
Curcumin and Weight Loss: Does It Work?

 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
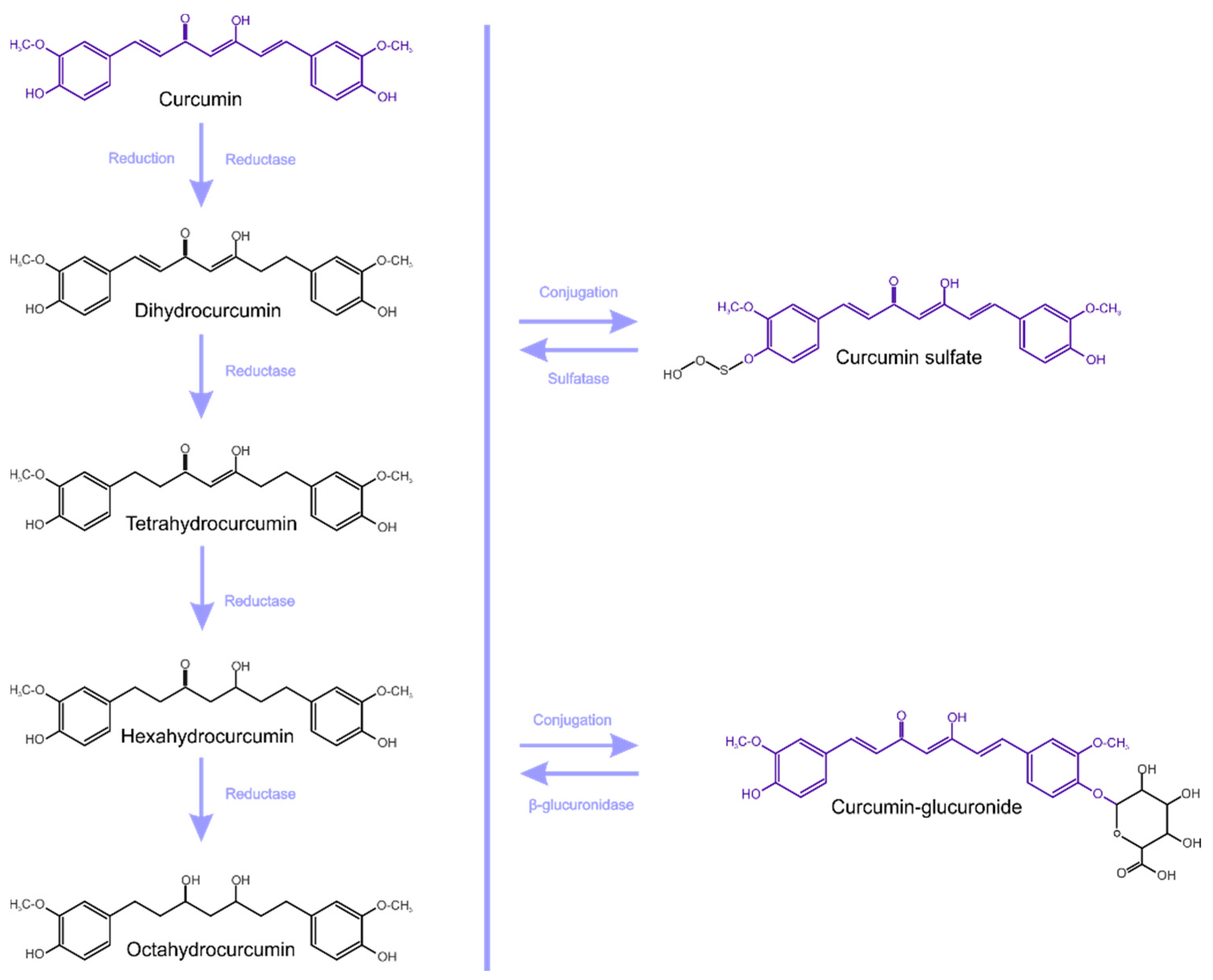
2. Medicinal Activities, Bioavailability and Metabolism of Curcumin—Brief Overview
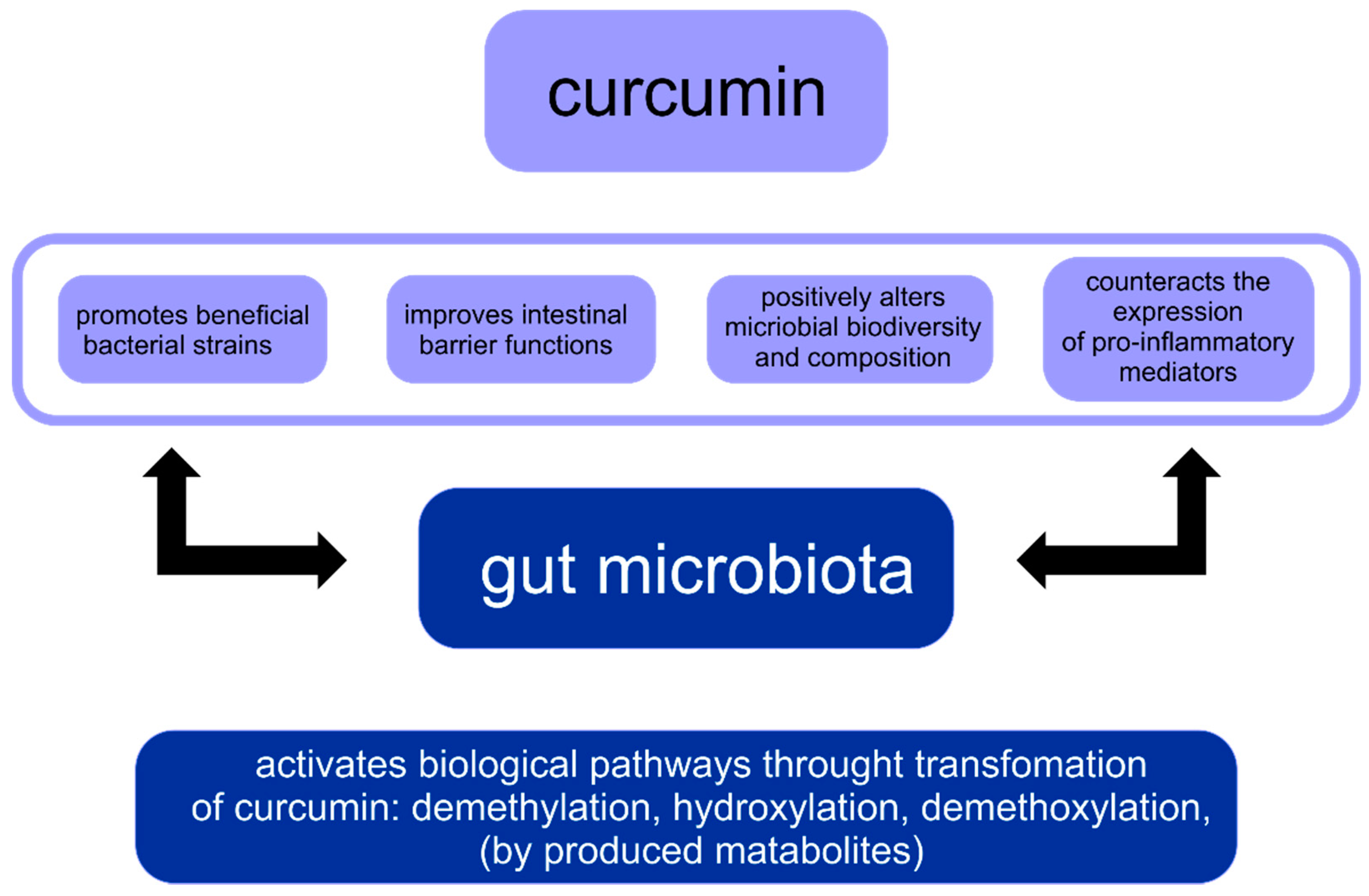
3. Reciprocal Interaction between Curcumin and Gut Microbiota (GM)
Curcumin, Gut Microbiota and Obesity
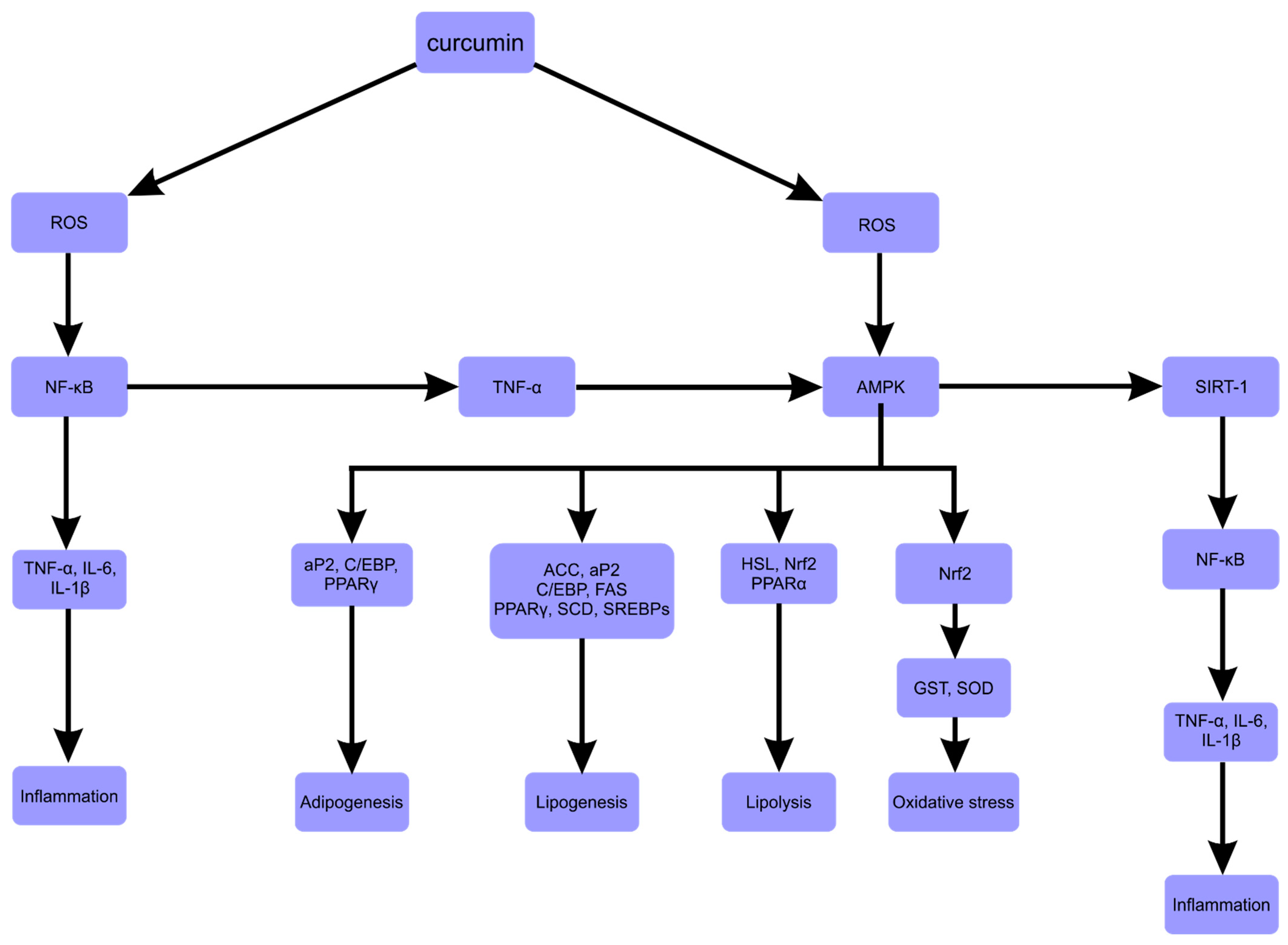
4. Redox Balance in Obesity, ROS and Inflammation in Obesity
5. The Inhibition of Adipogenesis
6. Regulation of Lipid Metabolism
7. Stimulation of Energy Expenditure
8. Biosafety, Toxic Reaction and Curcumin–Drug Interactions
9. Summary and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| ACC | Acetyl-CoA carboxylase |
| Acetyl-CoA | Acetyl-coenzyme A |
| AD | Alzheimer’s disease |
| ADI | Allowable daily intake |
| AMP | 5’ AMP-activated protein kinase |
| aP2 | Adipocyte protein 2 |
| Apo | Apolipoprotein |
| AUC | Area under the concentration time curve |
| BA | Bile acids |
| BAT | Brown adipose tissue |
| BMI | Body mass index |
| BW | Body weight |
| C/EBP | CCAAT/enhancer-binding proteins |
| cAMP | Cyclic adenosine monophosphate |
| CYP | Cytochrome P450 |
| DCA | Deoxycholic acid |
| EFSA | European Food Safety Authority |
| ER | Endoplasmic reticulum |
| FAS | Fatty acid synthase |
| FAs | Fatty acids |
| FFAs | Free Fatty acids |
| FGF21 | Fibroblast growth factor 21 |
| FIAF | Fasting-induced adipose factor |
| GM | Gut Microbiota |
| GPCR | G protein-coupled receptor |
| GPx | Glutathione peroxidase |
| GST | Glutathione S-transferase |
| HDL | High-density lipoprotein |
| HDL-C | High-density lipoprotein cholesterol |
| HFD | High-fat diet |
| IEC | Intestinal epithelial cells |
| IUPAC | International Union of Pure and Applied Chemistry |
| LCA | Lithocholic acid |
| LDL | Low-density lipoprotein |
| LDL-C | Low-density lipoprotein cholesterol |
| LFD | Low-fat diet |
| LPS | Lipopolysaccharide |
| MAPK | Mitogen-activated protein kinase |
| NF-κB | Nuclear factor-κ |
| NQO1 | NAD(P)H:quinone oxidoreductase 1-NQO1 |
| Nrf2 | Nuclear factor erythroid 2-like 2 |
| PKA | Protein kinase A |
| PPAR | Peroxisome proliferation-activated receptor |
| PRx | Peroxiredoxins |
| ROS | Reactive oxygen species |
| SCD | Stearoyl-CoA desaturase |
| SIRT | Sirtuin |
| SOD | Superoxide dismutase |
| SREBP | Sterol regulatory element binding proteins |
| STAT3 | Signal transducer and activator of transcription 3 |
| Tcf7l2 | Transcription factor 7-like 2 |
| TGFβ | Transforming growth factor beta |
| TGR5 | G-protein-coupled bile acid receptor |
| TLR | Toll-like receptor |
| TNF | Tumor necrosis factor |
| UCP | Uncoupling protein |
| WAT | White adipose tissue |
| WD | Western-type diet |
| WHO | World Health Organization |
References
- Singh, M.; Thrimawithana, T.; Shukla, R.; Adhikari, B. Managing Obesity through Natural Polyphenols: A Review. Future Foods 2020, 1–2, 100002. [Google Scholar] [CrossRef]
- Islam, T.; Koboziev, I.; Albracht-Schulte, K.; Mistretta, B.; Scoggin, S.; Yosofvand, M.; Moussa, H.; Zabet-Moghaddam, M.; Ramalingam, L.; Gunaratne, P.H.; et al. Curcumin Reduces Adipose Tissue Inflammation and Alters Gut Microbiota in Diet-Induced Obese Male Mice. Mol. Nutr. Food Res. 2021, 65, 2100274. [Google Scholar] [CrossRef]
- WHO. Diet, Nutrition, and the Prevention of Chronic Diseases: Report of a WHO-FAO Expert Consultation, Proceedings of the JoInt. WHO-FAO Expert Consultation on Diet, Nutrition, and the Prevention of Chronic Diseases, Geneva, Switzerland, 28 January–1 February 2002; WHO Technical Report Series; Weltgesundheitsorganisation, FAO, Eds.; World Health Organization: Geneva, Switzerland, 2003; ISBN 978-92-4-120916-8. [Google Scholar]
- Kasprzak, K.; Wojtunik-Kulesza, K.; Oniszczuk, T.; Kuboń, M.; Oniszczuk, A. Secondary Metabolites, Dietary Fiber and Conjugated Fatty Acids as Functional Food Ingredients against Overweight and Obesity. Nat. Prod. Commun. 2018, 13, 1073–1082. [Google Scholar] [CrossRef]
- Wadden, T.A.; Webb, V.L.; Moran, C.H.; Bailer, B.A. Lifestyle Modification for Obesity. Circulation 2012, 125, 1157–1170. [Google Scholar] [CrossRef]
- Poirier, P.; Giles, T.D.; Bray, G.A.; Hong, Y.; Stern, J.S.; Pi-Sunyer, F.X.; Eckel, R.H.; American Heart Association. Obesity Committee of the Council on Nutrition, Physical Activity, and Metabolism Obesity and Cardiovascular Disease: Pathophysiology, Evaluation, and Effect of Weight Loss: An Update of the 1997 American Heart Association Scientific Statement on Obesity and Heart Disease from the Obesity Committee of the Council on Nutrition, Physical Activity, and Metabolism. Circulation 2006, 113, 898–918. [Google Scholar] [CrossRef] [PubMed]
- Renehan, A.G.; Tyson, M.; Egger, M.; Heller, R.F.; Zwahlen, M. Body-Mass Index and Incidence of Cancer: A Systematic Review and Meta-Analysis of Prospective Observational Studies. Lancet 2008, 371, 569–578. [Google Scholar] [CrossRef]
- Després, J.-P. Abdominal Obesity: The Most Prevalent Cause of the Metabolic Syndrome and Related Cardiometabolic Risk. Eur. Heart J. Suppl. 2006, 8, B4–B12. [Google Scholar] [CrossRef]
- Zhang, C.; Rexrode, K.M.; van Dam, R.M.; Li, T.Y.; Hu, F.B. Abdominal Obesity and the Risk of All-Cause, Cardiovascular, and Cancer Mortality: Sixteen Years of Follow-up in US Women. Circulation 2008, 117, 1658–1667. [Google Scholar] [CrossRef]
- The Challenge of Obesity in the WHO European Region and the Strategies for Response: Summary; Branca, F., Nikogosian, H., Lobstein, T., World Health Organization, Eds.; World Health Organization, Regional Office for Europe: Copenhagen, Denmark, 2007; ISBN 978-92-890-1388-8. [Google Scholar]
