
High-Efficiency Reducing Strain for Producing Selenium Nanoparticles Isolated from Marine Sediment



Abstract
1. Introduction
2. Results and Discussion
2.1. Isolation and Characterization of a High-Efficiency Reducing Strain for Producing SeNPs
2.2. The Reduction Location of SeO32− in Bacillus sp. Q33
2.3. Synthesis and Characterization of SeNPs
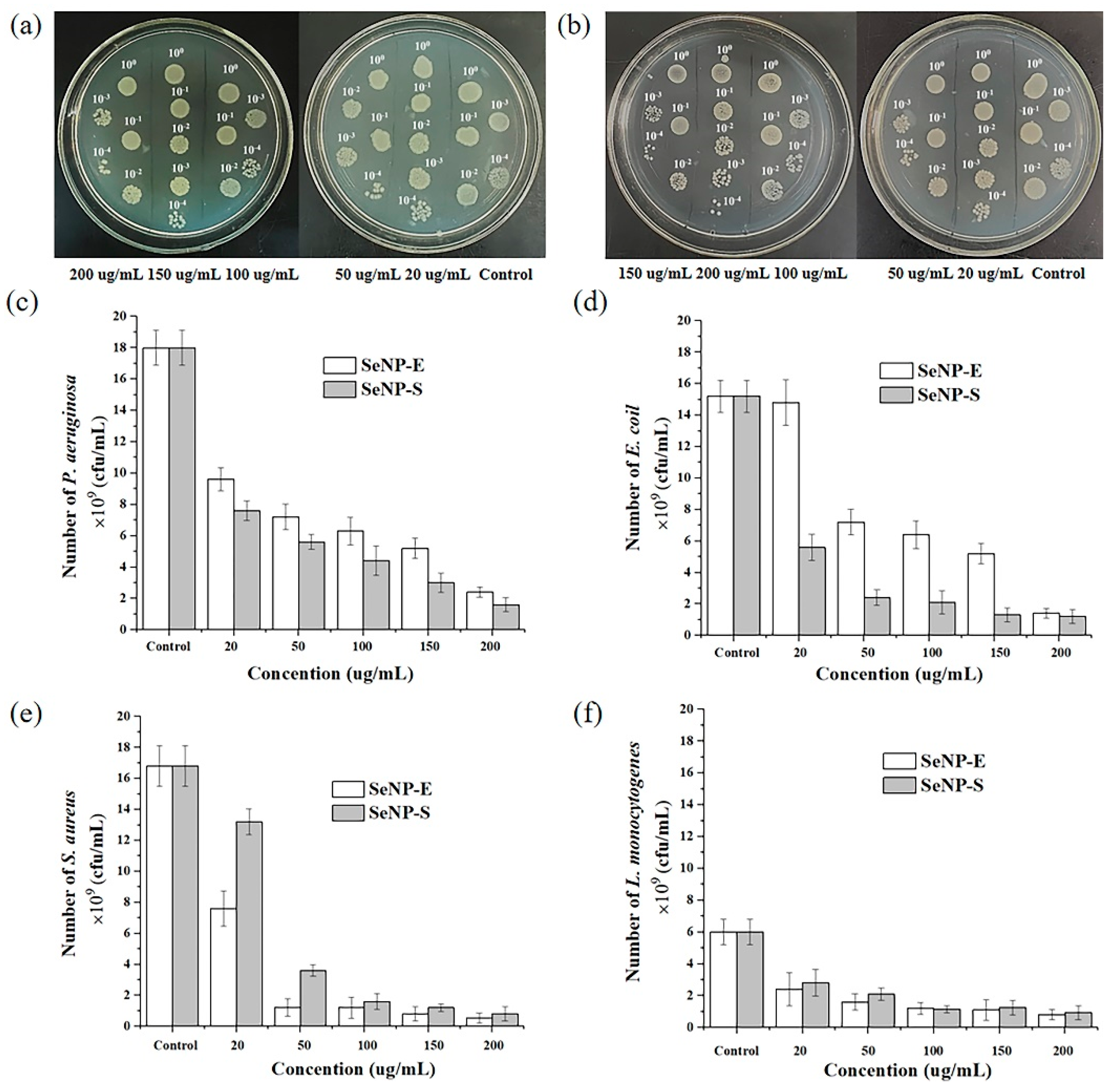
2.4. Antibacterial Activity of Selenium Nanoparticles
2.5. Cytotoxicity of Selenium Nanoparticles
3. Conclusions
4. Materials and Methods
4.1. Materials
4.2. Screening and Identification of High-Efficiency Reducing Strains for Producing Se Nanoparticles
4.2.1. Screening of Strains Capable of Reducing Sodium Selenite
4.2.2. Sodium Selenite Tolerance of the Strain
4.2.3. Reducing Rate of Sodium Selenite by the Strain
4.2.4. Strain Identification
4.3. Determination of SeO32− Reducing Location in Bacillus sp. Q33
4.4. Synthesis and Characterization of SeNPs
4.4.1. Synthesis of SeNPs-S by Whole Cells of Bacillus sp. Q33
4.4.2. Synthesis of SeNPs-E by the Extracellular Extract of Bacillus sp. Q33
4.4.3. Characterization of SeNPs-S and SeNPs-E
4.5. Biological Activity of SeNPs
4.5.1. Antibacterial Activity of SeNPs
Inhibitory rate (%) = (CFU Control − CFU Experimental group)/CFUControl
4.5.2. Cytotoxicity of SeNPs
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, J.S.; Spallholz, J.E. Toxicity of selenium compounds and nanoselenium particles. In General, Applied and Systems Toxicology; John Wiley & Sons: New York, NY, USA, 2011; pp. 787–802. [Google Scholar]
- Haug, A.; Graham, R.D.; Christophersen, O.A.; Lyons, G.H. How to use the world’s scarce selenium resources efficiently to increase the selenium concentration in food. Microb. Ecol. Health Dis. 2007, 19, 209–228. [Google Scholar] [PubMed]
- Stolz, J.F.; Basu, P.; Santini, J.M.; Oremland, R.S. Arsenic and selenium in microbial metabolism (Review). Annu. Rev. Microbiol. 2006, 60, 107–130. [Google Scholar] [CrossRef]
- Skalickova, S.; Milosavjevic, V.; Cihalova, K.; Horky, P.; Richtera, L.; Adam, V. Selenium nanoparticles as a nutritional supplement. Nutrition 2017, 33, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Zhang, H.; Hawthorn, L.; Ganther, H.E.; Ip, C. Delineation of the molecular basis for selenium-induced growth arrest in human prostate cancer cells by oligonucleotide array. Cancer Res. 2003, 63, 52–59. [Google Scholar] [PubMed]
- Turner, R.J.; Weiner, J.H.; Taylor, D.E. Selenium metabolism in Escherichia coli. BioMetals 1998, 11, 223–227. [Google Scholar] [CrossRef] [PubMed]
