRef] [PubMed]
- Femenia, T.; Gomez-Galan, M.; Lindskog, M.; Magara, S. Dysfunctional hippocampal activity affects emotion and cognition in mood disorders. Brain Res. 2012, 1476, 58–70. [Google Scholar] [CrossRef]
- Frey, B.N.; Andreazza, A.C.; Nery, F.G.; Martins, M.R.; Quevedo, J.; Soares, J.C.; Kapczinski, F. The role of hippocampus in the pathophysiology of bipolar disorder. Behav. Pharmacol. 2007, 18, 419–430. [Google Scholar] [CrossRef]
- Szonyi, A.; Mayer, M.I.; Cserep, C.; Takacs, V.T.; Watanabe, M.; Freund, T.F.; Nyiri, G. The ascending median raphe projections are mainly glutamatergic in the mouse forebrain. Brain Struct. Funct. 2016, 221, 735–751. [Google Scholar] [CrossRef]
- Klausberger, T.; Somogyi, P. Neuronal diversity and temporal dynamics: The unity of hippocampal circuit operations. Science 2008, 321, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Harris, K.D.; Hochgerner, H.; Skene, N.G.; Magno, L.; Katona, L.; Bengtsson Gonzales, C.; Somogyi, P.; Kessaris, N.; Linnarsson, S.; Hjerling-Leffler, J. Classes and continua of hippocampal CA1 inhibitory neurons revealed by single-cell transcriptomics. PLoS Biol. 2018, 16, e2006387. [Google Scholar] [CrossRef]
- Semyanov, A.; Kullmann, D.M. Modulation of GABAergic signaling among interneurons by metabotropic glutamate receptors. Neuron 2000, 25, 663–672. [Google Scholar] [CrossRef]
- Aznar, S.; Qian, Z.X.; Knudsen, G.M. Non-serotonergic dorsal and median raphe projection onto parvalbumin- and calbindin-containing neurons in hippocampus and septum. Neuroscience 2004, 124, 573–581. [Google Scholar] [CrossRef]
- McQuade, R.; Sharp, T. Functional mapping of dorsal and median raphe 5-hydroxytryptamine pathways in forebrain of the rat using microdialysis. J. Neurochem. 1997, 69, 791–796. [Google Scholar] [CrossRef]
- Kusljic, S.; van den Buuse, M. Functional dissociation between serotonergic pathways in dorsal and ventral hippocampus in psychotomimetic drug-induced locomotor hyperactivity and prepulse inhibition in rats. Eur. J. Neurosci. 2004, 20, 3424–3432. [Google Scholar] [CrossRef] [PubMed]
- Molliver, M.E. Serotonergic neuronal systems: What their anatomic organization tells us about function. J. Clin. Psychopharmacol. 1987, 7, 3S–23S. [Google Scholar] [CrossRef] [PubMed]
- Commons, K.G. Ascending serotonin neuron diversity under two umbrellas. Brain Struct. Funct. 2016, 221, 3347–3360. [Google Scholar] [CrossRef]
- McKenna, J.T.; Vertes, R.P. Collateral projections from the median raphe nucleus to the medial septum and hippocampus. Brain Res. Bull. 2001, 54, 619–630. [Google Scholar] [CrossRef]
- Vertes, R.P.; Fortin, W.J.; Crane, A.M. Projections of the median raphe nucleus in the rat. J. Comp. Neurol. 1999, 407, 555–582. [Google Scholar] [CrossRef]
- Jackson, J.; Bland, B.H.; Antle, M.C. Nonserotonergic projection neurons in the midbrain raphe nuclei contain the vesicular glutamate transporter VGLUT3. Synapse 2009, 63, 31–41. [Google Scholar] [CrossRef]
- Freund, T.F.; Gulyas, A.I.; Acsady, L.; Gorcs, T.; Toth, K. Serotonergic control of the hippocampus via local inhibitory interneurons. Proc. Natl. Acad. Sci. USA 1990, 87, 8501–8505. [Google Scholar] [CrossRef]
- Halasy, K.; Miettinen, R.; Szabat, E.; Freund, T.F. GABAergic Interneurons are the major postsynaptic targets of median raphe afferents in the rat dentate gyrus. Eur. J. Neurosci. 1992, 4, 144–153. [Google Scholar] [CrossRef]