- Meydani, M.; Hasan, S.T. Dietary Polyphenols and Obesity. Nutrients 2010, 2, 737–751. [Google Scholar] [CrossRef] [PubMed]
- Alsharif, F.J.; Almuhtadi, Y.A. The Effect of Curcumin Supplementation on Anthropometric Measures among Overweight or Obese Adults. Nutrients 2021, 13, 680. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, F.; Margarucci, S.; Galderisi, U.; Crispi, S.; Peluso, G. Curcumin, Gut Microbiota, and Neuroprotection. Nutrients 2019, 11, 2426. [Google Scholar] [CrossRef] [PubMed]
- Appel, K.; Meiser, P.; Millán, E.; Collado, J.A.; Rose, T.; Gras, C.C.; Carle, R.; Muñoz, E. Chokeberry (Aronia Melanocarpa (Michx.) Elliot) Concentrate Inhibits NF-ΚB and Synergizes with Selenium to Inhibit the Release of pro-Inflammatory Mediators in Macrophages. Fitoterapia 2015, 105, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Zhong, Y.; Duan, Y.; Chen, Q.; Li, F. Antioxidant Mechanism of Tea Polyphenols and Its Impact on Health Benefits. Anim. Nutr. 2020, 6, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Scazzocchio, B.; Minghetti, L.; D’Archivio, M. Interaction between Gut Microbiota and Curcumin: A New Key of Understanding for the Health Effects of Curcumin. Nutrients 2020, 12, 2499. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.-S.; Chen, T.-H.; Weng, L.; Huang, L.; Lai, D.; Weng, C.-F. Pharmacological Properties and Underlying Mechanisms of Curcumin and Prospects in Medicinal Potential. Biomed. Pharm. 2021, 141, 111888. [Google Scholar] [CrossRef]
- Wong, S.C.; Kamarudin, M.N.A.; Naidu, R. Anticancer Mechanism of Curcumin on Human Glioblastoma. Nutrients 2021, 13, 950. [Google Scholar] [CrossRef] [PubMed]
- Augusti, P.R.; Conterato, G.M.M.; Denardin, C.C.; Prazeres, I.D.; Serra, A.T.; Bronze, M.R.; Emanuelli, T. Bioactivity, Bioavailability, and Gut Microbiota Transformations of Dietary Phenolic Compounds: Implications for COVID-19. J. Nutr. Biochem. 2021, 97, 108787. [Google Scholar] [CrossRef] [PubMed]
- Den Hartogh, D.J.; Gabriel, A.; Tsiani, E. Antidiabetic Properties of Curcumin I: Evidence from In Vitro Studies. Nutrients 2020, 12, 118. [Google Scholar] [CrossRef]
- Tossetta, G.; Fantone, S.; Giannubilo, S.R.; Marzioni, D. The Multifaced Actions of Curcumin in Pregnancy Outcome. Antioxidants 2021, 10, 126. [Google Scholar] [CrossRef]
- Ireson, C.R.; Jones, D.J.L.; Orr, S.; Coughtrie, M.W.H.; Boocock, D.J.; Williams, M.L.; Farmer, P.B.; Steward, W.P.; Gescher, A.J. Metabolism of the Cancer Chemopreventive Agent Curcumin in Human and Rat Intestine. Cancer Epidemiol. Biomark. Prev. 2002, 11, 105–111. [Google Scholar]
- Shabbir, U.; Rubab, M.; Daliri, E.B.-M.; Chelliah, R.; Javed, A.; Oh, D.-H. Curcumin, Quercetin, Catechins and Metabolic Diseases: The Role of Gut Microbiota. Nutrients 2021, 13, 206. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.H.; Huang, T.M.; Lin, J.K. Biotransformation of Curcumin through Reduction and Glucuronidation in Mice. Drug Metab. Dispos. 1999, 27, 486–494. [Google Scholar] [PubMed]
- Sharma, R.A.; Euden, S.A.; Platton, S.L.; Cooke, D.N.; Shafayat, A.; Hewitt, H.R.; Marczylo, T.H.; Morgan, B.; Hemingway, D.; Plummer, S.M.; et al. Phase I Clinical Trial of Oral Curcumin: Biomarkers of Systemic Activity and Compliance. Clin. Cancer Res. 2004, 10, 6847–6854. [Google Scholar] [CrossRef]
- Dhillon, N.; Aggarwal, B.B.; Newman, R.A.; Wolff, R.A.; Kunnumakkara, A.B.; Abbruzzese, J.L.; Ng, C.S.; Badmaev, V.; Kurzrock, R. Phase II Trial of Curcumin in Patients with Advanced Pancreatic Cancer. Clin. Cancer Res. 2008, 14, 4491–4499. [Google Scholar] [CrossRef]
- Lao, C.D.; Ruffin, M.T.; Normolle, D.; Heath, D.D.; Murray, S.I.; Bailey, J.M.; Boggs, M.E.; Crowell, J.; Rock, C.L.; Brenner, D.E. Dose Escalation of a Curcuminoid Formulation. BMC Complement. Altern. Med. 2006, 6, 10. [Google Scholar] [CrossRef] [PubMed]
- Jabczyk, M.; Nowak, J.; Hudzik, B.; Zubelewicz-Szkodzińska, B. Curcumin and Its Potential Impact on Microbiota. Nutrients 2021, 13, 2004. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Chaturvedi, M.; Mishra, S.; Kumar, P.; Somvanshi, P.; Chaturvedi, R. Reductive Metabolites of Curcumin and Their Therapeutic Effects. Heliyon 2020, 6, e05469. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, T. Curcumin as a Functional Food-Derived Factor: Degradation Products, Metabolites, Bioactivity, and Future Perspectives. Food Funct. 2018, 9, 705–714. [Google Scholar] [CrossRef]
- Hassaninasab, A.; Hashimoto, Y.; Tomita-Yokotani, K.; Kobayashi, M. Discovery of the Curcumin Metabolic Pathway Involving a Unique Enzyme in an Intestinal Microorganism. Proc. Natl. Acad. Sci. USA 2011, 108, 6615–6620. [Google Scholar] [CrossRef]
- Tan, S.; Rupasinghe, T.W.T.; Tull, D.L.; Boughton, B.; Oliver, C.; McSweeny, C.; Gras, S.L.; Augustin, M.A. Degradation of Curcuminoids by in Vitro Pure Culture Fermentation. J. Agric. Food Chem. 2014, 62, 11005–11015. [Google Scholar] [CrossRef] [PubMed]
- Stachowska, E. Żywienie w Zaburzeniach Mikrobioty Jelitowej; PZWL: Warszawa, Poland, 2021; ISBN 978-83-200-6328-8. [Google Scholar]
- Kasprzak-Drozd, K.; Oniszczuk, T.; Stasiak, M.; Oniszczuk, A. Beneficial Effects of Phenolic Compounds on Gut Microbiota and Metabolic Syndrome. Int. J. Mol. Sci. 2021, 22, 3715. [Google Scholar] [CrossRef]
- Khan, M.J.; Gerasimidis, K.; Edwards, C.A.; Shaikh, M.G. Role of Gut Microbiota in the Aetiology of Obesity: Proposed Mechanisms and Review of the Literature. J. Obes. 2016, 2016, 7353642. [Google Scholar] [CrossRef]