- McCarty, S.; Chasteen, T.; Marshall, M.; Ray, F.; Reinhard, B. Phototrophic bacteria produce volatile, methylated sulfur and selenium compounds. FEMS Microbiol. Lett. 1993, 112, 93–98. [Google Scholar] [CrossRef]
- Dowdle, P.R.; Oremland, R.S. Microbial oxidation of elemental selenium in soil slurries and bacterial cultures. Environ. Sci. Technol. 1998, 32, 3749–3755. [Google Scholar] [CrossRef]
- Li, B.Z.; Liu, N.; Li, Y.Q.; Jing, W.X.; Fan, J.H.; Li, D.; Zhang, L.Y.; Zhang, X.F.; Zhang, X.F.; Zhang, Z.M.; et al. Reduction of selenite to red elemental selenium by Rhodopseudomonas palustris strain N. PLoS ONE 2014, 9, e95955. [Google Scholar] [CrossRef]
- Yu, B.; Zhang, Y.B.; Zheng, W.J.; Fan, C.; Chen, T. Positive surface charge enhances selective cellular uptake and anticancer efficacy of selenium nanoparticles. Inorg. Chem. 2012, 51, 8956–8963. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Li, N.; Chen, J. A subchronic toxicity study of elemental Nano-Se in Sprague-Dawley rats. Life Sci. 2005, 76, 1989–2003. [Google Scholar] [CrossRef]
- Savita, C.; Ahmad, U.; Mehta, S.K. Selenium nanomaterials: An overview of recent developments in synthesis, properties and potential applications (Review). Prog. Mater. Sci. 2016, 83, 270–329. [Google Scholar]
- Zhang, J.; Xu, Y.; Chen, S.; Li, J.; Han, W.; Sun, X.; Wu, D.; Hu, Z.; Wang, L. Enhanced antifouling and antibacterial properties of poly (ether sulfone) membrane modified through blending with sulfonated poly (aryl ether sulfone) and copper nanoparticles. Appl. Surf. Sci. 2018, 434, 806–815. [Google Scholar] [CrossRef]
- Liao, F.; Han, X.; Zhang, Y.; Chen, H.; Xu, C. CTAB-assisted solvothermal synthesis of ultralong t-selenium nanowires and bundles using glucose as green reducing agent. Mater. Lett. 2018, 214, 41–44. [Google Scholar] [CrossRef]
- Taniguchi, M.; Tanaka, T. Clarification of interactions among microorganisms and development of co-culture system for production of useful substances. Adv. Biochem. Eng. Biot. 2004, 90, 35–62. [Google Scholar]
- Oremland, R.; Herbel, M.J.; Blum, J.S.; Langley, S.; Beveridge, T.J.; Ajayan, P.M.; Sutto, T.; Ellis, A.V.; Curran, S. Structural and spectral features of selenium nanospheres produced by Se-respiring bacteria. Appl. Environ. Microb. 2004, 70, 52–60. [Google Scholar] [CrossRef]
- Dhanjal, S.; Cameotra, S.S. Aerobic, biogenesis of selenium nanospheres by Bacillus cereus isolated from coalmine soil. Microb. Cell Fact. 2010, 9, 52. [Google Scholar] [CrossRef]
