
Glucocorticoid Regulates the Synthesis of Porcine Muscle Protein through m6A Modified Amino Acid Transporter SLC7A7


Abstract
:1. Introduction
2. Results
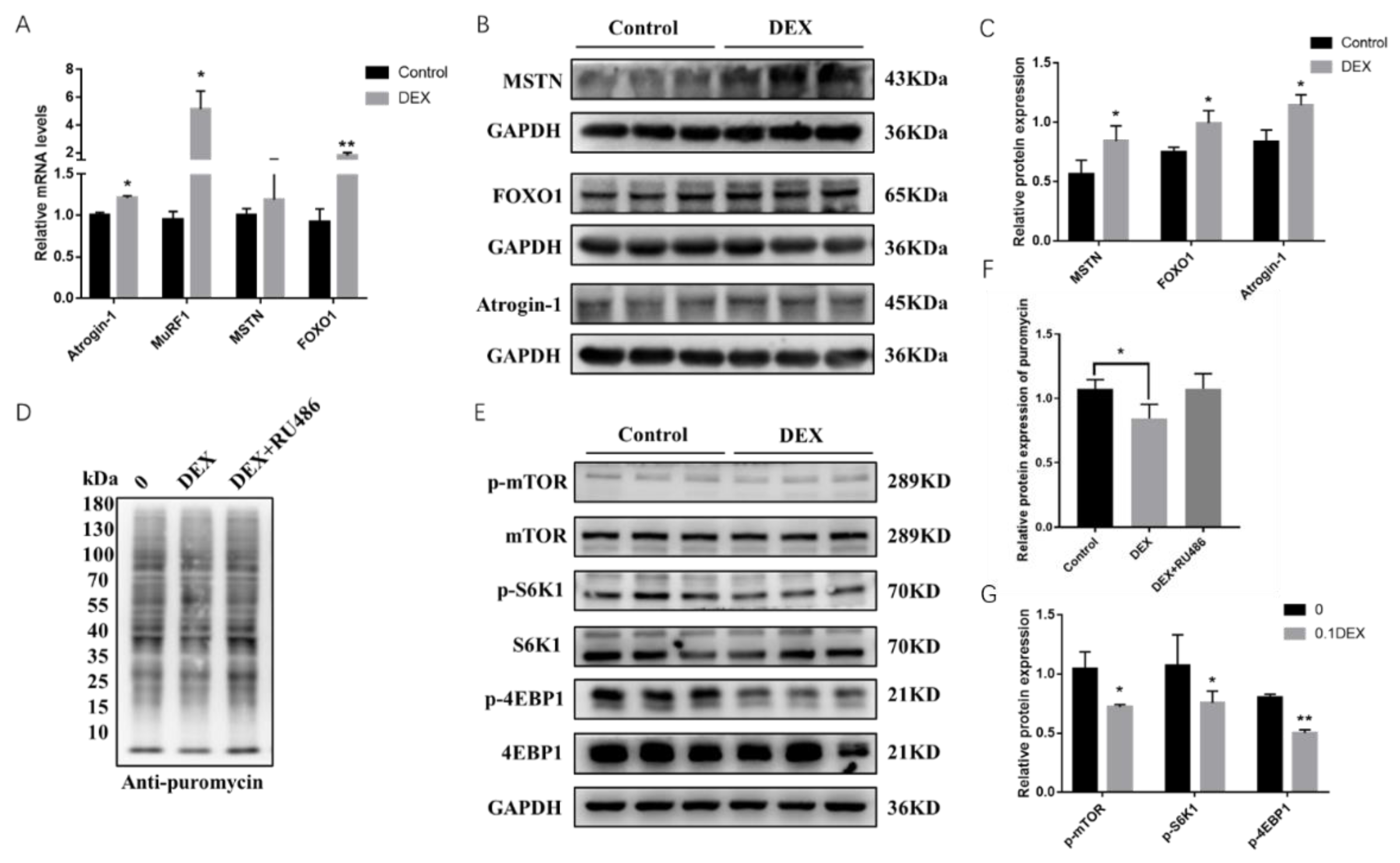
2.1. Effects of Glucocorticoid (GC) and RU486 on Protein Synthesis in Porcine Skeletal Muscle Cells
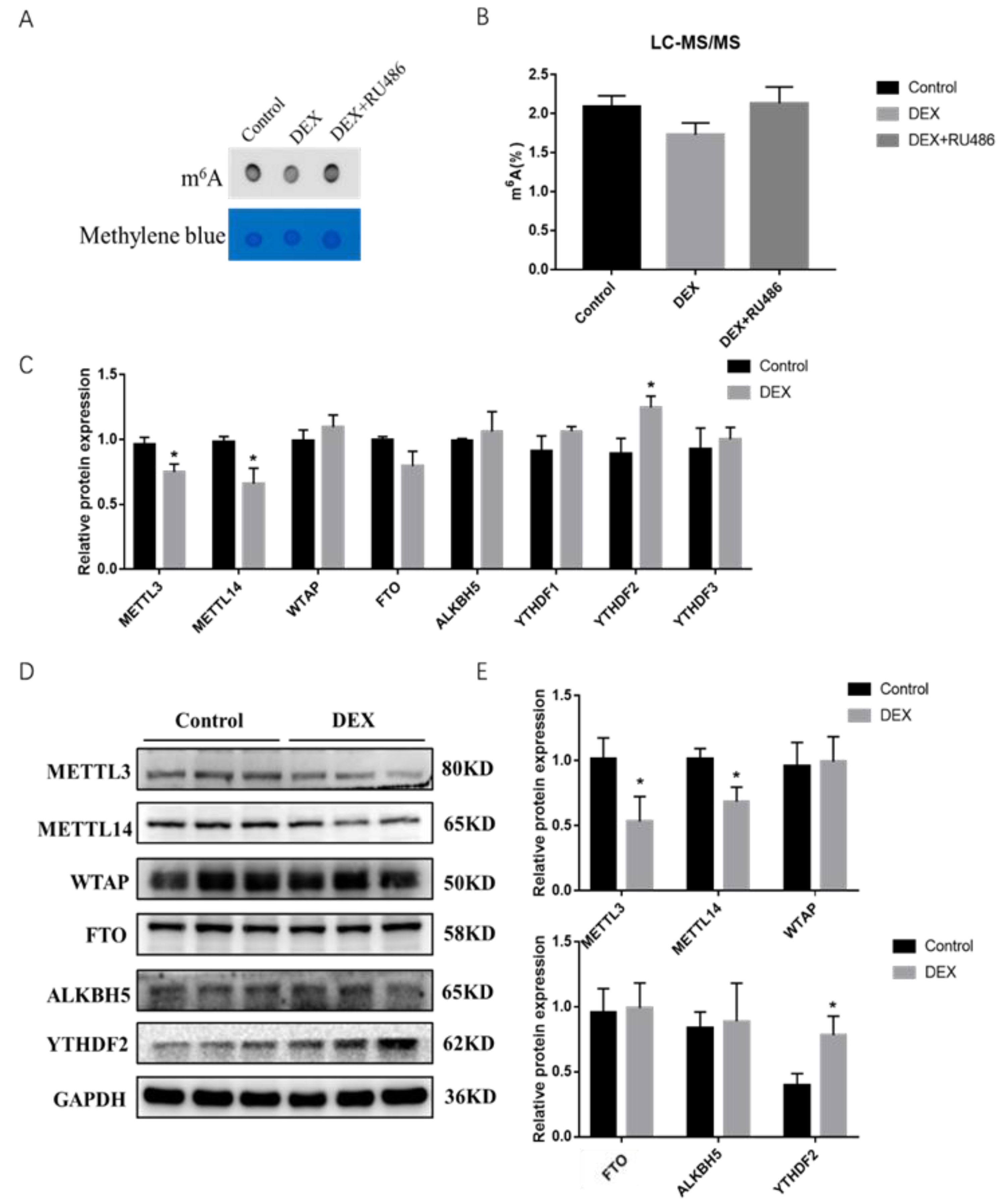
2.2. Effects of GCs and RU486 on Intracellular m6A Levels in Porcine Skeletal Muscle Cells
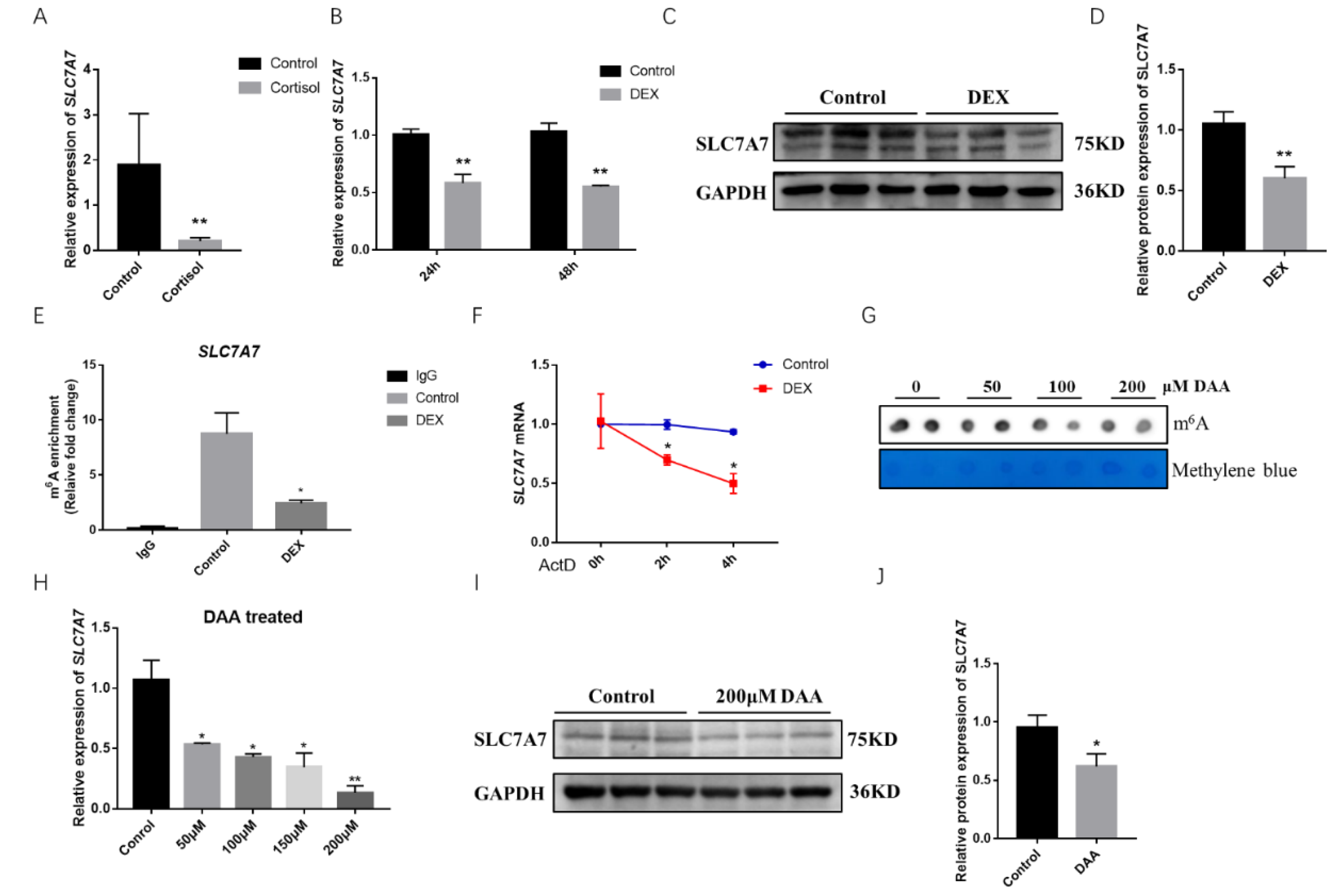
2.3. Effect of GCs on the Amino Acid Transporter SLC7A7
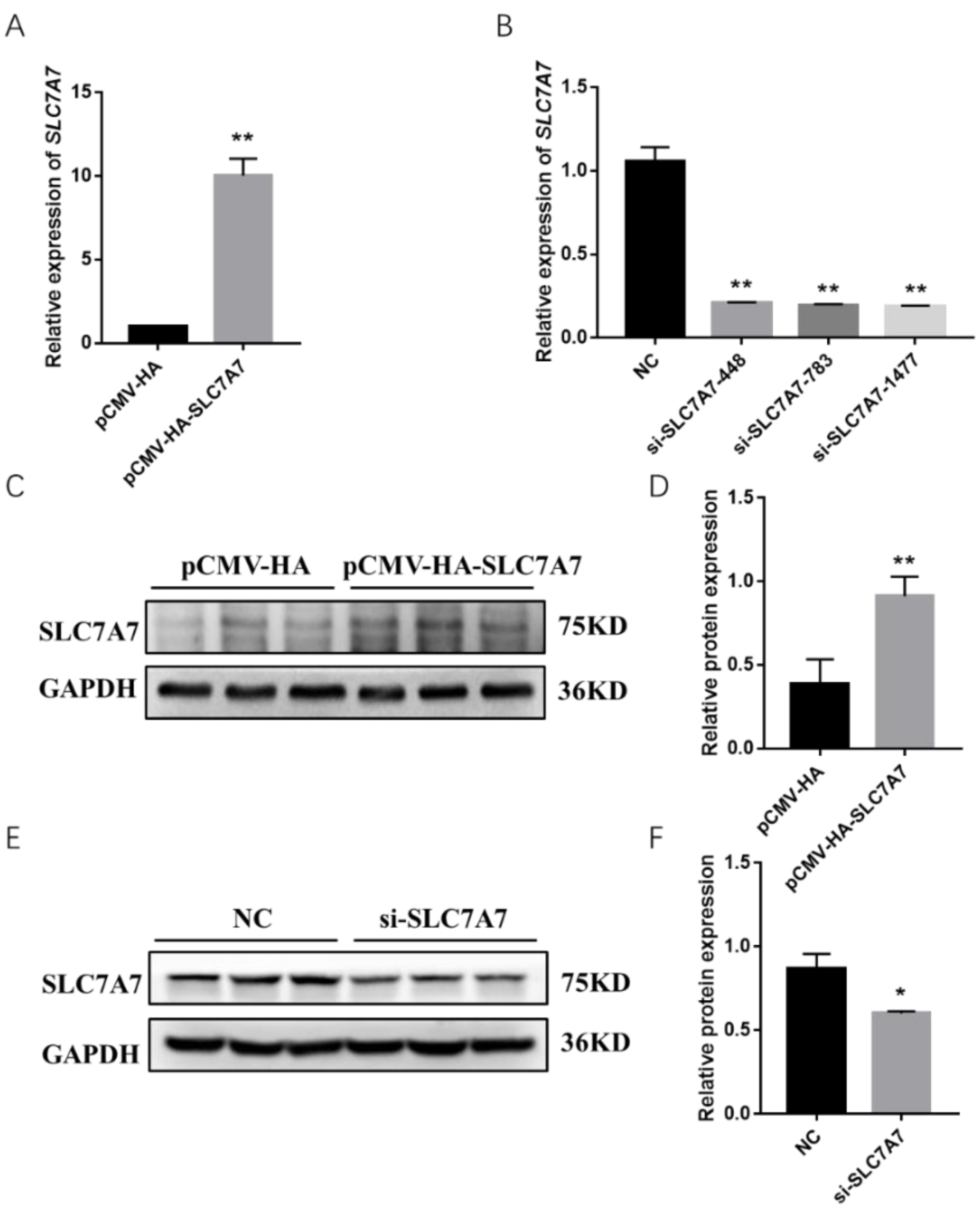
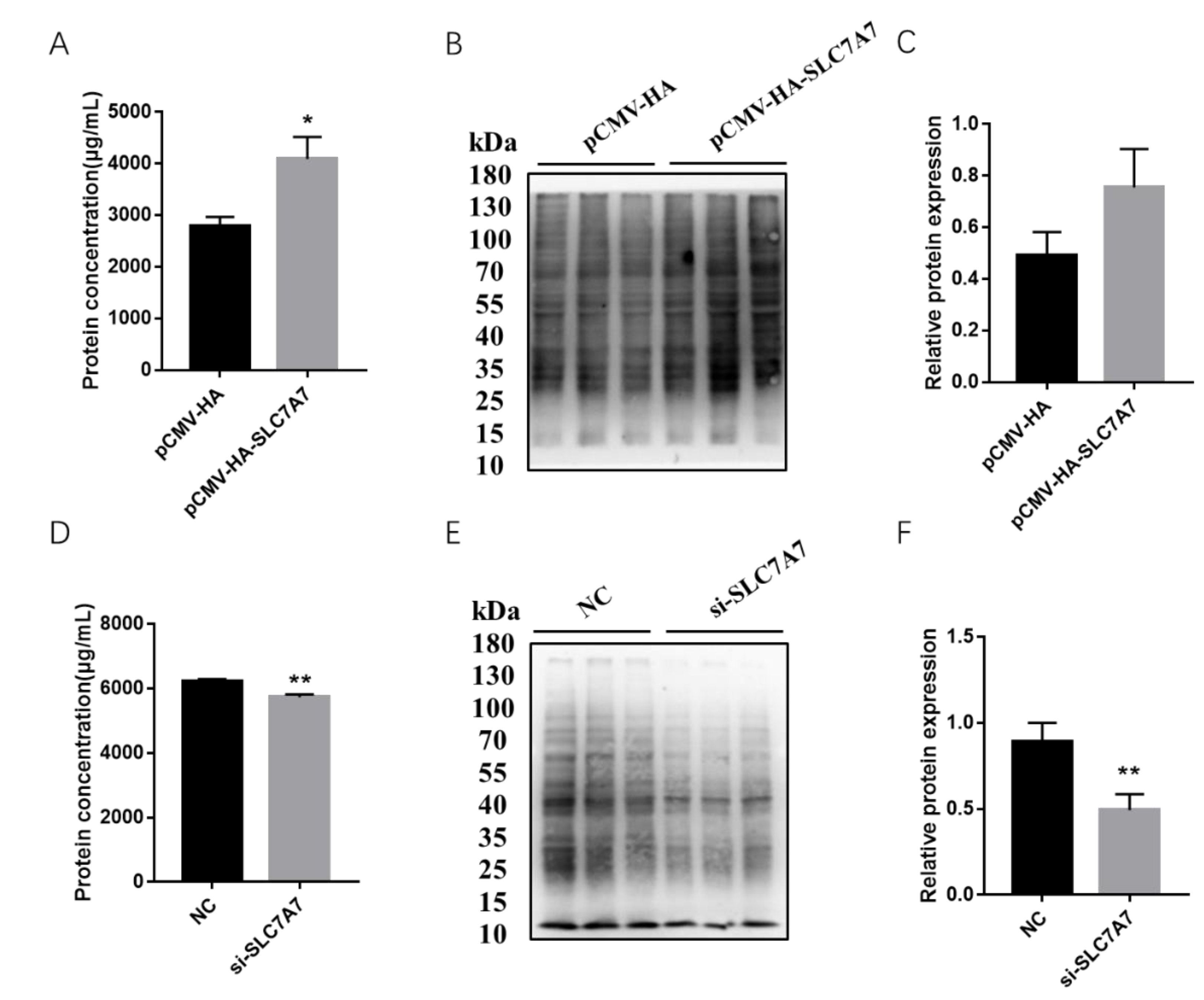
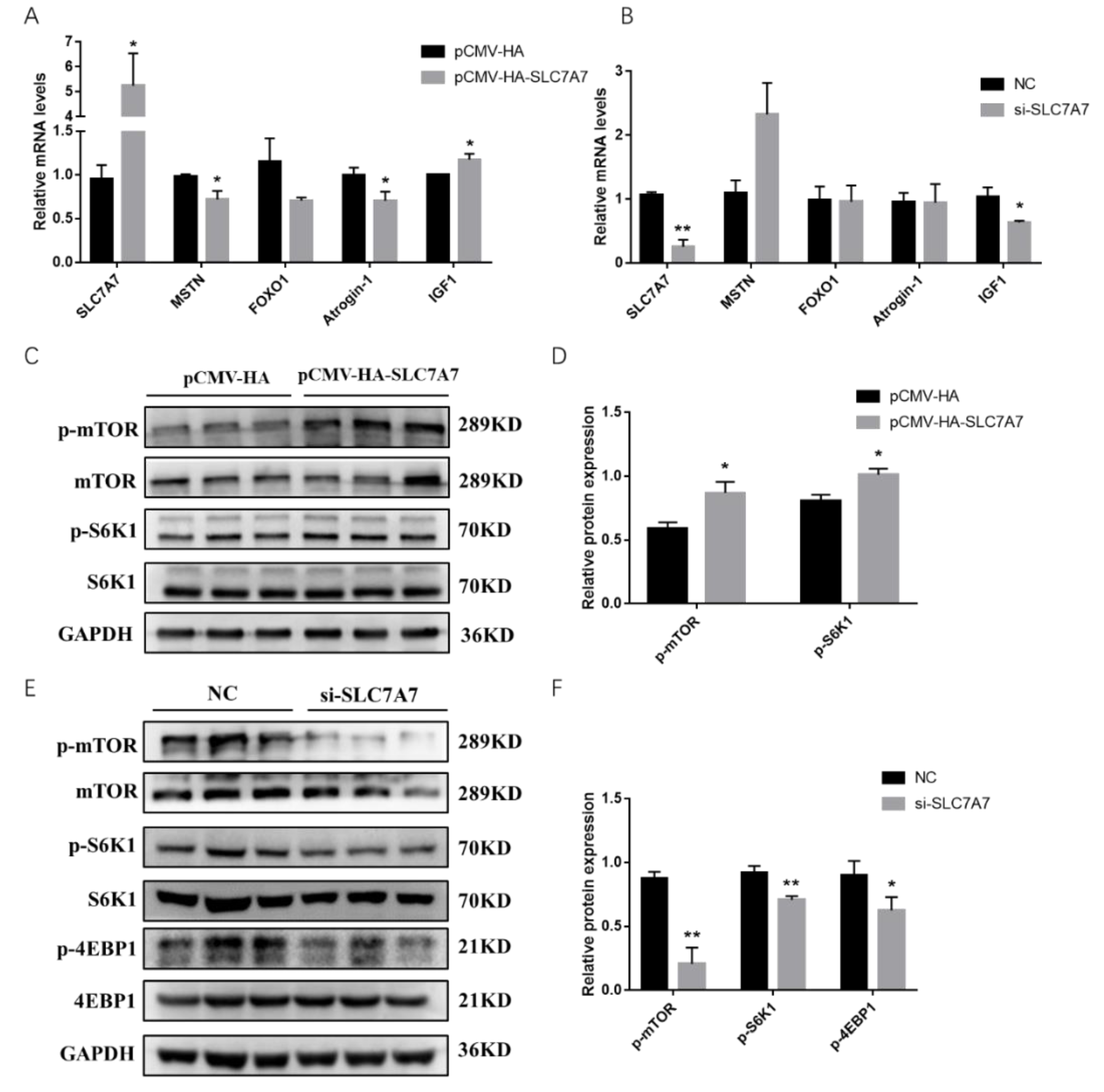
2.4. Effect of SLC7A7 on Protein Synthesis in Porcine Skeletal Muscle Cells
3. Discussion
4. Materials and Methods
4.1. Animals and Samples
4.2. Total RNA Preparation and cDNA Synthesis
4.3. Quantitative Real-Time Polymerase Chain Reaction (PCR)
4.4. BCA Method for Detecting Protein Concentration
4.5. Plasmid Construction, siRNA Synthesis and Cell Transfection
4.6. Western Blot Analysis
4.7. Surface Sensing of Translation (SUnSET) Non-Radioactive Method for Detecting Protein Synthesis Rate
4.8. m6A Immunoprecipitation (MeRIP) qPCR
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Li, Y.; Wang, P.; Yin, J.; Jin, S.; Su, W.; Tian, J.; Li, T.; Yao, K. Effects of ornithine alpha-ketoglutarate on growth performance and gut microbiota in a chronic oxidative stress pig model induced by d-galactose. Food Funct. 2020, 11, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Gjerstad, J.K.; Lightman, S.L.; Spiga, F. Role of glucocorticoid negative feedback in the regulation of HPA axis pulsatility. Stress 2018, 21, 403–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reser, J.E. Chronic stress, cortical plasticity and neuroecology. Behav. Processes 2016, 129, 105–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oyola, M.G.; Handa, R.J. Hypothalamic-pituitary-adrenal and hypothalamic-pituitary-gonadal axes: Sex differences in regulation of stress responsivity. Stress 2017, 20, 476–494. [Google Scholar] [CrossRef]
