
A Tale of Two Lobsters—Transcriptomic Analysis Reveals a Potential Gap in the RNA Interference Pathway in the Tropical Rock Lobster Panulirus ornatus

 ,
, 
Abstract
1. Introduction
1.1. Core RNAi Mechanism
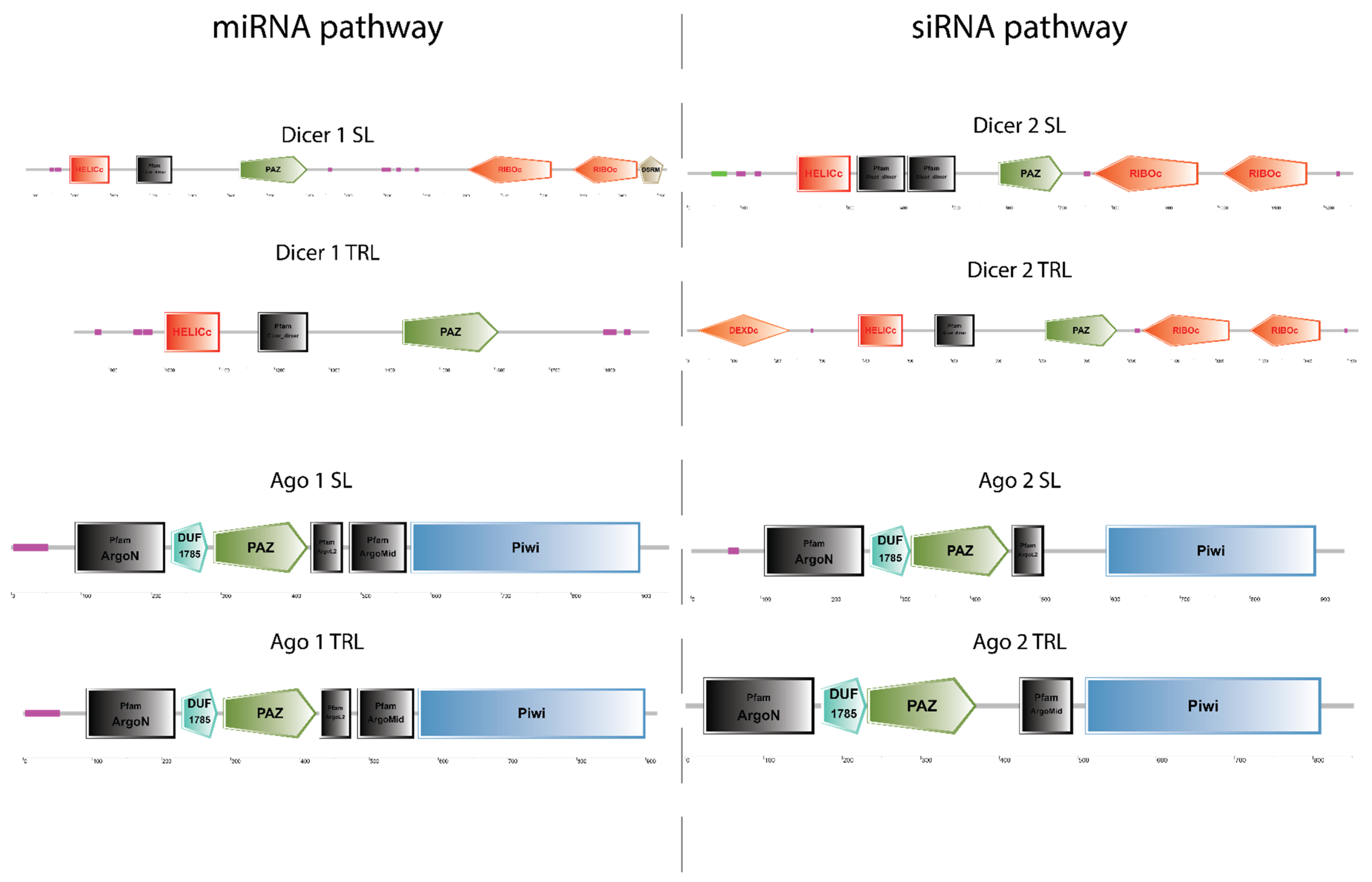
1.2. Dicer and Argonaute Domain Architecture
1.3. SID-1 dsRNA Channels and dsRNA Uptake
2. Results
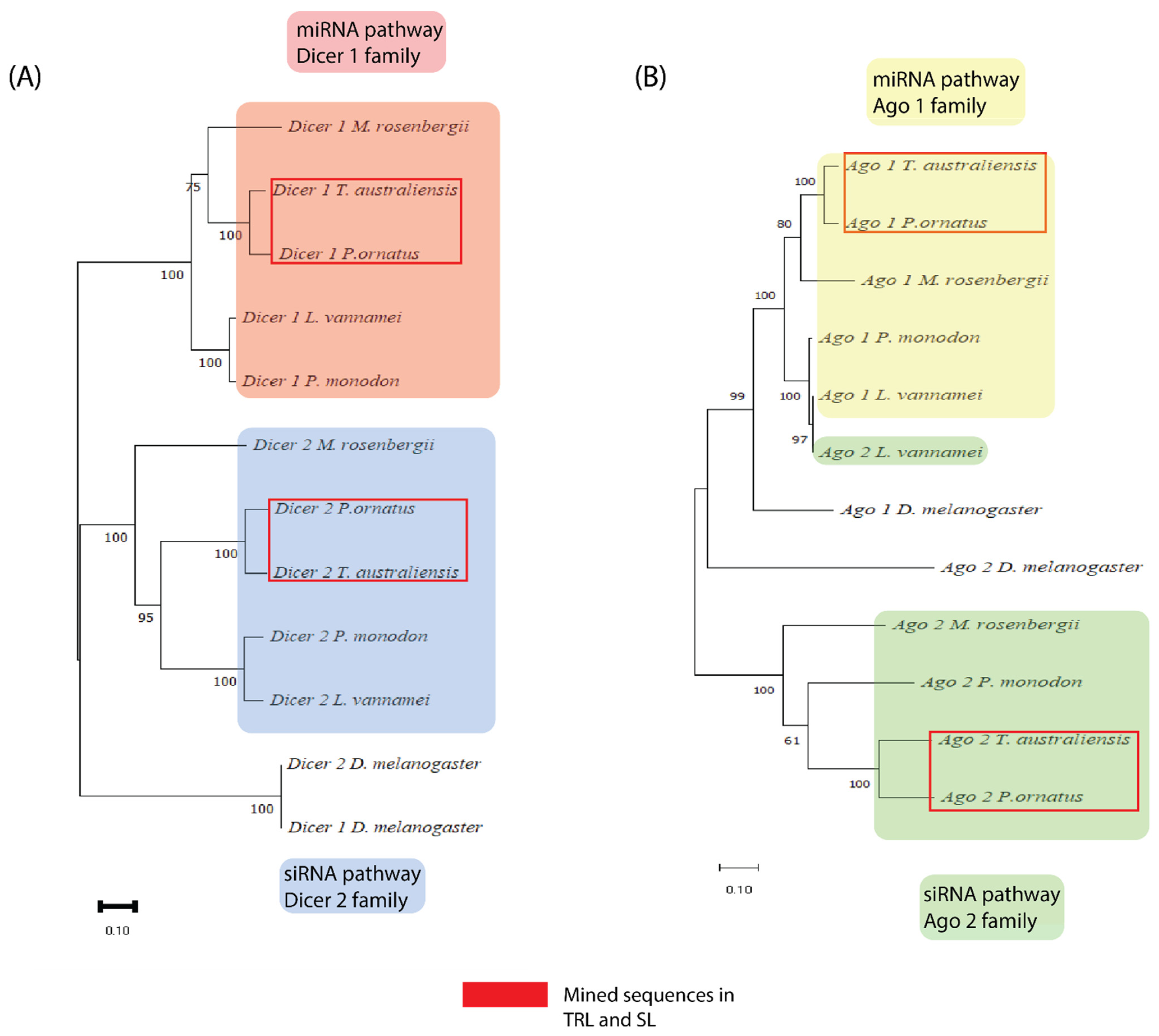
2.1. All Core RNAi Genes Are Present in Both P. ornatus and T. australiensis
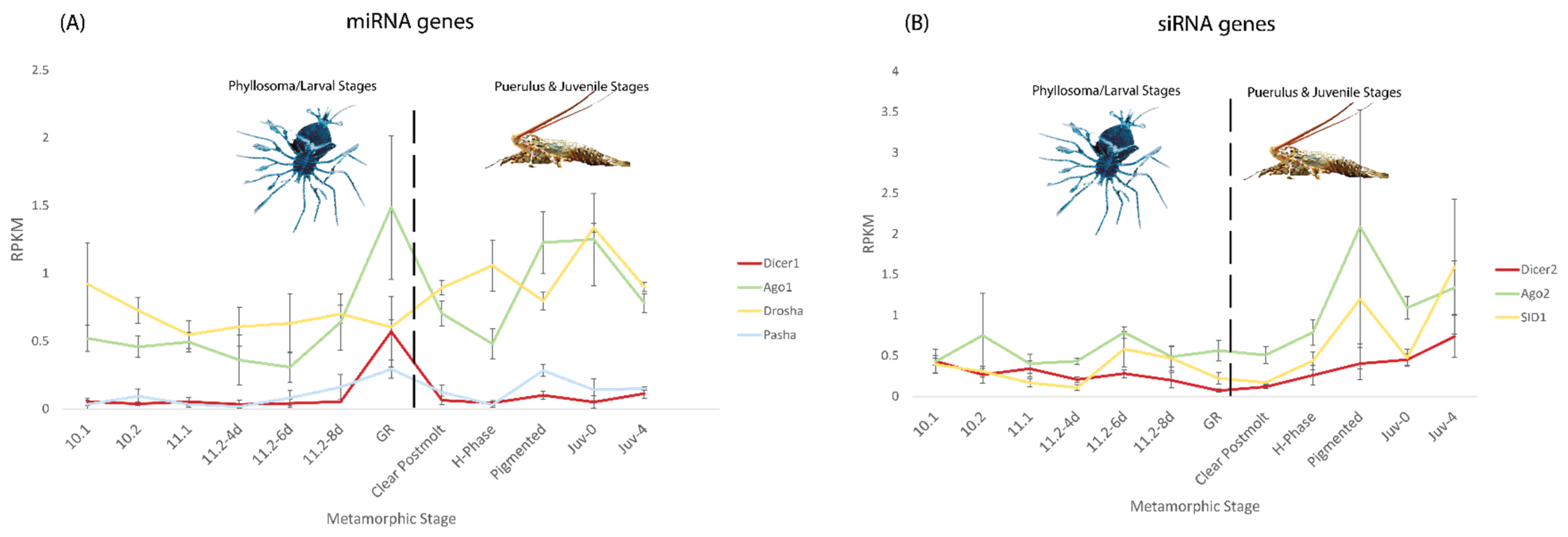
2.2. Variable Expression of siRNA and miRNA Genes in P. ornatus across Metamorphic Stages
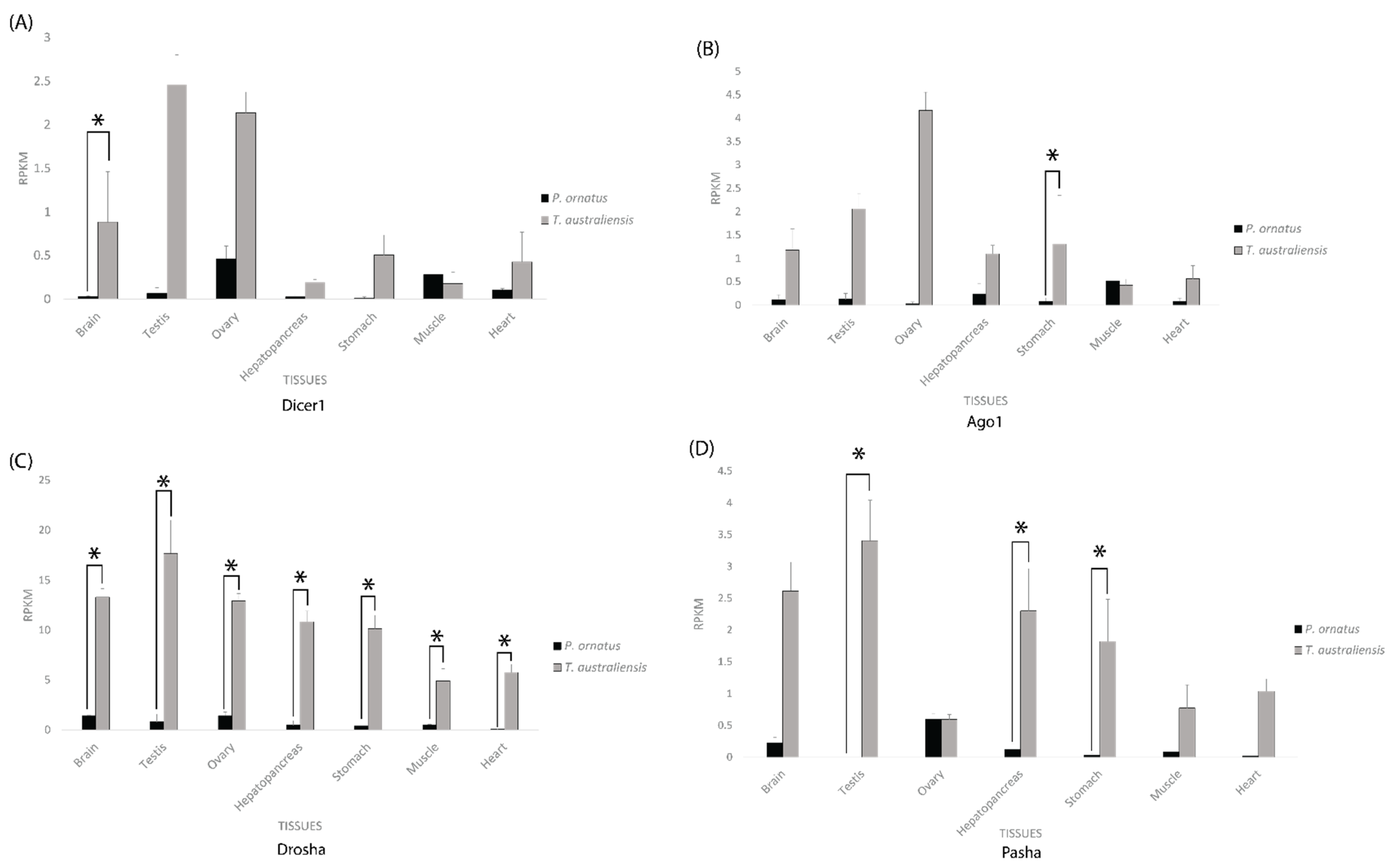
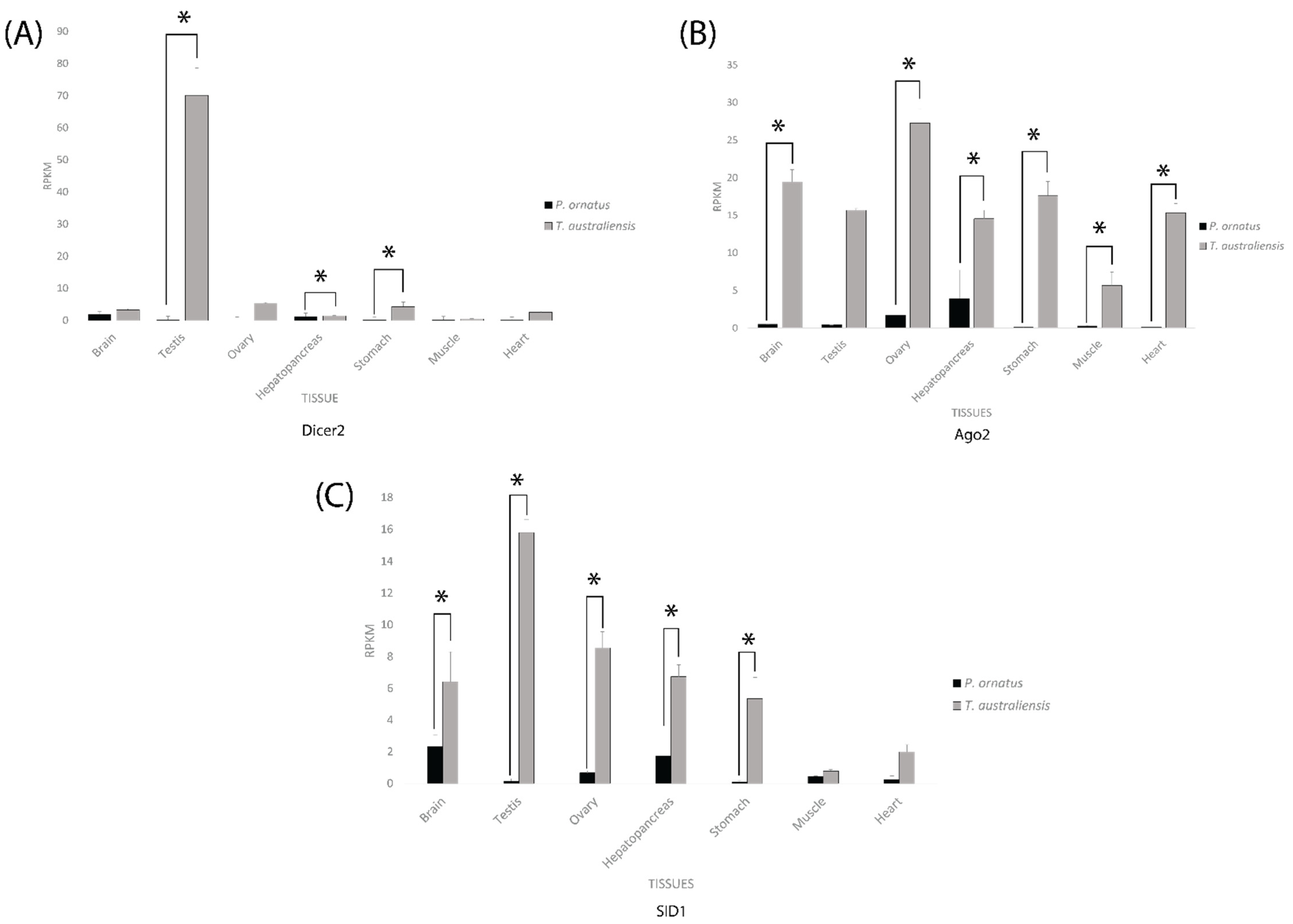
2.3. Core RNAi Machinery Is Expressed Significantly Higher in T. australiensis Than P. ornatus across Adult and Sub-Adult Tissues
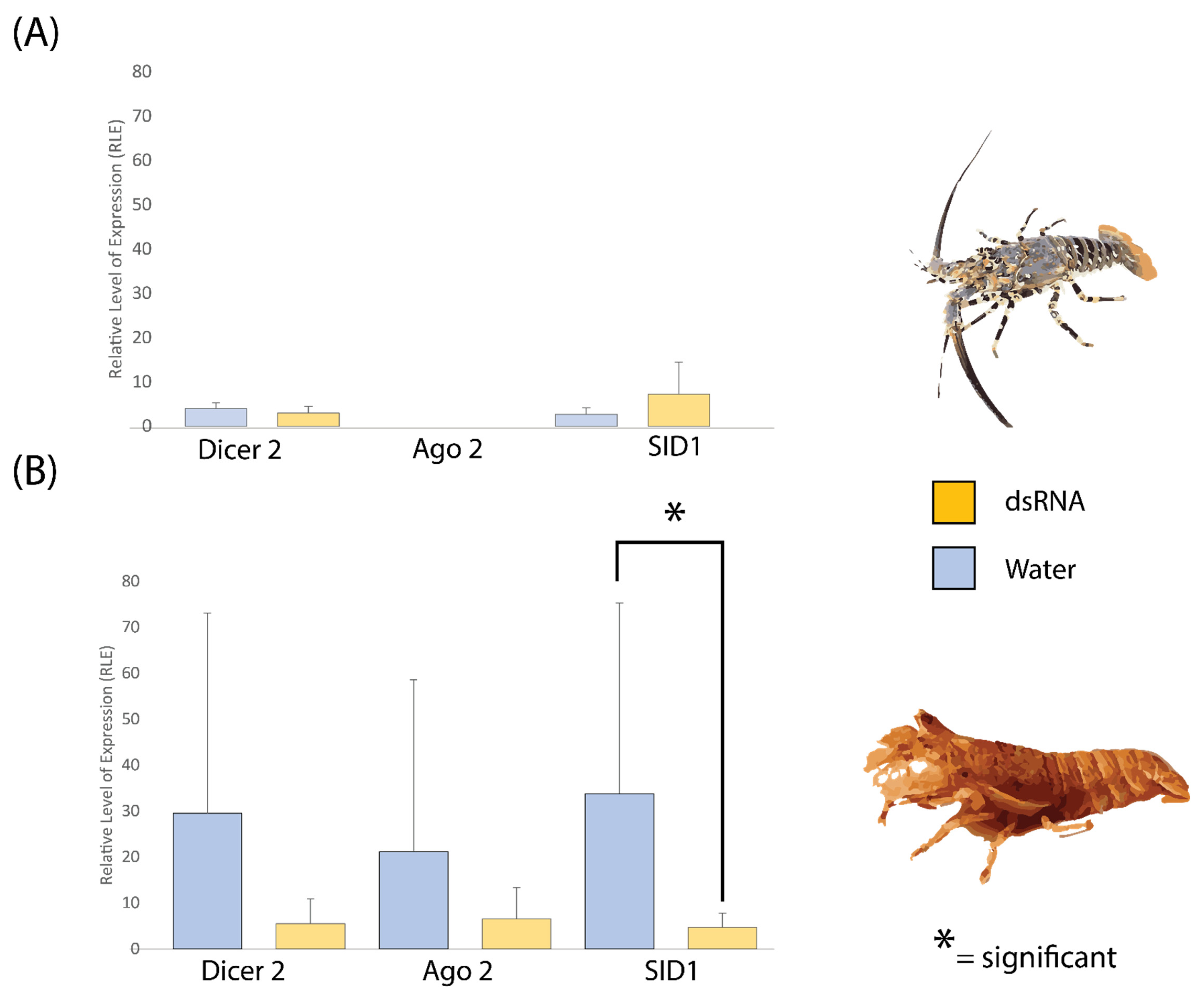
2.4. dsRNA Exposure Does Not Modulate siRNA Pathway Expression in P. ornatus and T. australiensis
3. Discussion
4. Materials and Methods
4.1. RNAi Gene orthologue Collection
4.2. Transcriptome Mining in P. ornatus and T. australiensis and SMART Annotation of RNAi Genes
4.3. Phylogenetic Analysis of Dicer and Argonaute Genes
4.4. Transcriptome Quantification and Tissue Specific RNAi Expression
4.5. RNAi Modulation, RNA Extraction, and RT qPCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Das, P.R.; Sherif, S.M. Application of Exogenous dsRNAs-induced RNAi in Agriculture: Challenges and Triumphs. Front. Plant Sci. 2020, 11, 946. [Google Scholar] [CrossRef] [PubMed]
- Setten, R.L.; Rossi, J.J.; Han, S.-P. The current state and future directions of RNAi-based therapeutics. Nat. Rev. Drug Discov. 2019, 18, 421–446. [Google Scholar] [CrossRef] [PubMed]
- Abo-Al-Ela, H.G. RNA Interference in Aquaculture: A Small Tool for Big Potential. J. Agric. Food Chem. 2021, 69, 4343–4355. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.V.; Christiaens, O.; Bossier, P.; Smagghe, G. RNA interference in shrimp and potential applications in aquaculture. Rev. Aquac. 2018, 10, 573–584. [Google Scholar] [CrossRef]
- Napoli, C.; Lemieux, C.; Jorgensen, R. Introduction of a Chimeric Chalcone Synthase Gene into Petunia Results in Reversible Co-Suppression of Homologous Genes in trans. Plant Cell 1990, 2, 279–289. [Google Scholar] [CrossRef]
- Agrawal, N.; Dasaradhi, P.V.N.; Mohmmed, A.; Malhotra, P.; Bhatnagar, R.K.; Mukherjee, S.K. RNA Interference: Biology, Mechanism, and Applications. Microbiol. Mol. Biol. Rev. 2003, 67, 657. [Google Scholar] [CrossRef]
- Zhang, R.; Jing, Y.; Zhang, H.; Niu, Y.; Liu, C.; Wang, J.; Zen, K.; Zhang, C.-Y.; Li, D. Comprehensive Evolutionary Analysis of the Major RNA-Induced Silencing Complex Members. Sci. Rep. 2018, 8, 14189. [Google Scholar] [CrossRef]