- Eszenyi, P.; Sztrik, A.; Babka, B.; Prokisch, J. Production of Lactomicrosel and nanosize (100–500 nm) selenium spheres by probiotic lactic acid bacteria. Int. Conf. Food Eng. Biotechnol. 2011, 9, 97–101. [Google Scholar]
- Dhillon, G.S.; Brar, S.K.; Kaur, S.; Verma, M. Green approach for nanoparticle biosynthesisy fungi: Current trends and applications. Crit. Rev. Biotechnol. 2012, 32, 49–73. [Google Scholar] [CrossRef]
- Narayanan, K.B.; Sakthivel, N. Biological synthesis of metal nanoparticles by microbe. Adv. Colloid. Interface. Sci. 2010, 156, 1–13. [Google Scholar] [CrossRef]
- Zare, B.; Babaie, S.; Setayesh, N.; Shahverdi, A.R. Isolation and characterization of a fungus for extracellular synthesis of small selenium nanoparticles. Nanomed. J. 2013, 1, 13–19. [Google Scholar]
- Vetchinkina, E.; Loshchinina, E.; Kursky, V.; Nikitina, V. Reduction of organic and inorganic selenium compounds by the edible medicinal basidiomycete Lentinula edodes and the accumulation of elemental selenium nanoparticles in its mycelium. J. Microbiol. 2013, 51, 829–835. [Google Scholar] [CrossRef]
- Ahmad, M.S.; Yasser, M.M.; Sholkamy, E.N.; Ali, A.M.; Mehanni, M.M. Anticancer activity of biostabilized selenium nanorods synthesized by Streptomyces bikiniensis strain Ess_amA-1. Int. J. Nanomed. 2015, 10, 3389–3401. [Google Scholar]
- Forootanfara, H.; Zare, B.; Fasihi-Bam, H.; Amirpour-Rostami, S.; Ameri, A.; Shakibaie, M.; Torabi, N.M. Biosynthesis and characterization of selenium nanoparticles produced by terrestrial actinomycete Streptomyces microflavus strain FSHJ31. Res. Rev. J. Microbiol. Biotechnol. 2014, 3, 47–53. [Google Scholar]
- Mustafa, T.Y.; Humeyra, I.; Osman, T.; Enes, D. A green nano-biosynthesis of selenium nanoparticles with Tarragon extract: Structural, thermal, and antimicrobial characterization. LWT Food Sci. Technol. 2021, 141, 110969. [Google Scholar]
- Menon, S.; Ks, S.D.; Agarwal, H.; Shanmugam, V.K. Efficacy of Biogenic Selenium Nanoparticles from an extract of ginger towards evaluation on antimicrobial and anti-oxidant activities. Colloid Interfac. Sci. 2019, 29, 1–8. [Google Scholar] [CrossRef]
- Ramamurthy, C.; Sampath, K.; Arunkumar, P.; Kumar, M.S.; Sujatha, V.; Premkumar, K.; Thirunavukkarasu, C. Green synthesis and characterization of selenium nanoparticles and its augmented cytotoxicity with doxorubicin on cancer cells. Bioproc. Biosyst. Eng. 2013, 36, 1131–1139. [Google Scholar] [CrossRef] [PubMed]
- Sowndarya, P.; Ramkumar, G.; Shivakumar, M. Green synthesis of selenium nanoparticles conjugated Clausena dentata plant leaf extract and their insecticidal potential against mosquito vectors. Artif. Cells Nanomed. B 2017, 45, 1490–1495. [Google Scholar] [CrossRef]