- Löfberg, E.; Gutierrez, A.; Wernerman, J.; Anderstam, B.; Mitch, W.E.; Price, S.R.; Bergström, J.; Alvestrand, A. Effects of high doses of GCs on free amino acids, ribosomes and protein turnover in human muscle. Eur. J. Clin. Invest. 2002, 32, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Tipton, K.D.; Gurkin, B.E.; Matin, S.; Wolfe, R.R. Nonessential amino acids are not necessary to stimulate net muscle protein synthesis in healthy volunteers. J. Nutr. Biochem. 1999, 10, 89–95. [Google Scholar] [CrossRef]
- Javed, K.; Fairweather, S.J. Amino acid transporters in the regulation of insulin secretion and signalling. Biochem. Soc. Trans. 2019, 47, 571–590. [Google Scholar] [CrossRef] [Green Version]
- Ji, X.; Yang, X.; Wang, N.; Kang, M.; Wang, Y.; Rong, L.; Fang, Y.; Xue, Y. Function of SLC7A7 in T-Cell Acute Lymphoblastic Leukemia. Cell. Physiol. Biochem. 2018, 48, 731–740. [Google Scholar] [CrossRef]
- Ishida, A.; Ashihara, A.; Nakashima, K.; Katsumata, M. Expression of cationic amino acid transporters in pig skeletal muscles during postnatal development. Amino Acids 2017, 49, 1805–1814. [Google Scholar] [CrossRef]
- de Baulny, H.O.; Schiff, M.; Dionisi-Vici, C. Lysinuric protein intolerance (LPI): A multi organ disease by far more complex than a classic urea cycle disorder. Mol. Genet. Metab. 2012, 106, 12–17. [Google Scholar] [CrossRef]
- Sperandeo, M.P.; Annunziata, P.; Bozzato, A.; Piccolo, P.; Maiuri, L.; D’Armiento, M.; Ballabio, A.; Corso, G.; Andria, G.; Borsani, G.; et al. Slc7a7 disruption causes fetal growth retardation by downregulating Igf1 in the mouse model of lysinuric protein intolerance. Am. J. Physiol.-Cell Physiol. 2007, 293, C191–C198. [Google Scholar] [CrossRef] [Green Version]