- Geurts, L.; Lazarevic, V.; Derrien, M.; Everard, A.; Van Roye, M.; Knauf, C.; Valet, P.; Girard, M.; Muccioli, G.G.; François, P.; et al. Altered Gut Microbiota and Endocannabinoid System Tone in Obese and Diabetic Leptin-Resistant Mice: Impact on Apelin Regulation in Adipose Tissue. Front. Microbiol. 2011, 2, 149. [Google Scholar] [CrossRef]
- Kimura, I.; Ozawa, K.; Inoue, D.; Imamura, T.; Kimura, K.; Maeda, T.; Terasawa, K.; Kashihara, D.; Hirano, K.; Tani, T.; et al. The Gut Microbiota Suppresses Insulin-Mediated Fat Accumulation via the Short-Chain Fatty Acid Receptor GPR43. Nat. Commun. 2013, 4, 1829. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-B.; Xin, S.-S.; Ding, L.-N.; Ding, W.-Y.; Hou, Y.-L.; Liu, C.-Q.; Zhang, X.-D. The Potential Role of Probiotics in Controlling Overweight/Obesity and Associated Metabolic Parameters in Adults: A Systematic Review and Meta-Analysis. Evid. -Based Complement. Altern. Med. 2019, 2019, e3862971. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.; Song, Z.; Weng, J.; Fantus, I.G. Curcumin and Other Dietary Polyphenols: Potential Mechanisms of Metabolic Actions and Therapy for Diabetes and Obesity. Am. J. Physiol. Endocrinol. Metab. 2018, 314, E201–E205. [Google Scholar] [CrossRef]
- Anhê, F.F.; Varin, T.V.; Le Barz, M.; Desjardins, Y.; Levy, E.; Roy, D.; Marette, A. Gut Microbiota Dysbiosis in Obesity-Linked Metabolic Diseases and Prebiotic Potential of Polyphenol-Rich Extracts. Curr. Obes. Rep. 2015, 4, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Liu, L.; Ji, H.-F. Regulative Effects of Curcumin Spice Administration on Gut Microbiota and Its Pharmacological Implications. Food Nutr. Res. 2017, 61, 1361780. [Google Scholar] [CrossRef]
- Zhong, Y.-B.; Kang, Z.-P.; Wang, M.-X.; Long, J.; Wang, H.-Y.; Huang, J.-Q.; Wei, S.-Y.; Zhou, W.; Zhao, H.-M.; Liu, D.-Y. Curcumin Ameliorated Dextran Sulfate Sodium-Induced Colitis via Regulating the Homeostasis of DCs and Treg and Improving the Composition of the Gut Microbiota. J. Funct. Foods 2021, 86, 104716. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, Y.; Xiang, L.; Wang, Z.; Xiao, G.G.; Hu, J. Effect of Curcumin on the Diversity of Gut Microbiota in Ovariectomized Rats. Nutrients 2017, 9, 1146. [Google Scholar] [CrossRef]
- Hersoug, L.-G.; Møller, P.; Loft, S. Gut Microbiota-Derived Lipopolysaccharide Uptake and Trafficking to Adipose Tissue: Implications for Inflammation and Obesity. Obes. Rev. 2016, 17, 297–312. [Google Scholar] [CrossRef]
- Wang, J.; Ghosh, S.S.; Ghosh, S. Curcumin Improves Intestinal Barrier Function: Modulation of Intracellular Signaling, and Organization of Tight Junctions. Am. J. Physiol. Cell Physiol. 2017, 312, C438–C445. [Google Scholar] [CrossRef]
- Han, Z.; Yao, L.; Zhong, Y.; Xiao, Y.; Gao, J.; Zheng, Z.; Fan, S.; Zhang, Z.; Gong, S.; Chang, S.; et al. Gut Microbiota Mediates the Effects of Curcumin on Enhancing Ucp1-Dependent Thermogenesis and Improving High-Fat Diet-Induced Obesity. Food Funct. 2021, 12, 6558–6575. [Google Scholar] [CrossRef]
- Broeders, E.P.M.; Nascimento, E.B.M.; Havekes, B.; Brans, B.; Roumans, K.H.M.; Tailleux, A.; Schaart, G.; Kouach, M.; Charton, J.; Deprez, B.; et al. The Bile Acid Chenodeoxycholic Acid Increases Human Brown Adipose Tissue Activity. Cell Metab. 2015, 22, 418–426. [Google Scholar] [CrossRef]
- Watanabe, M.; Houten, S.M.; Mataki, C.; Christoffolete, M.A.; Kim, B.W.; Sato, H.; Messaddeq, N.; Harney, J.W.; Ezaki, O.; Kodama, T.; et al. Bile Acids Induce Energy Expenditure by Promoting Intracellular Thyroid Hormone Activation. Nature 2006, 439, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.S.; Bie, J.; Wang, J.; Ghosh, S. Oral Supplementation with Non-Absorbable Antibiotics or Curcumin Attenuates Western Diet-Induced Atherosclerosis and Glucose Intolerance in LDLR−/−Mice—Role of Intestinal Permeability and Macrophage Activation. PLoS ONE 2014, 9, e108577. [Google Scholar] [CrossRef] [PubMed]
- Ellulu, M.S.; Patimah, I.; Khaza’ai, H.; Rahmat, A.; Abed, Y. Obesity and Inflammation: The Linking Mechanism and the Complications. Arch. Med. Sci. 2017, 13, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Neyrinck, A.M.; Alligier, M.; Memvanga, P.B.; Névraumont, E.; Larondelle, Y.; Préat, V.; Cani, P.D.; Delzenne, N.M. Curcuma Longa Extract Associated with White Pepper Lessens High Fat Diet-Induced Inflammation in Subcutaneous Adipose Tissue. PLoS ONE 2013, 8, e81252. [Google Scholar] [CrossRef]
- Carrera-Quintanar, L.; López Roa, R.I.; Quintero-Fabián, S.; Sánchez-Sánchez, M.A.; Vizmanos, B.; Ortuño-Sahagún, D. Phytochemicals That Influence Gut Microbiota as Prophylactics and for the Treatment of Obesity and Inflammatory Diseases. Mediat. Inflamm. 2018, 2018, e9734845. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Torres, I.; Castrejón-Téllez, V.; Soto, M.E.; Rubio-Ruiz, M.E.; Manzano-Pech, L.; Guarner-Lans, V. Oxidative Stress, Plant Natural Antioxidants, and Obesity. Int. J. Mol. Sci. 2021, 22, 1786. [Google Scholar] [CrossRef]
- Halliwell, B. Free Radicals and Antioxidants: Updating a Personal View. Nutr. Rev. 2012, 70, 257–265. [Google Scholar] [CrossRef]
- Pérez-Torres, I.; Guarner-Lans, V.; Rubio-Ruiz, M.E. Reductive Stress in Inflammation-Associated Diseases and the Pro-Oxidant Effect of Antioxidant Agents. Int. J. Mol. Sci. 2017, 18, 2098. [Google Scholar] [CrossRef] [PubMed]
- Meneses, M.J.; Silvestre, R.; Sousa-Lima, I.; Macedo, M.P. Paraoxonase-1 as a Regulator of Glucose and Lipid Homeostasis: Impact on the Onset and Progression of Metabolic Disorders. Int. J. Mol. Sci. 2019, 20, 4049. [Google Scholar] [CrossRef] [PubMed]
- Conner, E.M.; Grisham, M.B. Inflammation, Free Radicals, and Antioxidants. Nutrition 1996, 12, 274–277. [Google Scholar] [CrossRef]