- Nishida, K.M.; Saito, K.; Mori, T.; Kawamura, Y.; Nagami-Okada, T.; Inagaki, S.; Siomi, H.; Siomi, M.C. Gene silencing mechanisms mediated by Aubergine piRNA complexes in Drosophila male gonad. RNA 2007, 13, 1911–1922. [Google Scholar] [CrossRef]
- Lucas, K.; Raikhel, A.S. Insect microRNAs: Biogenesis, expression profiling and biological functions. Insect Biochem. Mol. Biol. 2013, 43, 24–38. [Google Scholar] [CrossRef]
- Asgari, S. MicroRNA functions in insects. Insect Biochem. Mol. Biol. 2013, 43, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Daniels, S.M.; Gatignol, A. The Multiple Functions of TRBP, at the Hub of Cell Responses to Viruses, Stress, and Cancer. Microbiol. Mol. Biol. Rev. 2012, 76, 652–666. [Google Scholar] [CrossRef] [PubMed]
- Hartig, J.V.; Förstemann, K. Loqs-PD and R2D2 define independent pathways for RISC generation in Drosophila. Nucleic Acids Res. 2011, 39, 3836–3851. [Google Scholar] [CrossRef] [PubMed]
- Marques, J.T.; Kim, K.; Wu, P.-H.; Alleyne, T.M.; Jafari, N.; Carthew, R.W. Loqs and R2D2 act sequentially in the siRNA pathway in Drosophila. Nat. Struct. Mol. Biol. 2010, 17, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Hur, I.; Park, S.-Y.; Kim, Y.-K.; Suh, M.R.; Kim, V.N. The role of PACT in the RNA silencing pathway. EMBO J. 2006, 25, 522–532. [Google Scholar] [CrossRef]
- Castillejo-López, C.; Cai, X.; Fahmy, K.; Baumgartner, S. Drosophila exoribonuclease nibbler is a tumor suppressor, acts within the RNAi machinery and is not enriched in the nuage during early oogenesis. Hereditas 2018, 155, 12. [Google Scholar] [CrossRef]
- Viet Dung, N.; Christiaens, O.; Le Van Bao, D.; De Vos, S.; MacRae, T.H.; Smagghe, G.; Bossier, P. Identification of RNAi-related genes and transgenerational efficiency of RNAi in Artemia franciscana. Aquaculture 2019, 501, 285–292. [Google Scholar] [CrossRef]
- Schuster, S.; Miesen, P.; Van Rij, R.P. Antiviral RNAi in Insects and Mammals: Parallels and Differences. Viruses 2019, 11, 448. [Google Scholar] [CrossRef]
- Mongelli, V.; Saleh, M.-C. Bugs Are Not to Be Silenced: Small RNA Pathways and Antiviral Responses in Insects. Annu. Rev. Virol. 2016, 3, 573–589. [Google Scholar] [CrossRef]
- Claycomb, J.M. Ancient Endo-siRNA Pathways Reveal New Tricks. Curr. Biol. 2014, 24, R703–R715. [Google Scholar] [CrossRef]