- Bodoy, S.; Sotillo, F.; Espino-Guarch, M.; Sperandeo, M.P.; Ormazabal, A.; Zorzano, A.; Sebastio, G.; Artuch, R.; Palacín, M. Inducible Slc7a7 Knockout Mouse Model Recapitulates Lysinuric Protein Intolerance Disease. Int. J. Mol. Sci. 2019, 20, 5294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engel, M.; Eggert, C.; Kaplick, P.M.; Eder, M.; Roh, S.; Tietze, L.; Namendorf, C.; Arloth, J.; Weber, P.; Rex-Haffner, M.; et al. The Role of m(6)A/m-RNA Methylation in Stress Response Regulation. Neuron 2018, 99, 389–403. [Google Scholar] [CrossRef]
- Heng, J.H.; Tian, M.; Zhang, W.F.; Chen, F.; Guan, W.T.; Zhang, S.H. Maternal heat stress regulates the early fat deposition partly through modification of m(6)A RNA methylation in neonatal piglets. Cell Stress Chaperones 2019, 24, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Li, Y.; Wang, T.; Zhong, X. Modification of N6-methyladenosine RNA methylation on heat shock protein expression. PLoS ONE 2018, 13, e0198604. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Z.; Yue, Y.N.; Han, D.L.; Wang, X.; Fu, Y.; Zhang, L.; Jia, G.F.; Yu, M.; Lu, Z.K.; Deng, X.; et al. A METTL3-METTL14 complex mediates mammalian nuclear RNA N-6-adenosine methylation. Nat. Chem. Biol. 2014, 10, 93–95. [Google Scholar] [CrossRef] [Green Version]
- Ping, X.L.; Sun, B.F.; Wang, L.; Xiao, W.; Yang, X.; Wang, W.J.; Adhikari, S.; Shi, Y.; Lv, Y.; Chen, Y.S.; et al. Mammalian WTAP is a regulatory subunit of the RNA N6-methyladenosine methyltransferase. Cell Res. 2014, 24, 177–189. [Google Scholar] [CrossRef] [Green Version]
- Zhong, H.; Tang, H.F.; Kai, Y. N6-methyladenine RNA Modification (m(6)A): An Emerging Regulator of Metabolic Diseases. Curr. Drug Targets 2020, 21, 1056–1067. [Google Scholar] [CrossRef]
- Dallman, M.F.; Akana, S.F.; Strack, A.M.; Hanson, E.S.; Sebastian, R.J. The neural network that regulates energy balance is responsive to GCs and insulin and also regulates HPA axis responsivity at a site proximal to CRF neurons. Ann. N. Y. Acad. Sci. 1995, 29, 730–742. [Google Scholar] [CrossRef]
- Chang, W.H.; Li, J.J.; Zhang, S.; Zheng, A.J.; Yuan, J.L.; Cai, H.Y.; Liu, G.H. Effects of glucocorticoid-induced stress on absorption of glycylsarcosine in jejunum of broilers. Poult. Sci. 2015, 94, 700–705. [Google Scholar] [CrossRef]