- Hornung, J.P.; Celio, M.R. The selective innervation by serotoninergic axons of calbindin-containing interneurons in the neocortex and hippocampus of the marmoset. J. Comp. Neurol. 1992, 320, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Varga, V.; Losonczy, A.; Zemelman, B.V.; Borhegyi, Z.; Nyiri, G.; Domonkos, A.; Hangya, B.; Holderith, N.; Magee, J.C.; Freund, T.F. Fast synaptic subcortical control of hippocampal circuits. Science 2009, 326, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Domonkos, A.; Nikitidou Ledri, L.; Laszlovszky, T.; Cserep, C.; Borhegyi, Z.; Papp, E.; Nyiri, G.; Freund, T.F.; Varga, V. Divergent in vivo activity of non-serotonergic and serotonergic VGluT3-neurones in the median raphe region. J. Physiol. 2016, 594, 3775–3790. [Google Scholar] [CrossRef]
- Leranth, C.; Vertes, R.P. Median raphe serotonergic innervation of medial septum/diagonal band of broca (MSDB) parvalbumin-containing neurons: Possible involvement of the MSDB in the desynchronization of the hippocampal EEG. J. Comp. Neurol. 1999, 410, 586–598. [Google Scholar] [CrossRef]
- Alreja, M. Excitatory actions of serotonin on GABAergic neurons of the medial septum and diagonal band of Broca. Synapse 1996, 22, 15–27. [Google Scholar] [CrossRef]
- Liu, W.; Alreja, M. Atypical antipsychotics block the excitatory effects of serotonin in septohippocampal neurons in the rat. Neuroscience 1997, 79, 369–382. [Google Scholar] [CrossRef]
- Assaf, S.Y.; Miller, J.J. The role of a raphe serotonin system in the control of septal unit activity and hippocampal desynchronization. Neuroscience 1978, 3, 539–550. [Google Scholar] [CrossRef]
- Kinney, G.G.; Kocsis, B.; Vertes, R.P. Injections of excitatory amino acid antagonists into the median raphe nucleus produce hippocampal theta rhythm in the urethane-anesthetized rat. Brain Res. 1994, 654, 96–104. [Google Scholar] [CrossRef]
- Okaty, B.W.; Freret, M.E.; Rood, B.D.; Brust, R.D.; Hennessy, M.L.; deBairos, D.; Kim, J.C.; Cook, M.N.; Dymecki, S.M. Multi-Scale molecular deconstruction of the serotonin neuron system. Neuron 2015, 88, 774–791. [Google Scholar] [CrossRef]
- Tavares, L.C.S.; Tort, A.B.L. Hippocampal-prefrontal interactions during spatial decision-making. Hippocampus 2022, 32, 38–54. [Google Scholar] [CrossRef]
- Ego-Stengel, V.; Wilson, M.A. Disruption of ripple-associated hippocampal activity during rest impairs spatial learning in the rat. Hippocampus 2010, 20, 1–10. [Google Scholar] [CrossRef]
- Girardeau, G.; Benchenane, K.; Wiener, S.I.; Buzsaki, G.; Zugaro, M.B. Selective suppression of hippocampal ripples impairs spatial memory. Nat. Neurosci. 2009, 12, 1222–1223. [Google Scholar] [CrossRef]
- Mizuseki, K.; Miyawaki, H. Hippocampal information processing across sleep/wake cycles. Neurosci. Res. 2017, 118, 30–47. [Google Scholar] [CrossRef]
- Khodagholy, D.; Gelinas, J.N.; Buzsaki, G. Learning-enhanced coupling between ripple oscillations in association cortices and hippocampus. Science 2017, 358, 369–372. [Google Scholar] [CrossRef]
- Szucs, A.; Ratkai, A.; Schlett, K.; Huerta, R. Frequency-dependent regulation of intrinsic excitability by voltage-activated membrane conductances, computational modeling and dynamic clamp. Eur. J. Neurosci. 2017, 46, 2429–2444. [Google Scholar] [CrossRef] [PubMed]
- Amilhon, B.; Huh, C.Y.; Manseau, F.; Ducharme, G.; Nichol, H.; Adamantidis, A.; Williams, S. parvalbumin interneurons of hippocampus tune population activity at theta frequency. Neuron 2015, 86, 1277–1289. [Google Scholar] [CrossRef] [PubMed]