- Masliah, G.; Maris, C.; König, S.L.; Yulikov, M.; Aeschimann, F.; Malinowska, A.L.; Mabille, J.; Weiler, J.; Holla, A.; Hunziker, J.; et al. Structural basis of si RNA recognition by TRBP double-stranded RNA binding domains. EMBO J. 2018, 37, e97089. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ye, X.; Jiang, F.; Liang, C.; Chen, D.; Peng, J.; Kinch, L.N.; Grishin, N.V.; Liu, Q. C3PO, an Endoribonuclease That Promotes RNAi by Facilitating RISC Activation. Science 2009, 325, 750–753. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Rand Tim, A.; Kalidas, S.; Du, F.; Kim, H.-E.; Smith Dean, P.; Wang, X. R2D2, a Bridge Between the Initiation and Effector Steps of the Drosophila RNAi Pathway. Science 2003, 301, 1921–1925. [Google Scholar] [CrossRef]
- Müller, M.; Fazi, F.; Ciaudo, C. Argonaute Proteins: From Structure to Function in Development and Pathological Cell Fate Determination. Front. Cell Dev. Biol. 2020, 7, 360. [Google Scholar] [CrossRef] [PubMed]
- Dowling, D.; Pauli, T.; Donath, A.; Meusemann, K.; Podsiadlowski, L.; Petersen, M.; Peters, R.S.; Mayer, C.; Liu, S.; Zhou, X.; et al. Phylogenetic Origin and Diversification of RNAi Pathway Genes in Insects. Genome Biol. Evol. 2017, 8, evw281. [Google Scholar] [CrossRef] [PubMed]
- Golyaev, V.; Candresse, T.; Rabenstein, F.; Pooggin, M.M. Plant virome reconstruction and antiviral RNAi characterization by deep sequencing of small RNAs from dried leaves. Sci. Rep. 2019, 9, 19268. [Google Scholar] [CrossRef]
- Chang, S.-S.; Zhang, Z.; Liu, Y. RNA Interference Pathways in Fungi: Mechanisms and Functions. Annu. Rev. Microbiol. 2012, 66, 305–323. [Google Scholar] [CrossRef]
- Ventura, T.; Manor, R.; Aflalo, E.D.; Weil, S.; Rosen, O.; Sagi, A. Timing Sexual Differentiation: Full Functional Sex Reversal Achieved Through Silencing of a Single Insulin-Like Gene in the Prawn, Macrobrachium rosenbergii1. Biol. Reprod. 2012, 86, 90. [Google Scholar] [CrossRef]
- Lopez, S.B.G.; Guimarães-Ribeiro, V.; Rodriguez, J.V.G.; Dorand, F.A.P.S.; Salles, T.S.; Sá-Guimarães, T.E.; Alvarenga, E.S.L.; Melo, A.C.A.; Almeida, R.V.; Moreira, M.F. RNAi-based bioinsecticide for Aedes mosquito control. Sci. Rep. 2019, 9, 4038. [Google Scholar] [CrossRef]
- Wynant, N.; Santos, D.; Vanden Broeck, J. The evolution of animal Argonautes: Evidence for the absence of antiviral AGO Argonautes in vertebrates. Sci. Rep. 2017, 7, 9230. [Google Scholar] [CrossRef]
- Kidwell, M.A.; Chan, J.M.; Doudna, J.A. Evolutionarily Conserved Roles of the Dicer Helicase Domain in Regulating RNA Interference Processing. J. Biol. Chem. 2014, 289, 28352–28362. [Google Scholar] [CrossRef] [PubMed]
- Tsutsumi, A.; Kawamata, T.; Izumi, N.; Seitz, H.; Tomari, Y. Recognition of the pre-miRNA structure by Drosophila Dicer-1. Nat. Struct. Mol. Biol. 2011, 18, 1153–1158. [Google Scholar] [CrossRef] [PubMed]
- Ciechanowska, K.; Pokornowska, M.; Kurzyńska-Kokorniak, A. Genetic Insight into the Domain Structure and Functions of Dicer-Type Ribonucleases. Int. J. Mol. Sci. 2021, 22, 616. [Google Scholar] [CrossRef] [PubMed]