- Zhao, L.D.; McMillan, R.P.; Xie, G.H.; Giridhar, S.; Raumgard, L.H.; El-Kadi, S.; Selsby, J.; Ross, J.; Gabler, N.; Hulver, M.W.; et al. Heat stress decreases metabolic flexibility in skeletal muscle of growing pigs. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2018, 315, R1096–R1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sartorelli, V.; Fulco, M. Molecular and cellular determinants of skeletal muscle atrophy and hypertrophy. Sci. STKE Signal Transduct. Knowl. Environ. 2004, 2004, re11. [Google Scholar] [CrossRef] [PubMed]
- Bonaldo, P.; Sandri, M. Cellular and molecular mechanisms of muscle atrophy. Dis. Models Mech. 2013, 6, 25–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schakman, O.; Kalista, S.; Barbe, C.; Loumaye, A.; Thissen, J.P. Glucocorticoid-induced skeletal muscle atrophy. Int. J. Biochem. Cell Biol. 2013, 45, 2163–2172. [Google Scholar] [CrossRef] [PubMed]
- Rom, O.; Reznick, A.Z. The role of E3 ubiquitin-ligases MuRF-1 and MAFbx in loss of skeletal muscle mass. Free. Radic. Biol. Med. 2016, 98, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Clarke, B.A.; Drujan, D.; Willis, M.S.; Murphy, L.O.; Corpina, R.A.; Burova, E.; Rakhilin, S.V.; Stitt, T.N.; Patterson, C.; Latres, E.; et al. The E3 ligase MuRF1 degrades myosin heavy chain protein in dexamethasone-treated skeletal muscle. Cell Metab. 2007, 6, 376–385. [Google Scholar] [CrossRef] [Green Version]
- Kuci, O.; Archambault, E.; Dodacki, A.; Nubret, E.; De Bandt, J.P.; Cynober, L. Effect of citrulline on muscle protein turnover in an in vitro model of muscle catabolism. Nutrition 2020, 71, 110597. [Google Scholar] [CrossRef]
- Bodine, S.C.; Stitt, T.N.; Gonzalez, M.; Kline, W.O.; Stover, G.L.; Bauerlein, R.; Zlotchenko, E.; Scrimgeour, A.; Lawrence, J.C.; Glass, D.J.; et al. Akt/mTOR pathway is a crucial regulator of skeletal muscle hypertrophy and can prevent muscle atrophy in vivo. Nat. Cell Biol. 2001, 3, 1014–1019. [Google Scholar] [CrossRef]