- Coelho, M.; Oliveira, T.; Fernandes, R. State of the Art Paper Biochemistry of Adipose Tissue: An Endocrine Organ. Arch. Med. Sci. 2013, 9, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Makki, K.; Froguel, P.; Wolowczuk, I. Adipose Tissue in Obesity-Related Inflammation and Insulin Resistance: Cells, Cytokines, and Chemokines. ISRN Inflamm. 2013, 2013, 139239. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.M.; Macedo-de la Concha, L.E.; Pantoja-Meléndez, C.A. Low-Grade Inflammation and Its Relation to Obesity and Chronic Degenerative Diseases. Rev. Médica Del. Hosp. Gen. De México 2017, 80, 101–105. [Google Scholar] [CrossRef]
- Boaru, S.G.; Borkham-Kamphorst, E.; Van de Leur, E.; Lehnen, E.; Liedtke, C.; Weiskirchen, R. NLRP3 Inflammasome Expression Is Driven by NF-ΚB in Cultured Hepatocytes. Biochem. Biophys. Res. Commun. 2015, 458, 700–706. [Google Scholar] [CrossRef]
- Nisr, R.B.; Shah, D.S.; Ganley, I.G.; Hundal, H.S. Proinflammatory NFkB Signalling Promotes Mitochondrial Dysfunction in Skeletal Muscle in Response to Cellular Fuel Overloading. Cell Mol. Life Sci. 2019, 76, 4887–4904. [Google Scholar] [CrossRef] [PubMed]
- Oliveira-Marques, V.; Marinho, H.S.; Cyrne, L.; Antunes, F. Role of Hydrogen Peroxide in NF-KappaB Activation: From Inducer to Modulator. Antioxid. Redox Signal. 2009, 11, 2223–2243. [Google Scholar] [CrossRef]
- Morgan, M.J.; Liu, Z. Crosstalk of Reactive Oxygen Species and NF-ΚB Signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Nam, M.; Kang, M.; Lee, J.; Hwang, G.-S.; Kim, H. Piperine Regulates UCP1 through the AMPK Pathway by Generating Intracellular Lactate Production in Muscle Cells. Sci. Rep. 2017, 7, 41066. [Google Scholar] [CrossRef]
- Böni-Schnetzler, M.; Boller, S.; Debray, S.; Bouzakri, K.; Meier, D.T.; Prazak, R.; Kerr-Conte, J.; Pattou, F.; Ehses, J.A.; Schuit, F.C.; et al. Free Fatty Acids Induce a Proinflammatory Response in Islets via the Abundantly Expressed Interleukin-1 Receptor I. Endocrinology 2009, 150, 5218–5229. [Google Scholar] [CrossRef]
- Akhter, N.; Madhoun, A.; Arefanian, H.; Wilson, A.; Kochumon, S.; Thomas, R.; Shenouda, S.; Al-Mulla, F.; Ahmad, R.; Sindhu, S. Oxidative Stress Induces Expression of the Toll-like Receptors (TLRs) 2 and 4 in the Human Peripheral Blood Mononuclear Cells: Implications for Metabolic Inflammation. Cell. Physiol. Biochem. 2019, 53, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Kim, F.; Pham, M.; Luttrell, I.; Bannerman, D.D.; Tupper, J.; Thaler, J.; Hawn, T.R.; Raines, E.W.; Schwartz, M.W. Toll-like Receptor-4 Mediates Vascular Inflammation and Insulin Resistance in Diet-Induced Obesity. Circ. Res. 2007, 100, 1589–1596. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Park, A.; Oh, K.-J.; Lee, S.C.; Kim, W.K.; Bae, K.-H. The Role of Adipose Tissue Mitochondria: Regulation of Mitochondrial Function for the Treatment of Metabolic Diseases. Int. J. Mol. Sci. 2019, 20, 4924. [Google Scholar] [CrossRef]
- Ali, A.T.; Hochfeld, W.E.; Myburgh, R.; Pepper, M.S. Adipocyte and Adipogenesis. Eur J. Cell Biol. 2013, 92, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Lefterova, M.I.; Lazar, M.A. New Developments in Adipogenesis. Trends Endocrinol. Metab. 2009, 20, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Brey, C.W.; Nelder, M.P.; Hailemariam, T.; Gaugler, R.; Hashmi, S. Krüppel-like Family of Transcription Factors: An Emerging New Frontier in Fat Biology. Int. J. Biol. Sci. 2009, 5, 622–636. [Google Scholar] [CrossRef] [PubMed]
- Schmid, B.; Rippmann, J.F.; Tadayyon, M.; Hamilton, B.S. Inhibition of Fatty Acid Synthase Prevents Preadipocyte Differentiation. Biochem. Biophys. Res. Commun. 2005, 328, 1073–1082. [Google Scholar] [CrossRef]
- Mohamed, G.A.; Ibrahim, S.R.M.; Elkhayat, E.S.; Dine, R.S.E. Natural Anti-Obesity Agents. Bull. Fac. Pharm. Cairo Univ. 2014, 2, 269–284. [Google Scholar] [CrossRef]
- Jin, T. Mechanisms Underlying the Metabolic Beneficial Effect of Curcumin Intervention: Beyond Anti-Inflammation and Anti-Oxidative Stress. Obes. Med. 2019, 13, 1–5. [Google Scholar] [CrossRef]
- Jin, T. Current Understanding on Role of the Wnt Signaling Pathway Effector TCF7L2 in Glucose Homeostasis. Endocr Rev. 2016, 37, 254–277. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.Y.; Le, T.T.; Chen, C.; Cheng, J.-X.; Kim, K.-H. Curcumin Inhibits Adipocyte Differentiation through Modulation of Mitotic Clonal Expansion. J. Nutr. Biochem. 2011, 22, 910–920. [Google Scholar] [CrossRef]
- Ahn, J.; Lee, H.; Kim, S.; Ha, T. Curcumin-Induced Suppression of Adipogenic Differentiation Is Accompanied by Activation of Wnt/Beta-Catenin Signaling. Am. J. Physiol. Cell Physiol. 2010, 298, C1510–C1516. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Li, Y.C.; Wang, J.; Kong, J.; Qi, Y.; Quigg, R.J.; Li, X. MiR-17-92 Cluster Accelerates Adipocyte Differentiation by Negatively Regulating Tumor-Suppressor Rb2/P130. Proc. Natl. Acad. Sci. USA 2008, 105, 2889–2894. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Song, Z.; Shao, W.; Du, W.W.; Zhao, L.R.; Zeng, K.; Yang, B.B.; Jin, T. Curcumin Represses Mouse 3T3-L1 Cell Adipogenic Differentiation via Inhibiting MiR-17-5p and Stimulating the Wnt Signalling Pathway Effector Tcf7l2. Cell Death Dis. 2017, 8, e2559. [Google Scholar] [CrossRef]
- Wang, T.; Xue, C.; Zhang, T.; Wang, Y. The Improvements of Functional Ingredients from Marine Foods in Lipid Metabolism. Trends Food Sci. Technol. 2018, 81, 74–89. [Google Scholar] [CrossRef]
- Karagianni, P.; Talianidis, I. Transcription Factor Networks Regulating Hepatic Fatty Acid Metabolism. Biochim. Biophys. Acta 2015, 1851, 2–8. [Google Scholar] [CrossRef]