- Elif; Fukunaga, R.; Lu, G.; Dutcher, R.; Wang, Y.; Traci; Phillip. Phosphate and R2D2 Restrict the Substrate Specificity of Dicer-2, an ATP-Driven Ribonuclease. Mol. Cell 2011, 42, 172–184. [Google Scholar] [CrossRef]
- Gleghorn, M.L.; Maquat, L.E. ‘Black sheep’ that don’t leave the double-stranded RNA-binding domain fold. Trends Biochem. Sci. 2014, 39, 328–340. [Google Scholar] [CrossRef]
- Pokornowska, M.; Milewski, M.C.; Ciechanowska, K.; Szczepańska, A.; Wojnicka, M.; Radogostowicz, Z.; Figlerowicz, M.; Kurzynska-Kokorniak, A. The RNA–RNA base pairing potential of human Dicer and Ago2 proteins. Cell. Mol. Life Sci. 2020, 77, 3231–3244. [Google Scholar] [CrossRef]
- Kurzynska-Kokorniak, A.; Koralewska, N.; Pokornowska, M.; Urbanowicz, A.; Tworak, A.; Mickiewicz, A.; Figlerowicz, M. The many faces of Dicer: The complexity of the mechanisms regulating Dicer gene expression and enzyme activities. Nucleic Acids Res. 2015, 43, 4365–4380. [Google Scholar] [CrossRef]
- MacRae Ian, J.; Zhou, K.; Li, F.; Repic, A.; Brooks Angela, N.; Cande, W.Z.; Adams Paul, D.; Doudna Jennifer, A. Structural Basis for Double-Stranded RNA Processing by Dicer. Science 2006, 311, 195–198. [Google Scholar] [CrossRef]
- Park, J.-E.; Heo, I.; Tian, Y.; Simanshu, D.K.; Chang, H.; Jee, D.; Patel, D.J.; Kim, V.N. Dicer recognizes the 5′ end of RNA for efficient and accurate processing. Nature 2011, 475, 201–205. [Google Scholar] [CrossRef]
- Lau, P.-W.; Guiley, K.Z.; De, N.; Potter, C.S.; Carragher, B.; Macrae, I.J. The molecular architecture of human Dicer. Nat. Struct. Mol. Biol. 2012, 19, 436–440. [Google Scholar] [CrossRef]
- Elbashir, S.M. Functional anatomy of siRNAs for mediating efficient RNAi in Drosophila melanogaster embryo lysate. EMBO J. 2001, 20, 6877–6888. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, A.W. Ribonuclease III mechanisms of double-stranded RNA cleavage. Wiley Interdiscip. Rev. RNA 2014, 5, 31–48. [Google Scholar] [CrossRef] [PubMed]
- Doyle, M.; Badertscher, L.; Jaskiewicz, L.; Güttinger, S.; Jurado, S.; Hugenschmidt, T.; Kutay, U.; Filipowicz, W. The double-stranded RNA binding domain of human Dicer functions as a nuclear localization signal. RNA 2013, 19, 1238–1252. [Google Scholar] [CrossRef] [PubMed]
- Schirle Nicole, T.; MacRae Ian, J. The Crystal Structure of Human Argonaute2. Science 2012, 336, 1037–1040. [Google Scholar] [CrossRef] [PubMed]
- Kwak, P.B.; Tomari, Y. The N domain of Argonaute drives duplex unwinding during RISC assembly. Nat. Struct. Mol. Biol. 2012, 19, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Ming, D.; Wall, M.E.; Sanbonmatsu, K.Y. Domain motions of Argonaute, the catalytic engine of RNA interference. BMC Bioinform. 2007, 8, 470. [Google Scholar] [CrossRef]