- Kim, J.; Park, M.Y.; Kim, H.K.; Park, Y.; Whang, K.Y. Cortisone and dexamethasone inhibit myogenesis by modulating the AKT/mTOR signaling pathway in C2C12. Biosci. Biotechnol. Biochem. 2016, 80, 2093–2099. [Google Scholar] [CrossRef] [Green Version]
- Shah, O.J.; Kimball, S.R.; Jefferson, L.S. Acute attenuation of translation initiation and protein synthesis by GCs in skeletal muscle. Am. J. Physiol.-Endocrinol. Metab. 2000, 278, E76–E82. [Google Scholar] [CrossRef]
- Shah, O.J.; Kimball, S.R.; Jefferson, L.S. Among translational effecters, p70(S6k) is uniquely sensitive to inhibition by GCs. Biochem. J. 2000, 347, 389–397. [Google Scholar] [CrossRef]
- Jellyman, J.K.; Martin-Gronert, M.S.; Cripps, R.L.; Giussani, D.A.; Ozanne, S.E.; Shen, Q.W.W.; Du, M.; Fowden, A.L.; Forhead, A.J. Effects of Cortisol and Dexamethasone on Insulin Signalling Pathways in Skeletal Muscle of the Ovine Fetus during Late Gestation. PLoS ONE 2012, 7, e52363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joha, S.; Nugues, A.L.; Hetuin, D.; Berthon, C.; Dezitter, X.; Dauphin, V.; Mahon, F.X.; Roche-Lestienne, C.; Preudhomme, C.; Quesnel, B.; et al. GILZ inhibits the mTORC2/AKT pathway in BCR-ABL(+) cells. Oncogene 2012, 31, 1419–1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.K.; Choi, J.W.; Choi, Y.H.; Nam, T.J. Protective Effect of Pyropia yezoensis Peptide on Dexamethasone-Induced Myotube Atrophy in C2C12 Myotubes. Mar. Drugs 2019, 17, 284. [Google Scholar] [CrossRef] [Green Version]
- de Kloet, E.R.; Joels, M.; Holsboer, F. Stress and the brain: From adaptation to disease. Nat. Rev. Neurosci. 2005, 6, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.K.; Ma, Y.J.; Li, Q.; Liu, E.M.; Jin, M.L.; Zhang, L.P.; Wei, C.H. The role of N-6-methyladenosine RNA methylation in the heat stress response of sheep (Ovis aries). Cell Stress Chaperones 2019, 24, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.Z.; Samanta, D.; Lu, H.Q.; Bullen, J.W.; Zhang, H.M.; Chen, I.; He, X.S.; Semenza, G.L. Hypoxia induces the breast cancer stem cell phenotype by HIF-dependent and ALKBH5-mediated m(6)A-demethylation of NANOG mRNA. Proc. Natl. Acad. Sci. USA 2016, 113, E2047–E2056. [Google Scholar] [CrossRef] [Green Version]
