
The Brain–Skin Axis in Psoriasis—Psychological, Psychiatric, Hormonal, and Dermatological Aspects

 , , ,
, , ,  , , and
, , and
Abstract
1. Introduction

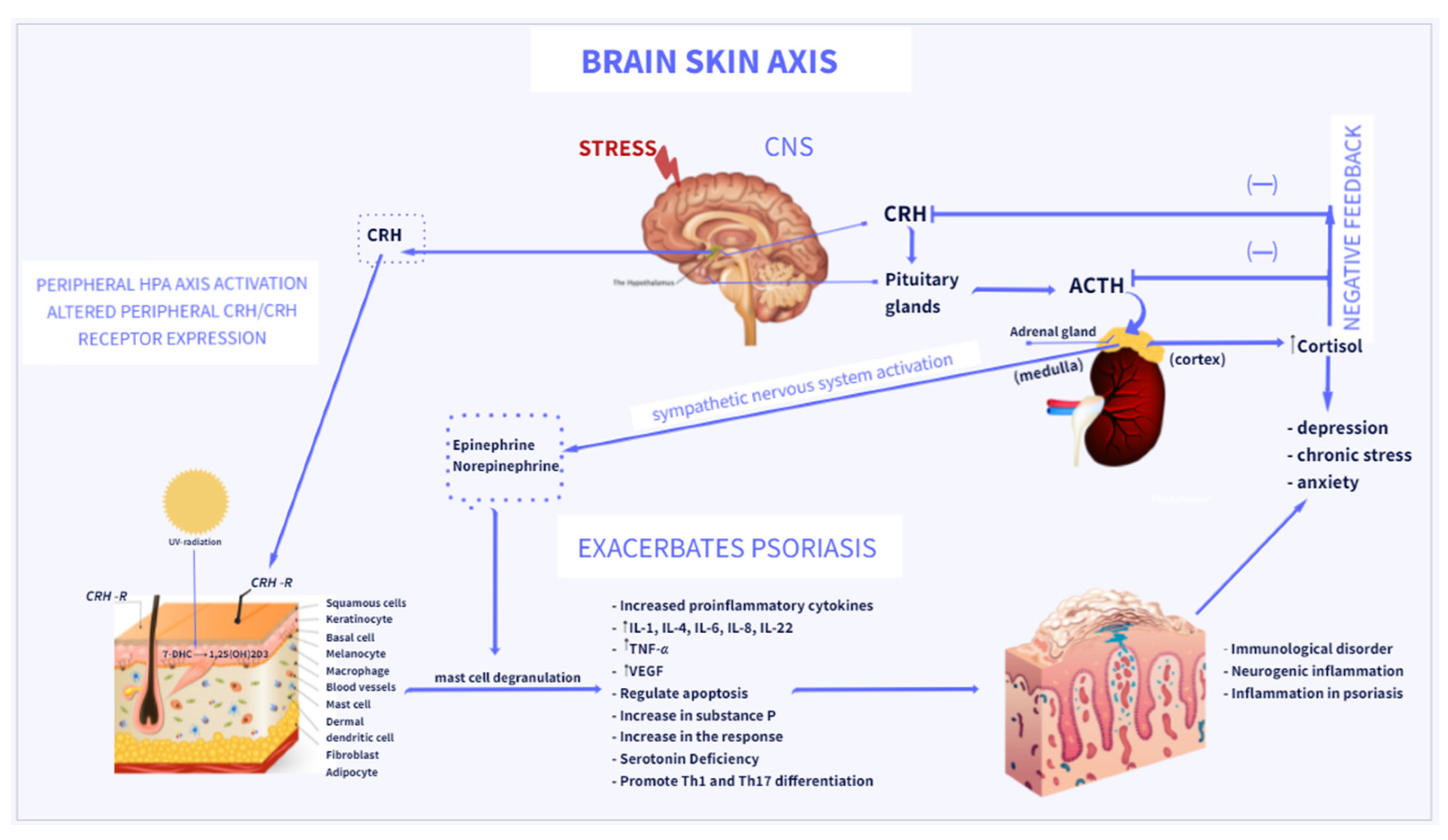{kind=link}
| Aspects | Consequences | Study |
|---|---|---|
| psychological | decrease in the quality of life | Randa et al. [2] |
| stressful life episodes | Rousset et al. [8] | |
| cognitive impairment | Innamorati et al. [12] | |
| psychiatric | anxiety, depression | Singh et al. [4], González-Parra et al. [14] |
| suicidal ideation | Dalgard et al. [6] | |
| hormonal, immunological | tissue inflammation, mood disorders | Slominski et al. [43], Kim et al. [44], González-Parra et al. [14], Ayasse et al. [45] |
| dermatological | exacerbates psoriasis | Kamiya et al. [46], Michalek et al. [7] |
2. Psychological Stress, Inflammation of Psoriasis
3. The Association between Psoriasis and Depression
4. The HPA Axis Links Psoriasis and Depression
5. Neuropeptides in Psoriasis and Depression
6. Sunlight Deficiency Influences Psoriasis and Depression
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jafferany, M.; Patel, A. Understanding Psychocutaneous Disease: Psychosocial & Psychoneuroimmunologic Perspectives. Int. J. Dermatol. 2020, 59, 8–15. [Google Scholar] [CrossRef]
- Randa, H.; Lomholt, J.J.; Skov, L.; Zachariae, R. Health-Related Quality of Life in Adolescents with Psoriasis: An Interview-Based Study. Br. J. Dermatol. 2018, 178, 1404–1411. [Google Scholar] [CrossRef] [PubMed]
- Otte, C.; Pariante, C.M.; Etkin, A.; Fava, M.; Mohr, D.C.; Schatzberg, A.F. Major Depressive Disorder. Nat. Rev. Dis. Primers 2016, 2, 16065. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Taylor, C.; Kornmehl, H.; Armstrong, A.W. Psoriasis and Suicidality: A Systematic Review and Meta-Analysis. J. Am. Acad. Dermatol. 2017, 77, 425–440.e2. [Google Scholar] [CrossRef] [PubMed]
- Tampa, M.; Sarbu, M.-I.; Mitran, M.-I.; Mitran, C.-I.; Matei, C.; Georgescu, S.-R. The Pathophysiological Mechanisms and the Quest for Biomarkers in Psoriasis, a Stress-Related Skin Disease. Dis. Markers 2018, 2018, 1–14. [Google Scholar] [CrossRef]
- Dalgard, F.J.; Gieler, U.; Tomas-Aragones, L.; Lien, L.; Poot, F.; Jemec, G.B.E.; Misery, L.; Szabo, C.; Linder, D.; Sampogna, F.; et al. The Psychological Burden of Skin Diseases: A Cross-Sectional Multicenter Study among Dermatological Out-Patients in 13 European Countries. J. Investig. Dermatol. 2015, 135, 984–991. [Google Scholar] [CrossRef]
- Michalek, I.M. A Systematic Review of Worldwide Epidemiology of Psoriasis. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 205–212. [Google Scholar] [CrossRef]
- Rousset, L.; Halioua, B. Stress and Psoriasis. Int. J. Dermatol. 2018, 57, 1165–1172. [Google Scholar] [CrossRef]
- Pettey, A.A.; Balkrishnan, R.; Rapp, S.R.; Fleischer, A.B.; Feldman, S.R. Patients with Palmoplantar Psoriasis Have More Physical Disability and Discomfort than Patients with Other Forms of Psoriasis: Implications for Clinical Practice. J. Am. Acad. Dermatol. 2003, 49, 271–275. [Google Scholar] [CrossRef]
- Damiani, G.; Kridin, K.; Pacifico, A.; Malagoli, P.; Pigatto, P.D.M.; Finelli, R.; Taccone, F.S.; Peluso, L.; Conic, R.R.Z.; Bragazzi, N.L.; et al. Antihistamines-Refractory Chronic Pruritus in Psoriatic Patients Undergoing Biologics: Aprepitant vs Antihistamine Double Dosage, a Real-World Data. J. Dermatol. Treat. 2020, 1–4. [Google Scholar] [CrossRef]
- Damiani, G.; Cazzaniga, S.; Conic, R.R.; Naldi, L. Pruritus Characteristics in a Large Italian Cohort of Psoriatic Patients. J. Eur. Acad. Dermatol. Venereol. 2019, 33, 1316–1324. [Google Scholar] [CrossRef] [PubMed]
- Innamorati, M.; Quinto, R.M.; Lester, D.; Iani, L.; Graceffa, D.; Bonifati, C. Cognitive Impairment in Patients with Psoriasis: A Matched Case-Control Study. J. Psychosom. Res. 2018, 105, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Moussavi, S.; Chatterji, S.; Verdes, E.; Tandon, A.; Patel, V.; Ustun, B. Depression, Chronic Diseases, and Decrements in Health: Results from the World Health Surveys. Lancet 2007, 370, 851–858. [Google Scholar] [CrossRef]
- González-Parra, S.; Daudén, E. Psoriasis and Depression: The Role of Inflammation. Actas Dermo-Sifiliogr. (Engl. Ed.) 2019, 110, 12–19. [Google Scholar] [CrossRef]
- Esposito, M.; Saraceno, R.; Giunta, A.; Maccarone, M.; Chimenti, S. An Italian Study on Psoriasis and Depression. Dermatology 2006, 212, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Koo, J.; Marangell, L.B.; Nakamura, M.; Armstrong, A.; Jeon, C.; Bhutani, T.; Wu, J.J. Depression and Suicidality in Psoriasis: Review of the Literature Including the Cytokine Theory of Depression. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 1999–2009. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-H.; Wang, W.-M.; Li, I.-H. Major Depressive Disorder Increased Risk of Psoriasis: A Propensity Score Matched Cohort Study. J. Affect. Disord. 2021, 278, 407–412. [Google Scholar] [CrossRef]