- Reddy Chichili, V.P.; Kumar, V.; Sivaraman, J. Linkers in the structural biology of protein-protein interactions. Protein Sci. 2013, 22, 153–167. [Google Scholar] [CrossRef]
- Ma, J.-B.; Ye, K.; Patel, D.J. Structural basis for overhang-specific small interfering RNA recognition by the PAZ domain. Nature 2004, 429, 318–322. [Google Scholar] [CrossRef]
- Boland, A.; Tritschler, F.; Heimstädt, S.; Izaurralde, E.; Weichenrieder, O. Crystal structure and ligand binding of the MID domain of a eukaryotic Argonaute protein. EMBO Rep. 2010, 11, 522–527. [Google Scholar] [CrossRef]
- Frank, F.; Sonenberg, N.; Nagar, B. Structural basis for 5′-nucleotide base-specific recognition of guide RNA by human AGO2. Nature 2010, 465, 818–822. [Google Scholar] [CrossRef]
- Wu, J.E.; Yang, J.; Cho, W.C.; Zheng, Y. Argonaute proteins: Structural features, functions and emerging roles. J. Adv. Res. 2020, 24, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Van Roessel, P.; Brand, A.H. Spreading silence with Sid. Genome Biol. 2004, 5, 208. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Koutmou, K.S.; Leahy, D.J.; Li, M. Systemic RNA Interference Deficiency-1 (SID-1) Extracellular Domain Selectively Binds Long Double-stranded RNA and Is Required for RNA Transport by SID-1. J. Biol. Chem. 2015, 290, 18904–18913. [Google Scholar] [CrossRef] [PubMed]
- Feinberg, E.H.; Hunter, C.P. Transport of dsRNA into Cells by the Transmembrane Protein SID-1. Science 2003, 301, 1545. [Google Scholar] [CrossRef] [PubMed]
- McEwan, D.L.; Weisman, A.S.; Hunter, C.P. Uptake of extracellular double-stranded RNA by SID-2. Mol. Cell 2012, 47, 746–754. [Google Scholar] [CrossRef]
- Saleh, M.-C.; van Rij, R.P.; Hekele, A.; Gillis, A.; Foley, E.; O’Farrell, P.H.; Andino, R. The endocytic pathway mediates cell entry of dsRNA to induce RNAi silencing. Nat. Cell Biol. 2006, 8, 793–802. [Google Scholar] [CrossRef]
- Tatematsu, M.; Funami, K.; Seya, T.; Matsumoto, M. Extracellular RNA Sensing by Pattern Recognition Receptors. J. Innate Immun. 2018, 10, 398–406. [Google Scholar] [CrossRef]
- Wynant, N.; Santos, D.; Van Wielendaele, P.; Vanden Broeck, J. Scavenger receptor-mediated endocytosis facilitates RNA interference in the desert locust, Schistocerca gregaria. Insect Mol. Biol. 2014, 23, 320–329. [Google Scholar] [CrossRef]
- Shih, J.D.; Hunter, C.P. SID-1 is a dsRNA-selective dsRNA-gated channel. RNA 2011, 17, 1057–1065. [Google Scholar] [CrossRef]
- Xu, W.; Han, Z. Cloning and Phylogenetic Analysis of Sid-1-Like Genes from Aphids. J. Insect Sci. 2008, 8, 30. [Google Scholar] [CrossRef]
- Maruekawong, K.; Tirasophon, W.; Panyim, S.; Attasart, P. Involvement of Lv SID-1 in dsRNA uptake in Litopenaeus vannamei. Aquaculture 2018, 482, 65–72. [Google Scholar] [CrossRef]