- Salvadó, L.; Palomer, X.; Barroso, E.; Vázquez-Carrera, M. Targeting Endoplasmic Reticulum Stress in Insulin Resistance. Trends Endocrinol. Metab. 2015, 26, 438–448. [Google Scholar] [CrossRef]
- Srivastava, G.; Apovian, C.M. Current Pharmacotherapy for Obesity. Nat. Rev. Endocrinol. 2018, 14, 12–24. [Google Scholar] [CrossRef]
- Takei, K.; Nakagawa, Y.; Wang, Y.; Han, S.-I.; Satoh, A.; Sekiya, M.; Matsuzaka, T.; Shimano, H. Effects of K-877, a Novel Selective PPARα Modulator, on Small Intestine Contribute to the Amelioration of Hyperlipidemia in Low-Density Lipoprotein Receptor Knockout Mice. J. Pharm. Sci. 2017, 133, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, S.; Asgary, S.; Askari, G.; Keshvari, M.; Hatamipour, M.; Feizi, A.; Sahebkar, A. Treatment of Non-Alcoholic Fatty Liver Disease with Curcumin: A Randomized Placebo-Controlled Trial. Phytother. Res. 2016, 30, 1540–1548. [Google Scholar] [CrossRef]
- Faghihzadeh, F.; Adibi, P.; Hekmatdoost, A. The Effects of Resveratrol Supplementation on Cardiovascular Risk Factors in Patients with Non-Alcoholic Fatty Liver Disease: A Randomised, Double-Blind, Placebo-Controlled Study. Br. J. Nutr. 2015, 114, 796–803. [Google Scholar] [CrossRef]
- Tian, L.; Zeng, K.; Shao, W.; Yang, B.B.; Fantus, I.G.; Weng, J.; Jin, T. Short-Term Curcumin Gavage Sensitizes Insulin Signaling in Dexamethasone-Treated C57BL/6 Mice. J. Nutr. 2015, 145, 2300–2307. [Google Scholar] [CrossRef] [PubMed]
- Zeng, K.; Tian, L.; Patel, R.; Shao, W.; Song, Z.; Liu, L.; Manuel, J.; Ma, X.; McGilvray, I.; Cummins, C.L.; et al. Diet Polyphenol Curcumin Stimulates Hepatic Fgf21 Production and Restores Its Sensitivity in High-Fat-Diet-Fed Male Mice. Endocrinology 2017, 158, 277–292. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kharitonenkov, A.; DiMarchi, R. FGF21 Revolutions: Recent Advances Illuminating FGF21 Biology and Medicinal Properties. Trends Endocrinol. Metab. 2015, 26, 608–617. [Google Scholar] [CrossRef]
- Vernia, S.; Cavanagh-Kyros, J.; Garcia-Haro, L.; Sabio, G.; Barrett, T.; Jung, D.Y.; Kim, J.K.; Xu, J.; Shulha, H.P.; Garber, M.; et al. The PPARα-FGF21 Hormone Axis Contributes to Metabolic Regulation by the Hepatic JNK Signaling Pathway. Cell Metab. 2014, 20, 512–525. [Google Scholar] [CrossRef]
- Lundåsen, T.; Hunt, M.C.; Nilsson, L.-M.; Sanyal, S.; Angelin, B.; Alexson, S.E.H.; Rudling, M. PPARalpha Is a Key Regulator of Hepatic FGF21. Biochem. Biophys Res. Commun. 2007, 360, 437–440. [Google Scholar] [CrossRef] [PubMed]
- Shao, M.; Yu, L.; Zhang, F.; Lu, X.; Li, X.; Cheng, P.; Lin, X.; He, L.; Jin, S.; Tan, Y.; et al. Additive Protection by LDR and FGF21 Treatment against Diabetic Nephropathy in Type 2 Diabetes Model. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E45–E54. [Google Scholar] [CrossRef] [PubMed]
- Woo, Y.C.; Xu, A.; Wang, Y.; Lam, K.S.L. Fibroblast Growth Factor 21 as an Emerging Metabolic Regulator: Clinical Perspectives. Clin. Endocrinol. 2013, 78, 489–496. [Google Scholar] [CrossRef]
- Bozic, M.; Guzmán, C.; Benet, M.; Sánchez-Campos, S.; García-Monzón, C.; Gari, E.; Gatius, S.; Valdivielso, J.M.; Jover, R. Hepatocyte Vitamin D Receptor Regulates Lipid Metabolism and Mediates Experimental Diet-Induced Steatosis. J. Hepatol. 2016, 65, 748–757. [Google Scholar] [CrossRef] [PubMed]
- Gariani, K.; Drifte, G.; Dunn-Siegrist, I.; Pugin, J.; Jornayvaz, F.R. Increased FGF21 Plasma Levels in Humans with Sepsis and SIRS. Endocr. Connect. 2013, 2, 146–153. [Google Scholar] [CrossRef]
- Berti, L.; Irmler, M.; Zdichavsky, M.; Meile, T.; Böhm, A.; Stefan, N.; Fritsche, A.; Beckers, J.; Königsrainer, A.; Häring, H.-U.; et al. Fibroblast Growth Factor 21 Is Elevated in Metabolically Unhealthy Obesity and Affects Lipid Deposition, Adipogenesis, and Adipokine Secretion of Human Abdominal Subcutaneous Adipocytes. Mol. Metab. 2015, 4, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Rusli, F.; Deelen, J.; Andriyani, E.; Boekschoten, M.V.; Lute, C.; van den Akker, E.B.; Müller, M.; Beekman, M.; Steegenga, W.T. Fibroblast Growth Factor 21 Reflects Liver Fat Accumulation and Dysregulation of Signalling Pathways in the Liver of C57BL/6J Mice. Sci. Rep. 2016, 6, 30484. [Google Scholar] [CrossRef] [PubMed]
- Baziar, N.; Parohan, M. The Effects of Curcumin Supplementation on Body Mass Index, Body Weight, and Waist Circumference in Patients with Nonalcoholic Fatty Liver Disease: A Systematic Review and Dose-Response Meta-Analysis of Randomized Controlled Trials. Phytother. Res. 2020, 34, 464–474. [Google Scholar] [CrossRef] [PubMed]
- Jalali, M.; Mahmoodi, M.; Mosallanezhad, Z.; Jalali, R.; Imanieh, M.H.; Moosavian, S.P. The Effects of Curcumin Supplementation on Liver Function, Metabolic Profile and Body Composition in Patients with Non-Alcoholic Fatty Liver Disease: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Complement. Med. 2020, 48, 102283. [Google Scholar] [CrossRef] [PubMed]
- Den Hartogh, D.J.; Gabriel, A.; Tsiani, E. Antidiabetic Properties of Curcumin II: Evidence from In Vivo Studies. Nutrients 2019, 12, 58. [Google Scholar] [CrossRef] [PubMed]
- Pivari, F.; Mingione, A.; Brasacchio, C.; Soldati, L. Curcumin and Type 2 Diabetes Mellitus: Prevention and Treatment. Nutrients 2019, 11, 1837. [Google Scholar] [CrossRef]