- Fardsadegh, B.; Jafarizadeh-Malmiri, H. Aloe vera leaf extract mediated green synthesis of selenium nanoparticles and assessment of their in vitro antimicrobial activity against spoilage fungi and pathogenic bacteria strains. Green Process. Synth. 2019, 8, 399–407. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Robert, A.K.; Michèle, R.P. Marine natural products. Nat. Prod. Rep. 2020, 37, 175–223. [Google Scholar] [CrossRef]
- Fehmida, B.; Muhammad, F.; Esam, I.A.; Muhammad, Y.; Sana, A.A.; Mohammad, A.K.; Ikram, U.; Muhammad, I.N. Bacteria from marine sponges: A source of new drugs. Curr. Drug Metab. 2016, 17, 11–15. [Google Scholar]
- Ashengroph, M.; Hosseini, S.R. A newly isolated Bacillus amyloliquefaciens SRB04 for the synthesis of selenium nanoparticles with potential antibacterial properties. Int. Microbiol. 2020, 24, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhao, H.Y.; Bi, Y.G.; Fan, X.D. Preparation and antioxidant activity of selenium nanoparticles decorated by polysaccharides from Sargassum fusiforme. J. Food Sci. 2021, 86, 977–986. [Google Scholar] [CrossRef] [PubMed]
- Hamza, F.; Vaidya, A.; Apte, M.; Kumara, A.R.; Zinjardea, S. Selenium nanoparticle-enriched biomass of Yarrowia lipolytica enhances growth and survival of Artemia salina. Enzym. Microb. Technol. 2017, 106, 48–54. [Google Scholar] [CrossRef]
- Sandhya, M.V.S.; Rajkumar, K.; Burgula, S. Efficient eco-friendly approach towards bimetallic nanoparticles synthesis and characterization using Exiguobacterium aestuarii by statistical optimization. Green Chem. Lett. Rev. 2019, 12, 420–434. [Google Scholar] [CrossRef]
- Kessi, J.; Hanselmann, K.W. Similarities between the abiotic reduction of selenite with glutathione and the dissimilatory reaction mediated by Rhodospirillum rubrum and Escherichiacoli. J. Biol. Chem. 2004, 24, 50662–50669. [Google Scholar] [CrossRef]
- Nancharaiah, Y.V.; Lens, P.N. Ecology and biotechnology of selenium-respiring bacteria (Review). Microbiol. Mol. Biol. Rev. 2015, 79, 61–80. [Google Scholar] [CrossRef] [PubMed]
- Staicu, L.C.; Ackerson, C.J.; Cornelis, P.; Ye, L.; Berendsen, R.L.; Hunter, W.J.; Noblitt, S.D.; Henry, C.S.; Cappa, J.J.; Montenieri, R.L.; et al. Pseudomonas moraviensis subsp. stanleyae, a bacterial endophyte of hyperaccumulator Stanleya pinnata, is capable of efficient selenite reduction to elemental selenium under aerobic conditions. J. Appl. Biotechnol. 2015, 119, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Lampis, S.; Zonaro, E.; Bertolini, C.; Bernardi, P.; Butler, C.S.; Vallini, G. Delayed formation of zero-valent selenium nanoparticles by Bacillus mycoides SelTE01 as a consequence of selenite reduction under aerobic conditions. Microb. Cell Fact. 2014, 13, 35. [Google Scholar] [CrossRef] [PubMed]