- Pratt, A.J.; Rambo, R.P.; Lau, P.-W.; Macrae, I.J. Preparation and Characterization of the Extracellular Domain of Human Sid-1. PLoS ONE 2012, 7, e33607. [Google Scholar] [CrossRef] [PubMed]
- Shih, J.D.; Fitzgerald, M.C.; Sutherlin, M.; Hunter, C.P. The SID-1 double-stranded RNA transporter is not selective for dsRNA length. RNA 2009, 15, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Winston, W.M.; Sutherlin, M.; Wright, A.J.; Feinberg, E.H.; Hunter, C.P. Caenorhabditis elegans SID-2 is required for environmental RNA interference. Proc. Natl. Acad. Sci. USA 2007, 104, 10565–10570. [Google Scholar] [CrossRef] [PubMed]
- Maruekawong, K.; Namlamoon, O.; Attasart, P. Systemic gene silencing from oral uptake of dsRNA in Litopenaeus vannamei requires both clathrin-mediated endocytosis and LvSID-1. Aquaculture 2022, 548, 737557. [Google Scholar] [CrossRef]
- Maruekawong, K.; Panyim, S.; Attasart, P. Involvement of endocytosis in cellular uptake of injected dsRNA into hepatopancreas but not in gill of Litopenaeus vannamei. Aquaculture 2019, 500, 393–397. [Google Scholar] [CrossRef]
- Banks, T.M.; Wang, T.; Fitzgibbon, Q.P.; Smith, G.G.; Ventura, T. Double-Stranded RNA Binding Proteins in Serum Contribute to Systemic RNAi Across Phyla—Towards Finding the Missing Link in Achelata. Int. J. Mol. Sci. 2020, 21, 6967. [Google Scholar] [CrossRef]
- Hyde, C.J.; Fitzgibbon, Q.P.; Elizur, A.; Smith, G.G.; Ventura, T. Transcriptional profiling of spiny lobster metamorphosis reveals three new additions to the nuclear receptor superfamily. BMC Genom. 2019, 20, 531. [Google Scholar] [CrossRef]
- Ventura, T.; Chandler, J.C.; Nguyen, T.V.; Hyde, C.J.; Elizur, A.; Fitzgibbon, Q.P.; Smith, G.G. Multi-Tissue Transcriptome Analysis Identifies Key Sexual Development-Related Genes of the Ornate Spiny Lobster (Panulirus ornatus). Genes 2020, 11, 1150. [Google Scholar] [CrossRef]
- Shpak, N.; Manor, R.; Abilevich, L.K.; Mantal, O.; Shavit, K.; Aflalo, E.D.; Toiber, D.; Sagi, A. Short versus long double-stranded RNA activation of a post-transcriptional gene knockdown pathway. RNA Biol. 2017, 14, 1766–1775. [Google Scholar] [CrossRef][Green Version]
- Pamuru, R.R.; Rosen, O.; Manor, R.; Chung, J.S.; Zmora, N.; Glazer, L.; Aflalo, E.D.; Weil, S.; Tamone, S.L.; Sagi, A. Stimulation of molt by RNA interference of the molt-inhibiting hormone in the crayfish Cherax quadricarinatus. Gen. Comp. Endocrinol. 2012, 178, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Rosen, O.; Manor, R.; Weil, S.; Gafni, O.; Linial, A.; Aflalo, E.D.; Ventura, T.; Sagi, A. A Sexual Shift Induced by Silencing of a Single Insulin-Like Gene in Crayfish: Ovarian Upregulation and Testicular Degeneration. PLoS ONE 2010, 5, e15281. [Google Scholar] [CrossRef] [PubMed]