- Soltani Zangbar, H.; Ghadiri, T.; Seyedi Vafaee, M.; Ebrahimi Kalan, A.; Fallahi, S.; Ghorbani, M.; Shahabi, P. Theta oscillations through hippocampal/prefrontal pathway: Importance in cognitive performances. Brain Connect. 2020, 10, 157–169. [Google Scholar] [CrossRef]
- Wang, D.V.; Yau, H.J.; Broker, C.J.; Tsou, J.H.; Bonci, A.; Ikemoto, S. Mesopontine median raphe regulates hippocampal ripple oscillation and memory consolidation. Nat. Neurosci. 2015, 18, 728–735. [Google Scholar] [CrossRef]
- Willner, P. Reliability of the chronic mild stress model of depression: A user survey. Neurobiol. Stress 2017, 6, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Willner, P.; Benton, D.; Brown, E.; Cheeta, S.; Davies, G.; Morgan, J.; Morgan, M. “Depression” increases “craving” for sweet rewards in animal and human models of depression and craving. Psychopharmacology 1998, 136, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Beecher, K.; Wang, J.; Jacques, A.; Chaaya, N.; Chehrehasa, F.; Belmer, A.; Bartlett, S.E. Sucrose consumption alters serotonin/glutamate co-localisation within the prefrontal cortex and hippocampus of mice. Front. Mol. Neurosci. 2021, 14, 678267. [Google Scholar] [CrossRef] [PubMed]
- Tukey, D.S.; Lee, M.; Xu, D.; Eberle, S.E.; Goffer, Y.; Manders, T.R.; Ziff, E.B.; Wang, J. Differential effects of natural rewards and pain on vesicular glutamate transporter expression in the nucleus accumbens. Mol. Brain 2013, 6, 32. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Han, Y.S.; Liu, L.; Tang, L.; Yang, H.; Meng, P.; Zhao, H.Q.; Wang, Y.H. Abnormal Glu/mGluR2/3/PI3K pathway in the hippocampal neurovascular unit leads to diabetes-related depression. Neural. Regen. Res. 2021, 16, 727–733. [Google Scholar] [CrossRef]
- Abela, A.R.; Browne, C.J.; Sargin, D.; Prevot, T.D.; Ji, X.D.; Li, Z.; Lambe, E.K.; Fletcher, P.J. Median raphe serotonin neurons promote anxiety-like behavior via inputs to the dorsal hippocampus. Neuropharmacology 2020, 168, 107985. [Google Scholar] [CrossRef]
- Van Liefferinge, J.; Jensen, C.J.; Albertini, G.; Bentea, E.; Demuyser, T.; Merckx, E.; Aronica, E.; Smolders, I.; Massie, A. Altered vesicular glutamate transporter expression in human temporal lobe epilepsy with hippocampal sclerosis. Neurosci. Lett. 2015, 590, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Llorente, I.L.; Perez-Rodriguez, D.; Burgin, T.C.; Gonzalo-Orden, J.M.; Martinez-Villayandre, B.; Fernandez-Lopez, A. Age and meloxicam modify the response of the glutamate vesicular transporters (VGLUTs) after transient global cerebral ischemia in the rat brain. Brain Res. Bull. 2013, 94, 90–97. [Google Scholar] [CrossRef]
- McLellan, M.A.; Rosenthal, N.A.; Pinto, A.R. Cre-loxP-Mediated recombination: General principles and experimental considerations. Curr. Protoc. Mouse Biol. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Sternberg, N.; Hamilton, D. Bacteriophage P1 site-specific recombination. I. Recombination between loxP sites. J. Mol. Biol. 1981, 150, 467–486. [Google Scholar] [CrossRef]
- Zelena, D.; Demeter, K.; Haller, J.; Balazsfi, D. Considerations for the use of virally delivered genetic tools for in-vivo circuit analysis and behavior in mutant mice: A practical guide to optogenetics. Behav. Pharmacol. 2017, 28, 598–609. [Google Scholar] [CrossRef]