- Kudou, K.; Komatsu, T.; Nogami, J.; Maehara, K.; Harada, A.; Saeki, H.; Oki, E.; Maehara, Y.; Ohkawa, Y. The requirement of Mettl3-promoted MyoD mRNA maintenance in proliferative myoblasts for skeletal muscle differentiation. Open Biol. 2017, 7, 170119. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.X.; Yao, Y.L.; Han, J.H.; Yang, Y.L.; Chen, Y.; Tang, Z.L.; Gao, F. Longitudinal epitranscriptome profiling reveals the crucial role of N-6-methyladenosine methylation in porcine prenatal skeletal muscle development. J. Genet. Genom. 2020, 47, 466–476. [Google Scholar] [CrossRef]
- Feng, Y.; Li, Y.; Jiang, W.; Hu, Y.; Jia, Y.; Zhao, R. GR-mediated transcriptional regulation of m(6)A metabolic genes contributes to diet-induced fatty liver in hens. J. Anim. Sci. Biotechnol. 2021, 12, 117. [Google Scholar] [CrossRef]
- Kostyo, J.L.; Redmond, A.F. Role of protein synthesis in the inhibitory action of adrenal steroid hormones on amino acid transport by muscle. Endocrinology 1966, 79, 531–540. [Google Scholar] [CrossRef]
- Li, Z.Y.; Zhang, H.F. Reprogramming of glucose, fatty acid and amino acid metabolism for cancer progression. Cell. Mol. Life Sci. 2016, 73, 377–392. [Google Scholar] [CrossRef] [PubMed]
- Poncet, N.; Halley, P.A.; Lipina, C.; Gierlinski, M.; Dady, A.; Singer, G.A.; Febrer, M.; Shi, Y.B.; Yamaguchi, T.P.; Taylor, P.M.; et al. Wnt regulates amino acid transporter Slc7a5 and so constrains the integrated stress response in mouse embryos. EMBO Rep. 2020, 21, e48469. [Google Scholar] [CrossRef] [PubMed]
- Habashy, W.S.; Milfort, M.C.; Adomako, K.; Attia, Y.A.; Rekaya, R.; Aggrey, S.E. Effect of heat stress on amino acid digestibility and transporters in meat-type chickens. Poult. Sci. 2017, 96, 2312–2319. [Google Scholar] [CrossRef] [PubMed]