- Perugini, J.; Di Mercurio, E.; Tossetta, G.; Severi, I.; Monaco, F.; Reguzzoni, M.; Tomasetti, M.; Dani, C.; Cinti, S.; Giordano, A. Biological Effects of Ciliary Neurotrophic Factor on HMADS Adipocytes. Front. Endocrinol. 2019, 10, 768. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Lv, J.; Han, M.; Yang, Z.; Li, T.; Jiang, S.; Yang, Y. STAT3: The Art of Multi-Tasking of Metabolic and Immune Functions in Obesity. Prog. Lipid Res. 2018, 70, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Di Pierro, F.; Bressan, A.; Ranaldi, D.; Rapacioli, G.; Giacomelli, L.; Bertuccioli, A. Potential Role of Bioavailable Curcumin in Weight Loss and Omental Adipose Tissue Decrease: Preliminary Data of a Randomized, Controlled Trial in Overweight People with Metabolic Syndrome. Preliminary Study. Eur. Rev. Med. Pharm. Sci. 2015, 19, 4195–4202. [Google Scholar]
- Ganjali, S.; Sahebkar, A.; Mahdipour, E.; Jamialahmadi, K.; Torabi, S.; Akhlaghi, S.; Ferns, G.; Parizadeh, S.M.R.; Ghayour-Mobarhan, M. Investigation of the Effects of Curcumin on Serum Cytokines in Obese Individuals: A Randomized Controlled Trial. Sci. World J. 2014, 2014, 898361. [Google Scholar] [CrossRef]
- Sahebkar, A.; Mohammadi, A.; Atabati, A.; Rahiman, S.; Tavallaie, S.; Iranshahi, M.; Akhlaghi, S.; Ferns, G.A.; Ghayour-Mobarhan, M. Curcuminoids Modulate Pro-Oxidant-Antioxidant Balance but Not the Immune Response to Heat Shock Protein 27 and Oxidized LDL in Obese Individuals. Phytother. Res. 2013, 27, 1883–1888. [Google Scholar] [CrossRef] [PubMed]
- Nieman, D.C.; Cialdella-Kam, L.; Knab, A.M.; Shanely, R.A. Influence of Red Pepper Spice and Turmeric on Inflammation and Oxidative Stress Biomarkers in Overweight Females: A Metabolomics Approach. Plant. Foods Hum. Nutr. 2012, 67, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Baum, L.; Cheung, S.K.K.; Mok, V.C.T.; Lam, L.C.W.; Leung, V.P.Y.; Hui, E.; Ng, C.C.Y.; Chow, M.; Ho, P.C.; Lam, S.; et al. Curcumin Effects on Blood Lipid Profile in a 6-Month Human Study. Pharm. Res. 2007, 56, 509–514. [Google Scholar] [CrossRef]
- Ramírez-Boscá, A.; Soler, A.; Carrión, M.A.; Díaz-Alperi, J.; Bernd, A.; Quintanilla, C.; Quintanilla Almagro, E.; Miquel, J. An Hydroalcoholic Extract of Curcuma Longa Lowers the Apo B/Apo A Ratio. Implications for Atherogenesis Prevention. Mech. Ageing Dev. 2000, 119, 41–47. [Google Scholar] [CrossRef]
- Öner-İyidoğan, Y.; Koçak, H.; Seyidhanoğlu, M.; Gürdöl, F.; Gülçubuk, A.; Yildirim, F.; Çevik, A.; Uysal, M. Curcumin Prevents Liver Fat Accumulation and Serum Fetuin-A Increase in Rats Fed a High-Fat Diet. J. Physiol. Biochem. 2013, 69, 677–686. [Google Scholar] [CrossRef]
- He, H.-J.; Wang, G.-Y.; Gao, Y.; Ling, W.-H.; Yu, Z.-W.; Jin, T.-R. Curcumin Attenuates Nrf2 Signaling Defect, Oxidative Stress in Muscle and Glucose Intolerance in High Fat Diet-Fed Mice. World J. Diabetes 2012, 3, 94–104. [Google Scholar] [CrossRef]
- Shao, W.; Yu, Z.; Chiang, Y.; Yang, Y.; Chai, T.; Foltz, W.; Lu, H.; Fantus, I.G.; Jin, T. Curcumin Prevents High Fat Diet Induced Insulin Resistance and Obesity via Attenuating Lipogenesis in Liver and Inflammatory Pathway in Adipocytes. PLoS ONE 2012, 7, e28784. [Google Scholar] [CrossRef]
- Leray, V.; Freuchet, B.; Le Bloc’h, J.; Jeusette, I.; Torre, C.; Nguyen, P. Effect of Citrus Polyphenol- and Curcumin-Supplemented Diet on Inflammatory State in Obese Cats. Br. J. Nutr. 2011, 106 (Suppl. S1), S198–S201. [Google Scholar] [CrossRef]
- Yekollu, S.K.; Thomas, R.; O’Sullivan, B. Targeting Curcusomes to Inflammatory Dendritic Cells Inhibits NF-ΚB and Improves Insulin Resistance in Obese Mice. Diabetes 2011, 60, 2928–2938. [Google Scholar] [CrossRef]
- Ejaz, A.; Wu, D.; Kwan, P.; Meydani, M. Curcumin Inhibits Adipogenesis in 3T3-L1 Adipocytes and Angiogenesis and Obesity in C57/BL Mice. J. Nutr. 2009, 139, 919–925. [Google Scholar] [CrossRef]
- Pongchaidecha, A.; Lailerd, N.; Boonprasert, W.; Chattipakorn, N. Effects of Curcuminoid Supplement on Cardiac Autonomic Status in High-Fat-Induced Obese Rats. Nutrition 2009, 25, 870–878. [Google Scholar] [CrossRef]
- Jang, E.-M.; Choi, M.-S.; Jung, U.J.; Kim, M.-J.; Kim, H.-J.; Jeon, S.-M.; Shin, S.-K.; Seong, C.-N.; Lee, M.-K. Beneficial Effects of Curcumin on Hyperlipidemia and Insulin Resistance in High-Fat-Fed Hamsters. Metabolism 2008, 57, 1576–1583. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, S.P.; Leibel, R.; Tortoriello, D.V. Dietary Curcumin Significantly Improves Obesity-Associated Inflammation and Diabetes in Mouse Models of Diabesity. Endocrinology 2008, 149, 3549–3558. [Google Scholar] [CrossRef]
- Yu, Y.; Hu, S.; Yan, H. The study of insulin resistance and leptin resistance on the model of simplicity obesity rats by curcumin. Chin. J. Prev. Med. 2008, 42, 818–822. [Google Scholar]
- Xie, X.; Kong, P.-R.; Wu, J.; Li, Y.; Li, Y. Curcumin Attenuates Lipolysis Stimulated by Tumor Necrosis Factor-α or Isoproterenol in 3T3-L1 Adipocytes. Phytomedicine 2012, 20, 3–8. [Google Scholar] [CrossRef]
- Zhao, J.; Sun, X.-B.; Ye, F.; Tian, W.-X. Suppression of Fatty Acid Synthase, Differentiation and Lipid Accumulation in Adipocytes by Curcumin. Mol. Cell Biochem. 2011, 351, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Zhao, S.; Wu, Z.; Yang, J.; Xie, X.; Yu, B.; Nie, S. Curcumin Promotes Cholesterol Efflux from Adipocytes Related to PPARgamma-LXRalpha-ABCA1 Passway. Mol. Cell Biochem. 2011, 358, 281–285. [Google Scholar] [CrossRef]
- Lee, Y.K.; Lee, W.S.; Hwang, J.T.; Kwon, D.Y.; Surh, Y.J.; Park, O.J. Curcumin Exerts Antidifferentiation Effect through AMPKalpha-PPAR-Gamma in 3T3-L1 Adipocytes and Antiproliferatory Effect through AMPKalpha-COX-2 in Cancer Cells. J. Agric. Food Chem. 2009, 57, 305–310. [Google Scholar] [CrossRef]
- Ohishi, T.; Fukutomi, R.; Shoji, Y.; Goto, S.; Isemura, M. The Beneficial Effects of Principal Polyphenols from Green Tea, Coffee, Wine, and Curry on Obesity. Molecules 2021, 26, 453. [Google Scholar] [CrossRef]