- Slominski, A.T.; Zmijewski, M.A.; Zbytek, B.; Tobin, D.J.; Theoharides, T.C.; Rivier, J. Key Role of CRF in the Skin Stress Response System. Endocr. Rev. 2013, 34, 827–884. [Google Scholar] [CrossRef]
- Slominski, A.; Wortsman, J. Neuroendocrinology of the Skin. Endocr. Rev. 2000, 21, 457–487. [Google Scholar] [CrossRef]
- Slominski, A.; Wortsman, J.; Luger, T.; Paus, R.; Solomon, S. Corticotropin Releasing Hormone and Proopiomelanocortin Involvement in the Cutaneous Response to Stress. Physiol. Rev. 2000, 80, 979–1020. [Google Scholar] [CrossRef]
- Slominski, A.; Zbytek, B.; Nikolakis, G.; Manna, P.R.; Skobowiat, C.; Zmijewski, M.; Li, W.; Janjetovic, Z.; Postlethwaite, A.; Zouboulis, C.C.; et al. Steroidogenesis in the Skin: Implications for Local Immune Functions. J. Steroid Biochem. Mol. Biol. 2013, 137, 107–123. [Google Scholar] [CrossRef] [PubMed]
- Slominski, R.M.; Tuckey, R.C.; Manna, P.R.; Jetten, A.M.; Postlethwaite, A.; Raman, C.; Slominski, A.T. Extra-Adrenal Glucocorticoid Biosynthesis: Implications for Autoimmune and Inflammatory Disorders. Genes Immun. 2020, 21, 150–168. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Zmijewski, M.A.; Skobowiat, C.; Zbytek, B.; Slominski, R.M.; Steketee, J.D. Sensing the Environment: Regulation of Local and Global Homeostasis by the Skin’s Neuroendocrine System; Advances in Anatomy, Embryology and Cell Biology; Springer Berlin Heidelberg: Berlin, Heidelberg, 2012; p. 212. ISBN 978-3-642-19682-9. [Google Scholar]
- Slominski, A.T.; Manna, P.R.; Tuckey, R.C. On the Role of Skin in the Regulation of Local and Systemic Steroidogenic Activities. Steroids 2015, 103, 72–88. [Google Scholar] [CrossRef] [PubMed]
- Slominski, R.M.; Raman, C.; Elmets, C.; Jetten, A.M.; Slominski, A.T.; Tuckey, R.C. The Significance of CYP11A1 Expression in Skin Physiology and Pathology. Mol. Cell. Endocrinol. 2021, 530, 111238. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Zmijewski, M.A.; Plonka, P.M.; Szaflarski, J.P.; Paus, R. How UV Light Touches the Brain and Endocrine System Through Skin, and Why. Endocrinology 2018, 159, 1992–2007. [Google Scholar] [CrossRef]
- Ferguson, F.J.; Lada, G.; Hunter, H.J.A.; Bundy, C.; Henry, A.L.; Griffiths, C.E.M.; Kleyn, C.E. Diurnal and Seasonal Variation in Psoriasis Symptoms. J. Eur. Acad. Dermatol. Venereol. 2021, 35, e45–e47. [Google Scholar] [CrossRef]
- Dowlati, Y.; Herrmann, N.; Swardfager, W.; Liu, H.; Sham, L.; Reim, E.K.; Lanctôt, K.L. A Meta-Analysis of Cytokines in Major Depression. Biol. Psychiatry 2010, 67, 446–457. [Google Scholar] [CrossRef]
- Jokela, M.; Virtanen, M.; Batty, G.D.; Kivimäki, M. Inflammation and Specific Symptoms of Depression. JAMA Psychiatry 2016, 73, 87. [Google Scholar] [CrossRef]
- Blalock, J.E. The Immune System as the Sixth Sense. J. Internal Med. 2005, 257, 126–138. [Google Scholar] [CrossRef]
- Blalock, J.E. The Syntax of Immune-Neuroendocrine Communication. Immunol. Today 1994, 15, 504–511. [Google Scholar] [CrossRef]
- Honeyman, J. Psychoneuroimmunology and the Skin. Acta Derm. Venerol. 2016, 96, 38–46. [Google Scholar] [CrossRef]
- Slominski, A.; Wortsman, J.; Tobin, D.J. The Cutaneous Serotoninergic/Melatoninergic System: Securing a Place under the Sun. FASEB J. 2005, 19, 176–194. [Google Scholar] [CrossRef]
- Slominski, A.; Pisarchik, A.; Semak, I.; Sweatman, T.; Wortsman, J.; Szczesniewski, A.; Slugocki, G.; McNulty, J.; Kauser, S.; Tobin, D.J.; et al. Serotoninergic and Melatoninergic Systems Are Fully Expressed in Human Skin. FASEB J. 2002, 16, 896–898. [Google Scholar] [CrossRef]
- Slominski, A.T.; Kim, T.; Kleszczyński, K.; Semak, I.; Janjetovic, Z.; Sweatman, T.; Skobowiat, C.; Steketee, J.D.; Lin, Z.; Postlethwaite, A.; et al. Characterization of Serotonin and N-acetylserotonin Systems in the Human Epidermis and Skin Cells. J. Pineal Res. 2020, 68, e12626. [Google Scholar] [CrossRef] [PubMed]
- Besedovsky, H.O.; Rey, A.D. Immune-Neuro-Endocrine Interactions: Facts and Hypotheses. Endocr. Rev. 1996, 17, 39. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A. A Nervous Breakdown in the Skin: Stress and the Epidermal Barrier. J. Clin. Investig. 2007, 117, 3166–3169. [Google Scholar] [CrossRef] [PubMed]
- Straub, R.H.; Cutolo, M. Psychoneuroimmunology—Developments in Stress Research. Wien Med. Wochenschr. 2018, 168, 76–84. [Google Scholar] [CrossRef]
- Blalock, J.E. Harnessing a Neural-Immune Circuit to Control Inflammation and Shock. J. Exp. Med. 2002, 195, F25–F28. [Google Scholar] [CrossRef] [PubMed]