- Sagi, A.; Manor, R.; Ventura, T. Gene silencing in crustaceans: From basic research to biotechnologies. Genes 2013, 4, 620–645. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; You, M.; Vasseur, L.; Zhao, Y.; Liu, C. Development of RNAi in Insects and RNAi-Based Pest Control; InTech: Vienna, Austria, 2011. [Google Scholar] [CrossRef]
- You, L.; Zhang, F.; Huang, S.; Merchant, A.; Zhou, X.; Li, Z. Over-expression of RNA interference ( RNAi) core machinery improves susceptibility to RNAi in silkworm larvae. Insect Mol. Biol. 2020, 29, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Garbutt, J.S.; Bellés, X.; Richards, E.H.; Reynolds, S.E. Persistence of double-stranded RNA in insect hemolymph as a potential determiner of RNA interference success: Evidence from Manduca sexta and Blattella germanica. J. Insect Physiol. 2013, 59, 171–178. [Google Scholar] [CrossRef]
- Yoon, J.-S.; Gurusamy, D.; Palli, S.R. Accumulation of dsRNA in endosomes contributes to inefficient RNA interference in the fall armyworm, Spodoptera frugiperda. Insect Biochem. Mol. Biol. 2017, 90, 53–60. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Cheloufi, S.; Dos Santos, C.O.; Chong, M.M.W.; Hannon, G.J. A dicer-independent miRNA biogenesis pathway that requires Ago catalysis. Nature 2010, 465, 584–589. [Google Scholar] [CrossRef]
- Yang, J.-S.; Maurin, T.; Lai, E.C. Functional parameters of Dicer-independent microRNA biogenesis. RNA 2012, 18, 945–957. [Google Scholar] [CrossRef]
- Lee, D.; Shin, C. Emerging roles of DROSHA beyond primary microRNA processing. RNA Biol. 2018, 15, 186–193. [Google Scholar] [CrossRef]
- De Smet, L.; Ravoet, J.; Wenseleers, T.; De Graaf, D.C. Expression of key components of the RNAi machinery are suppressed in Apis mellifera that suffer a high virus infection. Entomol. Sci. 2017, 20, 76–85. [Google Scholar] [CrossRef]
- Hyde, C.J.; Fitzgibbon, Q.P.; Elizur, A.; Smith, G.G.; Ventura, T. CrustyBase: An interactive online database for crustacean transcriptomes. BMC Genom. 2020, 21, 637. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent updates, new developments and status in 2020. Nucleic Acids Res. 2021, 49, D458–D460. [Google Scholar] [CrossRef] [PubMed]
- Schultz, J.; Milpetz, F.; Bork, P.; Ponting, C.P. SMART, a simple modular architecture research tool: Identification of signaling domains. Proc. Natl. Acad. Sci. USA 1998, 95, 5857–5864. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Thenus australiensis | Panulirus ornatus | RNAi Pathway | Best Blast Hit (NCBI) | E Value |
|---|---|---|---|---|---|
| Dicer1 | ✓ | ✓ | miRNA | endoribonuclease Dcr-1-like [Homarus americanus] | 0 |