- Helga, F.L.; Laura, C.; María, L.B.; Eduardo, D.; Manuel, C. Speeding up bioproduction of selenium nanoparticles by using Vibrio natriegens as microbial Factory. Sci. Rep. 2017, 7, 16046. [Google Scholar]
- Khoeia, N.S.; Lampis, S.; Zonaroa, E.; Yrjälä, K.; Bernardic, P.; Vallini, G. Insights into selenite reduction and biogenesis of elemental selenium nanoparticles by two environmental isolates of Burkholderia fungorum. New Biotechnol. 2017, 34, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Domokos-Szabolcsy, E.; Marton, L.; Sztrik, A.; Babka, B.; Prokisch, J.; Fari, M. Accumulation of red elemental selenium nanpparticles and their biological effects in tabacum. Plant Growth Regul. 2012, 68, 525–531. [Google Scholar] [CrossRef]
- Salem, S.S.; Moustafa, M.G.F.; Amr, F.; Mohamed, A.A.; Ebtesam, M.A.; Ahmed, A.A.; Tharwat, I.S. Antibacterial, cytotoxicity and larvicidal activity of green synthesized selenium nanoparticles Using Penicillium corylophilum. J. Clust. Sci. 2020, 32, 351–361. [Google Scholar] [CrossRef]
- Nishant, S.; Mausumi, M. Green synthesis and structural characterization of selenium nanoparticles and assessment of their antimicrobial property. Bioprocess Biosyst. Eng. 2015, 38, 1723–1730. [Google Scholar]
- An, C.; Tang, K.; Liu, X.; Qian, Y. Large-scale synthesis of high quality trigonal selenium nanowires. Eur. J. Inorg. Chem. 2003, 17, 3250–3255. [Google Scholar] [CrossRef]
- Senthil kumaran, C.K.; Agilan, S.; Velauthapillai, D.; Muthukumarasamy, N.; Thambidurai, M.; Senthil, T.S.; Balasundaraprabhu, R. Synthesis and characterization of selenium nanowires. ISRN Nanotechnol. 2011, 18, 405101. [Google Scholar] [CrossRef][Green Version]
- Zhang, W.J.; Chen, Z.J.; Liu, H.; Zhang, L.; Gao, P.; Li, D.P. Biosynthesis and structural characteristics of selenium nanoparticles by Pseudomonas alcaliphila. Colloids Surf. B. 2011, 88, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, N.; Mukhopadhyay, M. Biosynthesis and structural characterization of selenium nanoparticles mediated by Zooglea ramigera. Powder Technol. 2013, 244, 26–29. [Google Scholar] [CrossRef]
- Jeong, U.Y.; Xia, Y.N. Synthesis and crystallization of monodisperse spherical colloids of amorphous selenium. Adv. Mater. 2005, 17, 102–106. [Google Scholar]