- Vooijs, M.; Jonkers, J.; Berns, A. A highly efficient ligand-regulated Cre recombinase mouse line shows that LoxP recombination is position dependent. EMBO Rep. 2001, 2, 292–297. [Google Scholar] [CrossRef]
- Wunderlich, F.T.; Wildner, H.; Rajewsky, K.; Edenhofer, F. New variants of inducible Cre recombinase: A novel mutant of Cre-PR fusion protein exhibits enhanced sensitivity and an expanded range of inducibility. Nucleic Acids Res. 2001, 29, E47. [Google Scholar] [CrossRef]
- Karray, S.; Kress, C.; Cuvellier, S.; Hue-Beauvais, C.; Damotte, D.; Babinet, C.; Levi-Strauss, M. Complete loss of Fas ligand gene causes massive lymphoproliferation and early death, indicating a residual activity of gld allele. J. Immunol. 2004, 172, 2118–2125. [Google Scholar] [CrossRef]
- Raab, S.; Beck, H.; Gaumann, A.; Yuce, A.; Gerber, H.P.; Plate, K.; Hammes, H.P.; Ferrara, N.; Breier, G. Impaired brain angiogenesis and neuronal apoptosis induced by conditional homozygous inactivation of vascular endothelial growth factor. Thromb. Haemost. 2004, 91, 595–605. [Google Scholar] [CrossRef]
- Schmidt, E.E.; Taylor, D.S.; Prigge, J.R.; Barnett, S.; Capecchi, M.R. Illegitimate Cre-dependent chromosome rearrangements in transgenic mouse spermatids. Proc. Natl. Acad. Sci. USA 2000, 97, 13702–13707. [Google Scholar] [CrossRef] [PubMed]
- Papathanou, M.; Dumas, S.; Pettersson, H.; Olson, L.; Wallen-Mackenzie, A. Off-target effects in transgenic mice: Characterization of dopamine transporter (DAT)-Cre transgenic mouse lines exposes multiple non-dopaminergic neuronal clusters available for selective targeting within limbic neurocircuitry. eNeuro 2019, 6. [Google Scholar] [CrossRef]
- Schmidt-Supprian, M.; Rajewsky, K. Vagaries of conditional gene targeting. Nat. Immunol. 2007, 8, 665–668. [Google Scholar] [CrossRef] [PubMed]
- Heffner, C.S.; Herbert Pratt, C.; Babiuk, R.P.; Sharma, Y.; Rockwood, S.F.; Donahue, L.R.; Eppig, J.T.; Murray, S.A. Supporting conditional mouse mutagenesis with a comprehensive cre characterization resource. Nat. Commun. 2012, 3, 1218. [Google Scholar] [CrossRef] [PubMed]
- Guru, A.; Post, R.J.; Ho, Y.Y.; Warden, M.R. Making sense of optogenetics. Int. J. Neuropsychopharmacol. 2015, 18, pyv079. [Google Scholar] [CrossRef]
- Nagel, G.; Szellas, T.; Huhn, W.; Kateriya, S.; Adeishvili, N.; Berthold, P.; Ollig, D.; Hegemann, P.; Bamberg, E. Channelrhodopsin-2, a directly light-gated cation-selective membrane channel. Proc. Natl. Acad. Sci. USA 2003, 100, 13940–13945. [Google Scholar] [CrossRef]
- Jiang, J.; Cui, H.; Rahmouni, K. Optogenetics and pharmacogenetics: Principles and applications. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 313, R633–R645. [Google Scholar] [CrossRef]
- Chen, R.; Gore, F.; Nguyen, Q.A.; Ramakrishnan, C.; Patel, S.; Kim, S.H.; Raffiee, M.; Kim, Y.S.; Hsueh, B.; Krook-Magnusson, E.; et al. Deep brain optogenetics without intracranial surgery. Nat. Biotechnol. 2021, 39, 161–164. [Google Scholar] [CrossRef] [PubMed]
- Alexander, G.M.; Rogan, S.C.; Abbas, A.I.; Armbruster, B.N.; Pei, Y.; Allen, J.A.; Nonneman, R.J.; Hartmann, J.; Moy, S.S.; Nicolelis, M.A.; et al. Remote control of neuronal activity in transgenic mice expressing evolved G protein-coupled receptors. Neuron 2009, 63, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Armbruster, B.N.; Li, X.; Pausch, M.H.; Herlitze, S.; Roth, B.L. Evolving the lock to fit the key to create a family of G protein-coupled receptors potently activated by an inert ligand. Proc. Natl. Acad. Sci. USA 2007, 104, 5163–5168. [Google Scholar] [CrossRef]