- Pondeljak, N.; Lugović-Mihić, L. Stress-Induced Interaction of Skin Immune Cells, Hormones, and Neurotransmitters. Clin. Ther. 2020, 42, 757–770. [Google Scholar] [CrossRef]
- Slominski, A.T.; Hardeland, R.; Zmijewski, M.A.; Slominski, R.M.; Reiter, R.J.; Paus, R. Melatonin: A Cutaneous Perspective on Its Production, Metabolism, and Functions. J. Investig. Dermatol. 2018, 138, 490–499. [Google Scholar] [CrossRef]
- Younes, S.F. Immunohistochemical Evaluation of Role of Serotonin in Pathogenesis of Psoriasis. JCDR 2016. [Google Scholar] [CrossRef]
- Slominski, A.; Wortsman, J.; Tuckey, R.C.; Paus, R. Differential Expression of HPA Axis Homolog in the Skin. Mol. Cell. Endocrinol. 2007, 265–266, 143–149. [Google Scholar] [CrossRef]
- Kim, J.E.; Cho, D.H.; Kim, H.S.; Kim, H.J.; Lee, J.Y.; Cho, B.K.; Park, H.J. Expression of the Corticotropin-Releasing Hormone? Proopiomelanocortin Axis in the Various Clinical Types of Psoriasis. Exp. Dermatol. 2007, 16, 104–109. [Google Scholar] [CrossRef]
- Ayasse, M.T.; Buddenkotte, J.; Alam, M.; Steinhoff, M. Role of Neuroimmune Circuits and Pruritus in Psoriasis. Exp. Dermatol. 2020, 29, 414–426. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, K.; Kishimoto, M.; Sugai, J.; Komine, M.; Ohtsuki, M. Risk Factors for the Development of Psoriasis. IJMS 2019, 20, 4347. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zheng, J. Influence of Stress on the Development of Psoriasis. Clin. Exp. Dermatol. 2020, 45, 284–288. [Google Scholar] [CrossRef]
- Hunter, H.J.A.; Griffiths, C.E.M.; Kleyn, C.E. Does Psychosocial Stress Play a Role in the Exacerbation of Psoriasis? Br. J. Dermatol. 2013, 169, 965–974. [Google Scholar] [CrossRef]
- Connor, C.J.; Liu, V.; Fiedorowicz, J.G. Exploring the Physiological Link between Psoriasis and Mood Disorders. Dermatol. Res. Pract. 2015, 2015, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.M.; Podawiltz, A.; Mummert, D.I.; Jones, H.; Mummert, M.E. Psychological Stress and the Cutaneous Immune Response: Roles of the HPA Axis and the Sympathetic Nervous System in Atopic Dermatitis and Psoriasis. Dermatol. Res. Pract. 2012, 2012, 1–11. [Google Scholar] [CrossRef]
- Lin, T.-K.; Zhong, L.; Santiago, J. Association between Stress and the HPA Axis in the Atopic Dermatitis. IJMS 2017, 18, 2131. [Google Scholar] [CrossRef] [PubMed]
- Torales, J.; Echeverría, C.; Barrios, I.; García, O.; O’Higgins, M.; Castaldelli-Maia, J.M.; Ventriglio, A.; Jafferany, M. Psychodermatological Mechanisms of Psoriasis. Dermatol. Ther. 2020, 33, e13827. [Google Scholar] [CrossRef]
- Slominski, A. On the Role of the Corticotropin-Releasing Hormone Signalling System in the Aetiology of Inflammatory Skin Disorders. Br. J. Dermatol. 2009, 160, 229–232. [Google Scholar] [CrossRef]
- Greb, J.E.; Goldminz, A.M.; Elder, J.T.; Lebwohl, M.G.; Gladman, D.D.; Wu, J.J.; Mehta, N.N.; Finlay, A.Y.; Gottlieb, A.B. Psoriasis. Nat. Rev. Dis. Primers 2016, 2, 16082. [Google Scholar] [CrossRef]
- Kannan, S.; Heller, M.M.; Lee, E.S.; Koo, J.Y. The Role of Tumor Necrosis Factor-Alpha and Other Cytokines in Depression: What Dermatologists Should Know. J. Dermatol. Treat. 2013, 24, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C. The Mast Cell: A Neuroimmunoendocrine Master Player. Int. J. Tissue React. 1996, 18, 1–21. [Google Scholar]
- Theoharides, T.C. Neuroendocrinology of Mast Cells: Challenges and Controversies. Exp. Dermatol. 2017, 26, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Skobowiat, C.; Slominski, A.T. UVB Activates Hypothalamic–Pituitary–Adrenal Axis in C57BL/6 Mice. J. Investig. Dermatol. 2015, 135, 1638–1648. [Google Scholar] [CrossRef]
- Skobowiat, C.; Postlethwaite, A.E.; Slominski, A.T. Skin Exposure to Ultraviolet B Rapidly Activates Systemic Neuroendocrine and Immunosuppressive Responses. Photochem. Photobiol. 2017, 93, 1008–1015. [Google Scholar] [CrossRef] [PubMed]
- Dhabhar, F.S.; Saul, A.N.; Daugherty, C.; Holmes, T.H.; Bouley, D.M.; Oberyszyn, T.M. Short-Term Stress Enhances Cellular Immunity and Increases Early Resistance to Squamous Cell Carcinoma. Brain Behav. Immunity 2010, 24, 127–137. [Google Scholar] [CrossRef]
- Slominski, A.; Zbytek, B.; Zmijewski, M. Corticotropin Releasing Hormone and the Skin. Front. Biosci. 2006, 11, 2230. [Google Scholar] [CrossRef]
- Zbytek, B.; Slominski, A.T. CRH Mediates Inflammation Induced by Lipopolysaccharide in Human Adult Epidermal Keratinocytes. J. Investig. Dermatol. 2007, 127, 730–732. [Google Scholar] [CrossRef] [PubMed]
- Quevedo, M.-E.; Slominski, A.; Pinto, W.; Wei, E.; Wortsman, J. Pleiotropic Effects of Corticotropin Releasing Hormone on Normal Human Skin Keratinocytes. In Vitro Cell. Dev. Biol. Anim. 2001, 37, 50. [Google Scholar] [CrossRef]