- Rivastava, P.; Braganca, J.M.; Kowshik, M. In vivo synthesis of selenium nanoparticles by Halococcus salifodinae BK18 and their anti-proliferative properties against HeLa cell line. Biotechnol. Progr. 2014, 30, 1480–1487. [Google Scholar] [CrossRef] [PubMed]
- Diko, C.S.; Zhang, H.; Lian, S.; Fan, S.; Qu, Y. Optimal synthesis conditions and characterization of selenium nanoparticles in Trichoderma sp. WL-Go culture broth. Mater. Chem. Phys. 2020, 246, 122583. [Google Scholar] [CrossRef]
- Keskin, N.O.S.; Vural, O.A.; Abaci, S. Biosynthesis of noble selenium nanoparticles from Lysinibacillus sp. NOSK for antimicrobial, antibiofilm activity, and biocompatibility. Geomicrobiol. J. 2020, 37, 919–928. [Google Scholar] [CrossRef]
- ISO E. 10993-5; Biological Evaluation of Medical Devices. Tests for in vitro cytotoxicity. International Organization for Standardization: Geneva, Switzerland, 2009.
- Li, D.B.; Cheng, Y.Y.; Wu, C.; Li, W.W.; Li, N.; Yang, Z.C.; Tong, Z.H.; Yu, H.Q. Selenite reduction by Shewanella oneidensis MR-1 is mediated by fumarate reductase in periplasm. Sci. Rep. 2013, 4, 3735. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | k | R2 | t1/2 (h) | Reduction Rate of SeO32− |
|---|---|---|---|---|
| 72 | 0.1081 | 0.966 | 6.41 | 93.7% |
| 33 | 0.1297 | 0.973 | 5.34 | 96.7% |
| 21 | 0.1162 | 0.957 | 5.96 | 95.2% |
| Sample | Inhibition Rate (%) | |||
|---|---|---|---|---|
| P. aeruginosa | E. coli | S. aureus | L. monocytogenes | |
| SeNPs-S | 91.1 | 92.1 | 95.2 | 84.5 |
| SeNPs-E | 86.7 | 90.8 | 96.8 | 86.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Li, Z.; Zhang, L.; Lei, Z.; Jin, L.; Cao, J.; Quan, C. High-Efficiency Reducing Strain for Producing Selenium Nanoparticles Isolated from Marine Sediment. Int. J. Mol. Sci. 2022, 23, 11953. https://doi.org/10.3390/ijms231911953
Zhang L, Li Z, Zhang L, Lei Z, Jin L, Cao J, Quan C. High-Efficiency Reducing Strain for Producing Selenium Nanoparticles Isolated from Marine Sediment. International Journal of Molecular Sciences. 2022; 23(19):11953. https://doi.org/10.3390/ijms231911953
Chicago/Turabian StyleZhang, Liying, Zhuting Li, Lei Zhang, Zhixiao Lei, Liming Jin, Jijuan Cao, and Chunshan Quan. 2022. "High-Efficiency Reducing Strain for Producing Selenium Nanoparticles Isolated from Marine Sediment" International Journal of Molecular Sciences 23, no. 19: 11953. https://doi.org/10.3390/ijms231911953
APA StyleZhang, L., Li, Z., Zhang, L., Lei, Z., Jin, L., Cao, J., & Quan, C. (2022). High-Efficiency Reducing Strain for Producing Selenium Nanoparticles Isolated from Marine Sediment. International Journal of Molecular Sciences, 23(19), 11953. https://doi.org/10.3390/ijms231911953