- Drummond, M.J.; Fry, C.S.; Glynn, E.L.; Timmerman, K.L.; Dickinson, J.M.; Walker, D.K.; Gundermann, D.M.; Volpi, E.; Rasmussen, B.B. Skeletal muscle amino acid transporter expression is increased in young and older adults following resistance exercise. J. Appl. Physiol. 2011, 111, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Dorn, L.E.; Lasman, L.; Chen, J.; Xu, X.Y.; Hund, T.J.; Medvedovic, M.; Hanna, J.H.; van Berlo, J.H.; Accornero, F. The N-6-Methyladenosine mRNA Methylase METTL3 Controls Cardiac Homeostasis and Hypertrophy. Circulation 2019, 139, 533–545. [Google Scholar] [CrossRef]
- Ma, L.F.; Chen, T.X.; Zhang, X.; Miao, Y.Y.; Tian, X.T.; Yu, K.K.; Xu, X.; Niu, Y.J.; Guo, S.S.; Zhang, C.C.; et al. The m(6)A reader YTHDC2 inhibits lung adenocarcinoma tumorigenesis by suppressing SLC7A11-dependent antioxidant function. Redox Biol. 2021, 38, 101801. [Google Scholar] [CrossRef]
- Nicklin, P.; Bergman, P.; Zhang, B.; Triantafellow, E.; Wang, H.; Nyfeler, B.; Yang, H.; Hild, M.; Kung, C.; Wilson, C.; et al. Bidirectional Transport of Amino Acids Regulates mTOR and Autophagy. Cell 2009, 136, 521–534. [Google Scholar] [CrossRef] [Green Version]
- Asaoka, Y.; Nagai, Y.; Namae, M.; Furutani-Seiki, M.; Nishina, H. SLC7 family transporters control the establishment of left-right asymmetry during organogenesis in medaka by activating mTOR signaling. Biochem. Biophys. Res. Commun. 2016, 474, 146–153. [Google Scholar] [CrossRef]
- Wan, X.; Wang, D.; Xiong, Q.; Xiang, H.; Li, H.; Wang, H.; Liu, Z.; Niu, H.; Peng, J.; Jiang, S.; et al. Elucidating a molecular mechanism that the deterioration of porcine meat quality responds to increased cortisol based on transcriptome sequencing. Sci. Rep. 2016, 6, 36589. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence of Primer (5′–3′) | Size (bp) |
|---|---|---|
| METTL3 | F:CTGGGGTTACGAACGGGTAG | 243 |
| R:CCAGGGGACAGTCTCTCAATC | ||
| METTL14 | F:TGTGTTTACGCAAGTGGGGT | 179 |
| R:AATGAAGTCCCCGTCTGTGC | ||
| WTAP | F:GGCCAACGGACCAAGTAATG | 246 |
| R:TCATGTGAGTGGCGTGTGA | ||
| FTO | F:CCCCAGAAAATGCCGTACCT | 228 |