- Balaji, M.; Ganjayi, M.S.; Hanuma Kumar, G.E.N.; Parim, B.N.; Mopuri, R.; Dasari, S. A Review on Possible Therapeutic Targets to Contain Obesity: The Role of Phytochemicals. Obes. Res. Clin. Pract. 2016, 10, 363–380. [Google Scholar] [CrossRef] [PubMed]
- Wood dos Santos, T.; Cristina Pereira, Q.; Teixeira, L.; Gambero, A.; A Villena, J.; Lima Ribeiro, M. Effects of Polyphenols on Thermogenesis and Mitochondrial Biogenesis. Int. J. Mol. Sci. 2018, 19, 2757. [Google Scholar] [CrossRef] [PubMed]
- Lowell, B.B.; Spiegelman, B.M. Towards a Molecular Understanding of Adaptive Thermogenesis. Nature 2000, 404, 652–660. [Google Scholar] [CrossRef]
- Mele, L.; Bidault, G.; Mena, P.; Crozier, A.; Brighenti, F.; Vidal-Puig, A.; Del Rio, D. Dietary (Poly)Phenols, Brown Adipose Tissue Activation, and Energy Expenditure: A Narrative Review. Adv. Nutr. 2017, 8, 694–704. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, X.; Ye, Z.; Xu, C.; Zhang, M.; Ruan, B.; Wei, M.; Jiang, Y.; Zhang, Y.; Wang, L.; et al. Curcumin Promotes Browning of White Adipose Tissue in a Norepinephrine-Dependent Way. Biochem. Biophys. Res. Commun. 2015, 466, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Revelo, X.; Shao, W.; Tian, L.; Zeng, K.; Lei, H.; Sun, H.-S.; Woo, M.; Winer, D.; Jin, T. Dietary Curcumin Intervention Targets Mouse White Adipose Tissue Inflammation and Brown Adipose Tissue UCP1 Expression. Obesity 2018, 26, 547–558. [Google Scholar] [CrossRef] [PubMed]
- Xing, T.; Kang, Y.; Xu, X.; Wang, B.; Du, M.; Zhu, M.-J. Raspberry Supplementation Improves Insulin Signaling and Promotes Brown-Like Adipocyte Development in White Adipose Tissue of Obese Mice. Mol. Nutr. Food Res. 2018, 62, 1701035. [Google Scholar] [CrossRef]
- Arias, N.; Picó, C.; Teresa Macarulla, M.; Oliver, P.; Miranda, J.; Palou, A.; Portillo, M.P. A Combination of Resveratrol and Quercetin Induces Browning in White Adipose Tissue of Rats Fed an Obesogenic Diet. Obesity 2017, 25, 111–121. [Google Scholar] [CrossRef]
- Renaud, J.; Martinoli, M.-G. Considerations for the Use of Polyphenols as Therapies in Neurodegenerative Diseases. Int. J. Mol. Sci. 2019, 20, 1883. [Google Scholar] [CrossRef]
- Joint FAO/WHO Expert Committee on Food Additives; Meeting, Joint FAO/WHO Expert Committee on Food Additives; World Health Organization. Evaluation of Certain Food Additives and Contaminants: Sixty-First Report of the Joint FAO/WHO Expert Committee on Food Additives; World Health Organization: Geneva, Switzerland, 2004; ISBN 978-92-4-120922-9. [Google Scholar]
- Authority, E.F.S. Refined Exposure Assessment for Curcumin (E 100). EFSA J. 2014, 12, 3876. [Google Scholar] [CrossRef]
- Soleimani, V.; Sahebkar, A.; Hosseinzadeh, H. Turmeric (Curcuma Longa) and Its Major Constituent (Curcumin) as Nontoxic and Safe Substances: Review. Phytother. Res. 2018, 32, 985–995. [Google Scholar] [CrossRef] [PubMed]
- Fadus, M.C.; Lau, C.; Bikhchandani, J.; Lynch, H.T. Curcumin: An Age-Old Anti-Inflammatory and Anti-Neoplastic Agent. J. Tradit Complement. Med. 2017, 7, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Ringman, J.M.; Frautschy, S.A.; Teng, E.; Begum, A.N.; Bardens, J.; Beigi, M.; Gylys, K.H.; Badmaev, V.; Heath, D.D.; Apostolova, L.G.; et al. Oral Curcumin for Alzheimer’s Disease: Tolerability and Efficacy in a 24-Week Randomized, Double Blind, Placebo-Controlled Study. Alzheimer’s Res. Ther. 2012, 4, 43. [Google Scholar] [CrossRef] [PubMed]
- Kocher, A.; Schiborr, C.; Frank, J. Curcuminoid Drug Interactions. Ernahr. Umsch. 2015, 62, 188–195. [Google Scholar] [CrossRef]
- Bahramsoltani, R.; Rahimi, R.; Farzaei, M.H. Pharmacokinetic Interactions of Curcuminoids with Conventional Drugs: A Review. J. Ethnopharmacol. 2017, 209, 1–12. [Google Scholar] [CrossRef]
- Patel, S.S.; Acharya, A.; Ray, R.S.; Agrawal, R.; Raghuwanshi, R.; Jain, P. Cellular and Molecular Mechanisms of Curcumin in Prevention and Treatment of Disease. Crit. Rev. Food Sci. Nutr. 2020, 60, 887–939. [Google Scholar] [CrossRef]
- Shaikh, A.S.; Thomas, A.B.; Chitlange, S.S. Herb-Drug Interaction Studies of Herbs Used in Treatment of Cardiovascular Disorders-A Narrative Review of Preclinical and Clinical Studies. Phytother. Res. 2020, 34, 1008–1026. [Google Scholar] [CrossRef]
- Ramachandran, C.; Nair, S.M.; Escalon, E.; Melnick, S.J. Potentiation of Etoposide and Temozolomide Cytotoxicity by Curcumin and Turmeric ForceTM in Brain Tumor Cell Lines. J. Complement. Integr. Med. 2012, 9, 20. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Type of Study | Experimental Design and Treatments | Results | Trial Lenght | Ref. |
|---|---|---|---|---|
| Human study | Randomized, controlled trial; 44 obese subjects supplemented with curcumin complexed with phosphatidylserine in phytosome form | ↑BW loss ↑Enhance body fat reduction ↑Waistline reduction ↑Hip circumference reduction ↓BMI | 30 days | [105] |
| Human study | Randomized, double-blind, crossover trial; 30 obese individuals supplemented with capsules containing 500 mg curcuminoids plus 5mg bioperine; two capsules per day | ↓Serum IL1β and IL-4 levels | 4 weeks | [106] |
| Human study | Randomized, double-blind, placebo-controlled crossover trial; 30 obese individuals supplemented with capsules containing 500 mg curcuminoids plus 5 mg bioperine; one capsule per day | ↓Serum triglycerides level | 30 days | [107] |
| Human study | Randomized, doubled-blinded, placebo-controlled, crossover design, 62 overweight/obese females with systemic inflammation supplemented with turmeric; 800 mg per day | No significant changes in none of the investigated metabolic parameters or inflammation | 10 weeks | [108] |