- Zbytek, B.; Pfeffer, L.M.; Slominski, A.T. Corticotropin-Releasing Hormone Stimulates NF-ΚB in Human Epidermal Keratinocytes. J. Endocrinol. 2004, 181, R1–R7. [Google Scholar] [CrossRef]
- Slominski, A.; Wortsman, J.; Pisarchik, A.; Zbytek, B.; Linton, E.A.; Mazurkiewicz, J.E.; Wei, E.T. Cutaneous Expression of Corticotropin-Releasing Hormone (CRH), Urocortin, and CRH Receptors. FASEB J. 2001, 15, 1678–1693. [Google Scholar] [CrossRef]
- Fenix, K.; Wijesundara, D.K.; Cowin, A.J.; Grubor-Bauk, B.; Kopecki, Z. Immunological Memory in Imiquimod-Induced Murine Model of Psoriasiform Dermatitis. Int. J. Mol. Sci. 2020, 21, 7228. [Google Scholar] [CrossRef]
- Owczarczyk-Saczonek, A.; Krajewska-Włodarczyk, M.; Kasprowicz-Furmańczyk, M.; Placek, W. Immunological Memory of Psoriatic Lesions. Int. J. Mol. Sci. 2020, 21, 625. [Google Scholar] [CrossRef]
- Watanabe, R. Protective and Pathogenic Roles of Resident Memory T Cells in Human Skin Disorders. J. Dermatol. Sci. 2019, 95, 2–7. [Google Scholar] [CrossRef]
- Tokura, Y.; Phadungsaksawasdi, P.; Kurihara, K.; Fujiyama, T.; Honda, T. Pathophysiology of Skin Resident Memory T Cells. Front. Immunol. 2021, 11, 618897. [Google Scholar] [CrossRef]
- Patel, N.; Nadkarni, A.; Cardwell, L.A.; Vera, N.; Frey, C.; Patel, N.; Feldman, S.R. Psoriasis, Depression, and Inflammatory Overlap: A Review. Am. J. Clin. Dermatol. 2017, 18, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Matiushenko, V.; Kutasevych, Y.; Jafferany, M. Neurotransmitter Imbalance in Serum of Psoriatic Patients in Exacerbation Stage with Comorbid Psychoemotional Disorders. Dermatol. Ther. 2020, 33, e13337. [Google Scholar] [CrossRef] [PubMed]
- Baran, P.; Hansen, S.; Waetzig, G.H.; Akbarzadeh, M.; Lamertz, L.; Huber, H.J.; Ahmadian, M.R.; Moll, J.M.; Scheller, J. The Balance of Interleukin (IL)-6, IL-6·soluble IL-6 Receptor (SIL-6R), and IL-6·sIL-6R·sgp130 Complexes Allows Simultaneous Classic and Trans-Signaling. J. Biol. Chem. 2018, 293, 6762–6775. [Google Scholar] [CrossRef] [PubMed]
- Ganança, L.; Oquendo, M.A.; Tyrka, A.R.; Cisneros-Trujillo, S.; Mann, J.J.; Sublette, M.E. The Role of Cytokines in the Pathophysiology of Suicidal Behavior. Psychoneuroendocrinology 2016, 63, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Bergmans, R.S.; Kelly, K.M.; Mezuk, B. Inflammation as a Unique Marker of Suicide Ideation Distinct from Depression Syndrome among U.S. Adults. J. Affect. Disord. 2019, 245, 1052–1060. [Google Scholar] [CrossRef]
- Harden, J.L.; Lewis, S.M.; Lish, S.R.; Suárez-Fariñas, M.; Gareau, D.; Lentini, T.; Johnson-Huang, L.M.; Krueger, J.G.; Lowes, M.A. The Tryptophan Metabolism Enzyme L-Kynureninase Is a Novel Inflammatory Factor in Psoriasis and Other Inflammatory Diseases. J. Allergy Clin. Immunol. 2016, 137, 1830–1840. [Google Scholar] [CrossRef]
- Beurel, E.; Lowell, J.A. Th17 Cells in Depression. Brain Behav. Immunity 2018, 69, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, Y.; Wu, L.; Xiao, S.; Ji, Y.; Tan, Y.; Jiang, C.; Zhang, G. Dysregulation of the Gut-Brain-Skin Axis and Key Overlapping Inflammatory and Immune Mechanisms of Psoriasis and Depression. Biomed. Pharmacother. 2021, 137, 111065. [Google Scholar] [CrossRef]
- Raison, C.L.; Rutherford, R.E.; Woolwine, B.J.; Shuo, C.; Schettler, P.; Drake, D.F.; Haroon, E.; Miller, A.H. A Randomized Controlled Trial of the Tumor Necrosis Factor Antagonist Infliximab for Treatment-Resistant Depression: The Role of Baseline Inflammatory Biomarkers. JAMA Psychiatry 2013, 70, 31. [Google Scholar] [CrossRef]
- Rosenblat, J.D.; Cha, D.S.; Mansur, R.B.; McIntyre, R.S. Inflamed Moods: A Review of the Interactions between Inflammation and Mood Disorders. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 53, 23–34. [Google Scholar] [CrossRef]
- Pace, T.W.W.; Miller, A.H. Cytokines and Glucocorticoid Receptor Signaling: Relevance to Major Depression. Ann. N. Y. Acad. Sci. 2009, 1179, 86–105. [Google Scholar] [CrossRef]
- Herman, J.P.; Nawreen, N.; Smail, M.A.; Cotella, E.M. Brain Mechanisms of HPA Axis Regulation: Neurocircuitry and Feedback in Context Richard Kvetnansky Lecture. Stress 2020, 23, 617–632. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.T.; Khoo, B. Normal Physiology of ACTH and GH Release in the Hypothalamus and Anterior Pituitary in Man. Endotext 2000. [Google Scholar]
- Jiang, Z.; Rajamanickam, S.; Justice, N.J. CRF Signaling between Neurons in the Paraventricular Nucleus of the Hypothalamus (PVN) Coordinates Stress Responses. Neurobiol. Stress 2019, 11, 100192. [Google Scholar] [CrossRef]