| R:ACCAGGGGTCTCTATGTCCC | ||
| ALKBH5 | F:CGTGTCCGTGTCCTTCTTCA | 201 |
| R:AGGATGATGACAGCTCTGCG | ||
| YTHDF1 | F:CAAGTGGAAGGGCAAGTTCG | 182 |
| R:AGTGCTTGTAGGAGGCGATG | ||
| YTHDF2 | F:ATGCCTCGGCCATTGTGTG | 211 |
| R:CGCCGAGAGAAGGGAACAC | ||
| YTHDF3 | F:GCGCTTCGCCTTCAAGTGTA | 220 |
| R:TTCAGGGAACGGTAAGCTGC | ||
| SLC7A7 | F:TGTGCCTATGTCAAGTGGGG | 171 |
| R:CAGAGCGATGTCACCTGTTG | ||
| IGF1 | F:CTCTCCTTCACCAGCTCTGC | 200 |
| R:TCCAGCCTCCTCAGATCACA | ||
| MuRF1 | F:ATGGAGAACCTGGAGAAGCA | 219 |
| R:ACGGTCCATGATCACCTCAT | ||
| β-actin | F:CCAGGTCATCACCATCGG R:CCGTGTTGGCGTAGAGGT | 155 |
| Gene | Sequence of Primer (5′-3′) | Size (bp) |
|---|---|---|
| SLC7A7 | F:CCGGAATTCATGGTTGACGGCATGAAGTA | 1542 |
| R:CTAGCTAGCTTAATTAGACTTGGGATCTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, W.-J.; Guo, K.; Shi, J.-L.; Guo, C.-T.; Xu, J.-L.; Zheng, R.; Jiang, S.-W.; Chai, J. Glucocorticoid Regulates the Synthesis of Porcine Muscle Protein through m6A Modified Amino Acid Transporter SLC7A7. Int. J. Mol. Sci. 2022, 23, 661. https://doi.org/10.3390/ijms23020661
Xu W-J, Guo K, Shi J-L, Guo C-T, Xu J-L, Zheng R, Jiang S-W, Chai J. Glucocorticoid Regulates the Synthesis of Porcine Muscle Protein through m6A Modified Amino Acid Transporter SLC7A7. International Journal of Molecular Sciences. 2022; 23(2):661. https://doi.org/10.3390/ijms23020661
Chicago/Turabian StyleXu, Wei-Jing, Kai Guo, Jia-Li Shi, Chang-Tong Guo, Jia-Le Xu, Rong Zheng, Si-Wen Jiang, and Jin Chai. 2022. "Glucocorticoid Regulates the Synthesis of Porcine Muscle Protein through m6A Modified Amino Acid Transporter SLC7A7" International Journal of Molecular Sciences 23, no. 2: 661. https://doi.org/10.3390/ijms23020661
APA StyleXu, W.-J., Guo, K., Shi, J.-L., Guo, C.-T., Xu, J.-L., Zheng, R., Jiang, S.-W., & Chai, J. (2022). Glucocorticoid Regulates the Synthesis of Porcine Muscle Protein through m6A Modified Amino Acid Transporter SLC7A7. International Journal of Molecular Sciences, 23(2), 661. https://doi.org/10.3390/ijms23020661