- Campbell, E.J.; Marchant, N.J. The use of chemogenetics in behavioural neuroscience: Receptor variants, targeting approaches and caveats. Br. J. Pharmacol. 2018, 175, 994–1003. [Google Scholar] [CrossRef]
- Liguz-Lecznar, M.; Skangiel-Kramska, J. Vesicular glutamate transporters (VGLUTs): The three musketeers of glutamatergic system. Acta Neurobiol. Exp. (Wars) 2007, 67, 207–218. [Google Scholar]
- Kehrl, J.; Althaus, J.C.; Showalter, H.D.; Rudzinski, D.M.; Sutton, M.A.; Ueda, T. Vesicular glutamate transporter inhibitors: Structurally modified brilliant yellow analogs. Neurochem. Res. 2017, 42, 1823–1832. [Google Scholar] [CrossRef]
- Thompson, C.M.; Chao, C.K. VGLUT substrates and inhibitors: A computational viewpoint. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183175. [Google Scholar] [CrossRef] [PubMed]
- Gegelashvili, G.; Bjerrum, O.J. Glutamate transport system as a key constituent of glutamosome: Molecular pathology and pharmacological modulation in chronic pain. Neuropharmacology 2019, 161, 107623. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GABAergic Interneurons in the Cortex | GABAergic Interneurons in the Hippocampus | VGLUT3+ Interneurons in the Amygdala | VGLUT3+ Interneurons in the Hippocampus | |
|---|---|---|---|---|
| Resting membrane potential | −57.48–−49.40 mV | NA | NA | −59.00–−56.90 mV |
| Input resistance | 219.77–419.61 MΩ | 107.89 MΩ | 168.10 MΩ | 149.70–158.50 MΩ |
| Action potential threshold | −32.67–−27.82 mV | −42.81 mV | −38.80 mV | −41.90–−39.86 mV |
| Action potential amplitude | 71.30–86.11 mV | 74.27 mV | 71.60 mV | 55.70–57.40 mV |
| Firing frequency | 19.34–52.48 Hz (2×) | 15.00 Hz (steady trace) | 31.50 Hz (2×) | 31.30–34.90 Hz (2×) |
| Amplitude of after-hyperpolarisation | 8.60–17.63 mV | 12.68 mV (new method) | 14.70 mV | −11.80–−10.30 mV |
| Co-transmitters | CCK | CCK | CCK, GABA | CCK, GABA |
| Reference | [71], all subtypes displayed | [72] | [27] | [65], both subtypes displayed |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fazekas, C.L.; Szabó, A.; Török, B.; Bánrévi, K.; Correia, P.; Chaves, T.; Daumas, S.; Zelena, D. A New Player in the Hippocampus: A Review on VGLUT3+ Neurons and Their Role in the Regulation of Hippocampal Activity and Behaviour. Int. J. Mol. Sci. 2022, 23, 790. https://doi.org/10.3390/ijms23020790
Fazekas CL, Szabó A, Török B, Bánrévi K, Correia P, Chaves T, Daumas S, Zelena D. A New Player in the Hippocampus: A Review on VGLUT3+ Neurons and Their Role in the Regulation of Hippocampal Activity and Behaviour. International Journal of Molecular Sciences. 2022; 23(2):790. https://doi.org/10.3390/ijms23020790
Chicago/Turabian StyleFazekas, Csilla Lea, Adrienn Szabó, Bibiána Török, Krisztina Bánrévi, Pedro Correia, Tiago Chaves, Stéphanie Daumas, and Dóra Zelena. 2022. "A New Player in the Hippocampus: A Review on VGLUT3+ Neurons and Their Role in the Regulation of Hippocampal Activity and Behaviour" International Journal of Molecular Sciences 23, no. 2: 790. https://doi.org/10.3390/ijms23020790
APA StyleFazekas, C. L., Szabó, A., Török, B., Bánrévi, K., Correia, P., Chaves, T., Daumas, S., & Zelena, D. (2022). A New Player in the Hippocampus: A Review on VGLUT3+ Neurons and Their Role in the Regulation of Hippocampal Activity and Behaviour. International Journal of Molecular Sciences, 23(2), 790. https://doi.org/10.3390/ijms23020790