- Kim, J.S.; Han, S.Y.; Iremonger, K.J. Stress Experience and Hormone Feedback Tune Distinct Components of Hypothalamic CRH Neuron Activity. Nat. Commun. 2019, 10, 5696. [Google Scholar] [CrossRef]
- Davami, M.; Baharlou, R.; Ahmadi Vasmehjani, A.; Ghanizadeh, A.; Keshtkar, M.; Dezhkam, I.; Atashzar, M. Elevated IL-17 and TGF-β Serum Levels: A Positive Correlation between T-Helper 17 Cell-Related Pro-Inflammatory Responses with Major Depressive Disorder. BCN 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Lotrich, F.E.; El-Gabalawy, H.; Guenther, L.C.; Ware, C.F. The Role of Inflammation in the Pathophysiology of Depression: Different Treatments and Their Effects. J. Rheumatol. Suppl. 2011, 88, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Kreibig, S.D. Autonomic Nervous System Activity in Emotion: A Review. Biol. Psychol. 2010, 84, 394–421. [Google Scholar] [CrossRef] [PubMed]
- Gimeno, D.; Kivimäki, M.; Brunner, E.J.; Elovainio, M.; De Vogli, R.; Steptoe, A.; Kumari, M.; Lowe, G.D.O.; Rumley, A.; Marmot, M.G.; et al. Associations of C-Reactive Protein and Interleukin-6 with Cognitive Symptoms of Depression: 12-Year Follow-up of the Whitehall II Study. Psychol. Med. 2009, 39, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, A.V.; Rivier, C.L. Regulation of the Hypothalamic-Pituitary-Adrenal Axis by Cytokines: Actions and Mechanisms of Action. Physiol. Rev. 1999, 79, 1–71. [Google Scholar] [CrossRef] [PubMed]
- Chrousos, G.P. Stress and Disorders of the Stress System. Nat. Rev. Endocrinol. 2009, 5, 374–381. [Google Scholar] [CrossRef]
- Kim, J.; Cho, B.; Cho, D.; Park, H. Expression of Hypothalamic–Pituitary–Adrenal Axis in Common Skin Diseases: Evidence of Its Association with Stress-Related Disease Activity. Acta Derm. Venerol. 2013, 93, 387–393. [Google Scholar] [CrossRef]
- Slominski, A.; Mihm, M.C. Potential Mechanism of Skin Response to Stress. Int. J. Dermatol. 1996, 35, 849–851. [Google Scholar] [CrossRef] [PubMed]
- Bellavance Marc-André; Serge Rivest The HPA—Immune Axis and the Immunomodulatory Actions of Glucocorticoids in the Brain. Front. Immunol. 2014, 5, 136. [CrossRef]
- Slominski, A.; Roloff, B.; Curry, J.; Dahiya, M.; Szczesniewski, A.; Wortsman, J. The Skin Produces Urocortin. J. Clin. Endocrinol. Metab. 2000, 85, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Pisarchik, A.; Tobin, D.J.; Mazurkiewicz, J.E.; Wortsman, J. Differential Expression of a Cutaneous Corticotropin-Releasing Hormone System. Endocrinology 2004, 145, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Zbytek, B.; Slominski, A.T. Corticotropin-Releasing Hormone Induces Keratinocyte Differentiation in the Adult Human Epidermis. J. Cell. Physiol. 2005, 203, 118–126. [Google Scholar] [CrossRef]
- Slominski, A.; Zbytek, B.; Pisarchik, A.; Slominski, R.M.; Zmijewski, M.A.; Wortsman, J. CRH Functions as a Growth Factor/Cytokine in the Skin. J. Cell. Physiol. 2006, 206, 780–791. [Google Scholar] [CrossRef] [PubMed]
- Zbytek, B.; Pfeffer, L.M.; Slominski, A.T. CRH Inhibits NF-ΚB Signaling in Human Melanocytes. Peptides 2006, 27, 3276–3283. [Google Scholar] [CrossRef]
- Tagen, M.; Stiles, L.; Kalogeromitros, D.; Gregoriou, S.; Kempuraj, D.; Makris, M.; Donelan, J.; Vasiadi, M.; Staurianeas, N.G.; Theoharides, T.C. Skin Corticotropin-Releasing Hormone Receptor Expression in Psoriasis. J. Investig. Dermatol. 2007, 127, 1789–1791. [Google Scholar] [CrossRef][Green Version]
- Paus, R.; Theoharides, T.C.; Arck, P.C. Neuroimmunoendocrine Circuitry of the ‘Brain-Skin Connection’. Trends Immunol. 2006, 27, 32–39. [Google Scholar] [CrossRef]
- Antoniewicz, J.; Nedoszytko, B.; Lange, M.; Wierzbicka, J.; Górska-Ponikowska, M.; Niedoszytko, M.; Zabłotna, M.; Nowicki, R.J.; Żmijewski, M.A. Modulation of Dermal Equivalent of Hypothalamus-Pituitary-Adrenal Axis in Mastocytosis. Postepy Dermatol. Alergol. 2021, 38, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Pariante, C.M. Why Are Depressed Patients Inflamed? A Reflection on 20 Years of Research on Depression, Glucocorticoid Resistance and Inflammation. Eur. Neuropsychopharmacol. 2017, 27, 554–559. [Google Scholar] [CrossRef]
- Cruz-Topete, D.; Cidlowski, J.A. One Hormone, Two Actions: Anti- and Pro-Inflammatory Effects of Glucocorticoids. Neuroimmunomodulation 2015, 22, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Traks, T.; Keermann, M.; Karelson, M.; Rätsep, R.; Reimann, E.; Silm, H.; Vasar, E.; Kõks, S.; Kingo, K. Polymorphisms in Corticotrophin-Releasing Hormone-Proopiomelanocortin (CRH-POMC) System Genes Are Associated with Plaque Psoriasis. Acta Derm. Venerol. 2019, 99, 444–445. [Google Scholar] [CrossRef]