| Human study | Randomized, double-blinded, placebo-controlled trial. Elderly (n = 36, N 50 years); placebo group, 1 g per day curcumin group and 4 g per day curcumin group | ↔Serum lipid profile (triglycerides, total, LDL-C, HDL-C) | 6 months | [109] |
| Human study | A pre–post study. Healthy adults (n = 8, 43–70 years) received 10 mg per day | ↓Serum LDL, Apo B, Apo B/Apo A ↑Serum HDL and Apo A | 30 days | [110] |
| Animal study | Male Sprague–Dawley rats in an HFD-induced obesity model; control group, curcumin 100 mg/kg/BW per daygroup, 400 mg/kg/BW per day, HFD group, HFD + curcumin 100 or HFD + curcumin 400 | ↓Liver triglYcerydes ↓Serum fetuin-A | 8 weeks | [111] |
| Animal study | Male C57BL/6 J mice (8 weeks old) in an HFD-induced obesity and insulin resistance model; LFD group, HFD group and HFD + curcumin group—50 mg/kg BW by gavage | ↔BW ↑NrF2 in skeletal muscle ↑Glucose disposal and insulin sensitivity ↓MDA and ROS in skeletal muscle and mitochondria | 15 days | [112] |
| Animal study | Male C57BL/6 J mice (5 weeks old) in an HFD-induced obesity model; LFD group, HFD group or HFD + curcumin (4 g/kg diet, added 2 days/week) group | ↓BW and fat ↓NF-κB ↓SREBP-1c ↔Wnt signaling in mature adipocytes ↓Inflammatory in adipocytes | 28 weeks | [113] |
| Animal study | European obese cats (6.5 years old); control group, citrus group or curcumin group | ↓IFN-γ and IL-2 mRNA levels ↔mRNA expression of TNF-α, IL-1β, IL-4, IL-5, IL-10, IL-12, IL-18, TGF-β | 8 weeks | [114] |
| Animal study | Male C57BL/6 J, ob/ob mice and nonobese littermates in a model of steatosis; ob/ob control group, lipo group, ob/ob. + curcumin group, nonobese control group or nonobese + curcumin group | ↓NF-κB pathway ↓TNF ↓IL-6 ↑ IL-4 ↑Insulin sensitivity ↑Serum adiponectin | 24 or 72 h | [115] |
| Animal study | Male C57BL/6 mice (4 weeks old) in an HFD-induced obesity model; control group, HFD group or HFD + curcumin group (500 mg/kg of diet) | ↓BW, fat, microvessel density in adipose tissue. ↓Liver weights and hepatic steatosis ↓Serum glucose and triglycerides ↑Fatty acid and energy metabolism (↑ P-AMPK, P-ACC mRNA expression; ↓PPARγ and C/EBPα mRNA expression) | 12 weeks | [116] |
| Animal study | Male Wistar rats (100–120 g) in an HFD-induced obesity model; control group, HFD control group, HFD + 30 mg/kg BW curcuminoid group, HFD + 60 mg/kg BW curcuminoid group, HFD + 90 mg/kg BW curcuminoid group | ↓Plasma FFA ↓glucose levels | 12 weeks | [117] |
| Animal study | Male Golden-Syrian hamsters (4 weeks old) in an HFD-induced obesity model; HFD group or HFD + curcumin (0.05% in diet) | ↔BW, food intake, fat pad mass, plasma glucose Plasma FFA, triglycerydes, leptin, insulin ↑Plasma HDL-C, Apo A-I ↓Hepatic cholesterol and triglycerydes ↑FA β-oxidation activity ↓FAS, HMG-CoA reductase ↓Lipid peroxide levels | 10 weeks | [118] |
| Animal study | Male wild-type C57BL/6 J mice (8–10 weeks old) in a diet-induced-obesity (DIO) model. Male ob/ob C57BL/6 J mice (3–5 weeks old); groups including DIO control group, DIO + 3% curcumin, ob/ob control group or ob/ob. + 3% curcumin | ↓BW and body fat ↑Glycemic status and insulin sensitivity ↓Adipose, hepatic and systemic inflammation ↑mRNA expression of adiponectin ↓Hepatic NF-κB activity | 60 days | [119] |
| Animal study | Male Sprague–Dawley rats in an HF-diet-induced obesity model; control group, high curcumin (5.00 g/kg BW, HFD group, HFD + low curcumin (1.25 g/kg diet) group or HFD + high curcumin (5.00 g/kg diet) group | ↓BW, blood glucose, insulin, leptin, TNF-α ↓Insulin resistance and leptin resistance | 4 weeks | [120] |
| In vitro study | 3T3-L1 cells treated with curcumin (0–30 μM) | ↑Apoptosis at 30 μM ↓Glycerol release ↓MAPK phosphorylation | 2 and 24 h | [121] |
| In vitro study | Primary cell culture from epididymal fat pads treated with curcumin (0–20 μM) | ↔Wnt signaling ↓Inflammatory and oxidative pathway ↓NF-κB signaling | 0–60 min | [113] |
| In vitro study | 3T3-L1 cells treated with curcumin (0–100 μM) | ↓ Adipocyte differentiation and lipid accumulation ↓FAS, PPARγ | 0–8 days | [122] |
| In vitro study | Rabbit subcutaneous adipocytes treated with curcumin (0–20 μg/mL) | ↑Cholesterol efflux from adipocytes ↑PPARγ, LXRα, ABCA1 | 24 h | [123] |
| In vitro study | 3T3-L1 cells treated with curcumin (0–30 μM) | ↓Adipocyte differentiation and fat accumulation ↓Cell viability ↓C/EBPβ, PPARγ and C/EBPα | 18, 24 and 48 h, 6 days | [77] |
| In vitro study | 3T3-L1 cells treated with curcumin (0–50 μM) for | ↑AMPK PPARγ ↑Phosphorylation of ACC ↓Fat accumulation | 8 days | [124] |
| In vitro study | 3T3-L1 cells treated with curcumin (0, 5,10 and 20 μM) | ↓Adipocyte differentiation, fat accumulation and adipogenesis ↑fat oxidation ↓ACC ↑AMPK activation ↑Apoptosis | 24 h | [116] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasprzak-Drozd, K.; Oniszczuk, T.; Gancarz, M.; Kondracka, A.; Rusinek, R.; Oniszczuk, A. Curcumin and Weight Loss: Does It Work? Int. J. Mol. Sci. 2022, 23, 639. https://doi.org/10.3390/ijms23020639
Kasprzak-Drozd K, Oniszczuk T, Gancarz M, Kondracka A, Rusinek R, Oniszczuk A. Curcumin and Weight Loss: Does It Work? International Journal of Molecular Sciences. 2022; 23(2):639. https://doi.org/10.3390/ijms23020639
Chicago/Turabian StyleKasprzak-Drozd, Kamila, Tomasz Oniszczuk, Marek Gancarz, Adrianna Kondracka, Robert Rusinek, and Anna Oniszczuk. 2022. "Curcumin and Weight Loss: Does It Work?" International Journal of Molecular Sciences 23, no. 2: 639. https://doi.org/10.3390/ijms23020639
APA StyleKasprzak-Drozd, K., Oniszczuk, T., Gancarz, M., Kondracka, A., Rusinek, R., & Oniszczuk, A. (2022). Curcumin and Weight Loss: Does It Work? International Journal of Molecular Sciences, 23(2), 639. https://doi.org/10.3390/ijms23020639