| Dicer2 | ✓ | ✓ | siRNA | endoribonuclease Dicer-like [Homarus americanus] | 0 |
| Ago1 | ✓ | ✓ | miRNA | argonaute 1 [Panulirus interruptus] | 0 |
| Ago2 | ✓ | ✓ | siRNA | protein argonaute-2-like [Homarus americanus] | 0 |
| Drosha | ✓ | ✓ | miRNA | LOW QUALITY PROTEIN: ribonuclease 3-like [Penaeus monodon] | 0 |
| Pasha | ✓ | ✓ | miRNA | microprocessor complex subunit DGCR8-like [Homarus americanus] | 0 |
| TRBP | ✓ | ✓ | miRNA and siRNA | RISC-loading complex subunit tarbp2-like [Homarus americanus] | 0 |
| SID1 | ✓ | ✓ | siRNA | uncharacterized protein LOC121864209 isoform X1 [Homarus americanus] | 0 |
| Species | Gene Name | Forward Primer Sequence (5′–3′) | Reverse Primer Sequence (5′–3′) | Amplicon Size | Probe Used |
|---|---|---|---|---|---|
| Thenus australiensis | Dicer2 | TCATAACCGTCAGCAACCCA | GGGCCCTCACATCCATAAGG | 96 | 55 |
| Ago2 | TTAACCATCCACCTGCAGGC | GCGTACCTGTCCATAGAGGC | 73 | 58 | |
| SID1 | GGGGAAACGGAAGGAATGGA | GCATGTTGGGGTCATCCTCA | 79 | 120 | |
| 18S | GGTGCATGGCCGTTCTTA | TGGAGATCCGTCGACTAGTTAAT | 94 | 22 | |
| Panulirus ornatus | Dicer2 | GGGCACATGAACCTGGTACA | GAAGCTCTTTGTTCGGTCGC | 129 | 12 |
| Ago2 | CAAGAACGGGGGATGACCAT | TCTGGCAAATCTCCCTCTGG | 77 | 151 | |
| SID1 | TTTGCTGCCCTACCTACTGC | AAGCACCGATCCTCAACTCC | 80 | 89 | |
| 18S | AACGGACTTGACGGTTGGTT | CTGTTCGGAGCCTGACAGAA | 70 | 49 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banks, T.M.; Wang, T.; Fitzgibbon, Q.P.; Smith, G.G.; Ventura, T. A Tale of Two Lobsters—Transcriptomic Analysis Reveals a Potential Gap in the RNA Interference Pathway in the Tropical Rock Lobster Panulirus ornatus. Int. J. Mol. Sci. 2022, 23, 11752. https://doi.org/10.3390/ijms231911752
Banks TM, Wang T, Fitzgibbon QP, Smith GG, Ventura T. A Tale of Two Lobsters—Transcriptomic Analysis Reveals a Potential Gap in the RNA Interference Pathway in the Tropical Rock Lobster Panulirus ornatus. International Journal of Molecular Sciences. 2022; 23(19):11752. https://doi.org/10.3390/ijms231911752
Chicago/Turabian StyleBanks, Thomas M., Tianfang Wang, Quinn P. Fitzgibbon, Gregory G. Smith, and Tomer Ventura. 2022. "A Tale of Two Lobsters—Transcriptomic Analysis Reveals a Potential Gap in the RNA Interference Pathway in the Tropical Rock Lobster Panulirus ornatus" International Journal of Molecular Sciences 23, no. 19: 11752. https://doi.org/10.3390/ijms231911752
APA StyleBanks, T. M., Wang, T., Fitzgibbon, Q. P., Smith, G. G., & Ventura, T. (2022). A Tale of Two Lobsters—Transcriptomic Analysis Reveals a Potential Gap in the RNA Interference Pathway in the Tropical Rock Lobster Panulirus ornatus. International Journal of Molecular Sciences, 23(19), 11752. https://doi.org/10.3390/ijms231911752