- Slominski, A.; Zbytek, B.; Szczesniewski, A.; Semak, I.; Kaminski, J.; Sweatman, T.; Wortsman, J. CRH Stimulation of Corticosteroids Production in Melanocytes Is Mediated by ACTH. Am. J. Physiol.-Endocrinol. Metab. 2005, 288, E701–E706. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, H.; Jiang, B.; Yan, S.; Lu, J. A Promising Therapeutic Target for Psoriasis: Neuropeptides in Human Skin. Int. Immunopharmacol. 2020, 87, 106755. [Google Scholar] [CrossRef]
- Arck, P.C.; Slominski, A.; Theoharides, T.C.; Peters, E.M.J.; Paus, R. Neuroimmunology of Stress: Skin Takes Center Stage. J. Investig. Dermatol. 2006, 126, 1697–1704. [Google Scholar] [CrossRef]
- Harvima, I.T.; Nilsson, G.; Suttle, M.-M.; Naukkarinen, A. Is There a Role for Mast Cells in Psoriasis? Arch. Dermatol. Res. 2008, 300, 461–478. [Google Scholar] [CrossRef]
- Harvima, I.T.; Nilsson, G. Stress, the Neuroendocrine System and Mast Cells: Current Understanding of Their Role in Psoriasis. Expert Rev. Clin. Immunol. 2012, 8, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Neugebauer, V.; Mazzitelli, M.; Cragg, B.; Ji, G.; Navratilova, E.; Porreca, F. Amygdala, Neuropeptides, and Chronic Pain-Related Affective Behaviors. Neuropharmacology 2020, 170, 108052. [Google Scholar] [CrossRef]
- Huang, J.; Polgár, E.; Solinski, H.J.; Mishra, S.K.; Tseng, P.-Y.; Iwagaki, N.; Boyle, K.A.; Dickie, A.C.; Kriegbaum, M.C.; Wildner, H.; et al. Circuit Dissection of the Role of Somatostatin in Itch and Pain. Nat. Neurosci. 2018, 21, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Peters, E.M.J.; Liezmann, C.; Klapp, B.F.; Kruse, J. The Neuroimmune Connection Interferes with Tissue Regeneration and Chronic Inflammatory Disease in the Skin: Stress and Neuroimmune Plasticity. Ann. N. Y. Acad. Sci. 2012, 1262, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Kavalioti, M. Stress, Inflammation and Natural Treatments. J. Biol. Regul Homeost Agents 2018, 32, 1345–1347. [Google Scholar]
- Romana-Souza, B.; Otranto, M.; Almeida, T.F.; Porto, L.C.; Monte-Alto-Costa, A. Stress-Induced Epinephrine Levels Compromise Murine Dermal Fibroblast Activity through β-Adrenoceptors: Epinephrine-Stimulated Activation of β-Adrenoceptors Inhibits Dermal Fibroblast Activity. Exp. Dermatol. 2011, 20, 413–419. [Google Scholar] [CrossRef]
- Remröd, C.; Lonne-Rahm, S.; Nordlind, K. Study of Substance P and Its Receptor Neurokinin-1 in Psoriasis and Their Relation to Chronic Stress and Pruritus. Arch. Dermatol. Res. 2007, 299, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Alexopoulos, A.; Chrousos, G.P. Stress-Related Skin Disorders. Rev. Endocr. Metab. Disord. 2016, 17, 295–304. [Google Scholar] [CrossRef]
- Kramer, M.S.; Cutler, N.; Feighner, J.; Shrivastava, R.; Carman, J.; Sramek, J.J.; Reines, S.A.; Liu, G.; Snavely, D.; Wyatt-Knowles, E.; et al. Distinct Mechanism for Antidepressant Activity by Blockade of Central Substance P Receptors. Science 1998, 281, 1640–1645. [Google Scholar] [CrossRef] [PubMed]
- Reichardt, L.F. Neurotrophin-Regulated Signalling Pathways. Phil. Trans. R. Soc. B 2006, 361, 1545–1564. [Google Scholar] [CrossRef]
- JiaWen, W.; Hong, S.; ShengXiang, X.; Jing, L. Depression- and Anxiety-like Behaviour Is Related to BDNF/TrkB Signalling in a Mouse Model of Psoriasis. Clin. Exp. Dermatol. 2018, 43, 254–261. [Google Scholar] [CrossRef]
- Udina, M.; Navinés, R.; Egmond, E.; Oriolo, G.; Langohr, K.; Gimenez, D.; Valdés, M.; Gómez-Gil, E.; Grande, I.; Gratacós, M.; et al. Glucocorticoid Receptors, Brain-Derived Neurotrophic Factor, Serotonin and Dopamine Neurotransmission Are Associated with Interferon-Induced Depression. IJNPPY 2016, 19, pyv135. [Google Scholar] [CrossRef]
- Roy, M.; Tapadia, M.G.; Joshi, S.; Koch, B. Molecular and Genetic Basis of Depression. J. Genet. 2014, 93, 879–892. [Google Scholar] [CrossRef]
- Tejeda, G.; Díaz-Guerra, M. Integral Characterization of Defective BDNF/TrkB Signalling in Neurological and Psychiatric Disorders Leads the Way to New Therapies. IJMS 2017, 18, 268. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Wang, L.; Clark, J.D.; Kingery, W.S. Keratinocytes Express Cytokines and Nerve Growth Factor in Response to Neuropeptide Activation of the ERK1/2 and JNK MAPK Transcription Pathways. Regul. Pept. 2013, 186, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Jiang, T.; Chen, P.; Ouyang, J.; Xu, G.; Zeng, Z.; Sun, Y. Emerging Tendency towards Autoimmune Process in Major Depressive Patients: A Novel Insight from Th17 Cells. Psychiatry Research 2011, 188, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.E.; Di Nardo, A. Skin Neurogenic Inflammation. Semin. Immunopathol. 2018, 40, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, R.; Ghosh, A.K.; Ghosh, B.; Bhattacharyya, S.; Mondal, A.C. Decreased MRNA and Protein Expression of BDNF, NGF, and Their Receptors in the Hippocampus from Suicide: An Analysis in Human Postmortem Brain. Clin. Med. Insights Pathol. 2013, 6. [Google Scholar] [CrossRef]
- Dwivedi, Y.; Mondal, A.C.; Rizavi, H.S.; Conley, R.R. Suicide Brain Is Associated with Decreased Expression of Neurotrophins. Biol. Psychiatry 2005, 58, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Kechichian, E.; Ezzedine, K. Vitamin D and the Skin: An Update for Dermatologists. Am. J. Clin. Dermatol. 2018, 19, 223–235. [Google Scholar] [CrossRef]
- Bocheva, G.; Slominski, R.M.; Slominski, A.T. The Impact of Vitamin D on Skin Aging. IJMS 2021, 22, 9097. [Google Scholar] [CrossRef]
- Reichrath, J.; Saternus, R.; Vogt, T. Endocrine Actions of Vitamin D in Skin: Relevance for Photocarcinogenesis of Non-Melanoma Skin Cancer, and Beyond. Mol. Cell. Endocrinol. 2017, 453, 96–102. [Google Scholar] [CrossRef]
- Wirz-Justice, A.; Skene, D.J.; Münch, M. The Relevance of Daylight for Humans. Biochem. Pharmacol. 2021, 191, 114304. [Google Scholar] [CrossRef]
- Holick, M.F. The Vitamin D Deficiency Pandemic: Approaches for Diagnosis, Treatment and Prevention. Rev. Endocr. Metab. Disord. 2017, 18, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Kim, T.; Shehabi, H.Z.; Semak, I.; Tang, E.K.Y.; Nguyen, M.N.; Benson, H.A.E.; Korik, E.; Janjetovic, Z.; Chen, J.; et al. In Vivo Evidence for a Novel Pathway of Vitamin D3 Metabolism Initiated by P450scc and Modified by CYP27B1. FASEB J. 2012, 26, 3901–3915. [Google Scholar] [CrossRef]
- Bikle, D.; Christakos, S. New Aspects of Vitamin D Metabolism and Action—Addressing the Skin as Source and Target. Nat. Rev. Endocrinol. 2020, 16, 234–252. [Google Scholar] [CrossRef] [PubMed]
- Hanel, A.; Carlberg, C. Vitamin D and Evolution: Pharmacologic Implications. Biochem. Pharmacol. 2020, 173, 113595. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Kim, T.; Takeda, Y.; Janjetovic, Z.; Brożyna, A.A.; Skobowiat, C.; Wang, J.; Postlethwaite, A.; Li, W.; Tuckey, R.C.; et al. RORα and ROR γ Are Expressed in Human Skin and Serve as Receptors for Endogenously Produced Noncalcemic 20-hydroxy- and 20,23-dihydroxyvitamin D. FASEB J. 2014, 28, 2775–2789. [Google Scholar] [CrossRef]
- Slominski, A.; Kim, T.-K.; Janjetovic, Z.; Brożyna, A.; Żmijewski, M.; Xu, H.; Sutter, T.; Tuckey, R.; Jetten, A.; Crossman, D. Differential and Overlapping Effects of 20,23(OH)2D3 and 1,25(OH)2D3 on Gene Expression in Human Epidermal Keratinocytes: Identification of AhR as an Alternative Receptor for 20,23(OH)2D3. IJMS 2018, 19, 3072. [Google Scholar] [CrossRef]
- Slominski, A.T.; Kim, T.-K.; Qayyum, S.; Song, Y.; Janjetovic, Z.; Oak, A.S.W.; Slominski, R.M.; Raman, C.; Stefan, J.; Mier-Aguilar, C.A.; et al. Vitamin D and Lumisterol Derivatives Can Act on Liver X Receptors (LXRs). Sci. Rep. 2021, 11, 8002. [Google Scholar] [CrossRef]
- Umar, M.; Sastry, K.S.; Al Ali, F.; Al-Khulaifi, M.; Wang, E.; Chouchane, A.I. Vitamin D and the Pathophysiology of Inflammatory Skin Diseases. Skin. Pharmacol. Physiol. 2018, 31, 74–86. [Google Scholar] [CrossRef]
- Piotrowska, A.; Wierzbicka, J.; Żmijewski, M.A. Vitamin D in the Skin Physiology and Pathology. Acta Biochim. Pol. 2016, 63, 17–29. [Google Scholar] [CrossRef]
- Slominski, A.T.; Chaiprasongsuk, A.; Janjetovic, Z.; Kim, T.-K.; Stefan, J.; Slominski, R.M.; Hanumanthu, V.S.; Raman, C.; Qayyum, S.; Song, Y.; et al. Photoprotective Properties of Vitamin D and Lumisterol Hydroxyderivatives. Cell Biochem. Biophys. 2020, 78, 165–180. [Google Scholar] [CrossRef]
- Barrea, L.; Savanelli, M.C.; Di Somma, C.; Napolitano, M.; Megna, M.; Colao, A.; Savastano, S. Vitamin D and Its Role in Psoriasis: An Overview of the Dermatologist and Nutritionist. Rev. Endocr. Metab. Disord. 2017, 18, 195–205. [Google Scholar] [CrossRef]
- Megna, M.; Ferrillo, M.; Barrea, L.; Patruno, C.; Muscogiuri, G.; Savastano, S.; Fabbrocini, G.; Napolitano, M. Vitamin D and Psoriasis: An Update for Dermatologists and Nutritionists. Minerva Endocrinol. 2020, 45, 138–147. [Google Scholar] [CrossRef]
- Zhao, Y.; Asahina, A.; Asawanonda, P.; Frez, M.L.; Imafuku, S.; Hyun Kim, D.; Theng, C.; Wang, L.; Zhang, J.A.; Zimmo, S. Systematic Review and Practical Guidance on the Use of Topical Calcipotriol and Topical Calcipotriol with Betamethasone Dipropionate as Long-term Therapy for Mild-to-moderate Plaque Psoriasis. J. Dermatol. 2021, 48, 940–960. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, C.; Wang, J.; Chen, K.; Ding, Y.; Yan, G.; Lu, Q.; Li, W.; Fang, H.; Cheng, H.; et al. Comparison of Safety and Efficacy between Calcipotriol plus Betamethasone Dipropionate Gel and Calcipotriol Scalp Solution as Long-Term Treatment for Scalp Psoriasis in Chinese Patients: A National, Multicentre, Prospective, Randomized, Active-Controlled Phase 4 Trial. Eur. J. Dermatol. 2020, 30, 580–590. [Google Scholar] [CrossRef]
- Stanescu, A.M.A.; Simionescu, A.A.; Diaconu, C.C. Oral Vitamin D Therapy in Patients with Psoriasis. Nutrients 2021, 13, 163. [Google Scholar] [CrossRef] [PubMed]
- Theodoridis, X.; Grammatikopoulou, M.G.; Stamouli, E.-M.; Talimtzi, P.; Pagkalidou, E.; Zafiriou, E.; Haidich, A.-B.; Bogdanos, D.P. Effectiveness of Oral Vitamin D Supplementation in Lessening Disease Severity among Patients with Psoriasis: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrition 2021, 82, 111024. [Google Scholar] [CrossRef] [PubMed]
- Jalili, A.; Yosipovitch, G. Fixed-dose Combination Calcipotriol/Betamethasone Dipropionate Foam Provides a Rapid Onset of Action, Effective Itch Relief and Improves Patient Quality of Life. J. Eur. Acad. Dermatol. Venereol. 2021, 35, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Pink, A.E.; Jalili, A.; Berg, P.; Calzavara-Pinton, P.G.; Cueva Dobao, P.; Thaçi, D.; Torpet, M.; Jensen, K.L.; Segaert, S. Rapid Onset of Action of Calcipotriol/Betamethasone Dipropionate Cutaneous Foam in Psoriasis, Even in Patients with More Severe Disease. J. Eur. Acad. Dermatol. Venereol. 2019, 33, 1116–1123. [Google Scholar] [CrossRef]
- McCullough, P.; Amend, J. Results of Daily Oral Dosing with up to 60,000 International Units (Iu) of Vitamin D3 for 2 to 6 Years in 3 Adult Males. J. Steroid Biochem. Mol. Biol. 2017, 173, 308–312. [Google Scholar] [CrossRef]
- Warren, R.; Gold, M.; Gooderham, M.; Kircik, L.; Lacour, J.-P.; Laws, P.; Liljedahl, M.; Lynde, C.; Mørch, M.; Sondermann, W.; et al. Four-Week Daily Calcipotriene/Betamethasone Dipropionate Foam Is Highly Efficacious in Patients With Psoriasis (PSO-LONG Lead-in Phase). JDD 2021, 20, 436–441. [Google Scholar] [CrossRef]
- Stein Gold, L.; Green, L.; Dhawan, S.; Vestbjerg, B.; Praestegaard, M.; Selmer, J. A Phase 3, Randomized Trial Demonstrating the Improved Efficacy and Patient Acceptability of Fixed Dose Calcipotriene and Betamethasone Dipropionate Cream. JDD 2021, 20, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Zmijewski, M.A. Vitamin D and Human Health. IJMS 2019, 20, 145. [Google Scholar] [CrossRef] [PubMed]
- Parker, G.B.; Brotchie, H.; Graham, R.K. Vitamin D and Depression. J. Affect. Disord. 2017, 208, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Bang, M.; Wee, J.H.; Min, C.; Yoo, D.M.; Han, S.-M.; Kim, S.; Choi, H.G. Short- and Long-Term Exposure to Air Pollution and Lack of Sunlight Are Associated with an Increased Risk of Depression: A Nested Case-Control Study Using Meteorological Data and National Sample Cohort Data. Sci. Total Environ. 2021, 757, 143960. [Google Scholar] [CrossRef]
- Saleem, S.; Iqubal, M.K.; Garg, S.; Ali, J.; Baboota, S. Trends in Nanotechnology-Based Delivery Systems for Dermal Targeting of Drugs: An Enticing Approach to Offset Psoriasis. Expert Opin. Drug Deliv. 2020, 17, 817–838. [Google Scholar] [CrossRef]
- Damiani, G.; Pacifico, A.; Linder, D.M.; Pigatto, P.D.M.; Conic, R.; Grada, A.; Bragazzi, N.L. Nanodermatology-Based Solutions for Psoriasis: State-of-the Art and Future Prospects. Dermatol. Ther. 2019, 32, e13113. [Google Scholar] [CrossRef]
- Ramanunny, A.K.; Wadhwa, S.; Thakur, D.; Singh, S.K.; Kumar, R. Treatment Modalities of Psoriasis: A Focus on Requisite for Topical Nanocarrier. Endocr. Metab. Immune Disord. Drug Targets 2021, 21, 418–433. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marek-Jozefowicz, L.; Czajkowski, R.; Borkowska, A.; Nedoszytko, B.; Żmijewski, M.A.; Cubała, W.J.; Slominski, A.T. The Brain–Skin Axis in Psoriasis—Psychological, Psychiatric, Hormonal, and Dermatological Aspects. Int. J. Mol. Sci. 2022, 23, 669. https://doi.org/10.3390/ijms23020669
Marek-Jozefowicz L, Czajkowski R, Borkowska A, Nedoszytko B, Żmijewski MA, Cubała WJ, Slominski AT. The Brain–Skin Axis in Psoriasis—Psychological, Psychiatric, Hormonal, and Dermatological Aspects. International Journal of Molecular Sciences. 2022; 23(2):669. https://doi.org/10.3390/ijms23020669
Chicago/Turabian StyleMarek-Jozefowicz, Luiza, Rafał Czajkowski, Alina Borkowska, Bogusław Nedoszytko, Michał A. Żmijewski, Wiesław J. Cubała, and Andrzej T. Slominski. 2022. "The Brain–Skin Axis in Psoriasis—Psychological, Psychiatric, Hormonal, and Dermatological Aspects" International Journal of Molecular Sciences 23, no. 2: 669. https://doi.org/10.3390/ijms23020669
APA StyleMarek-Jozefowicz, L., Czajkowski, R., Borkowska, A., Nedoszytko, B., Żmijewski, M. A., Cubała, W. J., & Slominski, A. T. (2022). The Brain–Skin Axis in Psoriasis—Psychological, Psychiatric, Hormonal, and Dermatological Aspects. International Journal of Molecular Sciences, 23(2), 669. https://doi.org/10.3390/ijms23020669